อ่าน 7 นาที
เดลต้า-อะมิโนเลวูลินิก แอซิด ดีไฮดราเตส
อะมิโนเลวูลินิกแอซิดดีไฮดราเทส ( พอร์โฟบิลิ โนเจนซินเท สหรือ ALA ดีไฮดรา เทส หรืออะมิโนเลวูลิเนตดีไฮดราเทส ) เป็น เอนไซม์ ( EC 4.2.1.
เดลต้า-อะมิโนเลวูลินิก แอซิด ดีไฮดราเตส
| อาลัด | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
  | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | อลาด , อลาดห์, PBGS, อะมิโนเลวูลิเนต ดีไฮดราเตส, เอแอลเอ ดีไฮดราเทส | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 125270 ; เอ็มจีไอ : 96853 ; โฮโมโลยีน : 16 ; GeneCards : ALAD ; OMA : ALAD - orthologs | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






อะมิโนเลวูลินิกแอซิดดีไฮดราเทส ( พอร์โฟบิลิ โนเจนซินเท สหรือALA ดีไฮดรา เทส หรืออะมิโนเลวูลิเนตดีไฮดราเทส ) เป็นเอนไซม์ ( EC 4.2.1.24 ) ที่ในมนุษย์ถูกเข้ารหัสโดยยีนALAD [ 5 ] [ 6 ] พอร์ โฟบิลิโนเจนซินเทส (หรือALA ดีไฮดราเทสหรืออะมิโนเลวูลิเนตดีไฮดราเทส ) สังเคราะห์พอร์โฟบิลิโนเจนผ่านการควบแน่น แบบไม่สมมาตร ของ โมเลกุล อะมิโนเลวูลินิกแอซิดสองโมเลกุลเตตระไพร์โรลธรรมชาติทั้งหมดรวมถึงฮีม คลอโรฟิลล์และวิตามินบี12ต่างก็มีพอร์โฟบิลิโนเจนเป็นสารตั้งต้นร่วมกัน พอร์โฟบิลิโนเจนซินเทสเป็นต้นแบบของมอร์ฟีอิน[ 7 ]
การทำงาน
มันเร่งปฏิกิริยาต่อไปนี้ ซึ่งเป็นขั้นตอนที่สองของการสังเคราะห์ทางชีวภาพของพอร์ไฟริน : [ 8 ]
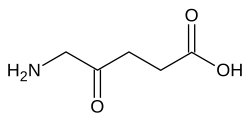


โมเลกุลของกรดอะมิโนเลวูลินิก 2 โมเลกุลจะควบแน่นกันเพื่อสร้างพอร์โฟบิลิโนเจน (สารตั้งต้นของฮีมไซโตโครม และฮีโมโปรตีนอื่นๆ) ปฏิกิริยานี้เป็นขั้นตอนแรกทั่วไปในการสังเคราะห์ทางชีวภาพของเตตระไพร์โรลทางชีวภาพทั้งหมด ในมนุษย์สังกะสีเป็นสิ่งจำเป็นสำหรับกิจกรรมของเอนไซม์[ 9 ]
โครงสร้าง
พื้นฐานเชิงโครงสร้างสำหรับการควบคุมแบบอัลโลสเตอริกของพอร์โฟบิลิโนเจนซินเทส (PBGS) คือการปรับ สมดุลโครงสร้างควอเทอร์ นารีระหว่างออกตาเมอร์และเฮกซาเมอร์ (ผ่านไดเมอร์) ซึ่งแสดงในเชิงแผนภาพเป็น 6mer* ↔ 2mer* ↔ 2mer ↔ 8mer เครื่องหมาย * แสดงถึงการจัดเรียงตัวใหม่ระหว่างสองโดเมนของแต่ละซับยูนิตที่เกิดขึ้นในสถานะแยกตัวออก เนื่องจากถูกห้ามในเชิงสเตอริกในมัลติเมอร์ขนาดใหญ่กว่า[ 7 ]

PBGS ถูกเข้ารหัสโดย ยีนเดียวและมัลติเมอร์ของ PBGS แต่ละตัวประกอบด้วยโปรตีนชนิดเดียวกันหลายสำเนา แต่ละซับยูนิตของ PBGS ประกอบด้วยโดเมน αβ-barrel ประมาณ 300 กรดอะมิโน ซึ่งเป็นที่ตั้งของตำแหน่งออกฤทธิ์ของเอนไซม์อยู่ตรงกลาง และโดเมนแขน N-terminal มากกว่า 25 กรดอะมิโน การควบคุมแบบอัลโลสเตอริกของ PBGS สามารถอธิบายได้ในแง่ของการวางตัวของโดเมน αβ-barrel เทียบกับโดเมนแขน N-terminal
แขน N -terminal แต่ละข้างจะมีปฏิสัมพันธ์กับหน่วยย่อยอื่น ๆ ในมัลติเมอร์ PBGS ได้มากถึงสองแบบ ปฏิสัมพันธ์แบบหนึ่งช่วยทำให้โครงสร้าง "ปิด" ของฝาปิดบริเวณออกฤทธิ์มีความเสถียร ส่วนปฏิสัมพันธ์อีกแบบหนึ่งจะจำกัดการเข้าถึงของตัวทำละลายจากปลายอีกด้านของโครงสร้าง αβ-barrel
ในสภาวะมัลติเมอร์ที่ไม่ทำงาน โดเมนแขน ส่วนปลาย Nไม่ได้มีส่วนร่วมในปฏิกิริยาการทำให้ฝาปิดมีเสถียรภาพ และในโครงสร้างผลึกของชุดประกอบที่ไม่ทำงาน ฝาปิดบริเวณตำแหน่งออกฤทธิ์จะอยู่ในสภาพที่ไม่เป็นระเบียบ
ตัวควบคุมอัลโลสเตอริก
เนื่องจาก PBGS เป็นเอนไซม์ที่พบได้เกือบทั่วไปและมีตำแหน่งออกฤทธิ์ที่มีการอนุรักษ์สูง จึงไม่ใช่เป้าหมายหลักสำหรับการพัฒนาสารต้านจุลชีพและ/หรือสารกำจัดวัชพืชในทางตรงกันข้าม ตำแหน่งอัลโลสเตอริกอาจมีความแปรผันทางวิวัฒนาการมากกว่าตำแหน่งออกฤทธิ์มาก จึงทำให้มีโอกาสในการพัฒนายามากขึ้น[ 7 ]
ความแปรผันทางวิวัฒนาการของกลไก อัลโลสเตอรีของ PBGS นำไปสู่การกำหนดกรอบการอภิปรายเกี่ยวกับการควบคุมอัลโลสเตอรีของ PBGS ในแง่ของปัจจัยภายในและภายนอก
ตัวควบคุมอัลโลสเตอริกภายใน
แมกนีเซียม
ไอออน แมกนีเซียมอัลโลสเตอริกอยู่ที่ส่วนต่อประสานที่มีการไฮเดรตสูงของไดเมอร์โปรออกตาเมอร์สองตัว ดูเหมือนว่าจะแยกตัวออกได้ง่าย และมีการแสดงให้เห็นว่าเฮกซาเมอร์สะสมเมื่อแมกนีเซียมถูกกำจัดออกไปในหลอดทดลอง[ 10 ]
ค่า pH
แม้ว่าโดยทั่วไปจะไม่นิยมพิจารณา ไอออน ไฮโดรเนียมเป็นตัวควบคุมแบบอัลโลสเตอริก แต่ในกรณีของ PBGS การโปรตอนของหมู่ข้างเคียงในตำแหน่งอื่นที่ไม่ใช่บริเวณเร่งปฏิกิริยาได้รับการพิสูจน์แล้วว่าส่งผลต่อสมดุลโครงสร้างระดับควอเทอร์นารี และด้วยเหตุนี้จึงส่งผลต่ออัตราการเกิดปฏิกิริยาเร่งปฏิกิริยาด้วยเช่นกัน
ตัวควบคุมอัลโลสเตอริกภายนอก
การทำให้เฮกซาเมอร์โมเลกุลขนาดเล็กเสถียร
จากการตรวจสอบ PBGS 6mer* พบว่ามีโพรงบนพื้นผิวซึ่งไม่มีอยู่ใน 8mer มีการเสนอว่าโมเลกุลขนาดเล็กที่จับกับโพรงที่มีการเปลี่ยนแปลงทางวิวัฒนาการนี้ จะช่วยทำให้ 6mer* ของ PBGS ที่เป็นเป้าหมายมีความเสถียรมากขึ้น และส่งผลให้ยับยั้งการทำงานได้
ตัวควบคุมอัลโลสเตอริกดังกล่าวเรียกว่ามอร์ฟล็อกเนื่องจากล็อก PBGS ในรูปแบบมอร์ฟีอินที่เฉพาะเจาะจง (6mer*) [ 11 ]
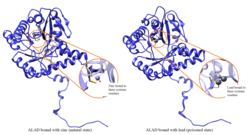
พิษตะกั่ว

กิจกรรมเอนไซม์ ALAD ถูกยับยั้งโดยตะกั่วโดยเริ่มตั้งแต่ระดับตะกั่วในเลือดที่เคยถือว่าปลอดภัย (<10 μg/dL) และยังคงมีความสัมพันธ์เชิงลบอย่างต่อเนื่องในช่วงตั้งแต่ 5 ถึง 95 μg/dL [ 12 ]การยับยั้ง ALAD โดยตะกั่วทำให้เกิดภาวะโลหิตจางเป็นหลัก เนื่องจากตะกั่วทั้งยับยั้งการสังเคราะห์ฮีมและลดอายุขัยของเซลล์เม็ดเลือดแดง ที่ไหลเวียน แต่ยังกระตุ้นการผลิตฮอร์โมนอิริโทรโปเอติน มากเกินไป ทำให้เซลล์เม็ดเลือดแดงเจริญเติบโตจากเซลล์ต้นกำเนิดไม่เพียงพอ ความบกพร่องในยีนโครงสร้าง ALAD อาจทำให้มีความไวต่อพิษตะกั่วและ โรคพอร์ฟิเรีย ตับเฉียบพลัน เพิ่มขึ้น มีการระบุตัวแปรการถอดรหัสที่ตัดต่อแบบอื่นซึ่งเข้ารหัสไอโซฟอร์มที่แตกต่างกัน[ 13 ]
ความขาดแคลน
การขาดพอร์โฟบิลิโนเจนซินเทสมักจะเกิดขึ้นภายหลัง (มากกว่าที่จะเป็นกรรมพันธุ์) และอาจเกิดจากพิษของโลหะหนักโดยเฉพาะพิษของตะกั่วเนื่องจากเอนไซม์ไวต่อการยับยั้งโดยโลหะหนักมาก[ 14 ]
ความบกพร่องทางพันธุกรรมของพอร์โฟบิลิโนเจนซินเทสเรียกว่าโรคพอร์ฟิเรียจากการขาดพอร์โฟบิลิโนเจนซินเทส (หรือ ALA ดีไฮดราเทส)เป็นสาเหตุของ โรค พอร์ฟิเรียที่ หายากมาก [ 15 ]โดยมีรายงานผู้ป่วยน้อยกว่า 10 ราย[ 16 ] โปรตีนกลายพันธุ์ที่เกี่ยวข้องกับโรคทั้งหมดมีแนวโน้มที่จะก่อตัวเป็นเฮกซาเมอร์เมื่อเทียบกับเอนไซม์ของมนุษย์ชนิดปกติ[ 15 ]
 |
PBGS เป็นต้นแบบของมอร์ฟีน
แบบจำลองมอร์ฟีอินของอัลโลสเตอรีที่ยกตัวอย่างโดย PBGS เพิ่มความเข้าใจอีกชั้นหนึ่งเกี่ยวกับกลไกที่เป็นไปได้ในการควบคุมการทำงานของโปรตีน และเสริมการมุ่งเน้นที่เพิ่มขึ้นของชุมชนวิทยาศาสตร์โปรตีนเกี่ยวกับพลวัตของโครงสร้างโปรตีน[ 7 ]
แบบจำลองนี้แสดงให้เห็นว่าพลวัตของปรากฏการณ์ต่างๆ เช่น การเปลี่ยนแปลงโครงสร้างของโปรตีน การเปลี่ยนแปลง สถานะของ โอลิโกเมอร์และปฏิกิริยาระหว่างโปรตีนชั่วคราว สามารถนำมาใช้เพื่อควบคุมการทำงานของเอนไซม์แบบอัลโลสเตอริกได้อย่างไร
ลิงก์ภายนอก
- หน้าแสดงตำแหน่งจีโนม ALADของมนุษย์และ รายละเอียดเกี่ยวกับยีน ALADในUCSC Genome Browser
- delta-Aminolevulinic+Acid+Dehydratase ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- http://www.omim.org/entry/125270?search=pbgs&highlight=pbgs
อ่านเพิ่มเติม
- Bernard A, Lauwerys R (1988). "การเปลี่ยนแปลงของเอนไซม์เดลต้า-อะมิโนเลวูลินิกแอซิดดีไฮดราเทสที่เกิดจากโลหะ". Annals of the New York Academy of Sciences . 514 : 41–7 . doi : 10.1111/j.1749-6632.1987.tb48759.x . PMID 3327436. S2CID 41966070 .
- Jaffe EK (ตุลาคม 2547). "กลไกปฏิกิริยาที่เร่งโดยพอร์โฟบิลิโนเจนซินเทส". เคมีชีวอินทรีย์ . 32 (5): 316– 25. doi : 10.1016/j.bioorg.2004.05.010 . PMID 15381398 .
- Roels HA, Buchet JP, Lauwerys RR, Sonnet J (สิงหาคม 1975). "การเปรียบเทียบผลกระทบในร่างกายของตะกั่วและแคดเมียมอนินทรีย์ต่อระบบกลูตาไธโอนรีดักเทสและเดลต้า-อะมิโนเลวูลินเนตดีไฮดราเทสในเม็ดเลือดแดงของมนุษย์"วารสารBritish Journal of Industrial Medicine 32 ( 3): 181– 92. doi : 10.1136/oem.32.3.181 . PMC 1008057 . PMID 1156566 .
- Ishida N, Fujita H, Fukuda Y, Noguchi T, Doss M, Kappas A, Sassa S (พฤษภาคม 1992). "การโคลนและการแสดงออกของยีนที่บกพร่องจากผู้ป่วยที่เป็นโรคพอร์ฟิเรียเดลต้า-อะมิโนเลวูลินเนตดีไฮดราเทส"วารสารการวิจัยทางคลินิก 89 ( 5): 1431– 7. doi : 10.1172/JCI115732 . PMC 443012 . PMID 1569184 .
- Dawson SJ, White LA (พฤษภาคม 1992). "การรักษาโรคเยื่อหุ้มหัวใจอักเสบจากเชื้อ Haemophilus aphrophilus ด้วยซิโปรฟลอกซาซิน". วารสารการติดเชื้อ . 24 (3): 317– 20. doi : 10.1016/S0163-4453(05)80037-4 . PMID 1602151 .
- Astrin KH, Kaya AH, Wetmur JG, Desnick RJ (สิงหาคม 1991). "RsaI polymorphism ในยีน delta-aminolevulinate dehydratase ของมนุษย์ที่ 9q34" . Nucleic Acids Research . 19 (15): 4307. doi : 10.1093/nar/19.15.4307-a . PMC 328595 . PMID 1678509 .
- Wetmur JG, Kaya AH, Plewinska M, Desnick RJ (ตุลาคม 1991). "ลักษณะทางโมเลกุลของอัลลีล delta-aminolevulinate dehydratase 2 (ALAD2) ของมนุษย์: นัยสำคัญสำหรับการคัดกรองทางโมเลกุลของบุคคลเพื่อความไวต่อพิษตะกั่วทางพันธุกรรม" American Journal of Human Genetics . 49 (4): 757– 63. PMC 1683158 . PMID 1716854 .
- Plewinska M, Thunell S, Holmberg L, Wetmur JG, Desnick RJ (กรกฎาคม 1991). "โรคพอร์ฟิเรียที่ขาดเอนไซม์เดลต้า-อะมิโนเลวูลินเนตดีไฮดราเทส: การระบุรอยโรคระดับโมเลกุลในผู้ป่วยโฮโมไซกัสที่มีอาการรุนแรง"วารสารพันธุศาสตร์มนุษย์อเมริกัน 49 ( 1): 167– 74. PMC 1683193 . PMID 2063868 .
- Potluri VR, Astrin KH, Wetmur JG, Bishop DF, Desnick RJ (กรกฎาคม 1987). "Human delta-aminolevulinate dehydratase: chromosomal localization to 9q34 by in situ hybridization". Human Genetics . 76 (3): 236– 9. doi : 10.1007/BF00283614 . PMID 3036687 . S2CID 32211471 .
- Gibbs PN, Jordan PM (มิถุนายน 1986). "การระบุไลซีนที่ตำแหน่งออกฤทธิ์ของ 5-aminolaevulinate dehydratase ของมนุษย์"วารสารชีวเคมี 236 ( 2): 447– 51. doi : 10.1042/bj2360447 . PMC 1146860 . PMID 3092810 .
- Wetmur JG, Bishop DF, Cantelmo C, Desnick RJ (ตุลาคม 1986). "Human delta-aminolevulinate dehydratase: ลำดับนิวคลีโอไทด์ของโคลน cDNA ความยาวเต็ม" . Proceedings of the National Academy of Sciences of the United States of America . 83 (20): 7703– 7. Bibcode : 1986PNAS...83.7703W . doi : 10.1073/pnas.83.20.7703 . PMC 386789 . PMID 3463993 .
- Wetmur JG, Bishop DF, Ostasiewicz L, Desnick RJ (1986). "การโคลนโมเลกุลของ cDNA สำหรับ delta-aminolevulinate dehydratase ของมนุษย์" Gene . 43 ( 1– 2): 123– 30. doi : 10.1016/0378-1119(86)90015-6 . PMID 3758678 .
- Doss M, von Tiepermann R, Schneider J (1981). "กลุ่มอาการพอร์ฟิเรียตับเฉียบพลันที่มีข้อบกพร่องของพอร์โฟบิลิโนเจนซินเทส" วารสารนานาชาติชีวเคมี 12 ( 5– 6 ): 823– 6. doi : 10.1016/0020-711X(80)90170-6 . PMID 7450139 .
- Kaya AH, Plewinska M, Wong DM, Desnick RJ, Wetmur JG (มกราคม 1994). "ยีน delta-aminolevulinate dehydratase (ALAD) ของมนุษย์: โครงสร้างและการตัดต่อทางเลือกของ mRNA เม็ดเลือดแดงและ mRNA พื้นฐาน" Genomics . 19 (2): 242– 8. doi : 10.1006/geno.1994.1054 . PMID 8188255 .
- Akagi R, Yasui Y, Harper P, Sassa S (กันยายน 1999). "การกลายพันธุ์ใหม่ของเดลต้า-อะมิโนเลวูลินเนตดีไฮดราเทสในเด็กสุขภาพดีที่มีกิจกรรมเอนไซม์ในเม็ดเลือดแดง 12%"วารสารโลหิตวิทยาของอังกฤษ 106 ( 4): 931– 7. doi : 10.1046/j.1365-2141.1999.01647.x . PMID 10519994 . S2CID 24044521 .
- อาคางิ อาร์, ชิมิสึ อาร์, ฟุรุยามะ เค, ดอสส์ โม, ซาสซ่า เอส (มีนาคม 2000) "ความบกพร่องทางโมเลกุลครั้งใหม่ของยีน delta-aminolevulinate dehydratase ในคนไข้ที่เป็นโรคพอร์ฟีเรียตับเฉียบพลันที่ถ่ายทอดทางพันธุกรรม " วิทยาตับ . 31 (3): 704– 8. ดอย : 10.1002/hep.510310321 . PMID 10706561 . S2CID 8998084 .
- Kervinen J, Jaffe EK, Stauffer F, Neier R, Wlodawer A, Zdanov A (กรกฎาคม 2544). "กลไกพื้นฐานสำหรับการยับยั้งการทำงานของเอนไซม์ porphobilinogen synthase โดย 4,7-dioxosebacic acid ซึ่งเป็นสารยับยั้งที่แสดงความจำเพาะต่อสายพันธุ์อย่างมาก" Biochemistry . 40 (28): 8227– 36. CiteSeerX 10.1.1.374.9639 . doi : 10.1021/bi010656k . PMID 11444968 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เดลต้า-อะมิโนเลวูลินิก แอซิด ดีไฮดราเตส
อะมิโนเลวูลินิกแอซิดดีไฮดราเทส ( พอร์โฟบิลิ โนเจนซินเท สหรือ ALA ดีไฮดรา เทส หรืออะมิโนเลวูลิเนตดีไฮดราเทส ) เป็น เอนไซม์ ( EC 4.2.1.
การทำงาน
มันเร่งปฏิกิริยาต่อไปนี้ ซึ่งเป็นขั้นตอนที่สองของ การสังเคราะห์ทางชีวภาพของพอร์ไฟริน : [ 8 ]
โครงสร้าง
พื้นฐานเชิงโครงสร้างสำหรับ การควบคุมแบบอัลโลสเตอริก ของพอร์โฟบิลิโนเจนซินเทส (PBGS) คือการปรับ สมดุลโครงสร้างควอเทอร์ นารี ระหว่างออกตาเมอร์และเฮกซาเมอร์ (ผ่านไดเมอร์) ซึ่งแสดงในเชิงแผนภาพเป็น 6mer* ↔ 2mer* ↔ 2mer ↔ 8mer เครื่องหมาย *...
ตัวควบคุมอัลโลสเตอริก
เนื่องจาก PBGS เป็นเอนไซม์ที่พบได้เกือบทั่วไปและมีตำแหน่งออกฤทธิ์ที่มีการอนุรักษ์สูง จึงไม่ใช่เป้าหมายหลักสำหรับการพัฒนา สารต้านจุลชีพ และ/หรือ สารกำจัดวัชพืช ในทางตรงกันข้าม ตำแหน่งอัลโลสเตอริกอาจมีความแปรผันทางวิวัฒนาการมากกว่าตำแหน่งออกฤทธิ์มาก...