อ่าน 8 นาที
อีแอลเค1
โปรตีน ETS Like-1 Elk-1เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยELK1 Elk -1 ทำหน้าที่เป็นตัวกระตุ้นการถอดรหัสมันถูกจัดอยู่ในกลุ่มปัจจัยเชิงซ้อนสามส่วน (TCF) ซึ่งเป็นกลุ่มย่อยของตระกูล...
อีแอลเค1
| อีแอลเค1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | ELK1 , ปัจจัยถอดรหัส ETS, ปัจจัยถอดรหัส ETS ELK1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 311040 ; เอ็มจีไอ : 101833 ; โฮโมโลยีน : 3832 ; การ์ดยีน : ELK1 ; OMA : ELK1 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





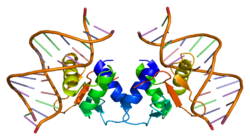
โปรตีน ETS Like-1 Elk-1เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยELK1 [ 5 ] Elk -1 ทำหน้าที่เป็นตัวกระตุ้นการถอดรหัสมันถูกจัดอยู่ในกลุ่มปัจจัยเชิงซ้อนสามส่วน (TCF) ซึ่งเป็นกลุ่มย่อยของตระกูล ETSซึ่งมีลักษณะเฉพาะคือโดเมนโปรตีน ทั่วไป ที่ควบคุม การจับ DNAกับลำดับเป้าหมาย Elk1 มีบทบาทสำคัญในบริบทต่างๆ รวมถึงการสร้างความทรงจำระยะยาวการติดยาเสพติดโรคอัลไซเมอร์ กลุ่ม อาการดาวน์ มะเร็งเต้านมและ ภาวะ ซึม เศร้า
โครงสร้าง

ดังแสดงในรูปที่ 1 โปรตีน Elk1 ประกอบด้วยโดเมนหลายส่วน โดเมน A ซึ่งอยู่บริเวณปลายด้าน N จำเป็นสำหรับการจับกันของ Elk1 กับ DNA บริเวณนี้ยังประกอบด้วยสัญญาณกำหนดตำแหน่งในนิวเคลียส (NLS) และสัญญาณส่งออกนิวเคลียส (NES) ซึ่งรับผิดชอบการนำเข้าและส่งออกนิวเคลียสตามลำดับ โดเมน B ช่วยให้ Elk1 จับกับไดเมอร์ของโคแฟคเตอร์ คือเซรัมรีสปอนส์แฟคเตอร์ (SRF) โดเมน R ซึ่งอยู่ติดกับโดเมน B มีส่วนเกี่ยวข้องกับการยับยั้งกิจกรรมการถอดรหัสของ Elk1 โดเมนนี้มีกรดอะมิโนไลซีนที่อาจเกิดการซูโมอิ เลชัน ซึ่ง เป็นกระบวนการหลังการแปลรหัสที่เสริมความแข็งแกร่งให้กับฟังก์ชันการยับยั้งของโดเมน R โดเมน D มีบทบาทสำคัญในการจับกับไมโทเจนแอคติเวตโปรตีนไคเนส (MAPK) ที่ทำงานอยู่ โดเมน C ซึ่งอยู่บริเวณปลายด้าน C ของ Elk1 ประกอบด้วยกรดอะมิโนที่ถูกฟอสฟอริเลตโดย MAPK ในบริเวณนี้ เซรีน 383 และ 389 เป็นไซต์สำคัญที่ต้องได้รับการฟอ สโฟรีเลต เพื่อให้การถอดรหัสที่ควบคุมโดย Elk1 เกิดขึ้นได้ สุดท้าย โดเมน DEF มีลักษณะเฉพาะสำหรับการโต้ตอบของไคเนสที่ควบคุมสัญญาณภายนอกเซลล์ ที่ถูกกระตุ้น (Erk) ซึ่งเป็น MAPK ชนิดหนึ่ง กับ Elk1 [ 6 ]
การแสดงออก
เนื่องจากบทบาทของ Elk1 ในฐานะปัจจัยการถอดรหัสจึงมีการแสดงออกในนิวเคลียสของเซลล์ที่ไม่ใช่เซลล์ประสาท โปรตีนนี้มีอยู่ในไซโตพลาสซึมเช่นเดียวกับในนิวเคลียสของเซลล์ประสาทที่เจริญ เต็มที่ [ 6 ]ในเซลล์ประสาทหลังการแบ่งตัว sElk1 ซึ่งเป็นรูปแบบหนึ่งของ Elk1 จะแสดงออกเฉพาะในนิวเคลียสเท่านั้น เนื่องจากขาดไซต์ NES ที่มีอยู่ในโปรตีนแบบเต็มความยาว[ 7 ]ยิ่งไปกว่านั้น แม้ว่า Elk1 จะแสดงออกอย่างกว้างขวาง แต่ระดับที่แท้จริงจะแตกต่างกันไปในแต่ละเนื้อเยื่อ ตัวอย่างเช่น สมองของหนูมี Elk1 สูงมาก แต่โปรตีนนี้แสดงออกเฉพาะในเซลล์ประสาทเท่านั้น[ 8 ]
รูปแบบการตัดต่อยีน
นอกเหนือจากโปรตีนแบบเต็มความยาวแล้ว ยีน Elk1 ยังสามารถสร้าง Elk1 เวอร์ชันที่สั้นลงได้สองแบบ คือ ∆Elk1 และ sElk1 การตัดต่อทางเลือกทำให้เกิด ∆Elk1 ตัวแปรนี้ขาดส่วนหนึ่งของโดเมนที่จับกับ DNA ซึ่งช่วยให้เกิดปฏิสัมพันธ์กับ SRF [ 9 ]ในทางกลับกัน sElk1 มีบริเวณที่จับกับ SRF ที่สมบูรณ์ แต่ขาดกรดอะมิโน 54 ตัวแรกที่มี NES พบเฉพาะในเซลล์ประสาท sElk1 ถูกสร้างขึ้นโดยใช้ไซต์เริ่มต้นการแปลภายใน[ 10 ]ทั้ง ∆Elk1 และ sElk1 ซึ่งเป็นเวอร์ชันที่สั้นลงของโปรตีนแบบเต็มความยาว สามารถจับกับ DNA และกระตุ้นการส่งสัญญาณของเซลล์ต่างๆ ได้ อันที่จริง sElk1 ต่อต้าน Elk1 ในการสร้างความแตกต่างของเซลล์ประสาทและการควบคุม การส่ง สัญญาณ ของปัจจัย การเจริญเติบโตของเส้นประสาท / ERK [ 8 ]
การส่งสัญญาณ
เป้าหมายปลายทางของ Elk1 คือองค์ประกอบการตอบสนองต่อซีรั่ม (SRE) ของโปรโตออนโคยีน c -fos [ 11 ] [ 12 ]ในการสร้างc-fosซึ่งเป็นโปรตีนที่เข้ารหัสโดย ยีน Fosนั้น Elk1 จำเป็นต้องได้รับการฟอสโฟรีเลตโดย MAPK ที่ปลาย C ของมัน[ 13 ] [ 14 ] MAPK เป็นตัวกระตุ้น ขั้นสุดท้าย ของเส้นทางการส่งสัญญาณที่เริ่มต้นที่เยื่อหุ้มเซลล์[ 15 ]การฟอสโฟรีเลตโดย MAPK ส่งผลให้เกิดการเปลี่ยนแปลงโครงสร้างของ Elk1 [ 16 ]ดังที่เห็นในรูปที่ 2 ไคเนส Rafทำงานอยู่เหนือ MAPK เพื่อกระตุ้นพวกมันโดยการฟอสโฟรีเลตและกระตุ้น MEK หรือไคเนส MAPK หรือ ERK [ 17 ] [ 18 ] [ 19 ] [ 20 ] Raf เองถูกกระตุ้นโดย Ras ซึ่งเชื่อมโยงกับตัวรับปัจจัยการเจริญเติบโตที่มี กิจกรรม ไทโรซีนไคเนสผ่านGrb2และ Sos [ 21 ] Grb2 และ Sos สามารถกระตุ้น Ras ได้ก็ต่อเมื่อมีการจับกันของปัจจัยการเจริญเติบโตกับตัวรับที่เกี่ยวข้องเท่านั้น อย่างไรก็ตาม การกระตุ้น Raf ไม่ได้ขึ้นอยู่กับ Ras เพียงอย่างเดียว โปรตีนไคเนส C ซึ่งถูกกระตุ้นโดยฟอร์บอลเอสเทอร์สามารถทำหน้าที่เดียวกันกับ Ras ได้[ 22 ] MEK ไคเนส (MEKK) ยังสามารถกระตุ้น MEK ซึ่งจากนั้นจะกระตุ้น MAPK ทำให้ Raf ไม่จำเป็นในบางครั้ง[ 23 ]ดังนั้น เส้นทางการส่งสัญญาณต่างๆ จึงผ่าน MEK และ MAPK และนำไปสู่การกระตุ้น Elk1 หลังจากกระตุ้น Elk1 แล้ว SRF ซึ่งช่วยให้ Elk1 จับกับ โปรโมเตอร์ c-fosจะต้องถูกดึงเข้ามา การจับกันของ Elk1 กับ SRF เกิดขึ้นเนื่องจากปฏิกิริยาระหว่างโปรตีนกับโปรตีนระหว่างโดเมน B ของ Elk1 และ SRF และปฏิกิริยาระหว่างโปรตีนกับ DNA ผ่านทางโดเมน A [ 6 ]
โปรตีนที่กล่าวถึงข้างต้นเปรียบเสมือนสูตรสำหรับเอาต์พุตการส่งสัญญาณเฉพาะอย่าง หากส่วนประกอบใดส่วนประกอบหนึ่ง เช่น SRF ขาดหายไป เอาต์พุตที่แตกต่างก็จะเกิดขึ้น ในกรณีนี้ การขาด SRF นำไปสู่การกระตุ้นยีนอื่นโดย Elk1 [ 16 ]ดังนั้น Elk1 จึงสามารถโต้ตอบกับไซต์การจับ ETS ได้อย่างอิสระ เช่นในกรณีของ โปรโตออนโคยีน lckในรูปที่ 2 [ 16 ]ยิ่งไปกว่านั้น ระยะห่างและการวางแนวสัมพัทธ์ของไซต์การจับ Elk1 กับ SRE ค่อนข้างยืดหยุ่น[ 24 ]ซึ่งบ่งชี้ว่ายีนต้นที่ควบคุมโดย SRE นอกเหนือจากc-fosอาจเป็นเป้าหมายของ Elk1 egr-1เป็นตัวอย่างของเป้าหมายของ Elk1 ที่ขึ้นอยู่กับการโต้ตอบกับ SRE [ 16 ]ในที่สุดการฟอสโฟรีเลชั่นของ Elk1 สามารถส่งผลให้เกิดการผลิตโปรตีนจำนวนมาก ขึ้นอยู่กับปัจจัยอื่นๆ ที่เกี่ยวข้องและการโต้ตอบเฉพาะระหว่างกัน
เมื่อศึกษาเส้นทางการส่งสัญญาณ การกลายพันธุ์สามารถเน้นย้ำถึงความสำคัญของแต่ละองค์ประกอบที่ใช้ในการกระตุ้นเป้าหมายปลายทางได้ ตัวอย่างเช่น การหยุดชะงักของโดเมน C-terminal ของ Elk1 ที่ MAPK ฟอสโฟรีเลตจะกระตุ้นการยับยั้งการทำงาน ของ c-fos [ 16 ]ในทำนองเดียวกัน SRF ที่ทำงานผิดปกติ ซึ่งปกติจะยึด Elk1 ไว้กับ SRE จะทำให้Fosไม่ถูกถอดรหัส[ 21 ]ในขณะเดียวกัน หากไม่มี Elk1 SRF ก็ไม่สามารถกระตุ้น การถอดรหัส c-fosได้หลังจากได้รับการกระตุ้นจาก MAPK [ 16 ]ด้วยเหตุผลเหล่านี้ Elk1 จึงเป็นตัวเชื่อมที่สำคัญระหว่างเส้นทางการส่งสัญญาณและการเริ่มต้นการถอดรหัสยีน
ความสำคัญทางคลินิก
ความทรงจำระยะยาว
การสร้างความทรงจำระยะยาวอาจขึ้นอยู่กับ Elk1 สารยับยั้ง MEKจะปิดกั้นการฟอสโฟรีเลชัน ของ Elk1 และ ทำให้ การเรียนรู้ การหลีกเลี่ยงรสชาติที่ไม่พึงประสงค์ลดลง นอกจากนี้การเรียนรู้การหลีกเลี่ยงซึ่งเกี่ยวข้องกับการที่ผู้ถูกทดลองเรียนรู้ว่าการตอบสนองเฉพาะอย่างจะนำไปสู่การป้องกันสิ่งเร้าที่ไม่พึงประสงค์นั้น มีความสัมพันธ์กับการเพิ่มขึ้นของการทำงานของ Erk, Elk1 และ c-fos ในฮิปโปแคมปัสอย่างชัดเจน บริเวณนี้ของสมองมีส่วนเกี่ยวข้องกับการจัดเก็บข้อมูลทั้งระยะสั้นและระยะยาว เมื่อการจับกันของ Elk1 หรือ SRF กับ DNA ถูกปิดกั้นในฮิปโปแคมปัสของหนู การแยกตัวของ SRF เท่านั้นที่จะรบกวนความทรงจำเชิงพื้นที่ ระยะยาว แม้ว่าการโต้ตอบของ Elk1 กับ DNA อาจไม่จำเป็นต่อการสร้างความทรงจำ แต่บทบาทเฉพาะของมันยังคงต้องได้รับการสำรวจต่อไป เนื่องจาก1การทำงานของ Elk1 สามารถกระตุ้นเหตุการณ์ระดับโมเลกุลอื่นๆ ที่ไม่จำเป็นต้องให้ Elk1 จับกับ DNA ได้ ตัวอย่างเช่น Elk1 มีส่วนเกี่ยวข้องกับการฟอสโฟรีเลชันของฮิสโตนปฏิสัมพันธ์ที่เพิ่มขึ้นกับ SRF และการดึงดูดกลไกการถอดรหัสพื้นฐาน ซึ่งทั้งหมดนี้ไม่จำเป็นต้องมีการจับกันโดยตรงของ Elk1 กับ DNA [ 6 ]
การติดยาเสพติด
การกระตุ้น Elk1 มีบทบาทสำคัญในการติดยาเสพติดหลังจากที่หนูได้รับโคเคนจะสังเกตเห็น การไฮเปอร์ฟอ สโฟรีเลชัน ของ Erk และ Elk1 อย่างรุนแรงและชั่วคราว ในสไตรอาตัม เมื่อหนูเหล่านี้ได้รับ สารยับยั้ง MEK การฟอสโฟรีเลชัน ของ Elk1 จะหายไป หากไม่มี Elk1 ที่ทำงานอยู่ การผลิต c-fos และ ความชอบสถานที่แบบมีเงื่อนไขที่เกิดจากโคเคนจะถูกปิดกั้น นอกจากนี้ การบริโภค เอทานอล เฉียบพลัน นำไปสู่การฟอสโฟรีเลชันของ Elk1 มากเกินไปในอะมิกดาลาการยับยั้งการทำงานของ Elk1 ยังพบว่าช่วยลดการตอบสนองของเซลล์ต่อสัญญาณการถอนยาและการรักษาด้วยโอปิออยด์ซึ่งเป็นยาที่เก่าแก่ที่สุดชนิดหนึ่งของโลก โดยรวมแล้ว ผลลัพธ์เหล่านี้เน้นย้ำว่า Elk1 เป็นองค์ประกอบสำคัญของการติดยาเสพติด[ 6 ]
พยาธิสรีรวิทยา
การสะสมของ เปปไทด์ เบต้าอะไมลอยด์ (Aβ) แสดงให้เห็นว่าก่อให้เกิดและ/หรือกระตุ้นโรคอัลไซเมอร์ Aβ ขัดขวางการฟอสโฟรีเลชันของ Elk1 ที่เกิดจากBDNF เมื่อการกระตุ้น Elk1 ถูกขัดขวางในเส้นทางนี้ การควบคุมยีนที่ขับเคลื่อนโดย SRE จะนำไปสู่ความเปราะบางของเซลล์ประสาทที่เพิ่มขึ้น Elk1 ยังยับยั้งการถอดรหัสของ พรีเซนิลิน 1 (PS1) ซึ่งเข้ารหัสโปรตีนที่จำเป็นสำหรับขั้นตอนสุดท้ายของการประมวลผลโปรตีโอไลติกแบบต่อเนื่องของโปรตีนต้นกำเนิดอะไมลอยด์ (APP) APP สร้าง Aβ รูปแบบต่างๆ (โพลีเปปไทด์ Aβ42/43) ยิ่งไปกว่านั้น PS1 ยังมีความเกี่ยวข้องทางพันธุกรรมกับกรณีเริ่มต้นเร็วส่วนใหญ่ของโรคอัลไซเมอร์ในครอบครัว ข้อมูลเหล่านี้เน้นย้ำถึงความเชื่อมโยงที่น่าสนใจระหว่าง Aβ, Elk1 และ PS1 [ 6 ]
ภาวะอื่นที่เกี่ยวข้องกับ Elk1 คือกลุ่มอาการดาวน์หนูทดลองในระยะตัวอ่อนและวัยชราที่มีภาวะทางพยาธิสรีรวิทยานี้แสดงให้เห็นถึงการลดลงของกิจกรรมของแคลซิเนอรินซึ่งเป็นฟอสฟาเทสหลักสำหรับ Elk1 หนูเหล่านี้ยังมีการเปลี่ยนแปลงที่ขึ้นอยู่กับอายุในการกระตุ้น ERK ยิ่งไปกว่านั้น การแสดงออกของSUMO3ซึ่งยับยั้งกิจกรรมของ Elk1 เพิ่มขึ้นในผู้ป่วยที่เป็นผู้ใหญ่ที่มีกลุ่มอาการดาวน์ ดังนั้น กลุ่มอาการดาวน์จึงมีความสัมพันธ์กับการเปลี่ยนแปลงในเส้นทาง ERK, แคลซิเนอริน และ SUMO ซึ่งทั้งหมดนี้ทำหน้าที่ต่อต้านกิจกรรมของ Elk1 [ 6 ]
นอกจากนี้ Elk1 ยังมีปฏิสัมพันธ์กับ ตัวแปรการต่อเชื่อมของ BRCA1ได้แก่ BRCA1a และ BRCA1b ปฏิสัมพันธ์นี้ช่วยเสริมการยับยั้งการเจริญเติบโตที่เกิดจาก BRCA1 ใน เซลล์ มะเร็งเต้านม Elk1 อาจเป็นเป้าหมายปลายทางของ BRCA1 ในเส้นทางการควบคุมการเจริญเติบโต วรรณกรรมล่าสุดเผยให้เห็นว่า กิจกรรมของโปรโมเตอร์ c-fos ถูกยับยั้ง ในขณะที่การแสดงออกมากเกินไปของ BRCA1a/1b ลดการกระตุ้น SRE ที่เกิดจาก MEK ผลลัพธ์เหล่านี้แสดงให้เห็นว่ากลไกหนึ่งของการยับยั้งการเจริญเติบโตและเนื้องอกโดยโปรตีน BRCA1a/1b ทำงานผ่านการยับยั้งการ แสดงออกของยีนเป้าหมายปลายทางของ Elk1 เช่นFos [ 25 ]
ภาวะซึมเศร้ามีความเชื่อมโยงกับ Elk1 การลดลงของการฟอสโฟรีเลชัน ของ Elk1 ที่เกิดจาก Erk พบได้ในฮิปโปแคมปัสและคอร์เทกซ์ส่วนหน้าของสมองหลังการเสียชีวิตของผู้ที่ฆ่าตัวตาย การส่งสัญญาณ Erk ที่ไม่สมดุลมีความสัมพันธ์กับภาวะซึมเศร้าและพฤติกรรมฆ่าตัวตาย การวิจัยในอนาคตจะเปิดเผยบทบาทที่แท้จริงของ Elk1 ในพยาธิสรีรวิทยาของภาวะซึมเศร้า[ 6 ]
ลิงก์ภายนอก
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ อีแอลเค1
โปรตีน ETS Like-1 Elk-1เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยELK1 Elk -1 ทำหน้าที่เป็นตัวกระตุ้นการถอดรหัสมันถูกจัดอยู่ในกลุ่มปัจจัยเชิงซ้อนสามส่วน (TCF) ซึ่งเป็นกลุ่มย่อยของตระกูล...
โครงสร้าง
ดังแสดงในรูปที่ 1 โปรตีน Elk1 ประกอบด้วยโดเมนหลายส่วน โดเมน A ซึ่งอยู่บริเวณปลายด้าน N จำเป็นสำหรับการจับกันของ Elk1 กับ DNA บริเวณนี้ยังประกอบด้วย สัญญาณกำหนดตำแหน่งในนิวเคลียส (NLS) และ สัญญาณส่งออกนิวเคลียส (NES)...
การแสดงออก
เนื่องจากบทบาทของ Elk1 ในฐานะ ปัจจัยการถอดรหัส จึงมีการแสดงออกในนิวเคลียสของเซลล์ที่ไม่ใช่เซลล์ประสาท โปรตีนนี้มีอยู่ในไซโตพลาสซึมเช่นเดียวกับในนิวเคลียสของเซลล์ประสาทที่เจริญ เต็มที่ [ 6 ] ในเซลล์ประสาทหลังการแบ่งตัว sElk1 ซึ่งเป็นรูปแบบหนึ่งของ Elk1...
รูปแบบการตัดต่อยีน
นอกเหนือจากโปรตีนแบบเต็มความยาวแล้ว ยีน Elk1 ยังสามารถสร้าง Elk1 เวอร์ชันที่สั้นลงได้สองแบบ คือ ∆Elk1 และ sElk1 การตัดต่อทางเลือก ทำให้เกิด ∆Elk1 ตัวแปรนี้ขาดส่วนหนึ่งของโดเมนที่จับกับ DNA ซึ่งช่วยให้เกิดปฏิสัมพันธ์กับ SRF [ 9 ] ในทางกลับกัน sElk1...