อ่าน 17 นาที
ปัจจัยยุติการแปลรหัสยูคาริโอต 1
ปัจจัยยุติการแปลของยูคาริโอต 1 (eRF1) หรือที่เรียกอีกอย่างว่า TB3-1 หรือ SUP45L1 เป็น โปรตีน ที่ถูกเข้ารหัสโดย ยีน ERF1 ในยูคาริโอต eRF1 เป็นโปรตีนที่จำเป็นซึ่งเกี่ยวข้องกับ...
ปัจจัยยุติการแปลรหัสยูคาริโอต 1
| อีทีเอฟพี1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | ETF1 , D5S1995, ERF, ERF1, RF1, SUP45L1, TB3-1, ปัจจัยยุติการแปลรหัสของยูคาริโอต 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 600285 ; เอ็มจีไอ : 2385071 ; โฮโมโลยีน : 3475 ; GeneCards : ETF1 ; OMA : ETF1 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




ปัจจัยยุติการแปลของยูคาริโอต1 (eRF1) หรือที่เรียกอีกอย่างว่าTB3-1หรือSUP45L1เป็นโปรตีน ที่ถูกเข้ารหัสโดย ยีน ERF1 ในยูคาริโอต eRF1 เป็นโปรตีนที่จำเป็นซึ่งเกี่ยวข้องกับ การจดจำ รหัสหยุดในการแปลการยุติการแปล และการสลายตัวของ mRNA ที่เกิดจากรหัสหยุดโดยผ่านคอมเพล็กซ์ SURF [ 5 ]
ความสำคัญ
เซลล์ทุกเซลล์จำเป็นต้องสร้างโปรตีนผ่านกระบวนการถอดรหัสและการแปล[ 6 ] eRF1 มีความสำคัญต่อการอยู่รอดและการบำรุงรักษาเซลล์เนื่องจากมีส่วนเกี่ยวข้องในการยุติการแปล การกลายพันธุ์ใดๆ ต่อตำแหน่งการจับและการเร่งปฏิกิริยาของ eRF1 จะทำให้การยุติการแปลไม่ถูกต้อง ซึ่งเป็นอันตรายถึงชีวิตต่อเซลล์ นอกจากนี้ eRF1 ยังช่วยปกป้องเซลล์จากการสร้างโปรตีนที่เป็นอันตรายจากการกลายพันธุ์แบบไร้ความหมาย[ 7 ]
โครงสร้าง
หลัก
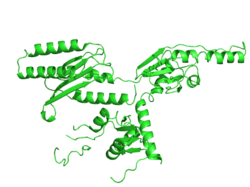
eRF1 ประกอบด้วยสายโพลีเปปไทด์ของกรดอะมิโนที่มีรูปร่างคล้ายตัวอักษร Y โปรตีนนี้ประกอบด้วยโดเมนหลัก 3 โดเมน ได้แก่ ลำต้นและกิ่ง 2 กิ่ง แต่ละโดเมนมีวัตถุประสงค์เฉพาะและรูปแบบการพับที่แตกต่างกัน ซึ่งช่วยให้โปรตีนทำงานได้อย่างถูกต้อง แม้ว่าแต่ละโดเมนจะมีเอกลักษณ์เฉพาะตัว แต่ทั้งหมดก็มีโครงสร้างพื้นฐานของคลาส α-β sandwich ซึ่งโดยพื้นฐานแล้วคือ แกน β sheetที่ล้อมรอบด้วยα helicesโดเมน 1 บางครั้งเรียกว่าโดเมน N สร้างขึ้นจากแกน β sheet ที่มี 4 สายล้อมรอบด้วย α-helices 2 อัน (α2 และ α3) หน่วยย่อย α2 และ α3 ขดตัวและจับกันเพื่อสร้างโครงสร้าง hairpin ที่มี motif NIKS [ 8 ] motif YxxCxxxF และ GTS loop [ 9 ]สันนิษฐานว่าไซต์เหล่านี้เป็นส่วนประกอบหลักในการจดจำรหัสหยุด นอกจากนี้ ปลาย N-terminus ยังอยู่ในโดเมน 1 ซึ่งมีปฏิสัมพันธ์กับโดเมน 3 เพื่อรักษาเสถียรภาพของโปรตีน โดเมน 3 บางครั้งเรียกว่าโดเมน C ประกอบด้วยปลาย C-terminus ของโพลีเปปไทด์ นอกจากนี้ โครงสร้างและหน้าที่ของโดเมน 3 ซึ่งเรียกว่าโดเมน M ยังไม่เป็นที่รู้จักมากนัก เนื่องจากข้อจำกัดทางแสงขัดขวางการวิจัยเพิ่มเติม โดเมน 2 ประกอบด้วยโครงสร้างแบบ α-β sandwich โดยที่สายด้านนอกเส้นหนึ่งของ sandwich ไม่มีโครงสร้างทุติยภูมิ ส่วนกรดอะมิโนหลักช่วยให้เกิดไซต์ GGQ ได้[ 8 ]
โครงสร้างแบบพับของ eRF1 นั้นเลียนแบบโครงสร้างของ โมเลกุล tRNA โดยพื้นฐานแล้ว ซึ่งทำให้มั่นใจได้ว่ากลไกของ eRF1 จะพอดีกับตำแหน่งอะมิโนเอซิลของไรโบโซม นอกจากนี้ eRF1 ยังมีความสามารถในการจดจำโคดอน ซึ่งเป็นหนึ่งในกระบวนการสำคัญที่ดำเนินการโดยโมเลกุล tRNA [ 10 ]เนื่องจากทั้ง tRNA และ eRF1 ต่างก็มีความสามารถในการจับกับ mRNA และ ศูนย์ เปปทิดิลทรานสเฟอเรส จึงมีขนาดใกล้เคียงกัน โดย eRF1 มีความกว้าง 71 Å เทียบกับ 70 Å ของฟีนิลอะลานีน tRNA [ 8 ] [ 11 ]โมเลกุลทั้งสองมีปฏิสัมพันธ์กับGTPases : eEF-1α ( EF-Tuในโปรคาริโอต) กับ tRNA และ eRF3 กับ eRF1 [ 8 ]
รูปแบบในชีววิทยาสรีรวิทยาที่เชื่อมโยงระหว่างโครงสร้างและหน้าที่การทำงานนั้นปรากฏให้เห็นอย่างชัดเจนระหว่างโมเลกุล eRF1 และ tRNA ไซต์ GGQ ใน eRF1 เทียบเท่ากับกลุ่มอะมิโนเอซิลที่ติดอยู่กับ tRNA โครงสร้างทั้งสองมีส่วนช่วยในการจดจำและจับกับไซต์เปปไทด์ทรานสเฟอเรสในไรโบโซม โครงสร้างทั้งสองจัดวางไซต์ที่มีความเชี่ยวชาญสูงให้ห่างจากส่วนที่เหลือของโมเลกุลเพื่อให้แน่ใจว่ามีการโต้ตอบแบบแยกเดี่ยว นอกจากนี้ โดเมน 2 ของ eRF1 ยังมีโครงสร้างคล้ายกับก้านอะมิโนเอซิลของ tRNA [ 11 ]ก้าน T ของ tRNA และโดเมน 3 ของ eRF1 ต่างก็ทำหน้าที่โต้ตอบกับโปรตีน GTPase
ลวดลาย GGQ
ลำดับกรดอะมิโน GGQ เป็นลำดับกรดอะมิโนที่คงที่ในปัจจัยปลดปล่อย (release factors) ในสิ่งมีชีวิตทุกโดเมน ตำแหน่ง GGQ ประกอบด้วยกรดอะมิโนไกลซีน 2 ตัว ตามด้วยกรดอะมิโนกลูตา มีน 1 ตัว ใน eRF1 ตำแหน่ง GGQ อยู่ที่ กรดอะมิโน ลำดับที่ 183-185 ของพอลิเปปไทด์ ซึ่งอยู่ในโดเมน 2 การวางตัวที่ห่างออกไปของตำแหน่ง GGQ นั้นได้รับการทำให้เสถียรโดยผลกระทบจากคุณสมบัติไม่ชอบน้ำของกรดอะมิโนข้างเคียง เช่นลิวซีน 176, โพรลีน 177, ฟีนิลอะลานีน 190 และลิวซีน 193 เชื่อกันว่ากรดอะมิโน gln185 ในลำดับ GGC เป็นตำแหน่งเร่งปฏิกิริยาหลักสำหรับการไฮโดรไลซิสของพันธะเอสเทอร์เปปทิดิล-tRNA ในเปปทิดิลทรานสเฟอเรสของไรโบโซม ตำแหน่ง GGQ ไม่เกี่ยวข้องกับหน้าที่ใดๆ ของการจดจำรหัสหยุด (stop codon recognition), การเชื่อมต่อกับไรโบโซม (ribosomal docking) หรือการจับกับ eRF3 [ 12 ] [ 8 ]
ลวดลาย NIKS
มอติฟ NIKS เป็นลำดับกรดอะมิโนที่มีการอนุรักษ์สูงซึ่งตั้งอยู่ที่ปลาย N ในโดเมน 1 (กรดอะมิโนตำแหน่งที่ 61-64) มอติฟ NIKS ประกอบด้วยกรดอะมิโนแอสปาราจีน (N), ไอโซลิวซีน (I), ไลซีน (K) และซีรีน (S) [ 13 ]หน้าที่หลักของมอติฟ NIKS คือการจดจำนิวคลีโอไทด์ตัวแรกในโคดอนหยุด ซึ่งมักจะเป็นยูราซิล นอกจากนี้ การกลายพันธุ์ในบริเวณนี้ยังเกี่ยวข้องกับการลดลงของการจับกับไรโบโซมและกิจกรรมเร่งปฏิกิริยา[ 14 ]
ลวดลาย YxxCxxxF และลูป GTS
มอติฟ YxxCxxxF และลูป GTS เป็นตำแหน่งกรดอะมิโนสองตำแหน่งที่อยู่ในโดเมน 1 ของ eRF1 มอติฟ YxxCxxxF พบในกรดอะมิโนตำแหน่งที่ 121-131 ในขณะที่ลูป GTS พบในกรดอะมิโนตำแหน่งที่ 31-33 มอติฟ YxxCxxxF ประกอบด้วยกรดอะมิโนที่ไม่เปลี่ยนแปลงสามตำแหน่ง ได้แก่ไทโรซีน (Y) ซิสเทอีน (C) และฟีนิลอะลานีน (F) ตำแหน่งเหล่านี้แยกออกจากกันในเชิงโครงสร้างในโปรตีน eRF1 ที่พับตัวแล้ว อย่างไรก็ตาม หน้าที่หลักของพวกมันคล้ายคลึงกันมาก พวกมันมีหน้าที่ในการ จดจำ พิวรีนในตำแหน่งที่ 2 และ 3 ของรหัสหยุด[ 15 ]

การจับกันของ eRF3
การจับกันของ eRF3 และ GTP กับ eRF1 เพื่อสร้างคอมเพล็กซ์เป็นสิ่งจำเป็นสำหรับการยุติการแปล การโต้ตอบระหว่างโดเมน C ของทั้ง eRF1 และ eRF3 เป็นแรงหลักที่ยึดคอมเพล็กซ์ไว้ด้วยกัน[ 17 ]อย่างไรก็ตาม ต่อมาได้มีการค้นพบว่าโดเมน M ก็มีส่วนช่วยในการรักษาเสถียรภาพของคอมเพล็กซ์ด้วย ตำแหน่งของ eRF3 ที่อยู่ใกล้กับโมทีฟ GGQ ซึ่งอยู่ในโดเมน C ช่วยให้ eRF1 มีผลเร่งปฏิกิริยาในการไฮโดรไลซ์เปปทิดิล-tRNA มากขึ้น[ 18 ]
โฮโมล็อก
แต่ละโดเมนของสิ่งมีชีวิต ( ยูแบคทีเรียอาร์เคียและยูคาริโอต ) มีปัจจัยปลดปล่อยที่แตกต่างกันซึ่งเกี่ยวข้องกับการยุติการแปล ยูแบคทีเรียมีปัจจัยปลดปล่อยหลายตัวเพื่อจดจำรหัสหยุด ในขณะที่ยูคาริโอต (eRF1) และอาร์เคีย (aRF1) มีโปรตีนเพียงตัวเดียวเพื่อจดจำรหัสหยุดทั้งสามตัว ความแตกต่างทางโครงสร้างและหน้าที่ระหว่างปัจจัยปลดปล่อยของยูแบคทีเรียและอาร์เคีย/ยูคาริโอตเชื่อว่าวิวัฒนาการแยกจากกันโดยมีจุดแยกตัวในช่วงต้น ความคล้ายคลึงกันทางหน้าที่ระหว่าง eRF1 และ aRF1 นำไปสู่ทฤษฎีของบรรพบุรุษร่วมกันที่โปรตีนทั้งสองวิวัฒนาการมาจาก[ 19 ]อย่างไรก็ตาม มีการศึกษาเกี่ยวกับปัจจัยปลดปล่อยของอาร์เคียเพียงเล็กน้อย[ 20 ]
ในโปรคาริโอต ปัจจัยการปลดปล่อยถูกจำแนกออกเป็น 2 ประเภท ประเภทที่ 1 ปัจจัยการปลดปล่อยจะจดจำรหัสหยุด และประเภทที่ 2 จะกระตุ้นการไฮโดรไลซิสโดยกิจกรรม GTPase อย่างไรก็ตาม โปรคาริโอตไม่มีโปรตีนตัวเดียวที่จดจำรหัสหยุดทั้งหมด รหัสหยุด UAG ถูกถอดรหัสโดยปัจจัยการปลดปล่อย 1 (RF1) และ UGA ถูกถอดรหัสโดยปัจจัยการปลดปล่อย 2รหัสหยุดสุดท้าย UAA ถูกถอดรหัสโดยทั้ง RF1 และ RF2 [ 21 ]ในยูคาริโอต eRF1 จดจำรหัสหยุดทั้งสามตัว[ 22 ]
แม้ว่าจะมีความแตกต่างที่ชัดเจนระหว่างการจดจำโคดอนของโปรคาริโอตและการจดจำโคดอนของอาร์เคียน/ยูคาริโอต แต่การทำงานของไซต์เร่งปฏิกิริยายังคงได้รับการอนุรักษ์ไว้ตลอดทุกโดเมน แต่ละโดเมนมีไซต์ GGQ ที่สำคัญเพื่อส่งเสริมการไฮโดรไลซิสของเปปไทด์[ 23 ]
การจดจำโคดอนหยุด
การสิ้นสุดของการแปลถูกกำหนดโดยการมีอยู่ของปัจจัยปลดปล่อยที่รู้จักรหัสหยุด ซึ่งจะเร่งปฏิกิริยาการปลดปล่อยโปรตีนที่สังเคราะห์ขึ้นใหม่ มีรหัสหยุดสามแบบที่พบในสิ่งมีชีวิตทุกโดเมน ได้แก่ UGA, UAG และ UAA [ 24 ]รหัสหยุดแต่ละรหัสเริ่มต้นด้วยนิวคลีโอไทด์ยูราซิลตามด้วยพิวรีนสองตัว (อะดีโนซีนและกัวนีน) ซึ่งจำเป็นสำหรับพื้นฐานระดับโมเลกุลของการจดจำรหัสหยุด โปรตีน eRF1 สามารถจดจำรหัสหยุดทั้งสามแบบได้ ซึ่งหมายความว่ามันต้องมีวิธีแยกแยะรหัสได้อย่างมีประสิทธิภาพมาก ตำแหน่งสามตำแหน่งที่ใช้สำหรับการจดจำรหัสหยุดคือ YxxCxxxF, ลูป GTS และตำแหน่ง NIKS
จุดประสงค์หลักของไซต์ที่มีความเชี่ยวชาญสูงเหล่านี้คือการสร้างเครือข่ายการเชื่อมโยงด้วยพันธะไฮโดรเจนที่กว้างขวางกับรหัสหยุดของ mRNA กระบวนการนี้แบ่งออกเป็นสองส่วน ได้แก่ การจดจำนิวคลีโอไทด์ตัวแรก (ยูราซิล) และการจดจำนิวคลีโอไทด์ในตำแหน่งที่สองและสาม
การจดจำนิวคลีโอไทด์ครั้งแรก
ไซต์ NIKS มีหน้าที่ในการจับกับนิวคลีโอไทด์ตัวแรกของรหัสหยุด: ยูราซิล โดยเกิดขึ้นจากการที่หมู่ Asn61 และ Lys63 ของ NIKS สร้างพันธะไฮโดรเจนกับหมู่คาร์บอนิลของยูราซิล นอกจากนี้ยังมีการสร้างพันธะไฮโดรเจนรองระหว่างหมู่ Asn61 กับหมู่คาร์บอนิลอีกหมู่หนึ่งที่พบในยูราซิล ยิ่งไปกว่านั้น ยังมีสมมติฐานว่าหมู่ Lys63 ทำปฏิกิริยากับโครงสร้างหลักของ mRNA ซึ่งมีส่วนช่วยให้เกิดความเสถียรและการจัดเรียงตัวที่เหมาะสมของ eRF1 ความจำเพาะของกลไกสำหรับยูราซิลหมายความว่านิวคลีโอไทด์อื่นๆ (กัวนีน อะดีโนซีน ไทมีน ไซโตซีน) จะสร้างโครงสร้างที่ไม่มีพันธะไฮโดรเจนมากพอที่จะทำให้การจับของ eRF1 มีเสถียรภาพ[ 25 ] [ 26 ] [ 27 ]
การจดจำนิวคลีโอไทด์ลำดับที่สองและสาม
นิวคลีโอไทด์ตำแหน่งที่สองและสามได้รับการจดจำโดยไซต์ YxxCxxxF และ GTS กระบวนการนี้มีความสำคัญมากเพราะทำให้ eRF1 มีความสามารถในการแยกแยะโคดอนหยุดจากโคดอนเริ่มต้นยูราซิลที่เข้ารหัสกรดอะมิโน[ 28 ]ตัวอย่างเช่น กรดอะมิโนซิสทีนถูกเข้ารหัสโดยโคดอน UGU ขั้นตอนแรกในการแยกแยะโคดอนหยุดจากโคดอนที่มีนัยสำคัญคือการแยกแยะพิวรีนจากไพริมิดีน เนื่องจากโคดอนหยุดทั้งหมดมีนิวคลีโอไทด์พิวรีนในตำแหน่ง +2 และ +3 กรดอะมิโนที่มีการอนุรักษ์สูงสองตัวคือ Glu55 และ Tyr125 (ตั้งอยู่ในโมทีฟ YxxCxxxF) ทำงานร่วมกันเพื่อสร้างพันธะไฮโดรเจนกับอะตอมไนโตรเจน N6 บนนิวคลีโอไทด์อะดีโนซีน/กัวนีน ปฏิสัมพันธ์นี้ตัดความเป็นไปได้ของไพริมิดีนในตำแหน่ง +2 และ +3 ออกไป[ 29 ] [ 30 ]
จำเป็นต้องมีการแยกแยะพิวรีนเพิ่มเติมในตำแหน่ง +2 และ +3 เนื่องจาก UGG เป็นรหัสพันธุกรรมสำหรับทริปโตแฟน ในกรณีของ UGG หมู่ Glu55 จะถูกผลักออกจากประจุลบที่รุนแรงจากนิวคลีโอไทด์กัวนีนสองตัว เนื่องจากไม่มีพันธะไฮโดรเจนเกิดขึ้นอย่างกว้างขวาง รหัสพันธุกรรมจึงไม่ได้รับการยอมรับว่าเป็นรหัสหยุด[ 30 ]
สารตกค้างที่สำคัญมากใน eRF1 คือ Cys127 ในโมทีฟ YxxCxxxF ซึ่งสร้างพันธะไฮโดรเจน 2 พันธะกับขอบ Watson และ Crick ที่อยู่บน mRNA พันธะไฮโดรเจนช่วยให้คอมเพล็กซ์ eRF1-stop codon มีเสถียรภาพมากขึ้นในหลายทิศทาง และช่วยให้เกิดการเรียงซ้อน/พันธะไฮโดรเจนของตำแหน่ง +2 และ +3 ของ stop codon ความแข็งแรงและจำนวนของการเรียงซ้อนในตำแหน่งที่สองและสามทำให้ eRF1 สามารถแยกแยะ stop codon ออกจาก sense codon ได้[ 30 ]
ไซต์ GTS มีความสามารถในการรับสองรูปแบบขึ้นอยู่กับปฏิสัมพันธ์ของอะดีนีนหรือกัวนีน สำหรับโคดอนหยุด UAG นั้น Thr32 จากไซต์ GTS จะสร้างพันธะไฮโดรเจนกับกัวนีนในตำแหน่ง +3 หากโคดอนหยุดมีกัวนีนในตำแหน่งที่สอง (UGA) โมทีฟ YxxCxxxF จะซ้อนทับบนโคดอน ทำให้ไซต์ GTS หันออกจากโคดอน[ 30 ] [ 29 ]
การกลายพันธุ์ที่ไร้สาระ
เกือบ 11% ของความผิดปกติทางพันธุกรรมที่ถ่ายทอดทางกรรมพันธุ์ทั้งหมดเกิดจากรหัสหยุดก่อนกำหนด (การกลายพันธุ์แบบไร้ความหมาย) ตัวอย่างที่โดดเด่นของการกลายพันธุ์แบบไร้ความหมายที่ได้รับการวิจัยอย่างกว้างขวาง ได้แก่CFTRซึ่งเป็นสาเหตุของโรคซิสติกไฟโบร ซิส และการกลายพันธุ์ของไดสโทรฟิ น ซึ่งเป็นสาเหตุ ของ โรคกล้ามเนื้อเสื่อมดูเชน การวิจัยด้านการรักษาในปัจจุบันมุ่งเน้นไปที่การบังคับให้เกิดการอ่านผ่านรหัสหยุดก่อนกำหนด ซึ่งจะทำให้สาย mRNA ที่กลายพันธุ์ก่อนหน้านี้สามารถเข้ารหัสโปรตีนที่พับตัวได้อย่างถูกต้อง วิธีทั่วไปในการบังคับให้เกิดการอ่านผ่านคือการจำกัดกิจกรรมของ eRF1 และ eRF3 กลไกหนึ่งที่เสนอคือการจำกัดความเข้มข้นของ eRF1 และ eRF3 ในเซลล์ ซึ่งในทางทฤษฎีจะช่วยลดการจดจำรหัสหยุด[ 31 ]อย่างไรก็ตาม กลไกนี้ยังไม่ได้ถูกนำมาใช้ในการรักษา แต่กลไกที่น่าสนใจที่สุดเกี่ยวข้องกับการย่อยสลายทางเคมีของ eRF1 เพื่อให้เกิดการอ่านผ่าน สารเคมีเหล่านี้อยู่ในกลุ่มยาที่เรียกว่าตัวส่งเสริมการอ่านผ่าน กลไกของโปรโมเตอร์แบบอ่านผ่านนั้นมีความหลากหลาย อย่างไรก็ตามกลไกทั่วไปคือการป้องกันไม่ให้ eRF1 หลุดออกจากไรโบโซม ส่งผลให้ไรโบโซมหยุดชะงัก จากนั้นจึงเกิดการชนกับไรโบโซมอื่นในที่สุด[ 32 ]
กลไกการยุติ
คอมเพล็กซ์การยุติ
เมื่อ eRF1 จดจำรหัสหยุดและจับกับไรโบโซมแล้ว eRF1 ก็พร้อมสำหรับขั้นตอนสุดท้ายของการยุติ: การไฮโดรไลซิสของพันธะเปปไทด์ เพื่อที่จะปลดปล่อยโพลีเปปไทด์ออกจากตำแหน่ง p ของไรโบโซม จำเป็นต้องมีโปรตีนเพิ่มเติม แหล่งพลังงาน และไอออนเพื่อช่วย eRF1 ซึ่งทำได้โดยการสร้างคอมเพล็กซ์ควอเทอร์นารี โปรตีนเพิ่มเติมคือ eRF3 ซึ่งเป็นGTPaseแหล่งพลังงานคือโมเลกุล GTP และไอออนคือ Mg 2+เมื่อ eRF3 จับกับ eRF1 แล้ว ความสัมพันธ์ของมันกับ GTP จะเพิ่มขึ้นอย่างมีนัยสำคัญเมื่อเทียบกับความสัมพันธ์ของโปรตีน eRF3 เพียงอย่างเดียว เป็นที่น่าสังเกตว่าการจดจำรหัสหยุดไม่จำเป็นต้องใช้ GTP ในขณะที่การไฮโดรไลซิสของเปปทิดิล tRNA และการปลดปล่อยคอมเพล็กซ์การยุติจำเป็นต้องใช้ GTP [ 33 ]
บทบาทของ eRF3
มีสมมติฐานมากมายเกี่ยวกับหน้าที่ของ eRF3 ในคอมเพล็กซ์การยุติ สมมติฐานแรกเริ่มคือ eRF3 ช่วยให้ eRF1 จับกับโคดอนหยุดได้ เนื่องจาก eRF3 มีโครงสร้างคล้ายกับEF-TUซึ่งเป็น GTPase ที่นำโมเลกุล tRNA ที่มีประจุไปยังตำแหน่งอะมิโนเอซิลของไรโบโซมในเซลล์โปรคาริโอต[ 34 ] [ 35 ]สมมติฐานอื่นๆ มุ่งเน้นไปที่ผลกระทบของการไฮโดรไลซิสของ GTP ซึ่งถูกควบคุมโดย eRF3 ต่อ eRF1 การกำหนดค่า GTP ก่อนไฮโดรไลซิสของคอมเพล็กซ์การยุติเอื้อต่อการจับของ eRF1 กับโคดอนหยุดและจัดวาง eRF1 ให้ตรงกับเปปไทด์ tRNA การกำหนดค่า GDP หลังไฮโดรไลซิสส่งเสริมการปลดปล่อยคอมเพล็กซ์และการแยกตัวของไรโบโซม[ 36 ]
การศึกษาเพิ่มเติมตั้งสมมติฐานว่าการไฮโดรไลซิสของ GTP เนื่องมาจาก eRF3 ช่วยให้ไซต์เร่งปฏิกิริยาของ eRF1 เข้าสู่ไซต์ p ของไรโบโซม จึงส่งเสริมการปล่อยโพลีเปปไทด์ที่เกิดขึ้นใหม่[ 33 ]
ตำแหน่งเร่งปฏิกิริยา GGQ
ตำแหน่งเร่งปฏิกิริยาที่รับผิดชอบการไฮโดรไลซิสของ peptidyl tRNA คือตำแหน่ง GGQ บน eRF1 ความเข้าใจในปัจจุบันเกี่ยวกับตำแหน่ง GGQ ระบุว่ามันเข้าสู่ตำแหน่ง P ของไรโบโซม ซึ่งเป็นที่ตั้งของ peptidyl tRNA หลังจากเกิดการเปลี่ยนแปลงโครงสร้างเนื่องจากการไฮโดรไลซิสของ GTP โดย eRF3 นอกจากนี้ การกลายพันธุ์ใดๆ ในตำแหน่ง GGQ จะทำให้ eRF1 ไม่ทำงาน ส่งผลให้เซลล์ไม่สามารถยุติการแปลรหัสได้อย่างสำเร็จ เนื่องจากกรดอะมิโนไกลซีนสองตัวใน GGQ จะทำมุมบิดที่เหมาะสม ซึ่งเป็นไปได้เฉพาะเมื่อมีไกลซีนสองตัวเท่านั้น หากไม่มีมุมที่เหมาะสม ตำแหน่งเร่งปฏิกิริยาจะไม่สามารถทำงานได้อย่างถูกต้อง เพื่อให้โปรตีนที่เกิดขึ้นใหม่ถูกปล่อยออกมา ตำแหน่ง GGQ จำเป็นต้องดึงโมเลกุลน้ำเข้าไปในตำแหน่งเร่งปฏิกิริยา วิธีการดึงโมเลกุลน้ำเพียงโมเลกุลเดียวยังไม่เป็นที่เข้าใจอย่างสมบูรณ์ เมื่ออยู่ในตำแหน่งแล้ว โมเลกุลของน้ำจะทำหน้าที่เป็นนิวคลีโอไฟล์และโจมตีกลุ่มคาร์บอนิลของพันธะเอสเทอร์ระหว่างโปรตีนที่เกิดขึ้นใหม่และ tRNA การไฮโดรไลซิสของพันธะเอสเทอร์ทำให้โปรตีนที่เกิดขึ้นใหม่หลุดออก และไรโบโซมและคอมเพล็กซ์การยุติถูกแยกออกจากกัน[ 37 ]

การเสื่อมสลายที่เกิดจากความไร้สาระ
เนื่องจาก eRF1 มีความสามารถในการจดจำและจับกับรหัสหยุด (stop codon) จึงวิวัฒนาการมาเป็นส่วนประกอบสำคัญในกลไกการตรวจสอบคุณภาพ mRNA กลไกหนึ่งของการตรวจสอบคุณภาพ mRNA คือ กระบวนการสลายตัวที่เกิดจากรหัสหยุด (nonsense mediated decay หรือ NMD)

NMD ใช้เพื่อปกป้องเซลล์จากการสร้างโปรตีนที่ถูกตัดทอนที่เป็นอันตรายอันเป็นผลมาจากการกลายพันธุ์แบบไร้ความหมาย เมื่อเร็วๆ นี้พบว่า NMD มีผลต่อการแยกเซลล์ของเซลล์ต้นกำเนิดเนื่องจากการสลายตัวของปัจจัยการเข้ารหัส mRNA [ 39 ] [ 40 ]เส้นทาง NMD แยกความแตกต่างระหว่างรหัสหยุดก่อนกำหนด (PTC) กับรหัสหยุดปกติโดยการโจมตีเฉพาะสาย mRNA ที่ถูกตัดก่อนเท่านั้น ซึ่งหมายความว่า mRNA มีเอ็กซอนและอินทรอนอยู่ในสาย เนื่องจากกลไกของ NMD จดจำ คอมเพล็กซ์ รอยต่อเอ็กซอน แตกต่างจากการยุติการแปล NMD ใช้คอมเพล็กซ์โปรตีนระดับกลางจำนวนมากเพื่อให้เกิดการสลายตัวของ mRNA ขั้นตอนเริ่มต้นของ NMD คือการสร้างคอมเพล็กซ์ SURP คอมเพล็กซ์นี้ประกอบด้วยโปรตีน 4 ชนิด ได้แก่ SMG-1, Upf1, eRF1 และ eRF3 (SURF) คอมเพล็กซ์นี้เกิดขึ้นเมื่อ Upf1 จับกับ SMG-1 จากนั้น SMG-1 จะจับกับคอมเพล็กซ์การยุติ eRF1 และ eRF3 [ 41 ] [ 42 ]จากนั้นคอมเพล็กซ์ SURF จะเชื่อมโยงกับคอมเพล็กซ์ปลายน้ำซึ่งประกอบด้วย Upf2, Upf3 และ EJC เพื่อสร้างคอมเพล็กซ์ใหม่: คอมเพล็กซ์เหนี่ยวนำการสลายตัว (DECID) เมื่อคอมเพล็กซ์ทั้งสองรวมกัน คอมเพล็กซ์ DECID จะแยก eRF1, eRF3 และไรโบโซมออก[ 43 ]คอมเพล็กซ์ใหม่นี้ประกอบด้วย EJC, Upf2, Upf3, SMG-1 และ Upf1 ที่ถูกฟอสฟอริเลต โปรตีน Upf1 ที่ถูกฟอสฟอริเลตจะดึงดูดโปรตีน SMG เพิ่มเติม ซึ่งอยู่ใน ตระกูลเอนไซม์ เอนโดนิวคลีเอสจากนั้นโปรตีน SMG จะตัดสาย mRNA ใกล้กับโคดอนหยุดก่อนกำหนด เหตุการณ์นี้โดยพื้นฐานแล้วคือการกำจัดกลุ่มหัวป้องกันบนสาย mRNA ซึ่งจะทำให้ส่วนที่เหลือของสายถูกย่อยสลายโดยเอ็กโซโซม[ 44 ]
eRF1 การเฝ้าระวัง mRNA อิสระ
NMD ไม่ใช่เส้นทางเดียวสำหรับการตรวจสอบ mRNA เส้นทาง No-Go Decay (NGD) ใช้ในการย่อยสลายสาย mRNA ที่ไม่มีรหัสหยุดที่ใช้งานได้ กลไกนี้ใช้โปรตีนสองชนิดคือ Dom34p และ Hbs1p ซึ่งคล้ายกับ eRF1 และ eRF3 ตามลำดับ โปรตีน Dom34p และ Hbs1p จะจดจำไรโบโซมที่หยุดชะงักเพื่อกระตุ้นการตัดด้วยเอนโดนิวคลีโอไทด์[ 45 ] Non -Stop Decay (NSD) เป็นอีกเส้นทางหนึ่งที่จัดการกับสาย mRNA ที่ไม่มีรหัสหยุดที่ใช้งานได้ กลไกนี้ไม่รวม eRF1 แต่รวมโปรตีน Ski7p ที่เป็นโฮโมล็อกของ eRF3 กลไกนี้ขึ้นอยู่กับการสังเคราะห์หางโพลี-A ซึ่งทำให้ไรโบโซมหยุดชะงัก จากนั้นไรโบโซมที่หยุดชะงักจะถูกจดจำโดย Ski7p เพื่อย่อยสลาย[ 46 ] [ 47 ]
อ่านเพิ่มเติม
- Kashima I, Yamashita A, Izumi N, Kataoka N, Morishita R, Hoshino S และคณะ (กุมภาพันธ์ 2549) "การจับกันของคอมเพล็กซ์ SMG-1-Upf1-eRF1-eRF3 (SURF) แบบใหม่กับคอมเพล็กซ์จุดเชื่อมต่อเอ็กซอน กระตุ้นการฟอสโฟรีเลชันของ Upf1 และการสลายตัวของ mRNA ที่เกิดจากรหัสหยุด" Genes & Development 20 ( 3): 355– 367. doi : 10.1101/gad.1389006 . PMC 1361706 . PMID 16452507 .
- Chavatte L, Seit-Nebi A, Dubovaya V, Favre A (ตุลาคม 2545). "ยูริดีนที่ไม่เปลี่ยนแปลงของรหัสหยุดสัมผัสกับลูป NIKSR ที่ได้รับการอนุรักษ์ของ eRF1 ในไรโบโซมของมนุษย์"วารสารEMBO 21 ( 19): 5302– 5311. doi : 10.1093/emboj/cdf484 . PMC 129024 . PMID 12356746 .
- Janzen DM, Geballe AP (2004). "ผลของการลดปริมาณปัจจัยปลดปล่อยยูคาริโอตต่อการยุติการแปลในสายเซลล์มนุษย์" . Nucleic Acids Research . 32 (15): 4491– 4502. doi : 10.1093/nar/gkh791 . PMC 516063 . PMID 15326224 .
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N และคณะ (ตุลาคม 2548). "มุ่งสู่แผนที่ระดับโปรตีโอมของเครือข่ายปฏิสัมพันธ์ระหว่างโปรตีนของมนุษย์" Nature . 437 (7062): 1173– 1178. Bibcode : 2005Natur.437.1173R . doi : 10.1038/nature04209 . PMID 16189514 . S2CID 4427026 .
- Ivanova EV, Alkalaeva EZ, Birsdall B, Kolosov PM, Pol'shakov VI, Kiselev LL (2008) "[ส่วนต่อประสานของอันตรกิริยาของโดเมนกลางของปัจจัยการยุติการแปลโดยมนุษย์ eRF1 กับไรโบโซมยูคาริโอต]" อณูคูเลียร์ไนอา ไบโอโลจิยา . 42 (6): 1,056– 1,066 ดอย : 10.1134/ S0026893308060162 PMID 19140327 . S2CID 38843938 .
- Hauryliuk V, Zavialov A, Kisselev L, Ehrenberg M (กรกฎาคม 2549). "ปัจจัยปลดปล่อยคลาส 1 eRF1 ส่งเสริมการจับ GTP โดยปัจจัยปลดปล่อยคลาส 2 eRF3". Biochimie . 88 (7): 747– 757. doi : 10.1016/j.biochi.2006.06.001 . PMID 16797113 .
- Inge-Vechtomov S, Zhouravleva G, Philippe M (2003). "ประวัติของปัจจัยปลดปล่อยยูคาริโอต (eRFs)"ชีววิทยาของเซลล์95 ( 3– 4): 195– 209. doi : 10.1016/S0248-4900(03)00035-2 . PMID 12867083 . S2CID 19468756 .
- Andér M, Aqvist J (เมษายน 2552). "การเมทิลเลชันของกลูตามีนส่งผลต่อโครงสร้างภายในของโมทีฟ GGQ ที่ได้รับการอนุรักษ์ไว้ทั่วโลกในปัจจัยปลดปล่อยไรโบโซมหรือไม่?" ชีวเคมี48 ( 15): 3483– 3489. doi : 10.1021/bi900117r . PMID 19265422 .
- Kobayashi Y, Zhuang J, Peltz S, Dougherty J (มิถุนายน 2010). "การระบุปัจจัยเซลล์ที่ปรับเปลี่ยนการเลื่อนเฟรมของไรโบโซมตามโปรแกรมของ HIV-1"วารสารเคมีชีวภาพ 285 ( 26): 19776– 19784. doi : 10.1074/jbc.M109.085621 . PMC 2888388 . PMID 20418372 .
- Sowa ME, Bennett EJ, Gygi SP, Harper JW (กรกฎาคม 2552). "การกำหนดภูมิทัศน์ปฏิสัมพันธ์ของเอนไซม์ดีอุบิควิตินเนตของมนุษย์" . Cell . 138 (2): 389– 403. doi : 10.1016/j.cell.2009.04.042 . PMC 2716422 . PMID 19615732 .
- Ilegems E, Pick HM, Vogel H (ธันวาคม 2004). "การลดระดับ eRF1 โดยการแทรกแซง RNA เพิ่มประสิทธิภาพการยับยั้ง tRNA ที่มีการเติมหมู่แอซิลผิดพลาดในเซลล์มนุษย์"วิศวกรรมโปรตีน การออกแบบและการคัดเลือก 17 ( 12): 821– 827. doi : 10.1093/protein/gzh096 . PMID 15716307 .
- Kolosov P, Frolova L, Seit-Nebi A, Dubovaya V, Kononenko A, Oparina N, และคณะ (2548) "กรดอะมิโนคงที่ที่จำเป็นสำหรับการถอดรหัสฟังก์ชันของโพลีเปปไทด์รีลีสแฟกเตอร์ eRF1 " การวิจัยกรดนิวคลีอิก33 (19): 6418– 6425. ดอย : 10.1093/nar/ gki927 PMC 1283522 . PMID 16282590 .
- Andersen JS, Lam YW, Leung AK, Ong SE, Lyon CE, Lamond AI, Mann M (มกราคม 2548). "พลวัตของโปรตีนในนิวเคลียส" Nature . 433 (7021): 77– 83. Bibcode : 2005Natur.433...77A . doi : 10.1038/nature03207 . PMID 15635413 . S2CID 4344740 .
- Figaro S, Scrima N, Buckingham RH, Heurgué-Hamard V (กรกฎาคม 2551). "โปรตีน HemK2 ที่เข้ารหัสบนโครโมโซม 21 ของมนุษย์ ทำการเมทิลเลชั่นปัจจัยการยุติการแปล eRF1" FEBS Letters . 582 (16): 2352– 2356. Bibcode : 2008FEBSL.582.2352F . doi : 10.1016/j.febslet.2008.05.045 . PMID 18539146 . S2CID 38589664 .
- Chavatte L, Frolova L, Laugâa P, Kisselev L, Favre A (สิงหาคม 2546). "รหัสหยุดและ UGG ส่งเสริมการจับตัวของปัจจัยปลดปล่อยโพลีเปปไทด์ eRF1 กับไซต์ A ของไรโบโซมอย่างมีประสิทธิภาพ" วารสารชีววิทยาโมเลกุล 331 ( 4): 745– 758. doi : 10.1016/S0022-2836(03)00813-1 . PMID 12909007 .
- Bohnsack MT, Regener K, Schwappach B, Saffrich R, Paraskeva E, Hartmann E, Görlich D (พฤศจิกายน 2545). "Exp5 ส่งออก eEF1A ผ่าน tRNA จากนิวเคลียสและทำงานร่วมกับเส้นทางการขนส่งอื่นๆ เพื่อจำกัดการแปลรหัสให้อยู่ในไซโตพลาสซึม"วารสารEMBO 21 ( 22): 6205– 6215. doi : 10.1093/emboj/cdf613 . PMC 137205 . PMID 12426392 .
- Gevaert K, Goethals M, Martens L, Van Damme J, Staes A, Thomas GR, Vandekerckhove J (พฤษภาคม 2546). "การสำรวจโปรตีโอมและการวิเคราะห์กระบวนการโปรตีนโดยการระบุเปปไทด์ปลาย N ที่คัดแยกด้วยสเปกโทรเมตรีมวล" Nature Biotechnology . 21 (5): 566– 569. doi : 10.1038/nbt810 . PMID 12665801 . S2CID 23783563 .
- Funakoshi Y, Doi Y, Hosoda N, Uchida N, Osawa M, Shimada I และคณะ (ธันวาคม 2550). "กลไกการกำจัดอะดีนีนของ mRNA: หลักฐานสำหรับปฏิสัมพันธ์ระดับโมเลกุลระหว่างปัจจัยการยุติการแปล eRF3 และเอนไซม์กำจัดอะดีนีนของ mRNA" Genes & Development . 21 (23): 3135– 3148. doi : 10.1101/gad.1597707 . PMC 2081979 . PMID 18056425 .
- Ivanova EV, Kolosov PM, Birdsall B, Kelly G, Pastore A, Kisselev LL, Polshakov VI (สิงหาคม 2550). "ปัจจัยยุติการแปลรหัสคลาส 1 ของยูคาริโอต eRF1 - โครงสร้าง NMR และพลวัตของโดเมนกลางที่เกี่ยวข้องกับการกระตุ้นการไฮโดรไลซิสของเปปทิดิล-tRNA ที่ขึ้นอยู่กับไรโบโซม" The FEBS Journal . 274 (16): 4223– 4237. doi : 10.1111/j.1742-4658.2007.05949.x . PMID 17651434 . S2CID 6429986 .
- Mantsyzov AB, Ivanova EV, Birdsall B, Alkalaeva EZ, Kryuchkova PN, Kelly G และคณะ (มิถุนายน 2010). "โครงสร้างและหน้าที่ของโดเมนปลาย C ของตัวปลดปล่อยโซ่โพลีเปปไทด์คลาส 1 ของยูคาริโอตโดยวิธี NMR"วารสารFEBS . 277 (12): 2611– 2627. doi : 10.1111/j.1742-4658.2010.07672.x . PMC 2984548 . PMID 20553496 .
ลิงก์ภายนอก
- Termination+Release+Factorที่ US National Library of Medicine Medical Subject Headings (MeSH)
บทความนี้ได้นำข้อความจากหอสมุดแห่งชาติสหรัฐอเมริกาด้านการแพทย์ มา ใช้ ซึ่งเป็นข้อมูลสาธารณะ
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ปัจจัยยุติการแปลรหัสยูคาริโอต 1
ปัจจัยยุติการแปลของยูคาริโอต 1 (eRF1) หรือที่เรียกอีกอย่างว่า TB3-1 หรือ SUP45L1 เป็น โปรตีน ที่ถูกเข้ารหัสโดย ยีน ERF1 ในยูคาริโอต eRF1 เป็นโปรตีนที่จำเป็นซึ่งเกี่ยวข้องกับ...
ความสำคัญ
เซลล์ทุกเซลล์จำเป็นต้องสร้างโปรตีนผ่านกระบวนการ ถอดรหัส และการแปล [ 6 ] eRF1 มีความสำคัญต่อการอยู่รอดและการบำรุงรักษาเซลล์เนื่องจากมีส่วนเกี่ยวข้องในการยุติการแปล การกลายพันธุ์ใดๆ ต่อตำแหน่งการจับและการเร่งปฏิกิริยาของ eRF1 จะทำให้การยุติการแปลไม่ถูกต้อง...
หลัก
eRF1 ประกอบด้วยสายโพลีเปปไทด์ของ กรดอะมิโน ที่มีรูปร่างคล้ายตัวอักษร Y โปรตีนนี้ประกอบด้วยโดเมนหลัก 3 โดเมน ได้แก่ ลำต้นและกิ่ง 2 กิ่ง แต่ละโดเมนมีวัตถุประสงค์เฉพาะและรูปแบบการพับที่แตกต่างกัน ซึ่งช่วยให้โปรตีนทำงานได้อย่างถูกต้อง...
ลวดลาย GGQ
ลำดับกรดอะมิโน GGQ เป็นลำดับกรดอะมิโนที่คงที่ในปัจจัยปลดปล่อย (release factors) ในสิ่งมีชีวิตทุกโดเมน ตำแหน่ง GGQ ประกอบด้วยกรดอะมิโนไกลซีน 2 ตัว ตามด้วยกรดอะมิโน กลูตา มีน 1 ตัว ใน eRF1 ตำแหน่ง GGQ อยู่ที่ กรดอะมิโน ลำดับที่ 183-185 ของพอลิเปปไทด์...