อ่าน 9 นาที
เอนโดกลิน
เอนโดกลิน (ENG) เป็นไกลโค โปรตีนเมมเบรน ชนิดที่ 1 ที่อยู่บนพื้นผิว เซลล์ และเป็นส่วนหนึ่งของคอมเพล็กซ์ ตัวรับ TGF เบต้า นอกจากนี้ยังเรียกกันทั่วไปว่า CD105, END, FLJ41744, HHT1,...
เอนโดกลิน
| ENG | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | ENG , Eng, AI528660, AI662476, CD105, Endo, S-endoglin, END, HHT1, ORW1, endoglin | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | OMIM : 131195 ; MGI : 95392 ; HomoloGene : 92 ; GeneCards : ENG ; OMA : ENG - orthologs | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




เอนโดกลิน (ENG) เป็นไกลโค โปรตีนเมมเบรนชนิดที่ 1 ที่อยู่บนพื้นผิวเซลล์ และเป็นส่วนหนึ่งของคอมเพล็กซ์ ตัวรับ TGF เบต้านอกจากนี้ยังเรียกกันทั่วไปว่า CD105, END, FLJ41744, HHT1, ORW และ ORW1 [ 5 ]มีบทบาทสำคัญในการสร้างหลอดเลือดใหม่ดังนั้นจึงทำให้เป็นโปรตีนที่สำคัญต่อการเจริญเติบโตของเนื้องอก การอยู่รอด และการแพร่กระจายของเซลล์มะเร็งไปยังตำแหน่งอื่น ๆ ในร่างกาย
ยีนและการแสดงออก
ยีนเอนโดกลินของมนุษย์ตั้งอยู่บนโครโมโซม 9 ของมนุษย์ โดยมีตำแหน่งของแถบไซโตเจนิกอยู่ที่ 9q34.11 [ 6 ] [ 7 ]ไกลโคโปรตีนเอนโดกลินถูกเข้ารหัสโดย 39,757 bp และแปลเป็นกรดอะมิโน 658 ตัว[ 5 ]
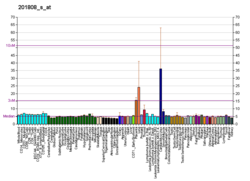
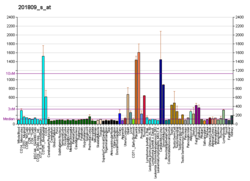
การแสดงออกของยีนเอนโดกลินมักจะต่ำในเซลล์เอนโดธีเลียมที่อยู่ในสภาวะพัก อย่างไรก็ตาม สถานการณ์จะเปลี่ยนไปเมื่อเริ่มมีการสร้างหลอดเลือดใหม่ และเซลล์เอนโดธีเลียมจะทำงานในบริเวณต่างๆ เช่น หลอดเลือดของเนื้องอก เนื้อเยื่อที่อักเสบ ผิวหนังที่เป็นโรคสะเก็ดเงิน การบาดเจ็บของหลอดเลือด และในระหว่างการเจริญเติบโตของตัวอ่อน[ 5 ] การแสดงออกของระบบหลอดเลือดเริ่มขึ้นเมื่ออายุประมาณ 4 สัปดาห์และดำเนินต่อไปหลังจากนั้น[ 5 ]เซลล์อื่นๆ ที่มีการแสดงออกของเอนโดกลิน ได้แก่ โมโนไซต์ โดยเฉพาะอย่างยิ่งเซลล์ที่กำลังเปลี่ยนไปเป็นแมโครฟาจ การแสดงออกในระดับต่ำในเซลล์กล้ามเนื้อเรียบปกติ การแสดงออกในระดับสูงในเซลล์กล้ามเนื้อเรียบของหลอดเลือด และในเนื้อเยื่อไตและตับที่กำลังเกิดพังผืด[ 5 ] [ 8 ]
โครงสร้าง
ไกลโคโปรตีนประกอบด้วยโฮโมไดเมอร์ขนาด 180 kDA ที่เสถียรด้วยพันธะไดซัลไฟด์ ระหว่างโมเลกุล [ 9 ]มีโดเมนภายนอกเซลล์ขนาดใหญ่ประมาณ 561 กรดอะมิโน โดเมนทรานส์เมมเบรนแบบไฮโดรโฟบิก และโดเมนหางไซโตพลาสมิกสั้นๆ ที่ประกอบด้วยกรดอะมิโน 45 ตัว[ 9 ]บริเวณ 260 กรดอะมิโนที่อยู่ใกล้กับเยื่อหุ้มเซลล์ภายนอกมากที่สุดเรียกว่าโดเมน ZP (หรือที่ถูกต้องกว่าคือโมดูล ZP ) [ 10 ] [ 11 ]บริเวณภายนอกเซลล์ที่อยู่นอกสุดเรียกว่าโดเมนกำพร้า (หรือที่ถูกต้องกว่าคือบริเวณกำพร้า (OR)) และเป็นส่วนที่จับกับลิแกนด์เช่นBMP- 9 [ 12 ] [ 13 ]
เอนโดกลินมีไอโซฟอร์ม 2 ชนิดที่เกิดจากการสลับการตัดต่อยีน ได้แก่ ไอโซฟอร์มยาว (L-เอนโดกลิน) และไอโซฟอร์มสั้น (S-เอนโดกลิน) [ 14 ]อย่างไรก็ตาม ไอโซฟอร์ม L มีการแสดงออกมากกว่าไอโซฟอร์ม S เอนโดกลินในรูปแบบที่ละลายน้ำได้สามารถผลิตได้จากการตัดแยกโปรตีนโดยเมทัลโลโปรตีเนส MMP-14 ในโดเมนภายนอกเซลล์ใกล้กับเยื่อหุ้มเซลล์[ 5 ] พบได้ในเซลล์บุผนังหลอดเลือดในเนื้อเยื่อทั้งหมด[ 9 ]แมโครฟาจที่ถูกกระตุ้นโมโนไซต์ที่ถูกกระตุ้น ลิมโฟบลาสต์ ไฟโบรบ ลาสต์ และเซลล์กล้ามเนื้อเรียบเอนโดกลินถูกระบุครั้งแรกโดยใช้โมโนโคลนอลแอนติบอดี (mAb) 44G4 แต่มีการค้นพบ mAb ต่อต้านเอนโดกลินมากขึ้น ทำให้มีวิธีการระบุในเนื้อเยื่อได้มากขึ้น[ 15 ]
มีการเสนอแนะว่าเอนโดกลินมีตำแหน่งไกลโคซิเลชันแบบ N-linked ที่เป็นไปได้ 5 ตำแหน่งในโดเมน N-terminal (ซึ่ง N102 ได้รับการสังเกตจากการทดลองในโครงสร้างผลึกของบริเวณที่ไม่มีคู่ ( PDB : 5I04 )) และโดเมน O-glycan ใกล้กับโดเมนเยื่อหุ้มเซลล์ซึ่งอุดมไปด้วยซีรีนและทรีโอนีน[ 9 ]หางไซโตพลาสมิกมีโมทีฟที่จับกับ PDZ ซึ่งช่วยให้สามารถจับกับโปรตีนที่มี PDZ และโต้ตอบกับพวกมันได้[ 16 ]มันมีลำดับไตรเปปไทด์อาร์จินีน-ไกลซีน-แอสปาร์ติกแอซิด (RGD) ที่ช่วยให้เกิดการยึดเกาะของเซลล์ผ่านการจับกับอินทิกรินหรือตัวรับที่จับกับ RGD อื่นๆ ที่มีอยู่ในเมทริกซ์นอกเซลล์ (ECM) [ 9 ] ลำดับ RGD บนเอนโดกลินนี้เป็นลำดับ RGD แรกที่ระบุได้ในเนื้อเยื่อบุผนังหลอดเลือด[ 9 ]
โครงสร้างผลึกเอกซ์เรย์ของเอนโดกลินของมนุษย์ ( PDB : 5I04 , 5HZV ) และสารประกอบเชิงซ้อนกับลิแกนด์BMP-9 ( PDB : 5HZW ) เผยให้เห็นว่าบริเวณที่ไม่มีคู่ของโปรตีน (กรดอะมิโน E26-S337) ประกอบด้วยสองโดเมน (OR1 และ OR2 ซึ่งสอดคล้องกับกรดอะมิโน E36-T46 + T200-C330 และกรดอะมิโน S47-R199 ตามลำดับ) ที่มีโครงสร้างพับแบบใหม่ซึ่งเป็นผลมาจากการจำลองยีนและการเรียงลำดับแบบวงกลม[ 13 ]โมดูลZP (สารตกค้าง P338-G581) ซึ่งมีส่วนประกอบ ZP-N และ ZP-C (สารตกค้าง T349-L443 และ N444-S576 ตามลำดับ) เรียงชิดกันอย่างใกล้ชิด ทำหน้าที่เป็นตัวกลางในการสร้างโฮโมไดเมอร์ของเอนโดกลินโดยการสร้างพันธะไดซัลไฟด์ระหว่างโมเลกุลที่เกี่ยวข้องกับซิสเทอีน 516 [ 13 ]เมื่อรวมกับพันธะไดซัลไฟด์ระหว่างโมเลกุลที่สองซึ่งเกี่ยวข้องกับซิสเทอีน 582 [ 16 ]จะสร้างแคลมป์โมเลกุลที่ยึดลิแกนด์ไว้โดยผ่านปฏิสัมพันธ์ของ OR1 สองชุดกับบริเวณข้อต่อของBMP-9โฮ โมไดเมอร์ [ 13 ]นอกจากจะอธิบายการกลายพันธุ์ HHT1 จำนวนมากแล้ว โครงสร้างผลึกของเอนโดกลินยังแสดงให้เห็นว่าเอพิโทปของแอนติบอดีโมโนโคลนอลต่อต้าน ENG TRC105 ทับซ้อนกับตำแหน่งการจับของBMP- 9 [ 13 ]
ปฏิสัมพันธ์
พบว่าเอนโดกลินมีปฏิสัมพันธ์กับตัวรับ TGF เบต้า 3 ที่ มีความสัมพันธ์สูง [ 6 ] [ 17 ]และตัวรับ TGF เบต้า 1 [ 16 ] [ 18 ]และมีความสัมพันธ์ต่ำกว่ากับตัวรับ TGF เบต้า 2 [ 6 ] เอนโดกลินมีความคล้ายคลึงกันในลำดับสูงกับโปรตีนที่จับกับ TGF เบต้าอีกตัวหนึ่งคือ เบตาไกลแคน ซึ่งเป็นหนึ่งในเบาะแสแรกที่บ่งชี้ว่าเอนโดกลินเป็นโปรตีนที่จับกับ TGF เบต้า[ 19 ]อย่างไรก็ตาม พบว่า TGF เบต้าจับกับเอนโดกลินที่มีอยู่เพียงเล็กน้อยด้วยความสัมพันธ์สูง ซึ่งแสดงให้เห็นว่ามีปัจจัยอื่นที่ควบคุมการจับนี้[ 19 ]
เอนโดกลินเองไม่ได้จับกับลิแกนด์ TGF เบต้า แต่จะปรากฏอยู่กับตัวรับ TGF เบต้าเมื่อลิแกนด์จับอยู่ ซึ่งบ่งชี้ถึงบทบาทสำคัญของเอนโดกลิน[ 16 ]เอนโดกลินแบบเต็มความยาวจะจับกับคอมเพล็กซ์ตัวรับ TGF เบต้าไม่ว่า TGF เบต้าจะจับอยู่หรือไม่ก็ตาม แต่เอนโดกลินในรูปแบบที่ถูกตัดทอนจะมีการจับที่จำเพาะเจาะจงมากกว่า[ 16 ]บริเวณกรดอะมิโน (aa) 437–558 ในโดเมนภายนอกเซลล์ของเอนโดกลินจะจับกับตัวรับ TGF เบต้า II ตัวรับ TGF เบต้า I จับกับบริเวณ aa 437-588 และบริเวณ aa ระหว่าง 437 กับปลาย N [ 16 ]แตกต่างจากตัวรับ TGF เบต้า I ซึ่งสามารถจับกับหางไซโตพลาสมิกได้เฉพาะเมื่อโดเมนไคเนสไม่ทำงานเท่านั้น ตัวรับ TGF เบต้า II สามารถจับกับเอนโดกลินได้ทั้งที่มีโดเมนไคเนสที่ไม่ทำงานและทำงานอยู่[ 16 ]ไคเนสจะทำงานเมื่อได้รับการฟอสโฟรีเลต นอกจากนี้ ตัวรับ TGF เบต้า I จะแยกตัวออกจากเอนโดกลินหลังจากที่ฟอสโฟรีเลตส่วนหางไซโตพลาสมิกของมัน ทำให้ตัวรับ TGF เบต้า I ไม่ทำงาน[ 16 ]เอนโดกลินได้รับการฟอสโฟรีเลตอย่างต่อเนื่องที่หมู่เซรินและทรีโอนีนในโดเมนไซโตพลาสมิก ปฏิสัมพันธ์ที่สูงระหว่างส่วนหางไซโตพลาสมิกและส่วนหางนอกเซลล์ของเอนโดกลินกับคอมเพล็กซ์ตัวรับ TGF เบต้า บ่งชี้ถึงบทบาทสำคัญของเอนโดกลินในการปรับเปลี่ยนการตอบสนองของ TGF เบต้า เช่น การกำหนดตำแหน่งของเซลล์และการเคลื่อนย้ายของเซลล์[ 16 ]
นอกจากนี้ Endoglin ยังสามารถเป็นตัวกลางในการเปลี่ยนแปลงของ F-actin, การยึดเกาะแบบโฟกัส, โครงสร้างไมโครทูบูลาร์ , การขนส่งเวสิเคิลเอนโดไซติกผ่านการโต้ตอบกับ zyxin, ZRP-1, beta-arrestin และ Tctex2beta, LK1, ALK5, ตัวรับ TGF beta II และ GIPC [ 5 ]ในการศึกษาหนึ่งกับไฟโบรบลาสต์ของหนู การแสดงออกของ Endoglin ที่มากเกินไปส่งผลให้ส่วนประกอบ ECM บางอย่างลดลง การเคลื่อนที่ของเซลล์ลดลง การเปลี่ยนแปลงรูปร่างของเซลล์ และการก่อตัวของกลุ่มระหว่างเซลล์[ 20 ]
การทำงาน
พบว่าเอนโดกลินเป็นตัวรับเสริมสำหรับคอมเพล็กซ์ตัวรับ TGF-beta [ 16 ] ดังนั้นจึงมีส่วนเกี่ยวข้องในการปรับการตอบสนองต่อการจับของTGF-beta1 , TGF-beta3 , แอคติวิน -A, BMP-2 , BMP-7และ BMP-9 นอกจากการส่งสัญญาณ TGF-beta แล้วเอนโดกลินอาจมีหน้าที่อื่นๆ อีก มีการตั้งสมมติฐานว่าเอนโดกลินมีส่วนเกี่ยวข้องกับการจัดระเบียบโครงสร้างเซลล์ที่มีผลต่อรูปร่างและการเคลื่อนที่ของเซลล์[ 21 ] เอนโดกลินมีบทบาทในการพัฒนาของระบบหัวใจและหลอดเลือดและการปรับโครงสร้างหลอดเลือด การแสดงออกของมันถูกควบคุมในระหว่างการพัฒนาของหัวใจ หนูทดลองที่ไม่มียีนเอนโดกลินจะตายเนื่องจากความผิดปกติของระบบหัวใจและหลอดเลือด[ 21 ]
ความสำคัญทางคลินิก
ในมนุษย์ เอนโดกลินอาจมีส่วนเกี่ยวข้องกับ โรค ทางพันธุกรรมแบบออโตโซมั ลโดมิแนนต์ ที่เรียกว่าโรคหลอดเลือดฝอยแตกง่ายทางพันธุกรรม (HHT) ชนิดที่ 1 [ 9 ] HHT เป็นโรคแรกในมนุษย์ที่เชื่อมโยงกับคอมเพล็กซ์ตัวรับ TGF เบต้า[ 22 ]ภาวะนี้ทำให้เกิดเลือดกำเดาไหลบ่อย หลอดเลือดฝอยแตกง่ายบนผิวหนังและเยื่อบุ และอาจทำให้เกิดความผิดปกติของหลอดเลือดแดงและหลอดเลือดดำในอวัยวะต่างๆ รวมถึงสมอง ปอด และตับ
การกลายพันธุ์ที่ทำให้เกิดโรค HHT
การกลายพันธุ์บางอย่างที่นำไปสู่ความผิดปกตินี้ได้แก่: [ 22 ]
- การแทนที่ไซโตซีน (C) ด้วยกัวนีน (G) ซึ่งเปลี่ยนไทโรซีนให้เป็นรหัสหยุด
- การลบเบสคู่ 39 คู่
- การลบเบสคู่ 2 คู่ ซึ่งทำให้เกิดรหัสหยุดก่อนกำหนด
พบว่าระดับเอนโดกลินสูงขึ้นในหญิงตั้งครรภ์ที่ต่อมาเป็นโรคครรภ์เป็นพิษ[ 23 ]
บทบาทในโรคมะเร็ง
บทบาทของเอนโดกลินในการสร้างหลอดเลือดใหม่[ 24 ]และการปรับเปลี่ยนการส่งสัญญาณของตัวรับ TGF เบต้า ซึ่งเป็นตัวกลางในการกำหนดตำแหน่งของเซลล์ การเคลื่อนย้ายของเซลล์ รูปร่างของเซลล์ การแพร่กระจายของเซลล์ การสร้างกลุ่มเซลล์ ฯลฯ ทำให้เอนโดกลินเป็นผู้เล่นสำคัญในการเจริญเติบโตและการแพร่กระจายของเนื้องอก[ 25 ] [ 26 ]การสามารถกำหนดเป้าหมายและลดหรือหยุดการสร้างหลอดเลือดใหม่ในเนื้องอกได้อย่างมีประสิทธิภาพจะช่วยป้องกันการแพร่กระจายของเซลล์มะเร็งปฐมภูมิไปยังส่วนอื่นๆ ของร่างกาย[ 25 ]นอกจากนี้ ยังมีข้อเสนอแนะว่าเอนโดกลินสามารถใช้สำหรับการถ่ายภาพเนื้องอกและการพยากรณ์โรคได้[ 25 ]
บทบาทของเอนโดกลินในมะเร็งอาจมีความขัดแย้งกันในบางครั้ง เนื่องจากจำเป็นต่อการสร้างหลอดเลือดใหม่ในเนื้องอก ซึ่งจำเป็นต่อการเจริญเติบโตและการอยู่รอดของเนื้องอก แต่การลดลงของการแสดงออกของเอนโดกลินในมะเร็งหลายชนิดมีความสัมพันธ์กับผลลัพธ์ที่ไม่ดีของมะเร็งนั้น[ 5 ]ตัวอย่างเช่น ในมะเร็งเต้านม การลดลงของเอนโดกลินในรูปแบบสมบูรณ์ และการเพิ่มขึ้นของเอนโดกลินในรูปแบบที่ละลายได้มีความสัมพันธ์กับการแพร่กระจายของเซลล์มะเร็ง[ 27 ] คอมเพล็กซ์ตัวรับ TGF เบต้า-เอนโดกลินยังส่งสัญญาณที่ขัดแย้งกันจาก TGF เบต้าด้วย TGF เบต้าสามารถทำหน้าที่เป็นตัวยับยั้งเนื้องอกในระยะก่อนเป็นมะเร็งของเนื้องอกที่ไม่ร้ายแรงโดยการยับยั้งการเจริญเติบโตและกระตุ้นให้เกิดอะพอพโทซิส[ 5 ] อย่างไรก็ตาม เมื่อเซลล์มะเร็งผ่านขั้นตอน Hallmarks of Cancer และสูญเสียการตอบสนองการเจริญเติบโตที่ยับยั้งแล้ว TGF beta จะเป็นตัวกลางในการบุกรุกเซลล์ การสร้างหลอดเลือดใหม่ (โดยความช่วยเหลือของเอนโดกลิน) การหลีกเลี่ยงระบบภูมิคุ้มกัน และองค์ประกอบ ECM ของเซลล์ ทำให้เซลล์เหล่านั้นกลายเป็นมะเร็งได้[ 5 ]
มะเร็งต่อมลูกหมากและการแสดงออกของเอนโดกลิน
มีการแสดงให้เห็นว่าการแสดงออกของเอนโดกลินและการหลั่ง TGF-beta ลดลงในเซลล์สโตรมาของไขกระดูกเมื่อนำมาเพาะเลี้ยงร่วมกับเซลล์มะเร็งต่อมลูกหมาก[ 28 ]นอกจากนี้ เส้นทางการส่งสัญญาณ TGF-beta/bone morphogenic protein (BMP) ซึ่งรวมถึง Smad1 และ Smad2/3 ก็ลดลงเช่นกัน พร้อมกับการถอดรหัสยีนที่ขึ้นอยู่กับ Smad [ 28 ]ผลลัพธ์อีกประการหนึ่งในการศึกษานี้คือ การแสดงออกของสารยับยั้งการจับกับ DNA ที่ขึ้นอยู่กับ Smad1/5/8 และสารยับยั้งการทำงานของพลาสมิโนเจนที่ขึ้นอยู่กับ Smad2/3 มีการลดลงของการแสดงออกและการแพร่กระจายของเซลล์[ 28 ]ในที่สุด เซลล์มะเร็งต่อมลูกหมากที่เพาะเลี้ยงร่วมกันได้เปลี่ยนแปลงการส่งสัญญาณ TGF-beta ในเซลล์สโตรมาของกระดูก ซึ่งชี้ให้เห็นว่าการปรับเปลี่ยนนี้เป็นกลไกของการแพร่กระจายของมะเร็งต่อมลูกหมากที่ช่วยส่งเสริมการเจริญเติบโตและการอยู่รอดในสโตรมาของกระดูกที่ตอบสนอง[ 28 ]การศึกษานี้เน้นย้ำถึงความสำคัญของเอนโดกลินในเส้นทางการส่งสัญญาณ TGF-beta ในเซลล์ประเภทอื่นนอกเหนือจากเซลล์เยื่อบุผนังหลอดเลือด
ในฐานะเป้าหมายของยา
TRC105เป็นแอนติบอดีทดลองที่มุ่งเป้าไปที่เอนโดกลินเพื่อ ใช้เป็นการรักษา ต้านการสร้างหลอดเลือดใหม่สำหรับมะเร็งเนื้อเยื่ออ่อน[ 29 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
- ข้อมูลเกี่ยวกับโรค Hereditary Hemorrhagic Telangiectasia จาก GeneReviews/NCBI/NIH/UW
- CD105+Antigen ใน หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เอนโดกลิน
เอนโดกลิน (ENG) เป็นไกลโค โปรตีนเมมเบรน ชนิดที่ 1 ที่อยู่บนพื้นผิว เซลล์ และเป็นส่วนหนึ่งของคอมเพล็กซ์ ตัวรับ TGF เบต้า นอกจากนี้ยังเรียกกันทั่วไปว่า CD105, END, FLJ41744, HHT1,...
ยีนและการแสดงออก
ยีนเอนโดกลินของมนุษย์ตั้งอยู่บนโครโมโซม 9 ของมนุษย์ โดยมีตำแหน่งของแถบไซโตเจนิกอยู่ที่ 9q34.11 [ 6 ] [ 7 ] ไกลโคโปรตีนเอนโดกลินถูกเข้ารหัสโดย 39,757 bp และแปลเป็นกรดอะมิโน 658 ตัว [ 5 ]
โครงสร้าง
ไกลโคโปรตีนประกอบด้วยโฮโม ไดเมอร์ ขนาด 180 kDA ที่เสถียรด้วยพันธะ ไดซัลไฟด์ ระหว่างโมเลกุล [ 9 ] มีโดเมนภายนอกเซลล์ขนาดใหญ่ประมาณ 561 กรดอะมิโน โดเมนทรานส์เมมเบรนแบบไฮโดรโฟบิก และโดเมนหางไซโตพลาสมิกสั้นๆ ที่ประกอบด้วยกรดอะมิโน 45 ตัว [ 9 ] บริเวณ 260...
ปฏิสัมพันธ์
พบว่าเอนโดกลินมี ปฏิสัมพันธ์ กับ ตัวรับ TGF เบต้า 3 ที่ มีความสัมพันธ์สูง [ 6 ] [ 17 ] และ ตัวรับ TGF เบต้า 1 [ 16 ] [ 18 ] และมีความสัมพันธ์ต่ำกว่ากับ ตัวรับ TGF เบต้า 2 [ 6 ] เอน โดกลินมีความคล้ายคลึงกันในลำดับสูงกับโปรตีนที่จับกับ TGF เบต้าอีกตัวหนึ่งคือ...