อ่าน 7 นาที
แคปซิด
แค ปซิด คือเปลือกโปรตีนของ ไวรัส ซึ่งห่อหุ้ม สารพันธุกรรม ของไวรัสไว้มันประกอบด้วย หน่วยย่อยโครงสร้าง แบบโอลิโกเมอร์ (ที่ซ้ำกัน) หลายหน่วยที่ทำจาก โปรตีน เรียกว่า โปรโตเมอร์...
แคปซิด


แคปซิดคือเปลือกโปรตีนของไวรัส ซึ่งห่อหุ้ม สารพันธุกรรมของไวรัสไว้มันประกอบด้วย หน่วยย่อยโครงสร้าง แบบโอลิโกเมอร์ (ที่ซ้ำกัน) หลายหน่วยที่ทำจากโปรตีนเรียกว่าโปรโตเมอร์หน่วยย่อยทางสัณฐานวิทยาแบบ 3 มิติที่สังเกตได้ ซึ่งอาจตรงหรือไม่ตรงกับโปรตีนแต่ละตัว เรียกว่าแคปโซ เม อร์ โปรตีนที่ประกอบเป็นแคปซิดเรียกว่าโปรตีนแคปซิดหรือโปรตีนเปลือกไวรัส ( VCP ) ส่วนประกอบทางพันธุกรรมของไวรัสที่อยู่ภายในแคปซิด พร้อมกับโปรตีนแกนไวรัส ที่อาจมีอยู่บ้าง เรียกว่าแกนไวรัสแคปซิดและแกนไวรัสรวมกันเรียกว่านิวคลีโอแคปซิด (ดูเพิ่มเติมที่ไวริออน )
แคปซิดถูกจำแนกประเภทอย่างกว้างๆ ตามโครงสร้างของมัน ไวรัสส่วนใหญ่มีแคปซิดที่มีโครงสร้างแบบเกลียวหรือทรงยี่สิบหน้า[ 2 ] [ 3 ]ไวรัสบางชนิด เช่นแบคทีริโอเฟจได้พัฒนาโครงสร้างที่ซับซ้อนมากขึ้นเนื่องจากข้อจำกัดของความยืดหยุ่นและไฟฟ้าสถิต[ 4 ] รูปทรงทรงยี่สิบหน้า ซึ่งมีหน้าสามเหลี่ยมด้านเท่า 20 หน้า มีลักษณะคล้ายทรงกลมในขณะที่รูปทรงเกลียวคล้ายกับรูปทรงของสปริงโดยใช้พื้นที่ของทรงกระบอกแต่ไม่ใช่ทรงกระบอก[ 5 ] หน้าของแคปซิดอาจประกอบด้วยโปรตีนหนึ่งตัวหรือมากกว่า ตัวอย่างเช่น แคปซิดของไวรัส โรคปากและเท้าเปื่อยมีหน้าประกอบด้วยโปรตีนสามตัวชื่อ VP1–3 [ 6 ]
ไวรัสบางชนิดมีเปลือกหุ้มหมายความว่าแคปซิดถูกเคลือบด้วยเยื่อไขมันที่เรียกว่าเปลือกไวรัสเปลือกนี้ได้รับมาจากแคปซิดจากเยื่อหุ้มเซลล์ภายในของโฮสต์ของไวรัส ตัวอย่างเช่น เยื่อหุ้มนิวเคลียสชั้นใน เยื่อหุ้ม กอลจิ และ เยื่อหุ้มเซลล์ชั้นนอก[ 7 ]
เมื่อไวรัสเข้าสู่เซลล์และเริ่มจำลองตัวเองแล้ว หน่วยย่อยแคปซิดใหม่จะถูกสังเคราะห์ขึ้นโดยใช้ กลไก การสังเคราะห์โปรตีนของเซลล์ ในไวรัสบางชนิด รวมถึงไวรัสที่มีแคปซิดแบบเกลียว และโดยเฉพาะอย่างยิ่งไวรัสที่มีจีโนม RNA โปรตีนแคปซิดจะประกอบรวมกับจีโนมของไวรัส ในไวรัสชนิดอื่น โดยเฉพาะไวรัสที่ซับซ้อนกว่าที่มีจีโนม DNA แบบสองสาย โปรตีนแคปซิดจะประกอบกันเป็นโปรแคปซิด ต้นแบบที่ว่างเปล่า ซึ่งมีโครงสร้างพอร์ทัลพิเศษอยู่ที่จุดยอดจุดหนึ่งDNA ของไวรัส จะถูกส่งผ่านพอร์ทัลนี้เข้าไปในแคปซิด[ 8 ]
การวิเคราะห์โครงสร้างของสถาปัตยกรรมโปรตีนแคปซิดหลัก (MCP) ถูกนำมาใช้เพื่อจำแนกไวรัสออกเป็นสายพันธุ์ ตัวอย่างเช่น แบคทีริโอเฟจ PRD1 ไวรัสสาหร่ายParamecium bursaria Chlorella virus-1 (PBCV-1) ไวรัส มิมิไวรัสและอะเดโนไวรัส ของสัตว์เลี้ยงลูกด้วยนม ถูกจัดอยู่ในสายพันธุ์เดียวกัน ในขณะที่แบคทีริโอเฟจ DNA สองสายแบบมีหาง ( Caudovirales ) และเฮอร์พีสไวรัสอยู่ในสายพันธุ์ที่สอง[ 9 ] [ 10 ] [ 11 ] [ 12 ]
รูปทรงเฉพาะ
ทรงยี่สิบหน้า


โครงสร้างทรงยี่สิบหน้าพบได้บ่อยมากในไวรัสทรงยี่สิบหน้าประกอบด้วยหน้าสามเหลี่ยม 20 หน้าที่ถูกล้อมรอบด้วยจุดยอดห้าเหลี่ยม 12 จุด และประกอบด้วยหน่วยอสมมาตร 60 หน่วย ดังนั้น ไวรัสทรงยี่สิบหน้าจึงประกอบด้วยหน่วยย่อยโปรตีน 60N หน่วย จำนวนและการจัดเรียงของแคปโซเมอร์ในแคปซิดทรงยี่สิบหน้าสามารถจำแนกประเภทได้โดยใช้ "หลักการสมมูลเสมือน" ที่เสนอโดยDonald CasparและAaron Klug [ 13 ] เช่นเดียวกับทรงหลายเหลี่ยมของ Goldbergโครงสร้างทรงยี่สิบหน้าสามารถถือได้ว่าสร้างขึ้นจากเพนทาเมอร์และเฮกซาเมอร์ โครงสร้างสามารถจัดทำดัชนีได้ด้วยจำนวนเต็มสองจำนวนhและkโดยที่และ; โครงสร้างสามารถคิดได้ว่าก้าวhขั้นจากขอบของเพนทาเมอร์ หมุนทวนเข็มนาฬิกา 60 องศา จากนั้นก้าวkขั้นเพื่อไปยังเพนทาเมอร์ถัดไปจำนวนการสร้างสามเหลี่ยมTสำหรับแคปซิดถูกกำหนดดังนี้:



ในแผนผังนี้ แคปซิดทรงยี่สิบหน้าประกอบด้วยเพนทาเมอร์ 12 ตัว บวกเฮกซาเมอร์ 10( T − 1) ตัว[ 14 ] [ 15 ]ตัวเลขTเป็นตัวแทนของขนาดและความซับซ้อนของแคปซิด[ 16 ]ตัวอย่างทางเรขาคณิตสำหรับค่าh , kและT หลายค่า สามารถพบได้ที่รายการโพลีเฮดราจีโอเดสิกและโพลีเฮดราโกลด์เบิร์ก
มีข้อยกเว้นมากมายสำหรับกฎนี้ ตัวอย่างเช่น ไวรัสโพลีโอมาและไวรัสพาพิลโลมา มีเพนตาเมอร์แทนที่จะเป็นเฮกซาเมอร์ในตำแหน่งเฮกซาวาเลนต์บนแลตทิซ T = 7 เสมือน สมาชิกของสายพันธุ์ ไวรัส RNAสองสายรวมถึงรีโอไวรัสโรตาไวรัสและแบคทีริโอเฟจ φ6 มีแคปซิดที่สร้างขึ้นจากโปรตีนแคปซิด 120 ชุด ซึ่งสอดคล้องกับแคปซิด T = 2 หรืออาจกล่าวได้ว่าเป็นแคปซิด T = 1 ที่มีไดเมอร์ในหน่วยอสมมาตร ในทำนองเดียวกัน ไวรัสขนาดเล็กจำนวนมากมีแคปซิด T = 3 เสมือน (หรือ P = 3) ซึ่งจัดเรียงตามแลตทิซ T = 3 แต่มีโพลีเปปไทด์ที่แตกต่างกันครอบครองตำแหน่งเสมือนเทียบเท่าสามตำแหน่ง[ 17 ]
โพรเลท
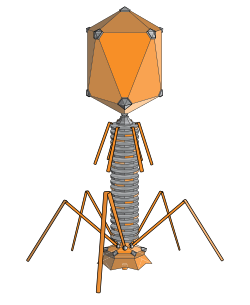
ทรงยี่สิบหน้าแบบยาวเป็นรูปทรงทั่วไปของหัวของแบคทีริโอเฟจ โครงสร้างดังกล่าวประกอบด้วยทรงกระบอกที่มีฝาปิดอยู่ที่ปลายทั้งสองข้าง ทรงกระบอกประกอบด้วยหน้าสามเหลี่ยมยาว 10 หน้า ตัวเลข Q (หรือ T mid ) ซึ่งสามารถเป็นจำนวนเต็มบวกใดๆ ก็ได้[ 18 ]ระบุจำนวนสามเหลี่ยมที่ประกอบด้วยหน่วยย่อยที่ไม่สมมาตรซึ่งประกอบเป็นสามเหลี่ยม 10 รูปของทรงกระบอก ฝาปิดจะถูกจำแนกตามตัวเลข T (หรือ T end ) [ 19 ]
แบคทีเรียE. coliเป็นโฮสต์ของแบคทีริโอเฟจ T4ซึ่งมีโครงสร้างหัวแบบยาวรี โปรตีน gp31 ที่เข้ารหัสโดยแบคทีริโอเฟจดูเหมือนจะมีหน้าที่คล้ายคลึงกับ โปรตีนชาเปอโรน GroES ของ E. coliและสามารถใช้แทนกันได้ในการประกอบไวรัสแบคทีริโอเฟจ T4 ระหว่างการติดเชื้อ[ 20 ]เช่นเดียวกับ GroES โปรตีน gp31 จะสร้างคอมเพล็กซ์ที่เสถียรกับชาเปอโรนินGroEL ซึ่งจำเป็นอย่างยิ่งสำหรับการพับและการประกอบโปรตีนแคปซิดหลัก gp23 ของแบคทีริโอเฟจ T4 ในร่างกาย[ 20 ]
เกลียว

ไวรัสพืชรูปแท่งและเส้นใยจำนวนมากมีแคปซิดที่มีสมมาตรแบบเกลียว [ 21 ] โครงสร้างเกลียวสามารถอธิบายได้ว่าเป็นชุดของ เกลียวโมเลกุล 1 มิติ nเกลียวที่สัมพันธ์กันด้วยสมมาตรแกนn เท่า [ 22 ]การแปลงเกลียวถูกจำแนกออกเป็นสองประเภท ได้แก่ ระบบเกลียวหนึ่งมิติและสองมิติ[ 22 ]การสร้างโครงสร้างเกลียวทั้งหมดขึ้นอยู่กับชุดของเมทริกซ์การแปลและการหมุนซึ่งถูกเข้ารหัสไว้ในธนาคารข้อมูลโปรตีน[ 22 ]สมมาตรแบบเกลียวแสดงด้วยสูตรP = μ x ρโดยที่μคือจำนวนหน่วยโครงสร้างต่อรอบของเกลียวρคือการเพิ่มขึ้นตามแกนต่อหน่วย และPคือระยะห่างของเกลียว โครงสร้างนี้กล่าวได้ว่าเปิดเนื่องจากลักษณะเฉพาะที่สามารถล้อมรอบปริมาตรใดๆ ได้โดยการเปลี่ยนความยาวของเกลียว[ 23 ]ไวรัสเกลียวที่เข้าใจมากที่สุดคือไวรัสโมเสกยาสูบ[ 21 ]ไวรัสเป็นโมเลกุลเดี่ยวของ RNA สายบวก โปรตีนหุ้มแต่ละอันที่อยู่ภายในเกลียวจะจับกับนิวคลีโอไทด์สามตัวของจีโนม RNA โดยพอลิเมอร์โดยรวมมีค่า μ เท่ากับ 16.33 หน่วยย่อยโปรตีนต่อรอบเกลียว[ 21 ] ไวรัสไข้หวัดใหญ่ A แตกต่างออกไปตรงที่ประกอบด้วยไรโบนิวคลีโอโปรตีนหลายตัวซึ่งจัดระเบียบ RNA ที่แบ่งส่วนให้เป็นโครงสร้างเกลียวคู่[ 24 ]
ฟังก์ชัน
หน้าที่ของแคปซิดมีดังนี้:
- ปกป้องจีโนม
- ส่งมอบจีโนม และ
- โต้ตอบกับโฮสต์
ไวรัสต้องสร้างเปลือกโปรตีนที่มั่นคงและป้องกันเพื่อปกป้องจีโนมจากสารเคมีและปัจจัยทางกายภาพที่เป็นอันตราย ซึ่งรวมถึงค่าpH หรืออุณหภูมิที่สูงหรือต่ำเกินไป และเอนไซม์ที่ย่อยสลายโปรตีนและ นิวคลีโอไท ด์ สำหรับไวรัสที่ไม่มีเปลือกหุ้ม แคปซิดเองอาจมีส่วนร่วมในการโต้ตอบกับตัวรับบนเซลล์เจ้าบ้าน ซึ่งนำไปสู่การเจาะผ่านเยื่อหุ้มเซลล์เจ้าบ้านและการนำแคปซิดเข้าไปภายใน การส่งมอบจีโนมเกิดขึ้นโดยการถอดเปลือกหรือการแยกส่วนของแคปซิดในภายหลังและการปล่อยจีโนมเข้าสู่ไซโตพลาสซึมหรือ โดยการขับจีโนมผ่านโครงสร้างช่องทางพิเศษโดยตรงเข้าไปในนิวเคลียสของเซลล์ เจ้าบ้าน
ที่มาและวิวัฒนาการ
มีการเสนอแนะว่าโปรตีนแคปซิดของไวรัสจำนวนมากได้วิวัฒนาการมาจากโปรตีนเซลล์ที่มีหน้าที่หลากหลายในหลายโอกาส[ 25 ]การนำโปรตีนเซลล์มาใช้ดูเหมือนจะเกิดขึ้นในขั้นตอนวิวัฒนาการที่แตกต่างกัน ดังนั้นโปรตีนเซลล์บางชนิดจึงถูกจับและปรับเปลี่ยนหน้าที่ก่อนการแยกตัวของสิ่งมีชีวิตเซลล์ออกเป็นสามโดเมนของสิ่งมีชีวิตในปัจจุบัน ในขณะที่โปรตีนอื่นๆ ถูกนำมาใช้เมื่อไม่นานมานี้ ผลก็คือ โปรตีนแคปซิดบางชนิดแพร่หลายในไวรัสที่ติดเชื้อสิ่งมีชีวิตที่มีความสัมพันธ์ห่างไกลกัน (เช่น โปรตีนแคปซิดที่มีโครงสร้างแบบม้วนเจลลี่ ) ในขณะที่บางชนิดจำกัดอยู่เฉพาะกลุ่มไวรัสบางกลุ่ม (เช่น โปรตีนแคปซิดของอัลฟาไวรัส) [ 25 ] [ 26 ]
แบบจำลองการคำนวณ (2015) แสดงให้เห็นว่าแคปซิดอาจมีต้นกำเนิดมาก่อนไวรัส และทำหน้าที่เป็นวิธีการถ่ายโอนแนวนอนระหว่างชุมชนจำลองตัวเอง เนื่องจากชุมชนเหล่านี้ไม่สามารถอยู่รอดได้หากจำนวนปรสิตยีนเพิ่มขึ้น โดยยีนบางตัวมีหน้าที่ในการสร้างโครงสร้างเหล่านี้ และยีนบางตัวที่เอื้อต่อการอยู่รอดของชุมชนที่จำลองตัวเองได้[ 27 ]การแทนที่ยีนบรรพบุรุษเหล่านี้ระหว่างสิ่งมีชีวิตเซลล์เดียวอาจเอื้อต่อการปรากฏของไวรัสใหม่ในระหว่างวิวัฒนาการ[ 26 ]
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Williams R (1 มิถุนายน 1979). รากฐานทางเรขาคณิตของโครงสร้างธรรมชาติ: หนังสือแหล่งข้อมูลด้านการออกแบบ . สำนักพิมพ์โดเวอร์. หน้า 142–144, รูปที่ 4-49, 50, 51: คัสเตอร์ทรงกลม 12 ลูก, ทรงกลม 42 ลูก, ทรงกลม 92 ลูก. ISBN 978-0-486-23729-9.
- Pugh A (1 กันยายน 1976). โพลีเฮดรา: แนวทางเชิงภาพ . สำนักพิมพ์มหาวิทยาลัยแคลิฟอร์เนีย. บทที่ 6. โพลีเฮดราเชิงเรขาคณิตของ R. Buckminster Fuller และโพลีเฮดราที่เกี่ยวข้อง. ISBN 978-0-520-02926-2.
- Almansour I, Alhagri M, Alfares R, Alshehri M, Bakhashwain R, Maarouf A (มกราคม 2019). "IRAM: ฐานข้อมูลแคปซิดของไวรัสและแหล่งข้อมูลการวิเคราะห์" . Database: The Journal of Biological Databases and Curation . 2019 . doi : 10.1093/database/baz079 . PMC 6637973 . PMID 31318422 .
ลิงก์ภายนอก
- IRAM - ฐานข้อมูลและแหล่งวิเคราะห์แคปซิดของไวรัสถูกเก็บถาวรเมื่อวันที่ 23 ตุลาคม 2019 ที่Wayback Machine
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ แคปซิด
แค ปซิด คือเปลือกโปรตีนของ ไวรัส ซึ่งห่อหุ้ม สารพันธุกรรม ของไวรัสไว้มันประกอบด้วย หน่วยย่อยโครงสร้าง แบบโอลิโกเมอร์ (ที่ซ้ำกัน) หลายหน่วยที่ทำจาก โปรตีน เรียกว่า โปรโตเมอร์...
ทรงยี่สิบหน้า
โครงสร้างทรงยี่สิบหน้าพบได้บ่อยมากในไวรัส ทรงยี่สิบหน้า ประกอบด้วยหน้าสามเหลี่ยม 20 หน้าที่ถูกล้อมรอบด้วยจุดยอดห้าเหลี่ยม 12 จุด และประกอบด้วยหน่วยอสมมาตร 60 หน่วย ดังนั้น ไวรัสทรงยี่สิบหน้าจึงประกอบด้วยหน่วยย่อยโปรตีน 60N หน่วย จำนวนและการจัดเรียงของ...
โพรเลท
ทรงยี่สิบหน้าแบบยาวเป็นรูปทรงทั่วไปของหัวของแบคทีริโอเฟจ โครงสร้างดังกล่าวประกอบด้วยทรงกระบอกที่มีฝาปิดอยู่ที่ปลายทั้งสองข้าง ทรงกระบอกประกอบด้วยหน้าสามเหลี่ยมยาว 10 หน้า ตัวเลข Q (หรือ T mid ) ซึ่งสามารถเป็นจำนวนเต็มบวกใดๆ ก็ได้ [ 18 ]...
เกลียว
ไวรัสพืชรูปแท่งและเส้นใยจำนวนมากมีแคปซิดที่มี สมมาตรแบบเกลียว [ 21 ] โครงสร้าง เกลียวสามารถอธิบายได้ว่าเป็นชุดของ เกลียวโมเลกุล 1 มิติ n เกลียวที่สัมพันธ์กันด้วยสมมาตรแกน n เท่า [ 22 ] การแปลงเกลียวถูกจำแนกออกเป็นสองประเภท ได้แก่ ระบบเกลียวหนึ่งมิติและสองมิติ...