อ่าน 11 นาที
จุดตรวจสอบวงจรเซลล์
จุดตรวจสอบวงจรเซลล์เป็นกลไกควบคุมในวงจรเซลล์ของยูคาริโอต ซึ่งช่วยให้มั่นใจได้ว่าวงจรเซลล์ดำเนินไปอย่างถูกต้อง...
จุดตรวจสอบวงจรเซลล์

จุดตรวจสอบวงจรเซลล์เป็นกลไกควบคุมในวงจรเซลล์ของยูคาริโอต ซึ่งช่วยให้มั่นใจได้ว่าวงจรเซลล์ดำเนินไปอย่างถูกต้อง จุดตรวจสอบแต่ละจุดทำหน้าที่เป็นจุดสิ้นสุดที่เป็นไปได้ในวงจรเซลล์ซึ่งในระหว่างนั้นจะมีการประเมินสภาวะของเซลล์ และการดำเนินไปในระยะต่างๆ ของวงจรเซลล์จะเกิดขึ้นได้ก็ต่อเมื่อมีสภาวะที่เหมาะสมเท่านั้น มีจุดตรวจสอบหลายจุดในวงจรเซลล์[ 1 ]แต่จุดตรวจสอบหลักสามจุด ได้แก่ จุดตรวจสอบ G1 หรือที่รู้จักกันในชื่อจุดตรวจสอบเริ่มต้นหรือจุดตรวจสอบการจำกัดหรือจุดตรวจสอบหลักจุดตรวจสอบ G2/Mและการเปลี่ยนผ่านจากระยะเมตาเฟสไปสู่ระยะแอนาเฟส หรือที่รู้จักกันในชื่อจุดตรวจสอบสปินเดิล [ 2 ] การดำเนินไปผ่านจุดตรวจสอบเหล่านี้ส่วนใหญ่ถูกกำหนดโดยการกระตุ้นของไคเนสที่ขึ้นอยู่กับไซคลินโดยหน่วยย่อยโปรตีน ควบคุม ที่เรียกว่าไซคลินซึ่งมีรูปแบบที่แตกต่างกันถูกผลิตขึ้นในแต่ละระยะของวงจรเซลล์เพื่อควบคุมเหตุการณ์เฉพาะที่เกิดขึ้นในนั้น[ 3 ] [ 4 ]
พื้นหลัง
สิ่งมีชีวิตทั้งหมดเป็นผลผลิตจากการเจริญเติบโตและการแบ่งเซลล์ซ้ำๆ[ 5 ]ในระหว่างกระบวนการนี้ ซึ่งเรียกว่าวัฏจักรเซลล์เซลล์จะจำลองเนื้อหาภายในแล้วแบ่งตัวออกเป็นสองส่วน จุดประสงค์ของวัฏจักรเซลล์คือการจำลอง DNA ของสิ่งมีชีวิตแต่ละชนิดอย่างแม่นยำ จากนั้นจึงแบ่งเซลล์และเนื้อหาภายในเซลล์อย่างเท่าเทียมกันระหว่างเซลล์สองเซลล์ที่เกิดขึ้น ในยูคาริโอต วัฏจักรเซลล์ประกอบด้วยสี่ขั้นตอนหลัก ได้แก่G1ซึ่งเซลล์มีการทำงานทางเมตาบอลิซึมและเจริญเติบโตอย่างต่อเนื่องระยะ Sซึ่งมีการจำลอง DNA เกิดขึ้นG2ซึ่งเซลล์ยังคงเจริญเติบโตและสังเคราะห์โปรตีนต่างๆ เพื่อเตรียมพร้อมสำหรับการแบ่งตัว และระยะ M ( ไมโทซิส ) ซึ่งโครโมโซมที่จำลองแล้ว (เรียกว่าโครมาทิดคู่ ) จะแยกออกเป็นนิวเคลียสลูกสองนิวเคลียส และเซลล์จะแบ่งออกเป็นสองเซลล์ลูก โดยแต่ละเซลล์จะมี DNA ครบชุด[ 6 ]เมื่อเปรียบเทียบกับวัฏจักรเซลล์ของยูคาริโอต วัฏจักร เซลล์ของโปรคาริโอต (ที่รู้จักกันในชื่อ การแบ่งตัวแบบไบนารี ) นั้นค่อนข้างง่ายและรวดเร็ว: โครโมโซมจำลองตัวเองจากจุดเริ่มต้นของการจำลอง เยื่อหุ้มเซลล์ใหม่ถูกสร้างขึ้น และผนังเซลล์ก่อตัวเป็นเซปตัมซึ่งแบ่งเซลล์ออกเป็นสองส่วน[ 7 ]
เนื่องจากวงจรเซลล์ของยูคาริโอตเป็นกระบวนการที่ซับซ้อน ยูคาริโอตจึงได้พัฒนาเครือข่ายของโปรตีนควบคุมที่เรียกว่าระบบควบคุมวงจรเซลล์ซึ่งทำหน้าที่ตรวจสอบและกำหนดความคืบหน้าของเซลล์ผ่านวงจรเซลล์[ 5 ]ระบบนี้ทำหน้าที่เหมือนตัวจับเวลาหรือนาฬิกา ซึ่งกำหนดเวลาที่แน่นอนให้เซลล์ใช้ในแต่ละระยะของวงจรเซลล์ ในขณะเดียวกันก็ตอบสนองต่อข้อมูลที่ได้รับจากกระบวนการที่ควบคุมด้วย จุดตรวจสอบวงจรเซลล์มีบทบาทสำคัญในระบบควบคุมโดยการตรวจจับข้อบกพร่องที่เกิดขึ้นระหว่างกระบวนการที่จำเป็น เช่นการจำลองดีเอ็นเอหรือการแยกโครโมโซมและกระตุ้นให้วงจรเซลล์หยุดชะงักเพื่อตอบสนองจนกว่าข้อบกพร่องจะได้รับการแก้ไข[ 8 ]กลไกการทำงานหลักของจุดตรวจสอบวงจรเซลล์คือการควบคุมกิจกรรมของกลุ่มโปรตีนไคเนสที่เรียกว่าไคเนสที่ขึ้นอยู่กับไซคลิน (CDK) ซึ่งจับกับโปรตีนควบคุมประเภทต่างๆ ที่เรียกว่าไซคลินโดยจะมีการสร้างและกระตุ้นคอมเพล็กซ์ไซคลิน-CDK ที่เฉพาะเจาะจงในระยะต่างๆ ของวงจรเซลล์ คอมเพล็กซ์เหล่านี้จะกระตุ้นเป้าหมายปลายทางต่างๆ เพื่อส่งเสริมหรือป้องกันการดำเนินไปของวงจรเซลล์[ 9 ]
จุดตรวจ G1 (ข้อจำกัด)
จุดตรวจสอบ G1 หรือที่รู้จักกันในชื่อจุดจำกัดในเซลล์สัตว์เลี้ยงลูกด้วยนมและจุดเริ่มต้นในยีสต์ คือจุดที่เซลล์ตัดสินใจที่จะเข้าสู่รอบการแบ่งเซลล์ เมื่อเซลล์ดำเนินไปในระยะ G1 ขึ้นอยู่กับสภาวะภายในและภายนอก เซลล์อาจชะลอระยะ G1 เข้าสู่สภาวะสงบที่เรียกว่าG0หรือดำเนินเลยจุดจำกัดไป[ 5 ]ความเสียหายของ DNA เป็นตัวบ่งชี้หลักที่ทำให้เซลล์ "หยุด" และไม่เข้าสู่รอบการแบ่งเซลล์ การตัดสินใจที่จะเริ่มต้นการแบ่งเซลล์รอบใหม่เกิดขึ้นเมื่อเซลล์กระตุ้นการถอดรหัสที่ขึ้นอยู่กับไซคลิน-CDK ซึ่งส่งเสริมการเข้าสู่ระยะ S จุดตรวจสอบนี้ทำให้มั่นใจได้ว่ากระบวนการจะดำเนินต่อไป[ 10 ]
ในช่วงต้นของ G1 จะมีตัวยับยั้งการถอดรหัสสามตัวที่เรียกว่าโปรตีนพ็อกเก็ต ซึ่งจะจับกับ ปัจจัยการถอดรหัส E2Fตระกูลยีน E2F เป็นกลุ่มของปัจจัยการถอดรหัสที่กำหนดเป้าหมายยีนหลายตัวที่สำคัญต่อการควบคุมวงจรเซลล์ รวมถึงไซคลิน CDK ตัวควบคุมจุดตรวจสอบ และโปรตีนซ่อมแซม DNA การควบคุมที่ผิดปกติของตระกูล E2F มักพบในกรณีของมะเร็ง ซึ่งเป็นหลักฐานที่แสดงว่าตระกูล E2F มีความสำคัญต่อการควบคุมการจำลองแบบ DNA และการแบ่งเซลล์อย่างเข้มงวด[ 10 ]โปรตีนพ็อกเก็ตทั้งสามตัวคือRetinoblastoma (Rb), p107 และ p130 ซึ่งจับกับปัจจัยการถอดรหัส E2F เพื่อป้องกันการผ่านจุดตรวจสอบ G1
ยีนตระกูล E2F ประกอบด้วยโปรตีนบางชนิดที่มีกลไกการกระตุ้นและโปรตีนบางชนิดที่มีกลไกการยับยั้ง P107 และ p130 ทำหน้าที่เป็นตัวยับยั้งร่วมสำหรับ E2F 4 และ E2F 5 ซึ่งทำงานเพื่อยับยั้งการถอดรหัสของปัจจัยส่งเสริม G1-to-S โปรตีนกระเป๋าตัวที่สาม Rb จับกับและยับยั้ง E2F 1, E2F 2 และ E2F 3 ซึ่งเป็นโปรตีน E2F ที่มีความสามารถในการกระตุ้น[ 10 ]
การตอบสนองเชิงบวกมีบทบาทสำคัญในการควบคุมความก้าวหน้าจากระยะ G1 ไปสู่ระยะ S โดยเฉพาะอย่างยิ่งเกี่ยวข้องกับการฟอสโฟรีเลชันของ Rb โดยโปรตีนคอมเพล็กซ์ Cyclin/CDK Rb ที่ไม่มีฟอสเฟต หรือ Rb ที่ไม่ได้รับการฟอสโฟรีเลชัน จะควบคุมการออกจากวงจรเซลล์ G0 และการแยกความแตกต่าง ในช่วงเริ่มต้นของระยะ G1 ปัจจัยการเจริญเติบโตและความเสียหายของ DNA จะส่งสัญญาณให้ระดับของไซคลิน D เพิ่มขึ้น ซึ่งจากนั้นจะจับกับ Cdk4 และ Cdk6 เพื่อสร้างคอมเพล็กซ์ CyclinD:Cdk4/6 [ 11 ]คอมเพล็กซ์นี้เป็นที่ทราบกันดีว่าทำให้ Rb ไม่ทำงานโดยการฟอสโฟรีเลชัน อย่างไรก็ตาม รายละเอียดของการฟอสโฟรีเลชันของ Rb นั้นค่อนข้างซับซ้อนและเฉพาะเจาะจงเมื่อเทียบกับความรู้ก่อนหน้านี้เกี่ยวกับจุดตรวจสอบ G1 CyclinD:Cdk4/6 จะวางฟอสเฟตเพียงหนึ่งตัว หรือโมโนฟอสโฟรีเลต Rb ที่หนึ่งในสิบสี่ตำแหน่งฟอสโฟรีเลชันที่เข้าถึงได้และเป็นเอกลักษณ์ ไอโซฟอร์มโมโนฟอสโฟรีเลตเฉพาะทั้งสิบสี่ชนิดมีความชอบในการจับกับสมาชิกในตระกูล E2F ที่แตกต่างกัน ซึ่งน่าจะเพิ่มความหลากหลายของกระบวนการเซลล์ภายในร่างกายของสัตว์เลี้ยงลูกด้วยนม[ 11 ]
E2F 4 และ E2F 5 ขึ้นอยู่กับ p107 และ p130 เพื่อรักษาตำแหน่งภายในนิวเคลียส อย่างไรก็ตาม Cyclin D:Cdk 4/6 ยังฟอสโฟรีเลต p107 และ p130 ซึ่งเป็นกระบวนการที่ปลดปล่อยการจับจาก E2F 4 และ 5 (ซึ่งจะหลุดออกไปยังไซโตพลาสซึม) และทำให้ E2F 1–3 สามารถจับกับ DNA และเริ่มต้นการถอดรหัสของ Cyclin E ได้[ 10 ]โปรตีน Rb รักษาสถานะโมโนฟอสโฟรีเลตในช่วงระยะ G1 ตอนต้น ในขณะที่ Cyclin E กำลังสะสมและจับกับ Cdk2
CyclinE:Cdk2 มีบทบาทสำคัญเพิ่มเติมในการฟอสโฟรีเลชันในช่วงการเปลี่ยนผ่านจาก G1 ไปสู่ S โดยเฉพาะอย่างยิ่ง CyclinE:Cdk2 ส่งเสริมวงจรป้อนกลับเชิงบวกซึ่งสร้างสวิตช์ "ทั้งหมดหรือไม่มีเลย" ในเครือข่ายควบคุมทางพันธุกรรมจำนวนมาก การป้อนกลับเชิงบวกทำให้มั่นใจได้ว่าเซลล์จะไม่เลื่อนไปมาระหว่างระยะต่างๆ ของวงจรเซลล์[ 12 ] Cyclin E:Cdk2 ดำเนินการฟอสโฟรีเลต Rb ที่ตำแหน่งฟอสโฟรีเลชันทั้งหมด หรือที่เรียกว่า "ไฮเปอร์ฟอสโฟรีเลต" ซึ่งทำให้มั่นใจได้ว่า Rb จะถูกปิดใช้งานอย่างสมบูรณ์ การไฮเปอร์ฟอสโฟรีเลตของ Rb ถือเป็นจุดจำกัด G1 ตอนปลาย หลังจากนั้นเซลล์จะไม่สามารถย้อนกลับไปในวงจรเซลล์ได้ ณ จุดนี้ โปรตีน E2F 1-3 จะจับกับ DNA และถอดรหัส Cyclin A และ Cdc 6 [ 11 ]
สารยับยั้งไคเนสที่ขึ้นอยู่กับไซคลิน 1B (CDKN1B) หรือที่รู้จักกันในชื่อ p27 จะจับกับและป้องกันการทำงานของ CyclinE:Cdk2 โดยการยับยั้ง อย่างไรก็ตาม เมื่อไซคลิน A สะสมและจับกับ Cdk2 พวกมันจะสร้างคอมเพล็กซ์และยับยั้ง p27 ไคเนสที่ขึ้นอยู่กับไซคลินในระยะ G1 จะทำงานร่วมกับไคเนสที่ขึ้นอยู่กับไซคลินในระยะ S โดยกำหนดเป้าหมาย p27 เพื่อการย่อยสลาย ในทางกลับกัน สิ่งนี้จะช่วยให้ Cyclin A:Cdk2 ซึ่งเป็นคอมเพล็กซ์ที่ฟอสโฟรีเลต E2F 1-3 เริ่มต้นการแยกตัวออกจากไซต์โปรโมเตอร์ DNA ทำให้ E2F 6–8 สามารถจับกับ DNA และยับยั้งการถอดรหัสได้[ 10 ] วงจรป้อนกลับเชิงลบที่ใช้ในการยับยั้งสารยับยั้ง p27 ได้สำเร็จ เป็นกระบวนการสำคัญอีกอย่างหนึ่งที่เซลล์ใช้เพื่อให้แน่ใจว่ามีการเคลื่อนที่ไปในทิศทางเดียวและไม่มีการย้อนกลับผ่านวงจรเซลล์
เมื่อเกิดความเสียหายต่อ DNA หรือเมื่อเซลล์ตรวจพบความบกพร่องใดๆ ที่ทำให้จำเป็นต้องชะลอหรือหยุดวงจรเซลล์ในระยะ G1 การหยุดชะงักจะเกิดขึ้นผ่านกลไกหลายอย่าง การตอบสนองอย่างรวดเร็วเกี่ยวข้องกับกระบวนการฟอสโฟรีเลชันที่เริ่มต้นด้วยไคเนส ATM ( Ataxia telangiectasia mutated ) หรือ ATR ( Ataxia Telangiectasia and Rad3 related ) ซึ่งทำหน้าที่เป็นเซนเซอร์ ขึ้นอยู่กับประเภทของความเสียหาย ไคเนสเหล่านี้จะฟอสโฟรีเลตและกระตุ้นไคเนสตัวรับ Chk2 และ Chk1 ตามลำดับ ซึ่งจะฟอสโฟรีเลตฟอสฟาเทส Cdc25A ต่อไป ทำให้เกิดการยูบิควิตินเนชันและการสลายตัว เนื่องจาก Cdc25A กระตุ้นคอมเพล็กซ์ไซคลิน E-CDK2 ที่กล่าวถึงก่อนหน้านี้โดยการกำจัดฟอสเฟตที่ยับยั้งออกจาก CDK2 ดังนั้นหากไม่มี Cdc25A ไซคลิน E-CDK2 จะยังคงไม่ทำงาน และเซลล์จะยังคงอยู่ในระยะ G1
เพื่อรักษาการจับกุมไว้ การตอบสนองอีกอย่างหนึ่งจะเริ่มต้นขึ้น โดยที่ Chk2 หรือ Chk1 จะฟอสโฟรีเลต p53 ซึ่งเป็นตัวยับยั้งเนื้องอก และสิ่งนี้จะทำให้ p53 มีเสถียรภาพโดยป้องกันไม่ให้มันจับกับ Mdm2 ซึ่งเป็นยูบิควิตินไลเกสที่ยับยั้ง p53 โดยการกำหนดเป้าหมายให้มันถูกย่อยสลาย p53 ที่มีเสถียรภาพแล้วจะทำหน้าที่เป็นตัวกระตุ้นการถอดรหัสของยีนเป้าหมายหลายตัว รวมถึง p21 ซึ่งเป็นตัวยับยั้งคอมเพล็กซ์ไซคลิน E-CDK2 ที่ส่งเสริมการเปลี่ยนจากระยะ G1 ไปสู่ระยะ S นอกจากนี้ กลไกอีกอย่างหนึ่งที่ทำให้ p21 ถูกกระตุ้นคือผ่านการสะสมของ p16 เพื่อตอบสนองต่อความเสียหายของ DNA p16 จะรบกวนคอมเพล็กซ์ไซคลิน D-CDK4 ทำให้ p21 หลุดออกจากคอมเพล็กซ์ ซึ่งนำไปสู่การดีฟอสโฟรีเลชันและการกระตุ้นของ Rb ซึ่งทำให้ Rb สามารถจับและยับยั้ง E2F 1–3 ได้ จึงทำให้เซลล์ไม่สามารถเปลี่ยนไปสู่ระยะ S ได้[ 13 ]เมื่อเร็วๆ นี้ บางแง่มุมของแบบจำลองนี้ได้ถูกโต้แย้ง[ 14 ]
จุดตรวจ G2

หลังจากกระบวนการจำลองดีเอ็นเอในระยะ S เซลล์จะเข้าสู่ระยะการเจริญเติบโตที่เรียกว่า G2 ในช่วงเวลานี้ โปรตีนไมโทซิสที่จำเป็นจะถูกสร้างขึ้น และเซลล์จะถูกควบคุมอีกครั้งเพื่อให้แน่ใจว่าอยู่ในสถานะที่เหมาะสมสำหรับการเข้าสู่ระยะไมโทซิส (M) ซึ่งเป็นระยะการแบ่งตัว มีกลไกตรวจสอบหลายจุดที่เกี่ยวข้องกับการเปลี่ยนผ่านจาก G2 ไปสู่ M โดยมีปัจจัยร่วมที่สำคัญคือการทำงานของไซคลิน-Cdk
แม้ว่าจะมีรูปแบบที่แตกต่างกันในคอมเพล็กซ์ไซคลิน-Cdk ที่จำเป็นในสิ่งมีชีวิตต่างๆ แต่ความจำเป็นของกิจกรรมไคเนสยังคงได้รับการอนุรักษ์ไว้และโดยทั่วไปจะมุ่งเน้นไปที่การจับคู่เพียงคู่เดียว ในยีสต์ฟิชชันมีไซคลินไมโทติกสามรูปแบบที่แตกต่างกัน และในยีสต์ที่แตกหน่อมีหกรูปแบบ แต่ไซคลินหลักที่ใช้คือไซคลิน B [ 15 ]ไซคลิน B จะทำหน้าที่เป็นตัวอ้างอิงสำหรับการอภิปรายเกี่ยวกับการเปลี่ยนผ่านจุดตรวจสอบ G2/M
เช่นเดียวกับระยะ S ระยะ G2 จะประสบกับจุดตรวจสอบความเสียหายของ DNA เซลล์จะถูกตรวจสอบอีกครั้งเพื่อหาตำแหน่งที่มีความเสียหายของ DNA หรือการจำลองแบบไม่สมบูรณ์ และไคเนส ATR และ ATM จะถูกดึงไปยังตำแหน่งที่เสียหาย การกระตุ้น Chk1 และ Chk2 ก็เกิดขึ้นเช่นกัน รวมถึงการกระตุ้น p53 เพื่อเหนี่ยวนำให้เกิดการหยุดวงจรเซลล์และหยุดการดำเนินไปสู่ไมโทซิส ส่วนประกอบเพิ่มเติมของระยะ S คือ คอมเพล็กซ์ก่อนการจำลองแบบ จะต้องถูกทำให้ไม่ทำงานผ่านการฟอสโฟรีเลชันของไซคลิน B-Cdk1 [ 16 ]
เมื่อมีการประเมินจุดตรวจสอบก่อนหน้านี้ การสะสมโปรตีน G2 จะช่วยกระตุ้นการทำงานของไซคลิน B-Cdk1 ผ่านกลไกหลายอย่าง ไซคลิน A-Cdk2 จะกระตุ้น Cdc25 ซึ่งเป็นตัวกระตุ้นของไซคลิน B-Cdk1 จากนั้น Cdc25 จะปิดใช้งานตัวยับยั้งไซคลิน B-Cdk1 คือ Wee1 ส่งผลให้เกิดวงจรป้อนกลับเชิงบวก ทำให้การแสดงออกของไซคลิน B และการกระตุ้น Cdk1 เพิ่มขึ้นอย่างมีนัยสำคัญ เมื่อเซลล์ดำเนินไปในระยะ G2 และเข้าสู่ช่วงเปลี่ยนผ่าน G2/M ไคเนส Plk1 จะฟอสโฟรีเลต Wee1 ซึ่งจะกำหนดเป้าหมาย Wee1 สำหรับการย่อยสลายผ่านคอมเพล็กซ์ยูบิควิตินไลเกส SCF [ 17 ]หน้าที่เพิ่มเติมของ Plk1 คือการกระตุ้น Cdc25 ผ่านการฟอสโฟรีเลต ผลรวมของการย่อยสลาย Wee1 และการกระตุ้น Cdc25 คือการกำจัดฟอสโฟรีเลตที่ยับยั้งออกจาก cdc2 ซึ่งจะกระตุ้น cdc2 Plk1 จะถูกกระตุ้นในช่วงการเปลี่ยนผ่าน G2/M โดย Aurora A และ Bora ซึ่งสะสมในช่วง G2 และสร้างคอมเพล็กซ์การกระตุ้น คอมเพล็กซ์ Plk1-Cdc2-cdc25 จะเริ่มต้นวงจรป้อนกลับเชิงบวกซึ่งทำหน้าที่กระตุ้น Cdc2 เพิ่มเติม และร่วมกับการเพิ่มขึ้นของระดับไซคลิน B ในช่วง G2 คอมเพล็กซ์ cdc2-ไซคลิน B ที่เกิดขึ้นจะกระตุ้นเป้าหมายปลายทางซึ่งส่งเสริมการเข้าสู่ไมโทซิส[ 18 ]กิจกรรม Cdk1 ที่เกิดขึ้นยังกระตุ้นการแสดงออกของ Mem1-Fkh ซึ่งเป็นยีนการเปลี่ยนผ่าน G2/M [ 19 ]การเพิ่มขึ้นอย่างรวดเร็วของกิจกรรมไซคลิน B-Cdk1 เป็นสิ่งจำเป็น เนื่องจากการเริ่มต้นเฟส M เป็นเหตุการณ์แบบทั้งหมดหรือไม่มีเลยที่เกี่ยวข้องกับฮิสเทอรีซิส ฮิสเทอรีซิสของกิจกรรม Cdk1 ผ่านไซคลิน B ขับเคลื่อนการเข้าสู่เฟส M โดยการสร้างเกณฑ์ขั้นต่ำของความเข้มข้นของไซคลิน B สิ่งนี้มีอยู่ที่ระดับที่สูงกว่าขั้นต่ำที่จำเป็นสำหรับการดำเนินเฟส M ต่อไปหลังจากการเข้าสู่เฟส โดยทำหน้าที่ปกป้องเหตุการณ์แบบทั้งหมดหรือไม่มีเลย ความเข้มข้นของการเข้าสู่เฟสนี้จะเพิ่มขึ้นอีกในกรณีที่การจำลองดีเอ็นเอไม่สมบูรณ์ ซึ่งเป็นการเพิ่มกลไกการควบคุมอีกอย่างหนึ่งที่จุดเปลี่ยนผ่าน G2/M [ 20 ]การมีอยู่ของฮิสเทอรีซิสทำให้การเข้าสู่เฟส M ได้รับการควบคุมอย่างเข้มงวดตามฟังก์ชันของกิจกรรมไซคลิน B-Cdk1
กลไกที่ป้องกันการเข้าสู่ระยะไมโทซิสเพื่อตอบสนองต่อความเสียหายของ DNA นั้นคล้ายคลึงกับกลไกในจุดตรวจสอบ G1/S ความเสียหายของ DNA กระตุ้นการทำงานของเส้นทาง ATM/ATR ที่กล่าวถึงข้างต้น ซึ่ง ATM/ATR จะฟอสฟอริเลตและกระตุ้นเอนไซม์ไคเนส Chk1/Chk2 ที่ทำหน้าที่ตรวจสอบจุดควบคุม Chk1/2 จะฟอสฟอริเลต cdc25 ซึ่งนอกจากจะถูกยับยั้งแล้ว ยังถูกกักเก็บไว้ในไซโตพลาสซึมโดยโปรตีน 14-3-3 โปรตีน 14-3-3 ถูกควบคุมโดย p53 ซึ่งดังที่กล่าวไว้ก่อนหน้านี้ ถูกกระตุ้นโดย Chk1 และ ATM/ATR p53 ยังกระตุ้นการทำงานของ p21 และทั้ง p21 และ 14-3-3 จะยับยั้งคอมเพล็กซ์ cyclin B-cdc2 ผ่านการฟอสฟอริเลตและการกักเก็บ cdc2 ไว้ในไซโตพลาสซึม นอกจากนี้ การไม่ทำงานของ cdc25 ส่งผลให้ไม่สามารถดีฟอสฟอริเลตและกระตุ้น cdc2 ได้[ 21 ] [ 22 ]สุดท้าย กลไกการตอบสนองต่อความเสียหายอีกประการหนึ่งคือการควบคุมเชิงลบของ Plk1 โดย ATM/ATR ซึ่งส่งผลให้ Wee1 และ Myt1 มีเสถียรภาพ จากนั้นจึงสามารถฟอสโฟรีเลตและยับยั้ง cdc2 ทำให้เซลล์หยุดนิ่งอยู่ในระยะ G2 จนกว่าความเสียหายจะได้รับการแก้ไข[ 23 ]
การเปลี่ยนผ่านจากระยะ G2 ไปสู่ระยะ M ในเซลล์ไข่ของกบ Xenopus
เมื่อสิ้นสุดระยะ G2 เซลล์จะเปลี่ยนเข้าสู่ระยะไมโทซิส ซึ่งนิวเคลียสจะแบ่งตัว การเปลี่ยนจาก G2 ไปสู่ M เกิดขึ้นอย่างรวดเร็วและไม่สามารถย้อนกลับได้ นี่เป็นข้อดีของเซลล์ เพราะการเข้าสู่ระยะไมโทซิสเป็นขั้นตอนสำคัญในวงจรชีวิตของเซลล์ หากเซลล์ไม่เข้าสู่ระยะนี้อย่างสมบูรณ์ เซลล์จะประสบปัญหามากมายจากการแบ่งตัวที่ไม่สมบูรณ์ ซึ่งอาจนำไปสู่ความตายในที่สุด
ในเซลล์ไข่ของกบ กระบวนการส่งสัญญาณจะเริ่มต้นขึ้นเมื่อโปรเจสเตอโรนจับกับตัวรับที่อยู่บนเยื่อหุ้มเซลล์ จากนั้น Mos จะถูกกระตุ้น Mos จะทำการฟอสฟอริเลต MEK1 ซึ่งจะทำการฟอสฟอริเลต MAPK ต่อไป MAPK มีบทบาทสองอย่างคือ การกระตุ้นคอมเพล็กซ์ Cyclin B-Cdk1 เพื่อเริ่มต้นการเข้าสู่ระยะไมโทซิส และการกระตุ้น Mos การกระตุ้น Mos นำไปสู่กลไกป้อนกลับเชิงบวก และจึงทำหน้าที่เป็น "สวิตช์เปิดปิด" เพื่อสร้างการเข้าสู่ระยะไมโทซิสแบบ "ทั้งหมดหรือไม่มีเลย"
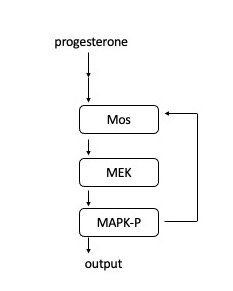
วงจรป้อนกลับนี้ถูกค้นพบครั้งแรกโดยแสดงให้เห็นว่าความเข้มข้นของ MAPK-P (MAPK ที่ถูกฟอสโฟรีเลต) เพิ่มขึ้นตามระดับของโปรเจสเตอโรนที่เพิ่มขึ้น[ 24 ]ในระดับเซลล์เดี่ยว เซลล์แต่ละเซลล์จะมี MAPK ที่ถูกฟอสโฟรีเลตทั้งหมดหรือไม่มี MAPK ที่ถูกฟอสโฟรีเลตเลย ซึ่งยืนยันว่ามันทำหน้าที่เป็นกลไกคล้ายสวิตช์ในแต่ละเซลล์ นอกจากนี้ยังแสดงให้เห็นว่าการปิดกั้นการสังเคราะห์โปรตีน Mos ทำให้การตอบสนองของ MAPK-P เป็นแบบค่อยเป็นค่อยไปมากขึ้น ซึ่งแสดงให้เห็นว่าการสังเคราะห์โปรตีน Mos มีความจำเป็นสำหรับลักษณะแบบทั้งหมดหรือไม่มีเลยของการกระตุ้น MAPK [ 25 ]
เสถียรภาพสองสถานะ
กระบวนการนี้สามารถทำความเข้าใจได้โดยใช้แนวคิดเรื่องความไม่เสถียร จากกราฟที่แสดงทางด้านขวา อัตราการสังเคราะห์ Mos จะเปลี่ยนแปลงไปเมื่อมีการเพิ่มโปรเจสเตอโรนมากขึ้น ในแต่ละเส้นโค้งจะมีจุดคงที่ที่เสถียรและจุดคงที่ที่ไม่เสถียร ณ จุดคงที่ที่ไม่เสถียร ระบบจะผลักดันไปสู่จุดคงที่ที่เสถียรจุดใดจุดหนึ่ง ดังนั้น ระบบจึงอยู่ในสถานะ "เปิด" หรือสถานะ "ปิด" เท่านั้น ไม่อยู่ระหว่างกลาง เมื่อระดับโปรเจสเตอโรนสูงพอ เส้นโค้ง Mos จะเลื่อนสูงขึ้นและในที่สุดจะตัดกับเส้นการสลายตัวที่จุดเดียวเท่านั้น ดังนั้นจึงมีสถานะ "เปิด" ที่เสถียรเพียงสถานะเดียว ซึ่งบ่งชี้ถึงการเข้าสู่ไมโทซิส
ความไม่สามารถย้อนกลับได้ที่เราเห็นในจุดเปลี่ยนผ่านของการแบ่งเซลล์แบบไมโทซิส เกิดจากการมีระดับโปรเจสเตอโรนในเซลล์สูงเพียงพอ เมื่อระดับโปรเจสเตอโรนสูงเพียงพอ ระบบจะเป็นแบบเสถียรเดียว (monostable) อันเป็นผลมาจากวงจรป้อนกลับเชิงบวกระหว่าง Mapk และ Mos จุดที่ระบบเปลี่ยนจากเสถียรสองแบบ (bistable) ไปเป็นแบบเสถียรเดียว (monostable) เรียกว่า จุดแยกแบบอานม้า (saddle node bifurcation)
ดังนั้น เราจึงสามารถเข้าใจปฏิกิริยาแบบ "ทั้งหมดหรือไม่มีเลย" และไม่สามารถย้อนกลับได้ของการเปลี่ยนผ่านในระยะไมโทซิส ด้วยแบบจำลองทางคณิตศาสตร์ของตัวควบคุมระดับโมเลกุลในฐานะระบบสองสถานะที่ขึ้นอยู่กับการมีอยู่ของปฏิกิริยาป้อนกลับเชิงบวก สถานะ "ปิด" จะถูกทำลายโดยระดับโปรเจสเตอโรนที่สูงพอ และเมื่อเซลล์ถูกผลักดันผ่านสถานะปิดไปแล้ว มันก็จะติดอยู่ในสถานะเปิด
ปรากฏการณ์ฮิสเทอรีซิสและแบบจำลองโนวัค-ไทสัน
จากแบบจำลองสองสถานะนี้ เราสามารถเข้าใจการเปลี่ยนแปลงของไมโทซิสโดยอาศัยฮิสเทอรีซิสเป็นตัวขับเคลื่อน ฮิสเทอรีซิสถูกนิยามว่าเป็นการพึ่งพาของสถานะของระบบต่อประวัติของมัน แบบจำลอง Novak–Tyson เป็นแบบจำลองทางคณิตศาสตร์ของความก้าวหน้าของวงจรเซลล์ที่ทำนายว่าการเปลี่ยนแปลงที่ไม่สามารถย้อนกลับได้ในการเข้าและออกจากไมโทซิสถูกขับเคลื่อนด้วยฮิสเทอรีซิส แบบจำลองนี้มีการคาดการณ์พื้นฐานสามประการที่ควรเป็นจริงในสารสกัดจากโอโอไซต์ที่มีวงจรซึ่งความก้าวหน้าของวงจรเซลล์ขึ้นอยู่กับฮิสเทอรีซิส: [ 26 ]
- ความเข้มข้นของไซคลิน บี ที่จำเป็นต่อการเข้าสู่ระยะไมโทซิสจะสูงกว่าความเข้มข้นที่จำเป็นต่อการคงสภาพของสารสกัดจากเซลล์ที่กำลังแบ่งตัวในระยะไมโทซิส
- ดีเอ็นเอที่ยังไม่ได้รับการจำลองแบบจะเพิ่มระดับของไซคลินซึ่งจำเป็นต่อการกระตุ้น Cdc2 และนำไปสู่การเข้าสู่ระยะไมโทซิส
- อัตราการกระตุ้นของ Cdc2 จะลดลงเมื่อความเข้มข้นของไซคลิน B สูงกว่าระดับเกณฑ์การกระตุ้นเล็กน้อย
Sha และคณะได้ทำการทดลองใน สารสกัดจากไข่ Xenopus laevisในปี 2546 เพื่อแสดงให้เห็นถึงลักษณะฮิสเทอรีซิสนี้[ 27 ]โดยใช้สารสกัดแบบวนรอบ พวกเขาพบว่าเกณฑ์การกระตุ้นสำหรับ Δcyclin B อยู่ระหว่าง 32 ถึง 42 nM ในขณะที่เกณฑ์การยับยั้งอยู่ระหว่าง 16 ถึง 24 nM Δcyclin B ดังนั้น การทดลองเหล่านี้จึงยืนยันถึงภาวะเสถียรสองสถานะของระบบนี้และความสำคัญของฮิสเทอรีซิสในการเปลี่ยนผ่านวงจรเซลล์นี้ ที่ความเข้มข้นของ cyclin B ระดับกลาง เซลล์อาจอยู่ในสถานะอินเตอร์เฟสหรือไมโทซิสก็ได้
การตอบสนองต่อความเครียดในการจำลองแบบ
เนื่องจากการเข้าสู่ระยะไมโทซิสเป็นการตัดสินใจครั้งใหญ่และมีค่าใช้จ่ายสูงสำหรับเซลล์ จึงเป็นเรื่องสมเหตุสมผลที่จะต้องมีระบบป้องกันการเข้าสู่ระยะนี้ก่อนกำหนด มีการแสดงให้เห็นแล้วว่าความผิดพลาดในขั้นตอนก่อนหน้า เช่น การมีส่วนของ DNA ที่ยังไม่ได้จำลอง จะขัดขวางความก้าวหน้าในวงจรเซลล์[ 28 ]แบบจำลองของ Novak–Tyson ทำนายว่าสิ่งนี้เกิดขึ้นโดยการเพิ่มระดับของไซคลิน B ซึ่งจำเป็นสำหรับการเข้าสู่ระยะไมโทซิส[ 26 ]
Sha และคณะได้ตรวจสอบว่าสิ่งนี้เป็นจริงใน สารสกัดจากไข่ Xenopusหรือไม่ พวกเขาใช้อะฟิดิโคลิน (APH) เพื่อยับยั้ง DNA polymerase และป้องกันการจำลอง DNA เมื่อได้รับการรักษาด้วย Cyclin B ในระยะอินเตอร์เฟส ค่าเกณฑ์การกระตุ้นจะเพิ่มขึ้นเป็นระหว่าง 80 ถึง 100 nM ตามที่แบบจำลอง Novak–Tyson ทำนายไว้[ 27 ]ดังนั้น การทดลองเหล่านี้จึงยืนยันว่าความเครียดของ DNA ที่ไม่ได้จำลองในเซลล์ส่งผลต่อวงจรฮิสเทอรีซิสและส่งผลให้ค่าเกณฑ์ของ Cyclin B สูงขึ้นมากในการเข้าสู่ไมโทซิส
จุดตรวจสอบระยะเมตาเฟส

จุด ตรวจสอบ แกนไมโทติกเกิดขึ้น ณ จุดในระยะเมตาเฟสที่โครโมโซมทั้งหมดควร/ได้เรียงตัวกันที่แผ่นไมโทติกและอยู่ภายใต้แรงตึงแบบไบโพลาร์ แรงตึงที่เกิดจากการยึดติดแบบไบโพลาร์นี้คือสิ่งที่ถูกตรวจจับ ซึ่งเป็นการเริ่มต้นการเข้าสู่ระยะแอนาเฟส ในการทำเช่นนี้ กลไกการตรวจจับจะทำให้แน่ใจว่าคอมเพล็กซ์ส่งเสริมแอนาเฟส (APC/C) ไม่ถูกยับยั้งอีกต่อไป ซึ่งตอนนี้สามารถย่อยสลายไซคลิน Bซึ่งมี D-box (กล่องทำลาย) และสลายเซคิวรินได้ [ 29 ] เซคิวรินเป็นโปรตีนที่มีหน้าที่ยับยั้งเซพาเรสซึ่งจะตัด โคฮี ซิน ซึ่งเป็น โปรตีนคอมโพสิตที่รับผิดชอบต่อการยึดเกาะของโครมาทิดคู่[ 30 ]เมื่อโปรตีนยับยั้งนี้ถูกย่อยสลายผ่านยูบิควิตินเนชันและการย่อยสลายโปรตีนในภายหลัง เซพาเรสก็จะทำให้โครมาทิดคู่แยกออกจากกัน[ 31 ]หลังจากที่เซลล์แบ่งตัวเป็นเซลล์ลูกสองเซลล์แล้ว เซลล์จะเข้าสู่ระยะG 1
มะเร็ง
กระบวนการ ซ่อมแซม DNAและจุดตรวจสอบวงจรเซลล์มีความเชื่อมโยงอย่างใกล้ชิดกับมะเร็งเนื่องจากหน้าที่ในการควบคุมความเสถียรของจีโนมและความก้าวหน้าของเซลล์ตามลำดับ กลไกโมเลกุลที่แม่นยำซึ่งเชื่อมโยงความผิดปกติในเส้นทางเหล่านี้กับการเกิดมะเร็งบางชนิดยังไม่เป็นที่เข้าใจดีในกรณีส่วนใหญ่[ 32 ] การสูญเสีย ATM แสดงให้เห็นว่าเกิดขึ้นก่อนการพัฒนาของมะเร็งต่อมน้ำเหลือง ซึ่งสันนิษฐานว่าเกิดจากการรวมตัวกันของโฮโมโลจัสมากเกินไป ทำให้เกิดความไม่เสถียรของจีโนมสูง[ 33 ]การหยุดชะงักของ Chk1 ในหนูทำให้เกิดการควบคุมจุดตรวจสอบวงจรเซลล์ผิดปกติอย่างมีนัยสำคัญ การสะสมของความเสียหายของ DNA และอุบัติการณ์ของการเกิดเนื้องอกเพิ่มขึ้น[ 34 ]การถ่ายทอดทางพันธุกรรมของBRCA1หรือBRCA2 ที่กลายพันธุ์เพียงตัวเดียว ทำให้เพศหญิงมีแนวโน้มที่จะเป็นมะเร็งเต้านมและมะเร็งรังไข่[ 35 ]เป็นที่ทราบกันว่า BRCA1 จำเป็นสำหรับการเปลี่ยนผ่าน S และ G2/M และเกี่ยวข้องกับการตอบสนองของเซลล์ต่อความเสียหายของ DNA เชื่อกันว่า BRCA2 มีส่วนเกี่ยวข้องกับการรวมตัวแบบโฮโมโลจัสและการควบคุมจุดตรวจสอบระยะ S และการกลายพันธุ์หรือการขาด BRCA2 มีความเชื่อมโยงอย่างมากกับการเกิดเนื้องอก[ 36 ]
ดูเพิ่มเติม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ จุดตรวจสอบวงจรเซลล์
จุดตรวจสอบวงจรเซลล์เป็นกลไกควบคุมในวงจรเซลล์ของยูคาริโอต ซึ่งช่วยให้มั่นใจได้ว่าวงจรเซลล์ดำเนินไปอย่างถูกต้อง...
พื้นหลัง
สิ่งมีชีวิตทั้งหมดเป็นผลผลิตจากการเจริญเติบโตและการแบ่งเซลล์ซ้ำๆ [ 5 ] ในระหว่างกระบวนการนี้ ซึ่งเรียกว่า วัฏจักรเซลล์ เซลล์จะจำลองเนื้อหาภายในแล้วแบ่งตัวออกเป็นสองส่วน จุดประสงค์ของวัฏจักรเซลล์คือการจำลอง DNA ของสิ่งมีชีวิตแต่ละชนิดอย่างแม่นยำ...
จุดตรวจ G1 (ข้อจำกัด)
จุดตรวจสอบ G1 หรือที่รู้จักกันในชื่อจุดจำกัดในเซลล์สัตว์เลี้ยงลูกด้วยนมและจุดเริ่มต้นในยีสต์ คือจุดที่เซลล์ตัดสินใจที่จะเข้าสู่รอบการแบ่งเซลล์ เมื่อเซลล์ดำเนินไปในระยะ G1 ขึ้นอยู่กับสภาวะภายในและภายนอก เซลล์อาจชะลอระยะ G1 เข้าสู่สภาวะสงบที่เรียกว่า G0...
จุดตรวจ G2
หลังจากกระบวนการจำลองดีเอ็นเอในระยะ S เซลล์จะเข้าสู่ระยะการเจริญเติบโตที่เรียกว่า G2 ในช่วงเวลานี้ โปรตีนไมโทซิสที่จำเป็นจะถูกสร้างขึ้น และเซลล์จะถูกควบคุมอีกครั้งเพื่อให้แน่ใจว่าอยู่ในสถานะที่เหมาะสมสำหรับการเข้าสู่ระยะไมโทซิส (M) ซึ่งเป็นระยะการแบ่งตัว...