อ่าน 12 นาที
ดีเมทิเลส
เอนไซม์ดีเมทิเลสเป็นเอนไซม์ที่กำจัดหมู่เมทิล (CH₃ ) ออกจาก กรดนิวคลีอิกโปรตีน (โดยเฉพาะฮิสโตน ) และโมเลกุลอื่นๆ
ดีเมทิเลส
เอนไซม์ดีเมทิเลสเป็นเอนไซม์ที่กำจัดหมู่เมทิล (CH₃ ) ออกจาก กรดนิวคลีอิกโปรตีน (โดยเฉพาะฮิสโตน ) และโมเลกุลอื่นๆ ดีเมทิเลสเป็นโปรตีนสำคัญทางด้านเอพิเจเนติกส์เนื่องจากมีหน้าที่ในการควบคุมการถอดรหัสของจีโนมโดยการควบคุมการเติมหมู่เมทิลในดีเอ็นเอและฮิสโตน และโดยนัยเดียวกันก็คือ สถานะของ โครมาตินในตำแหน่ง ยีน เฉพาะ
การกำจัดหมู่เมทิลออกจากไลซีนของฮิสโตน

ในตอนแรก การเมทิลเลชันของฮิสโตนถือเป็นกระบวนการที่ไม่สามารถย้อนกลับได้อย่างมีประสิทธิภาพ เนื่องจากครึ่งชีวิตของการเมทิลเลชันของฮิสโตนมีค่าประมาณเท่ากับครึ่งชีวิตของฮิสโตน[ 1 ] ฮิสโตนไลซีนดีเมทิเลส LSD1 (ต่อมาจัดอยู่ในกลุ่ม KDM1A) ได้รับการระบุครั้งแรกในปี 2547 ว่าเป็นโฮโมล็อกของนิวเคลียร์อะมีนออกซิเดส[ 2 ]ฮิสโตนไลซีนดีเมทิเลสมีสองกลุ่มหลัก โดยกำหนดจากกลไกการทำงาน ได้แก่อะมีนออกซิ เดส ที่ขึ้นอยู่ กับ ฟลาวินอะดีนีนไดนิวคลีโอไทด์ (FAD)และไฮดรอกซิเลสที่ขึ้นอยู่กับอัลฟา-คีโตกลูตาเรต
เอนไซม์ฮิสโตนไลซีนดีเมทิเลสมี โดเมนหลากหลายที่ทำหน้าที่ในการจดจำฮิสโตน การจับกับดีเอ็นเอ การจับกับสารตั้งต้น กรดอะมิโน ที่มีหมู่เมทิล และกิจกรรมเร่งปฏิกิริยา ซึ่งได้แก่:
- โดเมนอะมีนออกซิเดสที่ขึ้นอยู่กับ FAD ซึ่งประกอบด้วยไซต์เร่งปฏิกิริยาที่ใช้งานอยู่ของ KDM1
- โดเมน Jumonji-C ที่มีไซต์เร่งปฏิกิริยาที่ใช้งานอยู่ของ KDM2 ถึง KDM8 [ 3 ] [ 4 ]
- โดเมน Jumonji-N มีหน้าที่รับผิดชอบต่อความเสถียรของโครงสร้างโดเมน Jumonji-C
- โดเมน SWIRM (SWI3P, RSC8P และ Moira) ถูกเสนอให้เป็นจุดยึดสำหรับสารตั้งต้นฮิสโตนและรับผิดชอบต่อความเสถียรของโครมาติน
- โดเมน ซิงค์ฟิงเกอร์ PHD, CXXC และ C5HC2 มีหน้าที่ในการจดจำและจับกับฮิสโตน
เอนไซม์ฮิสโตนไลซีนดีเมทิเลสถูกจัดประเภทตามโดเมนและความจำเพาะของสารตั้งต้นที่ไม่เหมือนใคร สารตั้งต้นไลซีนถูกระบุตามตำแหน่งในลำดับกรดอะมิโนของฮิสโตนที่เกี่ยวข้องและสถานะการเมทิลเลชัน (ตัวอย่างเช่น H3K9me3 หมายถึงฮิสโตน 3 ไลซีน 9 ที่ถูกเมทิลเลชันสามตำแหน่ง)


- เคดีเอ็ม1
- โฮโมล็อกของ KDM1 ได้แก่KDM1AและKDM1B KDM1A ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K4me1/2 และ H3K9me1/2 ในขณะที่ KDM1B ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K4me1/2 กิจกรรมของ KDM1 มีความสำคัญต่อการ เกิดตัวอ่อนและ การแบ่งตัวของเนื้อเยื่อเฉพาะส่วนรวมถึงการเจริญเติบโตของไข่[ 1 ]การลบยีน KDM1A อาจส่งผลต่อการเจริญเติบโตและการแบ่งตัวของเซลล์ต้นกำเนิดตัวอ่อนและเป็นอันตรายถึงชีวิตในหนูทดลองที่ยีนถูกกำจัด[ 5 ] [ 6 ] พบว่าการแสดงออกของยีน KDM1A เพิ่มขึ้นในมะเร็งบางชนิด[ 7 ] [ 8 ]ดังนั้นการยับยั้ง KDM1A จึงถูกพิจารณาว่าเป็นวิธีการรักษาทางเอพิเจเนติกส์ที่เป็นไปได้สำหรับมะเร็ง[ 9 ] [ 10 ] [ 11 ]อย่างไรก็ตาม KDM1B ส่วนใหญ่เกี่ยวข้องกับการพัฒนาของไข่การลบยีนนี้ส่งผลให้เกิด การตายจาก ผลกระทบของแม่ในหนู[ 12 ] ออร์โธล็อกของ KDM1 ในD. melanogasterและC. elegansดูเหมือนจะทำงานคล้ายกับ KDM1B มากกว่า KDM1A [ 13 ] [ 14 ]
- เคดีเอ็ม2
- โฮโมล็อกของ KDM2 ได้แก่KDM2AและKDM2B KDM2A และ KDM2B ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K4me3 และ H3K36me2/3 KDM2A มีบทบาทในการส่งเสริมหรือยับยั้งการทำงานของเนื้องอก และ KDM2B มีบทบาทในการเกิดมะเร็ง[ 1 ] KDM2A และ KDM2B มีโดเมนซิงค์ฟิงเกอร์ CXXC ที่รับผิดชอบในการจับกับเกาะ CpG ที่ไม่ถูกเมทิลเลชั่น และเชื่อกันว่าพวกมันอาจจับกับองค์ประกอบควบคุมยีนจำนวนมากโดยปราศจากปัจจัยการถอดรหัสที่จำเพาะต่อลำดับ[ 15 ]มีการสังเกตพบ KDM2B ที่แสดงออกมากเกินไปในมะเร็งต่อมน้ำเหลืองและมะเร็งต่อมน้ำเหลือง ในมนุษย์ และมีการสังเกตพบ KDM2B ที่แสดงออกน้อยเกินไปในมะเร็งต่อมลูกหมากและมะเร็งสมองกลิโอบลาสโตมาในมนุษย์ นอกจากนี้ยังพบว่า KDM2B สามารถป้องกันการแก่ชราในบางเซลล์ผ่านการแสดงออกนอกตำแหน่ง[ 16 ]
- เคดีเอ็ม3
- โฮโมล็อกของ KDM3 ได้แก่KDM3A , KDM3BและKDM3C KDM3A, KDM3B และ KDM3C ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K9me1/2 KDM3A มีบทบาทในการสร้างสเปิร์มและหน้าที่การเผาผลาญ อย่างไรก็ตาม กิจกรรมของ KDM3B และ KDM3C ยังไม่เป็นที่ทราบแน่ชัด[ 1 ] : การศึกษาการลดระดับ KDM3A ในหนูทดลองส่งผลให้เกิดภาวะมีบุตรยากในเพศชายและโรคอ้วนในวัยผู้ใหญ่ การศึกษาเพิ่มเติมชี้ให้เห็นว่า KDM3A อาจมีบทบาทในการควบคุมยีนที่ขึ้นอยู่กับตัวรับแอนโดรเจน รวมถึงยีนที่เกี่ยวข้องกับ ความสามารถในการสร้าง เซลล์ต้นกำเนิดซึ่งบ่งชี้ถึงบทบาทที่เป็นไปได้ของ KDM3A ในการเกิดเนื้องอก[ 17 ]
- เคดีเอ็ม4
- โฮโมล็อกของ KDM4 ได้แก่KDM4A , KDM4B , KDM4C , KDM4D , KDM4E และ KDM4F KDM4A, KDM4B และ KDM4C ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K9me2/3, H3K9me3 และ H3K36me2/3 ในขณะที่ KDM4D, KDM4E และ KDM4F ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K9me2/3 KDM4A, KDM4B, KDM4C และ KDM4D มีบทบาทในการเกิดเนื้องอกอย่างไรก็ตาม กิจกรรมของ KDM4E และ KDM4F ยังไม่เป็นที่ทราบแน่ชัด[ 1 ]การเพิ่มขึ้นของ KDM4B พบได้ในเมดุลโลบลาสโตมา และการขยายตัวของ KDM4C ได้รับการบันทึกไว้ในมะเร็งเซลล์สความัสของหลอดอาหาร เมดุลโลบลาสโตมา และมะเร็งเต้านม[ 18 ] [ 19 ] [ 20 ] [ 21 ]ข้อมูลการแสดงออกของยีนอื่นๆ ยังชี้ให้เห็นว่า KDM4A, KDM4B และ KDM4C มีการแสดงออกมากเกินไปในมะเร็งต่อมลูกหมาก[ 22 ]
- เคดีเอ็ม5
- โฮโมล็อกของ KDM5 ประกอบด้วยKDM5A , KDM5B , KDM5CและKDM5D KDM5A, KDM5B, KDM5C และ KDM5D ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K4me2/3 [ 1 ]ตระกูล KDM5 ดูเหมือนจะควบคุมหน้าที่สำคัญในการพัฒนา รวมถึงการแยกแยะเซลล์ การทำงาน ของไมโทคอนเดรียและการดำเนินไปของวงจรเซลล์[ 23 ] [ 24 ] [ 25 ] [ 26 ] [ 27 ] [ 28 ] นอกจากนี้KDM5B และ KDM5C ยังแสดงให้เห็นถึงการโต้ตอบกับโปรตีน PcG ซึ่งเกี่ยวข้องกับการยับยั้งการถอดรหัส การกลายพันธุ์ของ KDM5C บนโครโมโซม X ยังพบในผู้ป่วยที่มีความบกพร่องทางสติปัญญาที่เชื่อมโยงกับโครโมโซม X [ 29 ]การลดลงของโฮโมล็อก KDM5C ในD. rerioแสดงให้เห็นถึงความบกพร่องของรูปแบบสมองและการตายของเซลล์ประสาท[ 30 ]
- เคดีเอ็ม6
- ตระกูล KDM6 ประกอบด้วยKDM6A , KDM6BและKDM6C KDM6A และ KDM6B ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K27me2/3 และ KDM4C ทำหน้าที่กำจัดหมู่เมทิลออกจาก H3K27me3 KDM6A และ KDM6B มีคุณสมบัติในการยับยั้งเนื้องอก การลดระดับ KDM6A ในไฟโบรบลาสต์ส่งผลให้จำนวนประชากรไฟโบรบลาสต์เพิ่มขึ้นทันที การแสดงออกของ KDM6B ในไฟโบรบลาสต์จะกระตุ้นยีนก่อมะเร็งของเส้นทาง Ras/Raf/MEK/ERK [ 31 ]การกลายพันธุ์แบบจุดของ KDM6A ได้รับการระบุว่าเป็นสาเหตุหนึ่งของกลุ่มอาการคาบูกิซึ่งเป็นความผิดปกติแต่กำเนิดที่ส่งผลให้เกิดความบกพร่องทางสติปัญญา[ 32 ] [ 33 ]การลบ KDM6A ในD. rerioส่งผลให้การแสดงออกของยีน HOX ลดลง ซึ่งมีบทบาทในการควบคุมรูปแบบของร่างกายในระหว่างการพัฒนา[ 34 ]ในการศึกษาในสัตว์เลี้ยงลูกด้วยนม พบว่า KDM6A ยังควบคุมยีน HOX อีกด้วย[ 35 ] [ 36 ]การกลายพันธุ์ของ KDM5B ขัดขวางการพัฒนาของอวัยวะสืบพันธุ์ในC.elegans [ 35 ]การศึกษาอื่นๆ แสดงให้เห็นว่าการแสดงออกของ KDM6B เพิ่มขึ้นในแมโครฟาจที่ ถูกกระตุ้น และแสดงออกอย่างมีพลวัตในระหว่างการแยกตัวของเซลล์ต้นกำเนิด[ 37 ] [ 38 ]
การกำจัดหมู่เมทิลของเอสเทอร์

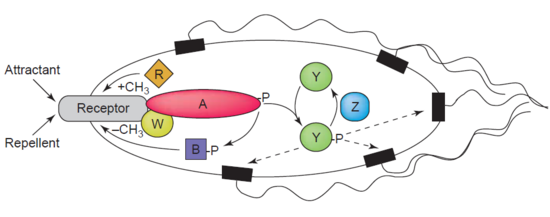
อีกตัวอย่างหนึ่งของดีเมทิเลสคือโปรตีนกลูตาเมตเมทิลเอสเทอเรสหรือที่รู้จักกันในชื่อโปรตีน CheB (EC 3.1.1.61) ซึ่งทำหน้าที่ดีเมทิเลส MCPs ( โปรตีนรับเมทิล ใน การเคลื่อนที่ของเซลล์ ) ผ่านการไฮโดรไลซิสของพันธะเอสเทอร์คาร์บอกซิลิก การเชื่อมโยงของตัวรับการเคลื่อนที่ของเซลล์กับตัวกระตุ้นนำไปสู่การฟอสโฟรีเลชันของ CheB การฟอสโฟรีเลชันของโปรตีน CheB ช่วยเพิ่มกิจกรรมการดีเมทิเลสของ MCP ส่งผลให้เซลล์ปรับตัวเข้ากับสิ่งเร้าจากสิ่งแวดล้อม[ 39 ] MCPs ตอบสนองต่อสารดึงดูดและสารขับไล่ภายนอกเซลล์ในแบคทีเรียเช่นE. coliใน การควบคุม การเคลื่อนที่ของเซลล์ CheB ถูกเรียกว่า เมทิลเอสเทอเรสโดยเฉพาะเนื่องจากมันกำจัดหมู่เมทิลออกจาก สารตกค้าง เมทิลกลูตาเมตที่อยู่บน MCPs ผ่านการไฮโดรไลซิส ทำให้เกิดกลูตาเมตพร้อมกับการปล่อยเมทานอล[ 40 ]
CheB เป็นที่สนใจเป็นพิเศษของนักวิจัย เนื่องจากอาจเป็นเป้าหมายในการรักษาเพื่อลดการแพร่กระจายของการติดเชื้อแบคทีเรีย[ 41 ]
ดูเพิ่มเติม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ดีเมทิเลส
เอนไซม์ดีเมทิเลสเป็นเอนไซม์ที่กำจัดหมู่เมทิล (CH₃ ) ออกจาก กรดนิวคลีอิกโปรตีน (โดยเฉพาะฮิสโตน ) และโมเลกุลอื่นๆ
การกำจัดหมู่เมทิลออกจากไลซีนของฮิสโตน
ในตอนแรก การเมทิลเลชันของฮิสโตน ถือเป็นกระบวนการที่ไม่สามารถย้อนกลับได้อย่างมีประสิทธิภาพ เนื่องจากครึ่งชีวิตของการเมทิลเลชันของฮิสโตนมีค่าประมาณเท่ากับครึ่งชีวิตของฮิสโตน [ 1 ] ฮิสโตนไลซีนดีเมทิเลส LSD1 (ต่อมาจัดอยู่ในกลุ่ม KDM1A) ได้รับการระบุครั้งแรกในปี...
การกำจัดหมู่เมทิลของเอสเทอร์
อีกตัวอย่างหนึ่งของดีเมทิเลสคือ โปรตีนกลูตาเมตเมทิลเอสเทอเรส หรือที่รู้จักกันในชื่อโปรตีน CheB (EC 3.1.1.
ดูเพิ่มเติม
เคโมแท็กซิส เอสเตอเรส ทรานสเฟอเรส เมทิลเลส ดึงข้อมูลมาจาก " https://en.wikipedia.org/w/index.php?title=Demethylase&oldid=1323550819 "