อ่าน 25 นาที
ดรอเมโอซอริเด
Dromaeosauridae ( / ˌ d r ɒ m i . ə ˈ s ɔːr ɪ d iː / ) เป็นวงศ์ของไดโนเสาร์เทอโรพอดโคเอลูโรซอเรียน ที่มีขน พวกมันโดยทั่วไปเป็น สัตว์กินเนื้อที่มีขนขนาดเล็กถึงขนาดกลางที่เฟื่องฟูใน.
ดรอเมโอซอริเด
| ดรอเมโอซอริเด ช่วงเวลา: | |
|---|---|
 | |
| ภาพโครงกระดูกฟอสซิลของไดโนเสาร์กลุ่มดรอมิโอ ซอ ริเดีย เรียงตามเข็มนาฬิกาจากซ้ายบน: ไดโนนิคัส แอนติร์โรปัส (ยูโดรมิโอซอร์ที่มีโครงสร้างใหญ่โต) , บิวเทรแรปเตอร์ กอนซาเลโซรัม (อูเนนลาไจนีนที่มีจมูกยาว) , เวโลซิแรปเตอร์ มงโกเลียน ซิส (เวโลซิแรปเตอร์รีนขนาดเล็ก), ไมโครแรปเตอร์ กุย (ไมโครแรปเตอร์เรียนมีปีก), ฮา ลส์คาแรปเตอร์ เอสคูยีอี (ฮาลส์คาแรปเตอร์รีนกึ่งน้ำ), เจิ้นหยวนหลง ซูนิ (ดรอมิโอซอริเดียมีปีกยาว) | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลื้อยคลาน |
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ |
| กลุ่มสายพันธุ์ : | ซอริสเชีย |
| กลุ่มสายพันธุ์ : | เทโรโปดา |
| กลุ่มสายพันธุ์ : | ปาราเวส |
| ตระกูล: | † Dromaeosauridae Matthew & Brown , 1922 |
| ชนิดต้นแบบ | |
| † โดรเมโอซอรัส อัลเบอร์เทนซิส แมทธิว แอนด์ บราวน์, 1922 | |
| กลุ่มย่อย[ 5 ] | |
| |
| คำพ้องความหมาย | |
| |
Dromaeosauridae ( / ˌ d r ɒ m i . ə ˈ s ɔːr ɪ d iː / ) เป็นวงศ์ของไดโนเสาร์เทอโรพอดโคเอลูโรซอเรียน ที่มีขน พวกมันโดยทั่วไปเป็น สัตว์กินเนื้อที่มีขนขนาดเล็กถึงขนาดกลางที่เฟื่องฟูใน ยุค ครีเทเชียสชื่อ Dromaeosauridae หมายถึง 'กิ้งก่าวิ่ง' มาจากภาษากรีกδρομαῖος ( dromaîos ) ซึ่งหมายถึง 'วิ่งด้วยความเร็วเต็มที่' 'รวดเร็ว' และσαῦρος ( saûros ) ซึ่งหมายถึง 'กิ้งก่า' ในการใช้งานแบบไม่เป็นทางการ พวกมันมักถูกเรียกว่าแรปเตอร์[ 6 ] (ตามชื่อเวโลซิแรปเตอร์ ) ซึ่งเป็นคำที่ได้รับความนิยมจากภาพยนตร์เรื่องจูราสสิกพาร์ค สกุล หลายสกุลมีคำว่า "แรปเตอร์" อยู่ในชื่อโดยตรง และวัฒนธรรมสมัยนิยมได้เน้นย้ำถึงรูปลักษณ์ที่คล้ายนกและการคาดเดาพฤติกรรมที่คล้ายนกของพวกมัน
ฟอสซิลของไดโนเสาร์กลุ่ม ดรอเมโอซอริเดที่แน่ชัดพบได้ในทวีปอเมริกาเหนือยุโรปและเอเชียฟอสซิลของพาราเวียนบางส่วนที่พบในทวีปอื่นๆ ได้รับการพิจารณาว่าเป็นดรอเมโอซอริเดมาแต่เดิม แต่เมื่อเร็วๆ นี้ได้รับการตีความใหม่ว่าเป็นวงศ์Unenlagiidae ที่ไม่เหมือนใคร นอกเหนือจากวงศ์ Dromaeosauridae โดยผู้เขียนบางคนพิจารณาว่าเป็นสายพันธุ์ที่แยกต่างหากของAvialae [ 7 ] ฟอสซิลร่างกายที่เก่าแก่ที่สุดเป็นที่รู้จักจากยุคค รีเทเชียสตอนต้น (145–140 ล้านปีก่อน) และมีชีวิตรอดจนถึงสิ้นสุดยุคครีเทเชียส ( ยุค มาสทริชเชียน 66 ล้านปีก่อน) ดำรงอยู่จนถึงเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคครีเทเชียส-พาลีโอจีนการปรากฏตัวของดรอเมโอซอริเดตั้งแต่ยุคจูราสสิกตอนกลางได้รับการเสนอแนะจากการค้นพบฟอสซิลฟันที่แยกออกมา แม้ว่าจะไม่พบฟอสซิลร่างกายของดรอเมโอซอริเดจากช่วงเวลานี้ก็ตาม[ 8 ] [ 9 ]
คำอธิบาย
การวินิจฉัยทางเทคนิค
Dromaeosaurids มีลักษณะเฉพาะดังต่อไปนี้: กระดูกหน้าผาก รูปตัว T สั้น ที่สร้าง ขอบเขต ด้านหน้าของช่อง supratemporal fenestra ; ชั้นวางที่ยื่นออกมาทางด้านท้ายและด้านข้างของกระดูก squamosal ; กระบวนการด้านข้างของกระดูก quadrateที่สัมผัสกับกระดูกquadratojugal ; parapophyses ที่ยกขึ้นและมีก้านบนกระดูกสันหลังส่วนหลัง ; นิ้วเท้าที่ 2 ที่ดัดแปลง; chevronsและprezygapophysisของกระดูกสันหลัง ส่วนหาง ที่ยาวและทอดข้ามกระดูกสันหลังหลายข้อ; การมีแอ่ง subglenoid บนกระดูกcoracoid [ 10 ]
ขนาดและโครงสร้างโดยทั่วไป


ไดโนเสาร์กลุ่มดรอเมโอซอริเดเป็นไดโนเสาร์ขนาดเล็กถึงขนาดกลาง มีความยาวตั้งแต่ 1.5–2.07 เมตร (4.9–6.8 ฟุต) (ในกรณีของเวโลซิแรปเตอร์ ) ไปจนถึงเกือบหรือเกิน 6 เมตร (20 ฟุต) (ในยูทาห์แรปเตอร์ดาโกตาแรปเตอร์และอะคิลโลเบเตอร์ ) [ 11 ] [ 12 ]ดูเหมือนว่าขนาดใหญ่จะวิวัฒนาการขึ้นอย่างน้อยสองครั้งในกลุ่มดรอเมโอซอริเด ครั้งหนึ่งในกลุ่มดรอเมโอซอรีนยูทาห์แรปเตอร์และอะคิลโลเบเตอร์และอีกครั้งในกลุ่มอูเนนลาไจน์ ( ออสโทรแรปเตอร์ ซึ่งมีความยาว 5–6 เมตร (16–20 ฟุต)) สายพันธุ์ที่สามที่เป็นไปได้ของดรอเมโอซอริเดขนาดยักษ์ นั้นแสดงให้เห็นได้จากฟันที่แยกออกมาซึ่งพบในเกาะไอล์ออฟไวต์ประเทศอังกฤษฟันเหล่านี้เป็นของสัตว์ที่มีขนาดเท่ากับUtahraptor ซึ่งเป็น ไดโนเสาร์กลุ่ม Dromaeosaurine แต่ดูเหมือนว่าจะเป็นของไดโนเสาร์กลุ่ม Velociraptorine เมื่อพิจารณาจากรูปร่างของฟัน[ 13 ] [ 14 ]
โครงสร้างร่างกายของไดโนเสาร์กลุ่มดรอเมโอซอริเดที่โดดเด่นช่วยจุดประกายทฤษฎีที่ว่าไดโนเสาร์อาจว่องไว วิ่งเร็ว และมีความสัมพันธ์ใกล้ชิดกับนกภาพประกอบของโรเบิร์ต บักเกอร์ สำหรับ งานวิจัยของจอห์น ออสตรอม ในปี 1969 [ 15 ]ที่แสดงให้เห็นไดโนนิคัสดรอเมโอซอริเดวิ่งเร็ว เป็นหนึ่งในการสร้างภาพทางบรรพชีวินวิทยาที่มีอิทธิพลมากที่สุดในประวัติศาสตร์[ 16 ]โครงสร้างร่างกายของไดโนเสาร์กลุ่มดรอเมโอซอริเดประกอบด้วยกะโหลกศีรษะขนาดค่อนข้างใหญ่ ฟันหยัก จมูกแคบ (ยกเว้นดรอเมโอซอรีนที่ วิวัฒนาการแล้ว ) และดวงตาที่หันไปข้างหน้าซึ่งบ่งชี้ถึงการมองเห็นแบบสองตาในระดับหนึ่ง[ 17 ]

ไดโนเสาร์กลุ่ม Dromaeosaurids เช่นเดียวกับไดโนเสาร์กลุ่ม theropods ส่วนใหญ่ มีคอยาวปานกลางโค้งเป็นรูปตัว S และลำตัวค่อนข้างสั้นและลึก เช่นเดียวกับไดโนเสาร์กลุ่ม maniraptorans อื่นๆ พวกมันมีแขนยาวที่สามารถพับแนบกับลำตัวได้ในบางชนิด และมีมือค่อนข้างใหญ่ที่มีนิ้วยาวสามนิ้ว (นิ้วกลางยาวที่สุดและนิ้วชี้สั้นที่สุด) ปลายนิ้วมีกรงเล็บขนาดใหญ่[ 10 ]โครงสร้างสะโพกของไดโนเสาร์กลุ่ม Dromaeosaurids มีลักษณะเด่นคือมีส่วนยื่นคล้ายรองเท้าขนาดใหญ่ที่ยื่นออกมาใต้โคนหาง เท้าของไดโนเสาร์กลุ่ม Dromaeosaurids มีกรงเล็บขนาดใหญ่โค้งงอที่นิ้วเท้าที่สอง หางของพวกมันเรียว มีกระดูกสันหลัง ยาวและต่ำ ไม่มีกระบวนการตามขวางและหนามประสาทหลังจากกระดูกสันหลังส่วนหางชิ้นที่ 14 [ 10 ] มีการระบุพบกระบวนการกระดูกรูป ตะขอของซี่โครงในไดโนเสาร์กลุ่ม Dromaeosaurids หลายชนิด[ 18 ] [ 19 ] [ 20 ]
เท้า


เช่นเดียวกับเทโรพอดอื่นๆ ดรอเมโอซอริเดเป็นสัตว์สองขา กล่าวคือ พวกมันเดินด้วยขาหลัง อย่างไรก็ตาม ในขณะที่เทโรพอดส่วนใหญ่เดินโดยใช้สามนิ้วสัมผัสพื้น ร่องรอยเท้าฟอสซิลยืนยันว่า กลุ่ม พาราเวียนยุค แรกหลาย กลุ่ม รวมถึงดรอเมโอซอริเด ยกนิ้วที่สองขึ้นจากพื้นในตำแหน่งที่ยืดออกมากเกินไป โดยมีเพียงนิ้วที่สามและสี่เท่านั้นที่รับน้ำหนักของสัตว์ นี่เรียกว่าดิดักทิลีเชิงฟังก์ชัน[ 21 ]นิ้วที่สองที่ขยายใหญ่ขึ้นมีกรงเล็บขนาดใหญ่ โค้งงอ รูป ทรงเคียว (รูปเคียว หรือ ด รี พานอยด์ ) ที่ผิดปกติ (ยกขึ้นจากพื้นหรือ 'หดกลับ' เมื่อเดิน) ซึ่งเชื่อกันว่าใช้ในการจับเหยื่อและปีนต้นไม้ (ดู "หน้าที่ของกรงเล็บ" ด้านล่าง) กรงเล็บนี้มีลักษณะคล้ายใบมีดเป็นพิเศษใน ยูโดรเมโอซอรัสที่เป็นนักล่าขนาดใหญ่[ 22 ] Balaur bondoc ซึ่ง เป็นไดโนเสาร์ดรอเมโอซอริเดชนิดหนึ่งที่เป็นไปได้ก็มีนิ้วเท้าแรกที่ได้รับการดัดแปลงอย่างมากในลักษณะขนานกับนิ้วเท้าที่สอง นิ้วเท้าแรกและนิ้วเท้าที่สองบนเท้าแต่ละข้างของB. bondoc นั้นหดเข้าและมีกรงเล็บรูปเคียวขนาดใหญ่ กรงเล็บที่คล้ายกันนี้สามารถพบได้ใน Seriemaในปัจจุบัน[ 23 ]
หาง

ไดโนเสาร์กลุ่ม Dromaeosaurids มีหางยาว กระดูกสันหลังส่วนหางส่วนใหญ่มีส่วนยื่นคล้ายแท่งกระดูก (เรียกว่า prezygapophyses) รวมถึงเอ็นกระดูกในบางชนิด ในการศึกษาDeinonychus ของ Ostrom เขาเสนอว่าลักษณะเหล่านี้ทำให้หางแข็งขึ้นจนสามารถงอได้เฉพาะที่โคนเท่านั้น และหางทั้งหมดจะเคลื่อนที่เหมือนคานแข็งๆ อันเดียว[ 15 ]อย่างไรก็ตาม ตัวอย่างVelociraptor mongoliensis ที่ได้รับการอนุรักษ์ไว้อย่างดี (IGM 100/986) มีโครงกระดูกหางที่เชื่อมต่อกันซึ่งโค้งงอในแนวนอนเป็นรูปตัว S ยาว สิ่งนี้ชี้ให้เห็นว่าในขณะที่ยังมีชีวิตอยู่ หางสามารถงอไปมาได้ด้วยความยืดหยุ่นในระดับหนึ่ง[ 24 ]มีการเสนอว่าหางนี้ใช้เป็นตัวช่วยในการทรงตัวหรือถ่วงน้ำหนักขณะวิ่งหรืออยู่ในอากาศ[ 24 ]ในMicroraptorมีพัดขนรูปเพชรยาวที่ได้รับการอนุรักษ์ไว้ที่ปลายหาง สิ่งนี้อาจถูกนำมาใช้เป็นตัวกันโคลงทางอากาศพลศาสตร์และหางเสือระหว่างการร่อนหรือการบินด้วยกำลัง (ดู "การบินและการร่อน" ด้านล่าง) [ 25 ]
ขนนก


มีหลักฐานจำนวนมากที่แสดงให้เห็นว่าไดโนเสาร์กลุ่มดรอเมโอซอริเดมีขน ปกคลุม ทั่วตัว ฟอสซิลของไดโนเสาร์กลุ่มดรอเมโอซอริเดบางส่วนยังคงรักษาขนยาวแบบเพนนาเซียสไว้บนมือและแขน ( เรมิจีส ) และหาง ( เรคทริซีส ) รวมถึงขนสั้นคล้ายขนปุยที่ปกคลุมทั่วร่างกาย[ 26 ] [ 27 ]ฟอสซิลอื่นๆ ที่ไม่ได้รักษาร่องรอยของขนไว้ แต่ยังคงรักษาปุ่มที่เกี่ยวข้องบนกระดูกปลายแขนซึ่งเป็นตำแหน่งที่ขนปีกยาวจะติดอยู่ขณะมีชีวิต[ 28 ] โดยรวมแล้ว รูปแบบขนนี้ดูคล้ายกับ อาร์คีออปเทอริกซ์มาก[ 26 ]
ไดโนเสาร์ดรอมิโอซอริเดตัวแรกที่มีหลักฐานแน่ชัดว่ามีขนคือซิโนร์นิโทซอรัส ซึ่ง รายงานจากประเทศจีนโดย Xu และคณะในปี 1999 [ 27 ]ฟอสซิลดรอมิโอซอริเดอื่นๆ อีกมากมายถูกพบว่ามีขนปกคลุมทั่วร่างกาย บางตัวมีปีกที่มีขนพัฒนาเต็มที่ไมโครแรปเตอร์ยังแสดงหลักฐานของปีกคู่ที่สองบนขาหลังอีกด้วย[ 26 ]แม้ว่ารอยประทับของขนโดยตรงจะเป็นไปได้เฉพาะในตะกอนที่มีเม็ดละเอียดเท่านั้น แต่ฟอสซิลบางส่วนที่พบในหินที่หยาบกว่าแสดงหลักฐานของขนโดยการมีอยู่ของปุ่มขน ซึ่งเป็นจุดยึดสำหรับขนปีกที่นกบางชนิดมี ดรอมิโอ ซอริ เด ราโฮนาวิสและเวโลซิแรปเตอร์ต่างก็ถูกพบว่ามีปุ่มขน แสดงให้เห็นว่าไดโนเสาร์เหล่านี้มีขนแม้ว่าจะไม่พบรอยประทับก็ตาม จากข้อมูลนี้ เป็นไปได้มากว่าแม้แต่ไดโนเสาร์ดรอเมโอซอริเดขนาดใหญ่ที่อาศัยอยู่บนพื้นดินก็ยังมีขน เนื่องจากแม้แต่นกที่บินไม่ได้ในปัจจุบันก็ยังคงมีขนอยู่เกือบทั้งหมด และไดโนเสาร์ดรอเมโอซอริเดที่มีขนาดค่อนข้างใหญ่ เช่นเวโลซิแรปเตอร์ก็เป็นที่ทราบกันว่ายังคงมีขนแบบเพนนาเซียสอยู่[ 28 ] [ 29 ]แม้ว่านักวิทยาศาสตร์บางคนจะเสนอแนะว่าไดโนเสาร์ดรอเมโอซอริเดขนาดใหญ่สูญเสียเกราะป้องกันบางส่วนหรือทั้งหมดไป แต่การค้นพบขนใน ตัวอย่างของ เวโลซิแรปเตอร์ได้รับการอ้างถึงว่าเป็นหลักฐานว่าสมาชิกทุกตัวในวงศ์นี้ยังคงมีขนอยู่[ 28 ] [ 30 ]
เมื่อไม่นานมานี้ การค้นพบZhenyuanlongได้ยืนยันการมีอยู่ของขนปกคลุมเต็มตัวในไดโนเสาร์ดรอเมโอซอริเดขนาดค่อนข้างใหญ่ นอกจากนี้ สัตว์ชนิดนี้ยังแสดงให้เห็นขนปีกขนาดใหญ่ตามสัดส่วนที่ลู่ลม รวมถึงหางที่แผ่กว้าง ซึ่งทั้งสองอย่างนี้เป็นลักษณะที่ไม่คาดคิดที่อาจช่วยให้เข้าใจถึงผิวหนังของไดโนเสาร์ดรอเมโอซอริเดขนาดใหญ่ได้[ 31 ] Dakotaraptorเป็นไดโนเสาร์ดรอเมโอซอริเดสายพันธุ์ที่ใหญ่กว่ามาก โดยมีหลักฐานของขน แม้ว่าจะทางอ้อมในรูปแบบของปุ่มขน[ 32 ]ถึงแม้ว่านักวิจัยคนอื่นๆ จะถือว่าแท็กซอนนี้เป็นไคเมียรา เนื่องจากองค์ประกอบของไดโนเสาร์ที่มีลักษณะเฉพาะของดรอเมโอซอรัสยังสามารถจัดอยู่ในกลุ่มซีนาแนทิดและออร์นิโทมิโมซอเรียนได้ อีกด้วย [ 33 ] [ 34 ]
การจำแนกประเภท
ความสัมพันธ์กับนก
ไดโนเสาร์กลุ่ม Dromaeosaurids มีลักษณะหลายอย่างร่วมกับนก ยุคแรก (กลุ่มAvialaeหรือAves ) ความสัมพันธ์ที่แน่ชัดระหว่างพวกมันกับนกได้รับการศึกษามาอย่างมาก และสมมติฐานเกี่ยวกับความสัมพันธ์นั้นก็เปลี่ยนแปลงไปตามหลักฐานใหม่จำนวนมากที่ปรากฏขึ้น จนกระทั่งปี 2001 Mark Norellและเพื่อนร่วมงานได้วิเคราะห์ฟอสซิลของไดโนเสาร์กลุ่ม Coelurosaur จำนวนมาก และได้ข้อสรุปเบื้องต้นว่า Dromaeosaurids มีความสัมพันธ์ใกล้ชิดกับนกมากที่สุด โดยมีTroodontidsเป็นกลุ่มนอกที่ห่างไกลออกไป พวกเขายังเสนอแนะว่า Dromaeosauridae อาจเป็น กลุ่ม ที่ไม่เป็นเอกภาพเมื่อเทียบกับ Avialae [ 35 ]ในปี 2002 Hwang และเพื่อนร่วมงานได้ใช้ผลงานของ Norell และคณะ รวมถึงลักษณะใหม่และหลักฐานฟอสซิลที่ดีกว่า เพื่อพิจารณาว่านก (avialans) ควรถูกมองว่าเป็นญาติกับ Dromaeosaurids และTroodontidsมากกว่า[ 11 ]ความเห็นพ้องของนักบรรพชีวินวิทยาคือ ยังไม่มีหลักฐานเพียงพอที่จะระบุได้ว่าไดโนเสาร์กลุ่มดรอเมโอซอริเดสามารถบินหรือร่อนได้หรือไม่ หรือว่าพวกมันวิวัฒนาการมาจากบรรพบุรุษที่สามารถทำได้[ 36 ]
ทฤษฎีทางเลือกและภาวะบินไม่ได้
ไดโนเสาร์กลุ่ม Dromaeosaurids มีลักษณะคล้ายนกมากจนนักวิจัยบางคนโต้แย้งว่าควรจัดประเภทพวกมันเป็นนกเสียมากกว่า ประการแรก เนื่องจากพวกมันมีขน ไดโนเสาร์กลุ่ม Dromaeosaurids (รวมถึงไดโนเสาร์เทอโรพอดกลุ่ม Coelurosaurian อื่นๆ อีกมากมาย) จึงจัดเป็น "นก" ตามความหมายดั้งเดิมของคำว่า "นก" หรือ "Aves" ซึ่งอิงจากการมีขน อย่างไรก็ตาม นักวิทยาศาสตร์คนอื่นๆ เช่นLawrence Witmerได้โต้แย้งว่าการเรียกเทอโรพอดอย่างCaudipteryxว่าเป็นนกเพียงเพราะมันมีขน อาจเป็นการขยายความหมายของคำจนเกินความหมายที่เป็นประโยชน์[ 37 ]

นักวิจัยอย่างน้อยสองกลุ่มเสนอว่าไดโนเสาร์กลุ่มดรอเมโอซอริเดอาจสืบเชื้อสายมาจากบรรพบุรุษที่บินได้ สมมติฐานเกี่ยวกับบรรพบุรุษที่บินได้ของดรอเมโอซอริเดบางครั้งเรียกว่า "นกมาก่อน" (Birds Came First หรือ BCF) โดยปกติแล้ว George Olshevsky ได้รับการยกย่องว่าเป็นผู้เขียนคนแรกของ BCF [ 38 ]ในงานของเขาเองGregory S. Paulชี้ให้เห็นลักษณะหลายประการของโครงกระดูกดรอเมโอซอริเดที่เขาตีความว่าเป็นหลักฐานว่ากลุ่มทั้งหมดได้วิวัฒนาการมาจากบรรพบุรุษไดโนเสาร์ที่บินได้ บางทีอาจเป็นสัตว์เช่นArchaeopteryxในกรณีนั้น ดรอเมโอซอริเดขนาดใหญ่จะบินไม่ได้ในภายหลัง เช่นเดียวกับนกกระจอกเทศในปัจจุบัน[ 29 ]ในปี 1988 Paul เสนอว่าดรอเมโอซอริเดอาจมีความสัมพันธ์ใกล้ชิดกับนก ในปัจจุบันมากกว่าArchaeopteryxอย่างไรก็ตาม ในปี 2002 พอลได้จัดให้โดรเมโอซอริเดและอาร์คีออปเทอริกซ์เป็นญาติใกล้ชิดกัน[ 39 ]
ในปี พ.ศ. 2545 Hwang และคณะพบว่าMicroraptorเป็นไดโนเสาร์ดรอมิโอซอริเดที่ดั้งเดิมที่สุด[ 11 ]ในปี พ.ศ. 2546 Xu และคณะได้อ้างถึงตำแหน่งพื้นฐานของMicroraptorพร้อมกับลักษณะของขนและปีก เป็นหลักฐานว่าไดโนเสาร์ดรอมิโอซอริเดบรรพบุรุษสามารถร่อนได้ ในกรณีนั้น ไดโนเสาร์ดรอมิโอซอริเดขนาดใหญ่จะเป็นสัตว์บกในภายหลัง โดยสูญเสียความสามารถในการร่อนไปในภายหลังในประวัติศาสตร์วิวัฒนาการของพวกมัน[ 26 ]
นอกจากนี้ในปี 2002 Steven Czerkas ได้บรรยายถึงCryptovolansแม้ว่าจะเป็นชื่อพ้องรองของMicroraptorก็ตาม เขาสร้างฟอสซิลขึ้นใหม่โดยไม่ถูกต้อง โดยมีเพียงปีกสองข้าง และโต้แย้งว่า dromaeosaurids เป็นสัตว์ที่บินได้โดยใช้แรงขับเคลื่อน ไม่ใช่สัตว์ที่ร่อนได้โดยใช้แรงขับเคลื่อน ต่อมาเขาได้ออกการสร้างใหม่ที่สอดคล้องกับของMicroraptor [ 40 ]

ในปี 2548 Mayr และ Peters ได้อธิบายกายวิภาคของตัวอย่างArchaeopteryx ที่ได้รับการอนุรักษ์ไว้อย่างดี และพบว่ากายวิภาคของมันคล้ายกับเทโรพอดที่ไม่ใช่นกมากกว่าที่เคยเข้าใจมาก่อน โดยเฉพาะอย่างยิ่ง พวกเขาพบว่าArchaeopteryxมีกระดูกเพดานปาก แบบดั้งเดิม นิ้วหัวแม่เท้าที่ไม่กลับด้านและนิ้วเท้าที่สองที่ยืดได้มาก การวิเคราะห์ทางวิวัฒนาการของพวกเขาทำให้เกิดผลลัพธ์ที่ขัดแย้งกันว่าConfuciusornisมีความใกล้ชิดกับMicroraptorมากกว่าArchaeopteryxทำให้ Avialae เป็นกลุ่มอนุกรมวิธานแบบพาราไฟเลติก พวกเขายังเสนอแนะว่าบรรพบุรุษของพาราเวียนสามารถบินหรือร่อนได้ และว่าดรอเมโอซอริเดและโทรโอโดนทิดไม่สามารถบินได้ในภายหลัง (หรือสูญเสียความสามารถในการร่อน) [ 42 ] [ 43 ] Corfe และ Butler วิพากษ์วิจารณ์งานนี้ในประเด็นด้านระเบียบวิธี[ 44 ]
ความท้าทายต่อสมมติฐานทางเลือกทั้งหมดเหล่านี้เกิดขึ้นเมื่อเทอร์เนอร์และเพื่อนร่วมงานในปี 2007 ได้อธิบายถึงไดโนเสาร์ดรอมิโอซอริเดชนิดใหม่ชื่อมหาคาลา (Mahakala ) ซึ่งพวกเขาพบว่าเป็นสมาชิกที่ดั้งเดิมที่สุดและพื้นฐานที่สุดของวงศ์ดรอมิโอซอริเด (Dromaeosauridae) ดั้งเดิมกว่าไมโครแรปเตอร์ (Microraptor ) มหาคาลามีแขนสั้นและไม่สามารถร่อนได้ เทอร์เนอร์และคณะยังสรุปได้ว่าการบินวิวัฒนาการขึ้นเฉพาะในกลุ่มอาวิอาเล (Avialae) และสองประเด็นนี้ชี้ให้เห็นว่าบรรพบุรุษของดรอมิโอซอริเดไม่สามารถร่อนหรือบินได้ จากการวิเคราะห์ทางคลัดิสติกนี้มหาคาลาชี้ให้เห็นว่าลักษณะดั้งเดิมของดรอมิโอซอริเดคือไม่สามารถบินได้[ 45 ]อย่างไรก็ตาม ในปี 2555 การศึกษาที่ขยายและแก้ไขใหม่โดยรวมถึงการค้นพบไดโนเสาร์ดรอเมโอซอริเดล่าสุด พบว่าXiaotingiaที่มีลักษณะคล้ายArchaeopteryxเป็นสมาชิกดั้งเดิมที่สุดของกลุ่ม Dromaeosauridae ซึ่งดูเหมือนจะบ่งชี้ว่าสมาชิกยุคแรกสุดของกลุ่มนี้อาจมีความสามารถในการบินได้[ 46 ]
อนุกรมวิธาน
วิลเลียม ดิลเลอร์ แมทธิวและบาร์นัม บราวน์เป็นผู้ตั้งวงศ์ Dromaeosauridae โดยตั้งวงศ์ย่อย (Dromaeosaurinae) ของวงศ์Deinodontidaeในปี พ.ศ. 2465 ซึ่งประกอบด้วยสกุลใหม่เพียงสกุลเดียวคือDromaeosaurus [ 47 ]
วงศ์ย่อยของ Dromaeosauridae มักมีการเปลี่ยนแปลงเนื้อหาตามการวิเคราะห์ใหม่ แต่โดยทั่วไปประกอบด้วยกลุ่มต่อไปนี้ ไดโนเสาร์กลุ่ม Dromaeosauridae จำนวนหนึ่งไม่ได้รับการจัดให้อยู่ในวงศ์ย่อยใดโดยเฉพาะ บ่อยครั้งเนื่องจากซากดึกดำบรรพ์ของพวกมันอยู่ในสภาพที่ไม่ดีเกินกว่าจะจัดวางได้อย่างมั่นใจใน การวิเคราะห์ ทางวิวัฒนาการ (ดูส่วนวิวัฒนาการด้านล่าง) หรือไม่สามารถระบุได้แน่ชัด โดยถูกจัดให้อยู่ในกลุ่มต่างๆ ขึ้นอยู่กับวิธีการที่ใช้ในเอกสารต่างๆ วงศ์ย่อยที่เก่าแก่ที่สุดของ Dromaeosauridae ที่รู้จักคือ Halszkaraptorinae ซึ่งเป็นกลุ่มของสิ่งมีชีวิตที่แปลกประหลาด มีนิ้วและคอยาว มีฟันขนาดเล็กจำนวนมาก และอาจมีพฤติกรรมกึ่งน้ำกึ่งบก[ 48 ]อีกกลุ่มหนึ่งที่ลึกลับคือ Unenlagiinae ซึ่งเป็นวงศ์ย่อยของ Dromaeosauridae ที่ได้รับการสนับสนุนน้อยที่สุด และเป็นไปได้ว่าสมาชิกบางส่วนหรือทั้งหมดอยู่ในกลุ่มอื่นนอกเหนือจาก Dromaeosauridae [ 49 ] [ 50 ]สมาชิกที่อาศัยอยู่บนพื้นดินขนาดใหญ่ เช่นBuitreraptorและUnenlagiaแสดงให้เห็นถึงการปรับตัวในการบินที่แข็งแกร่ง แม้ว่าพวกมันอาจจะมีขนาดใหญ่เกินกว่าที่จะ 'บินขึ้น' ได้ สมาชิกที่เป็นไปได้ตัวหนึ่งในกลุ่มนี้คือRahonavisซึ่งมีขนาดเล็กมาก มีปีกที่พัฒนาอย่างดีซึ่งแสดงหลักฐานของปุ่มขน (จุดยึดสำหรับขนปีก) และเป็นไปได้มากว่ามันสามารถบินได้ กลุ่มดรอเมโอซอริเดที่ดั้งเดิมที่สุดถัดไปคือ Microraptoria กลุ่มนี้รวมถึงดรอเมโอซอริเดขนาดเล็กที่สุดหลายตัว ซึ่งแสดงให้เห็นถึงการปรับตัวสำหรับการอาศัยอยู่บนต้นไม้ รอยประทับผิวหนังของดรอเมโอซอริเดที่รู้จักทั้งหมดมาจากกลุ่มนี้และทั้งหมดแสดงให้เห็นถึงการปกคลุมของขนอย่างกว้างขวางและปีกที่พัฒนาอย่างดี เช่นเดียวกับ unenlagiines บางชนิดอาจมีความสามารถในการบินอย่างคล่องแคล่ว กลุ่มย่อยที่ก้าวหน้าที่สุดของดรอเมโอซอริเด Eudromaeosauria รวมถึงสกุลที่มีรูปร่างกำยำและขา pendek ซึ่งน่าจะเป็นนักล่าแบบซุ่มโจมตี กลุ่มนี้ประกอบด้วย Velociraptorinae, Dromaeosaurinae และในบางการศึกษายังรวมถึงกลุ่มที่สามด้วย คือ Saurornitholestinae วงศ์ย่อย Velociraptorinae นั้นโดยทั่วไปแล้วจะรวมถึงVelociraptor , DeinonychusและSaurornitholestesและในขณะที่การค้นพบTsaaganให้การสนับสนุนการจัดกลุ่มนี้ การรวมDeinonychus , Saurornitholestesและสกุลอื่นๆ อีกเล็กน้อยยังคงไม่แน่นอน Dromaeosaurinae มักพบว่าประกอบด้วยสายพันธุ์ขนาดกลางถึงขนาดใหญ่ โดยทั่วไปแล้วจะมีกะโหลกรูปทรงกล่อง (วงศ์ย่อยอื่นๆ โดยทั่วไปจะมีจมูกที่แคบกว่า) [ 5 ]





การจำแนกประเภทสกุลต่างๆ ของดรอเมโอซอริเดต่อไปนี้เป็นไปตามตารางที่ให้ไว้ใน Holtz, 2011 เว้นแต่จะระบุไว้เป็นอย่างอื่น[ 5 ]
- วงศ์ Dromaeosauridae
วิวัฒนาการ
พอล เซเรโนเป็นผู้กำหนดกลุ่มสายพันธุ์ ( clade) เป็นครั้งแรก ในปี 1998 โดยเป็นกลุ่มธรรมชาติที่ครอบคลุมมากที่สุด ซึ่งประกอบด้วยDromaeosaurusแต่ไม่รวมTroodon , OrnithomimusหรือPasserวงศ์ย่อยต่างๆ ก็ได้รับการกำหนดใหม่เป็นกลุ่มสายพันธุ์เช่นกัน โดยปกติแล้วจะกำหนดให้เป็นกลุ่มสายพันธุ์ที่ใกล้เคียงกับชื่อกลุ่มมากกว่าDromaeosaurusหรือชื่อกลุ่มย่อยอื่นๆ (ตัวอย่างเช่น มาโควิคกี้ กำหนดกลุ่มสายพันธุ์ Unenlagiinae ว่าเป็นกลุ่มสายพันธุ์ Dromaeosauridae ที่ใกล้เคียงกับUnenlagiaมากกว่าVelociraptor ) Microraptoria เป็นกลุ่มสายพันธุ์ย่อย Dromaeosauridae เพียงกลุ่มเดียวที่ไม่ได้ถูกแปลงมาจากวงศ์ย่อย เซนเตอร์และคณะได้ตั้งชื่อนี้โดยไม่ใช้คำต่อท้ายวงศ์ย่อย-inaeเพื่อหลีกเลี่ยงปัญหาที่อาจเกิดขึ้นจากการจัด กลุ่มอนุกรมวิธาน แบบดั้งเดิมหากพบว่ากลุ่มนี้อยู่นอกกลุ่ม Dromaeosauridae อย่างแท้จริง[ 52 ]เซเรโนเสนอคำจำกัดความที่แก้ไขแล้วของกลุ่มย่อยที่มีไมโครแรปเตอร์เพื่อให้แน่ใจว่าจะอยู่ในวงศ์ Dromaeosauridae และตั้งวงศ์ย่อย Microraptorinae โดยอ้างอิงถึงเซนเตอร์และคณะ แม้ว่าการใช้งานนี้จะปรากฏเฉพาะในฐานข้อมูล TaxonSearch ออนไลน์ของเขาเท่านั้นและยังไม่ได้ตีพิมพ์อย่างเป็นทางการ[ 53 ]การวิเคราะห์คลัดิสติกอย่างละเอียดที่ดำเนินการโดยเทอร์เนอร์และคณะ (2012) ยังสนับสนุนความเป็นกลุ่มเดียวของวงศ์ Dromaeosauridae อีกด้วย[ 54 ]
แผนภูมิวิวัฒนาการด้านล่างนี้เป็นไปตามการวิเคราะห์ในปี 2015 โดย DePalma et al. โดยใช้ข้อมูลที่อัปเดตจาก Theropod Working Group [ 32 ]
| ดรอเมโอซอริเด | |


แผนภูมิวิวัฒนาการอีกอันที่สร้างขึ้นด้านล่างนี้เป็นไปตามการ วิเคราะห์ทางวิวัฒนาการที่ดำเนินการในปี 2017 โดย Cau et al. โดยใช้ข้อมูลที่อัปเดตจาก Theropod Working Group ในคำอธิบายของHalszkaraptor [ 48 ]
| ดรอเมโอซอริเด | |



บรรพชีววิทยา
ประสาทสัมผัส
การเปรียบเทียบระหว่างวงแหวนสเคลอรัลของไดโนเสาร์ดรอเมโอซอริเดหลายชนิด ( ไมโครแรปเตอร์ , ซิโนร์นิโท ซอรัส และเวโลซิแรปเตอร์ ) กับนกและสัตว์เลื้อยคลานในปัจจุบัน แสดงให้เห็นว่าไดโนเสาร์ดรอเมโอซอริเดบางชนิด (รวมถึงไมโครแรปเตอร์และเวโลซิแรปเตอร์ ) อาจเป็นนักล่าที่ออกหากินในเวลากลางคืน ในขณะที่ซิโนร์นิโท ซอรัส คาดว่าจะเป็น สัตว์ ที่ออกหากินทั้ง กลางวันและกลางคืน (ออกหากินในช่วงเวลาสั้นๆ ตลอดทั้งวัน) [ 55 ]อย่างไรก็ตาม การค้นพบขนสีรุ้งในไมโครแรปเตอร์ทำให้เกิดข้อสงสัยเกี่ยวกับการอนุมานว่าไดโนเสาร์สกุลนี้ออกหากินในเวลากลางคืน เนื่องจากไม่มีนกในปัจจุบันชนิดใดที่มีขนสีรุ้งที่ทราบว่าออกหากินในเวลากลางคืน[ 56 ]
การศึกษาเกี่ยวกับปุ่มรับกลิ่น ของไดโนเสาร์กลุ่มดรอเมโอซอริเดเผยให้เห็นว่าพวกมันมีอัตราส่วนการรับกลิ่นที่คล้ายคลึงกันเมื่อเทียบกับขนาดของพวกมันกับ ไดโนเสาร์เทอโรพอดที่ไม่ใช่นกชนิดอื่นๆและนกสมัยใหม่ที่มีประสาทรับกลิ่นที่เฉียบคม เช่นไดโนเสาร์กลุ่มไทแรนโนซอริเดและนกแร้งไก่งวงซึ่งอาจสะท้อนถึงความสำคัญของประสาทรับกลิ่นในกิจกรรมประจำวันของไดโนเสาร์กลุ่มดรอเมโอซอริเด เช่น การหาอาหาร[ 57 ] [ 58 ]
การให้อาหาร
พบว่าการกินอาหารของ Dromaeosaurid เป็นแบบทั่วไปของเทโรพอดกลุ่ม Coelurosauria โดยมีวิธีการกินแบบ "เจาะและดึง" ที่คล้ายกับมังกรโคโมโดในปัจจุบัน[ 59 ]การศึกษารูปแบบการสึกหรอของฟัน Dromaeosaurid โดย Angelica Torices และคณะ แสดงให้เห็นว่าฟันของ Dromaeosaurid มีรูปแบบการสึกหรอคล้ายกับที่พบใน Tyrannosauridae และ Troodontidae อย่างไรก็ตาม การสึกหรอระดับจุลภาคบนฟันบ่งชี้ว่า Dromaeosaurid น่าจะชอบเหยื่อที่มีขนาดใหญ่กว่า Troodontidae ซึ่งมักอาศัยอยู่ในสภาพแวดล้อมเดียวกัน การแบ่งแยกทางด้านอาหารเช่นนี้อาจทำให้พวกมันสามารถอาศัยอยู่ในสภาพแวดล้อมเดียวกันได้ การศึกษาเดียวกันนี้ยังระบุว่าไดโนเสาร์กลุ่มดรอเมโอซอริเด เช่นดรอเมโอซอรัสและซอโรร์นิโธเลสเตส (ไดโนเสาร์ดรอเมโอซอริเดสองชนิดที่วิเคราะห์ในการศึกษา) น่าจะกินกระดูกเป็นอาหาร และปรับตัวได้ดีกว่าในการจัดการกับเหยื่อที่ดิ้นรน ในขณะที่ไดโนเสาร์กลุ่มโทรโอโดนทิด ซึ่งมีขากรรไกรที่อ่อนแอกว่า จะล่าสัตว์ที่อ่อนนุ่มกว่าและเหยื่อประเภทอื่นๆ เช่น สัตว์ไม่มีกระดูกสันหลังและซากสัตว์[ 60 ]
ฟังก์ชันกรงเล็บ
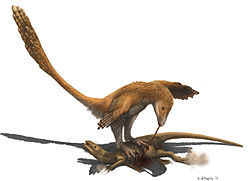

ปัจจุบันมีความเห็นไม่ตรงกันเกี่ยวกับหน้าที่ของ "กรงเล็บเคียว" ที่ขยายใหญ่ขึ้นบนนิ้วเท้าที่สอง เมื่อจอห์น ออสตรอม อธิบายถึงกรงเล็บนี้สำหรับไดโนนิคัสในปี 1969 เขาตีความกรงเล็บนี้ว่าเป็นอาวุธฟันคล้ายใบมีด คล้ายกับเขี้ยวของแมวเขี้ยวเสือ บางชนิด ใช้ร่วมกับการเตะที่ทรงพลังเพื่อตัดเหยื่อ อดัมส์ (1987) เสนอว่ากรงเล็บนี้ใช้สำหรับคว้านท้องไดโนเสาร์เซราทอปเซียนขนาด ใหญ่ [ 61 ]การตีความกรงเล็บเคียวว่าเป็นอาวุธสังหารนั้นใช้ได้กับไดโนเสาร์ดรอเมโอซอริเดทั้งหมด อย่างไรก็ตาม แมนนิงและคณะแย้งว่ากรงเล็บนี้ทำหน้าที่เป็นตะขอ โดยสร้างปลอกเคราตินขึ้นใหม่โดยมีหน้าตัดเป็นรูปวงรี แทนที่จะเป็นรูปหยดน้ำคว่ำอย่างที่เคยอนุมานไว้ก่อนหน้านี้[ 62 ]ในการตีความของแมนนิง กรงเล็บนิ้วเท้าที่สองจะใช้เป็นเครื่องช่วยปีนป่ายเมื่อปราบเหยื่อขนาดใหญ่ และยังใช้เป็นอาวุธแทงอีกด้วย
ออสตรอมเปรียบเทียบไดโนนิคัสกับนกกระจอกเทศและนกแคสโซวารีเขาตั้งข้อสังเกตว่านกชนิดนี้สามารถสร้างความเสียหายร้ายแรงได้ด้วยกรงเล็บขนาดใหญ่บนนิ้วเท้าที่สอง[ 15 ]นกแคสโซวารีมีกรงเล็บยาวถึง 125 มิลลิเมตร (4.9 นิ้ว) [ 63 ]ออสตรอมอ้างถึงกิลลิอาร์ด (1958) ที่กล่าวว่ากรงเล็บเหล่านั้นสามารถตัดแขนหรือคว้านท้องคนได้[ 64 ]คอฟรอน (1999 และ 2003) ศึกษาการโจมตีของนกแคสโซวารีที่บันทึกไว้ 241 ครั้ง และพบว่ามีมนุษย์ 1 คนและสุนัข 2 ตัวเสียชีวิต แต่ไม่มีหลักฐานว่านกแคสโซวารีสามารถคว้านท้องหรือฉีกร่างสัตว์อื่นได้[ 65 ] [ 66 ]นกแคสโซวารีใช้กรงเล็บเพื่อป้องกันตัวเอง โจมตีสัตว์ที่คุกคาม และในการแสดงการต่อสู้ เช่น การแสดงการข่มขู่ด้วยการโค้งคำนับ[ 63 ]เซริเอมายังมีกรงเล็บนิ้วเท้าที่สองที่ขยายใหญ่ขึ้น และใช้มันฉีกเหยื่อขนาดเล็กเพื่อกลืนกิน[ 67 ]
ฟิลิปป์ แมนนิงและเพื่อนร่วมงาน (2009) พยายามทดสอบหน้าที่ของกรงเล็บเคียวและกรงเล็บที่มีรูปร่างคล้ายกันบนแขนขาหน้า พวกเขาได้วิเคราะห์กลศาสตร์ชีวภาพของการกระจายแรงกดและแรงดึงไปตามกรงเล็บและไปยังแขนขา โดยใช้ การถ่ายภาพ รังสีเอกซ์เพื่อสร้างแผนที่โครงร่างสามมิติของกรงเล็บแขนขาหน้าจากเวโลซิแรปเตอร์เพื่อเปรียบเทียบ พวกเขาได้วิเคราะห์โครงสร้างของกรงเล็บจากนกนักล่าสมัยใหม่ เช่นนกฮูกเหยี่ยวพวกเขาพบว่า จากวิธีการส่งผ่านแรงกดไปตามกรงเล็บ กรงเล็บเหล่านั้นเหมาะสำหรับการปีนป่าย นักวิทยาศาสตร์พบว่าปลายแหลมของกรงเล็บเป็นเครื่องมือสำหรับเจาะและจับยึด ในขณะที่ฐานกรงเล็บที่โค้งและขยายออกช่วยถ่ายโอนแรงกดอย่างสม่ำเสมอ ทีมของแมนนิงยังได้เปรียบเทียบความโค้งของ "กรงเล็บเคียว" บนเท้าของไดโนเสาร์กลุ่มโดรเมโอซอริเดกับความโค้งในนกและสัตว์เลี้ยงลูกด้วยนมในปัจจุบันด้วย การศึกษาก่อนหน้านี้แสดงให้เห็นว่าปริมาณความโค้งของกรงเล็บสอดคล้องกับวิถีชีวิตของสัตว์: สัตว์ที่มีกรงเล็บโค้งมากในรูปทรงเฉพาะมักจะเป็นสัตว์ปีนป่าย ในขณะที่กรงเล็บที่ตรงกว่าบ่งบอกถึงวิถีชีวิตที่อาศัยอยู่บนพื้นดิน กรงเล็บเคียวของไดโนนิคัสในกลุ่มโดรเมโอซอริเดียมีความโค้ง 160 องศา ซึ่งอยู่ในช่วงของสัตว์ปีนป่าย กรงเล็บแขนขาหน้าที่พวกเขาศึกษาก็อยู่ในช่วงความโค้งของการปีนป่ายเช่นกัน[ 68 ]
นักบรรพชีวินวิทยา ปีเตอร์ แมคโควิคกี้ แสดงความคิดเห็นเกี่ยวกับการศึกษาของทีมแมนนิง โดยระบุว่าไดโนเสาร์ดรอเมโอซอริเดขนาดเล็กและดั้งเดิม (เช่นไมโครแรปเตอร์ ) น่าจะปีนต้นไม้ได้ แต่การปีนต้นไม้ไม่ได้อธิบายว่าทำไมไดโนเสาร์ดรอเมโอซอริเดขนาดยักษ์ในภายหลัง เช่นอะคิลโลเบเตอร์จึงยังคงมีกรงเล็บโค้งงอมาก ทั้งๆ ที่พวกมันมีขนาดใหญ่เกินกว่าจะปีนต้นไม้ได้ แมคโควิคกี้ตั้งข้อสันนิษฐานว่าไดโนเสาร์ดรอเมโอซอริเดขนาดยักษ์อาจปรับกรงเล็บให้ใช้สำหรับจับเหยื่อโดยเฉพาะ[ 69 ]

ในปี 2009 ฟิล เซนเตอร์ ได้ตีพิมพ์งานวิจัยเกี่ยวกับนิ้วเท้าของไดโนเสาร์ดรอเมโอซอริเด และแสดงให้เห็นว่าช่วงการเคลื่อนไหวของนิ้วเท้านั้นสอดคล้องกับการขุดรังแมลงที่แข็งแรง เซนเตอร์เสนอว่าไดโนเสาร์ดรอเมโอซอริเดขนาดเล็ก เช่นราโฮนาวิสและบิวเทอร์แรปเตอร์มีขนาดเล็กพอที่จะกินแมลงเป็นอาหาร บางส่วน ในขณะที่ไดโนเสาร์สกุลใหญ่กว่า เช่นไดโนนิคัสและนอยเควนแรปเตอร์อาจใช้ความสามารถนี้ในการจับ เหยื่อมี กระดูกสันหลังที่อาศัยอยู่ในรังแมลง อย่างไรก็ตาม เซนเตอร์ไม่ได้ทดสอบว่าความโค้งงอที่แข็งแรงของกรงเล็บไดโนเสาร์ดรอเมโอซอริเดนั้นเอื้อต่อกิจกรรมดังกล่าวด้วยหรือไม่[ 70 ]
ในปี 2011 เดนเวอร์ ฟาวเลอร์และเพื่อนร่วมงานได้เสนอวิธีการใหม่ที่ไดโนเสาร์กลุ่มดรอมิโอซอริเดอาจใช้ในการล่าเหยื่อขนาดเล็ก แบบจำลองนี้เรียกว่าแบบจำลองการล่าเหยื่อแบบ "การตรึงเหยื่อของแรปเตอร์" (RPR) ซึ่งเสนอว่าไดโนเสาร์กลุ่มดรอมิโอซอริเดฆ่าเหยื่อในลักษณะที่คล้ายคลึงกับ นกเหยี่ยว แอคซิพิทริ ดในปัจจุบัน โดยการกระโดดเข้าใส่เหยื่อ กดเหยื่อไว้ใต้ร่าง และจับเหยื่ออย่างแน่นหนาด้วยกรงเล็บรูปเคียวขนาดใหญ่ เช่นเดียวกับนกเหยี่ยวแอคซิพิทริด ไดโนเสาร์กลุ่มดรอมิโอซอริเดจะเริ่มกินเหยื่อที่ยังมีชีวิตอยู่ จนกระทั่งเหยื่อตายในที่สุดจากการเสียเลือดและอวัยวะล้มเหลว ข้อเสนอนี้อิงจากการเปรียบเทียบรูปร่างและสัดส่วนของเท้าและขาของไดโนเสาร์กลุ่มดรอมิโอซอริเดกับนกเหยี่ยวหลายกลุ่มในปัจจุบันที่มีพฤติกรรมการล่าเหยื่อที่เป็นที่รู้จัก ฟาวเลอร์พบว่าเท้าและขาของไดโนเสาร์กลุ่มดรอมิโอซอริเดนั้นคล้ายคลึงกับนกอินทรีและเหยี่ยว มากที่สุด โดยเฉพาะอย่างยิ่งในแง่ของการมีกรงเล็บที่สองที่ใหญ่ขึ้นและช่วงการเคลื่อนไหวในการจับที่คล้ายคลึงกัน อย่างไรก็ตาม กระดูกฝ่าเท้า ที่สั้น และความแข็งแรงของเท้าจะคล้ายคลึงกับนกฮูก มากกว่า วิธีการล่าเหยื่อแบบ RPR นั้นสอดคล้องกับลักษณะทางกายวิภาคอื่นๆ ของไดโนเสาร์กลุ่มดรอมิโอซอริเด เช่น ลักษณะฟันที่ผิดปกติและรูปร่างของแขน แขนซึ่งสามารถออกแรงได้มากแต่คาดว่าปกคลุมด้วยขนยาว อาจถูกใช้เป็นตัวช่วยในการทรงตัวขณะอยู่บนตัวเหยื่อที่กำลังดิ้นรน ร่วมกับหางที่แข็งแรงเพื่อช่วยในการทรงตัว ขากรรไกรของไดโนเสาร์กลุ่มดรอมิโอซอริเด ซึ่งฟาวเลอร์และเพื่อนร่วมงานคิดว่าค่อนข้างอ่อนแอ จะมีประโยชน์ในการกินเหยื่อที่ยังมีชีวิตอยู่ แต่ไม่ค่อยมีประโยชน์สำหรับการจัดการเหยื่ออย่างรวดเร็วและรุนแรง การปรับตัวเพื่อการล่าเหยื่อเหล่านี้ที่ทำงานร่วมกันอาจมีนัยสำคัญต่อต้นกำเนิดของการกระพือปีกในไดโนเสาร์กลุ่มพาราเวียนด้วย[ 71 ] [ 72 ]

ในปี 2019 ปีเตอร์ บิชอป ได้สร้างโครงกระดูกขาและกล้ามเนื้อของไดโนนิคัส ขึ้นใหม่ โดยใช้แบบจำลองสามมิติของกล้ามเนื้อเส้นเอ็นและกระดูกโดยการเพิ่มแบบจำลองทางคณิตศาสตร์และสมการ บิชอปได้จำลองสภาวะที่จะให้แรงสูงสุดที่ปลายกรงเล็บเคียว และดังนั้นจึงเป็นหน้าที่ที่น่าจะเป็นไปได้มากที่สุด ในบรรดารูปแบบการใช้งานกรงเล็บเคียวที่เสนอไว้ ได้แก่ การเตะเพื่อตัด ฟัน หรือคว้านท้องเหยื่อ การจับยึดด้านข้างของเหยื่อ การเจาะโดยใช้แรงจากน้ำหนักตัว การโจมตีบริเวณสำคัญของเหยื่อ การจับเหยื่อไว้การแข่งขันภายในหรือ ระหว่างสายพันธุ์ และการขุดหาเหยื่อจากที่ซ่อน ผลลัพธ์ที่บิชอปได้รับแสดงให้เห็นว่า ท่าทางหมอบลงจะเพิ่มแรงของกรงเล็บ อย่างไรก็ตาม แรงเหล่านี้ยังคงค่อนข้างอ่อน ซึ่งบ่งชี้ว่ากรงเล็บไม่แข็งแรงพอที่จะใช้ในการฟัน แทนที่จะใช้สำหรับการฟัน กรงเล็บเคียวมีแนวโน้มที่จะมีประโยชน์มากกว่าในมุมขาที่งอ เช่น การจับเหยื่อและการแทงเหยื่อในระยะใกล้ ผลลัพธ์เหล่านี้สอดคล้องกับ ตัวอย่าง Fighting Dinosaursซึ่งเก็บรักษาVelociraptorและProtoceratopsที่กำลังต่อสู้กัน โดย Velociraptor ใช้กรงเล็บจับอีกตัวหนึ่งในท่าขาที่ไม่เหยียดตรง แม้จะมีผลลัพธ์ที่ได้ Bishop ก็พิจารณาว่าความสามารถของกรงเล็บเคียวอาจแตกต่างกันไปในแต่ละกลุ่ม เนื่องจากในกลุ่ม dromaeosaurids นั้นAdasaurusมีกรงเล็บเคียวที่เล็กกว่าปกติซึ่งยังคงมีโครงสร้าง ginglymoid ที่เป็นลักษณะเฉพาะ ซึ่งเป็นโครงสร้างที่แบ่งออกเป็นสองส่วน และพื้นผิวข้อต่อที่ยืดหยุ่นได้มากของกระดูกนิ้วเท้าก่อนสุดท้าย เขาไม่สามารถยืนยันหรือปฏิเสธได้ว่านิ้วเท้าที่สองอาจสูญเสียหรือยังคงรักษาการทำงานไว้ได้[ 73 ]
งานวิจัยปี 2020 โดย Gianechini et al. ยังระบุว่า เวโลซิแรปทอรีน โดรเมโอซอรีน และยูโดรเมโอซอรัสอื่นๆ ในลอราเซีย มีความแตกต่างอย่างมากในเทคนิคการเคลื่อนที่และการล่าเหยื่อจากโดรเมโอซอริเดียกลุ่มอูเนนลาไจน์ในกอนด์วานา กระดูกนิ้วเท้าที่สองที่สั้นกว่าในนิ้วที่สองของเท้าช่วยให้สามารถสร้างแรงได้มากขึ้น ซึ่งเมื่อรวมกับกระดูกฝ่าเท้าที่สั้นและกว้างกว่า และลักษณะทางสัณฐานวิทยาที่คล้ายบานพับที่เห็นได้ชัดของพื้นผิวข้อต่อของกระดูกฝ่าเท้าและกระดูกนิ้วเท้า อาจทำให้ยูโดรเมโอซอรัสมีแรงยึดจับที่มากกว่าอูเนนลาไจน์ ทำให้สามารถปราบและฆ่าเหยื่อขนาดใหญ่ได้อย่างมีประสิทธิภาพมากขึ้น เมื่อเปรียบเทียบกันแล้ว ไดโนเสาร์ดรอมิโอซอริเดในกลุ่มอูเนนลาไจน์มีกระดูกฝ่าเท้าส่วนใต้ที่ยาวและเรียวกว่า และข้อต่อแบบบานพับที่เห็นได้ไม่ชัดเจนนัก ซึ่งลักษณะดังกล่าวอาจทำให้พวกมันมีความสามารถในการวิ่งที่มากขึ้นและมีความเร็วมากกว่า นอกจากนี้ กระดูกนิ้วเท้าที่สองที่ยาวกว่าของนิ้วที่สองยังช่วยให้ไดโนเสาร์กลุ่มอูเนนลาไจน์สามารถเคลื่อนไหวนิ้วที่สองของเท้าได้อย่างรวดเร็วเพื่อล่าเหยื่อที่มีขนาดเล็กกว่าและจับยากกว่า ความแตกต่างเหล่านี้ในด้านการเคลื่อนที่และความเชี่ยวชาญในการล่าเหยื่ออาจเป็นคุณลักษณะสำคัญที่มีอิทธิพลต่อเส้นทางการวิวัฒนาการที่ก่อให้เกิดไดโนเสาร์ดรอมิโอซอริเดทั้งสองกลุ่มในซีกโลกเหนือและซีกโลกใต้[ 74 ]
พฤติกรรมกลุ่ม

ฟอสซิล ของไดโนนิคัสถูกค้นพบเป็นกลุ่มเล็กๆ ใกล้กับซากของเทโนนโทซอ รัส ซึ่งเป็นไดโนเสาร์กินพืชขนาดใหญ่ใน กลุ่ม ออร์นิธิสเชียนสิ่งนี้ถูกตีความว่าเป็นหลักฐานว่าไดโนเสาร์ดรอเมโอซอริเดเหล่านี้ล่าเหยื่อเป็นฝูงอย่างเป็นระบบเช่นเดียวกับสัตว์เลี้ยงลูกด้วยนมใน ปัจจุบันบางชนิด [ 76 ]อย่างไรก็ตามนักบรรพชีวินวิทยา บางคนไม่ เห็นด้วยกับหลักฐานนี้ และการศึกษาต่อมาที่ตีพิมพ์ในปี 2550 โดย Roach และ Brinkman ชี้ให้เห็นว่าไดโนนิคัสอาจแสดงพฤติกรรมการล่าแบบรวมกลุ่มที่ไม่เป็นระเบียบไดแอพซิด ในปัจจุบัน รวมถึงนกและจระเข้ (ญาติสนิทที่สุดของดรอเมโอซอริเด) แสดงพฤติกรรมการล่าแบบร่วมมือกันในระยะยาวน้อยมาก (ยกเว้นเหยี่ยวอะพลอมาโดและเหยี่ยวแฮร์ริส ) โดยปกติแล้วพวกมันจะเป็นนักล่าเดี่ยว อาจรวมพลังกันเป็นครั้งคราวเพื่อเพิ่มโอกาสในการล่า (เช่นเดียวกับจระเข้บางครั้ง) หรือถูกดึงดูดไปยังซากสัตว์ที่ถูกฆ่าก่อนหน้านี้ ซึ่งมักเกิดความขัดแย้งระหว่างสัตว์ชนิดเดียวกัน ตัวอย่างเช่น ในสถานการณ์ที่มังกรโคโมโด กลุ่มหนึ่ง กำลังกินอาหารด้วยกัน ตัวที่ใหญ่ที่สุดจะกินก่อน และอาจโจมตีมังกรโคโมโดตัวเล็กกว่าที่พยายามจะกินอาหาร หากสัตว์ตัวเล็กกว่าตาย มันมักจะถูกกินเป็นอาหารของพวกเดียวกันเอง เมื่อนำข้อมูลนี้ไปใช้กับแหล่งที่พบพฤติกรรมการล่าเป็นฝูงของไดโนเสาร์กลุ่มดรอเมโอซอริเด มันดูค่อนข้างสอดคล้องกับกลยุทธ์การกินอาหารแบบมังกรโคโมโดโครงกระดูกของไดโนนิคัสที่พบในแหล่งเหล่านี้เป็นของตัวที่ยังไม่โตเต็มวัย โดยมีส่วนที่หายไปซึ่งอาจถูกไดโนนิคัสตัว อื่นกิน ซึ่งการศึกษาของ Roach และคณะได้นำเสนอเป็นหลักฐานที่ขัดแย้งกับแนวคิดที่ว่าสัตว์เหล่านี้ร่วมมือกันในการล่า [ 77 ]การศึกษาในปี 2020 โดย Frederickson และเพื่อนร่วมงานพบว่าความชอบด้านอาหารระหว่างไดโนนิคั สวัยเยาว์และวัยผู้ใหญ่ นั้นแตกต่างกัน แสดงให้เห็นว่าการให้อาหารของพ่อแม่สิ้นสุดลงก่อนที่ลูกจะโตพอที่จะกินอาหารแบบผู้ใหญ่ทั่วไปได้ ซึ่งจะบ่งชี้ว่าสกุลนี้ไม่ได้แสดงพฤติกรรมการล่าเป็นฝูงเหมือนสัตว์เลี้ยงลูกด้วยนม ถึงกระนั้น พวกเขาก็ยังพิจารณาว่าการอยู่รวมกันเป็นฝูงเป็นไปได้ในไดโนนิคัสวิถีชีวิตของมังกรโคโมโดก็ถูกวิพากษ์วิจารณ์เช่นกัน เนื่องจากขาดการกระจายตัวของลูกมังกรและมังกรโตเต็มวัยในพื้นที่ ซึ่งบ่งชี้ว่าวิถีชีวิตการกินพวกเดียวกันลดลง [ 78 ]
ในปี 2544 มีการค้นพบซากดึกดำบรรพ์ ของยูทาห์แรปเตอร์ หลายตัว ที่มีอายุตั้งแต่โตเต็มวัยไปจนถึงลูกอ่อนตัวเล็กยาวเพียงสามฟุต ณ สถานที่ที่บางคนเชื่อว่าเป็นกับดักนักล่าในทรายดูด บางคนถือว่านี่เป็นหลักฐานของพฤติกรรมการล่าแบบเป็นครอบครัว อย่างไรก็ตาม ก้อนหินทรายทั้งหมดยังไม่ถูกเปิดออก และนักวิจัยยังไม่แน่ใจว่าสัตว์เหล่านั้นตายพร้อมกันหรือไม่[ 79 ]เฟรเดอริคสันและเพื่อนร่วมงานแนะนำว่านี่อาจเป็นสัญญาณของพฤติกรรมอยู่รวมกันเป็นฝูงในยูทาห์แรปเตอร์และดรอเมโอซอริเดที่แสดงการดูแลลูกอ่อนหลังฟักออกจาก รัง [ 78 ]
ในปี 2550 นักวิทยาศาสตร์ได้อธิบายร่องรอยเท้า ของไดโนเสาร์ด รอเมโอซอริเด ขนาดใหญ่เป็นครั้งแรก ในมณฑลซานตง ประเทศจีน นอกจากจะยืนยันสมมติฐานที่ว่ากรงเล็บเคียวถูกเก็บกลับไว้เหนือพื้นดินแล้ว ร่องรอยเท้า (ที่สร้างโดยไดโนเสาร์สายพันธุ์ขนาดใหญ่เท่ากับ Achillobator) ยังแสดงหลักฐานว่ามีไดโนเสาร์ 6 ตัวที่มีขนาดใกล้เคียงกันเคลื่อนที่ไปด้วยกันตามแนวชายฝั่ง ไดโนเสาร์แต่ละตัวอยู่ห่างกันประมาณ 1 เมตร เดินทางไปในทิศทางเดียวกัน และเดินด้วยความเร็วค่อนข้างช้า ผู้เขียนบทความที่อธิบายรอยเท้าเหล่านี้ตีความร่องรอยเท้าว่าเป็นหลักฐานว่าไดโนเสาร์ดรอเมโอซอริเดบางสายพันธุ์อาศัยอยู่เป็นกลุ่ม แม้ว่าร่องรอยเท้าจะไม่แสดงถึงพฤติกรรมการล่าเหยื่ออย่างชัดเจน แต่ผู้เขียนก็ไม่สามารถตัดความเป็นไปได้ที่กลุ่มไดโนเสาร์ดรอเมโอซอริเดอาจล่าเหยื่อด้วยกันได้[ 21 ]
การบินและการร่อน

แขนส่วนหน้าของไดโนเสาร์กลุ่ม ดรอเมโอซอริเดดูเหมือนจะปรับตัวได้ดีในการต้านทานแรงบิดและแรงดัดที่เกี่ยวข้องกับการกระพือปีกและการร่อน[ 80 ]และความสามารถในการบินหรือร่อนได้รับการเสนอแนะสำหรับไดโนเสาร์กลุ่มดรอเมโอซอริเดอย่างน้อยห้าชนิด ชนิดแรกคือ Rahonavis ostromi (เดิมทีจัดอยู่ในกลุ่มนก แต่พบว่าเป็นไดโนเสาร์กลุ่มดรอเมโอซอริเดในการศึกษาครั้งต่อมา[ 17 ] [ 81 ] ) อาจมีความสามารถในการบิน ได้ ดังที่แสดงโดยแขนส่วนหน้าที่ยาวพร้อมหลักฐานการยึดติดของปุ่มขนสำหรับขนปีกที่ยาวและแข็งแรง[ 82 ]แขนส่วนหน้าของRahonavisมีโครงสร้างที่แข็งแรงกว่าArchaeopteryxและแสดงหลักฐานว่ามีเอ็นที่แข็งแรงซึ่งจำเป็นสำหรับการบินแบบกระพือปีก Luis Chiappe สรุปว่า ด้วยการปรับตัวเหล่านี้Rahonavisอาจบินได้ แต่คงจะไม่คล่องแคล่วในอากาศเท่ากับนกในปัจจุบัน[ 83 ]
ไดโนเสาร์ ดรอเมโอซอริเดอีกชนิดหนึ่งคือ ไมโครแรปเตอร์ กุย อาจมีความสามารถในการร่อนโดยใช้ปีกที่พัฒนาอย่างดีทั้งที่ขาหน้าและขาหลัง การศึกษาในปี 2005 โดยซานการ์ แชตเตอร์จี ชี้ให้เห็นว่าปีกของไมโครแรปเตอร์ ทำหน้าที่เหมือน " เครื่องบินสองปีก " แบบแยกส่วนและน่าจะใช้ รูปแบบการร่อน แบบฟูโกอิดซึ่งมันจะพุ่งตัวจากที่เกาะและโฉบลงมาเป็นรูปตัวยู จากนั้นก็ยกตัวขึ้นอีกครั้งเพื่อลงจอดบนต้นไม้อีกต้น โดยใช้หางและปีกหลังช่วยควบคุมตำแหน่งและความเร็ว แชตเตอร์จียังพบว่าไมโครแรปเตอร์มีคุณสมบัติพื้นฐานที่จำเป็นในการบินด้วยพลังงานในระดับคงที่ นอกเหนือจากการร่อน[ 25 ]
Changyuraptor yangiเป็นญาติใกล้ชิดกับ Microraptor guiและเชื่อกันว่าเป็นสัตว์ที่ร่อนหรือบินได้ เนื่องจากมีปีกสี่ปีกและสัดส่วนของแขนขาที่คล้ายคลึงกัน อย่างไรก็ตาม มันเป็นสัตว์ที่มีขนาดใหญ่กว่ามาก ประมาณขนาดไก่งวงป่าจัดอยู่ในกลุ่มพาราเวียนยุคมีโซโซอิกที่บินได้ขนาดใหญ่ที่สุดเท่าที่รู้จัก
ไดโนเสาร์ดรอเมโอซอริเดอีกชนิดหนึ่งคือDeinonychus antirrhopusอาจแสดงความสามารถในการบินได้บางส่วน ลูกของไดโนเสาร์ชนิดนี้มีแขนยาวกว่าและมีกระดูกอกที่แข็งแรงกว่าตัวเต็มวัย ซึ่งคล้ายกับที่พบในเทโรพอดที่กระพือปีกชนิดอื่น ๆ ซึ่งบ่งชี้ว่าพวกมันอาจสามารถบินได้เมื่อยังเล็กและสูญเสียความสามารถนั้นไปเมื่อโตขึ้น[ 84 ]
ความเป็นไปได้ที่Sinornithosaurus milleniiจะสามารถร่อนหรือแม้กระทั่งบินด้วยพลังงานได้นั้นถูกหยิบยกขึ้นมาหลายครั้ง[ 85 ] [ 86 ]แม้ว่าจะไม่มีการศึกษาเพิ่มเติมเกิดขึ้นก็ตาม
Zhenyuanlongรักษาขนปีกที่มีรูปร่างตามหลักอากาศพลศาสตร์ไว้ โดยเฉพาะอย่างยิ่งขนคลุมปีกที่คล้ายนก ซึ่งแตกต่างจากขนคลุมปีกที่ยาวและกว้างกว่าของรูปแบบต่างๆ เช่น Archaeopteryxและ Anchiornisรวมถึงแผ่นกระดูกอกที่เชื่อมติดกัน เนื่องจากขนาดและแขนสั้น จึงไม่น่าเป็นไปได้ที่ Zhenyuanlongจะสามารถบินได้ (แม้ว่าความสำคัญของการสร้างแบบจำลองทางชีวกลศาสตร์ในเรื่องนี้จะถูกเน้นย้ำ [ 31 ] ) แต่ก็อาจบ่งชี้ถึงการสืบเชื้อสายที่ค่อนข้างใกล้ชิดจากบรรพบุรุษที่บินได้ หรือแม้กระทั่งความสามารถในการร่อนหรือโดย ใช้ปีกช่วย
พยาธิวิทยาโบราณ
ในปี 2001 บรูซ รอธไชลด์และคณะได้ตีพิมพ์งานวิจัยที่ตรวจสอบหลักฐานเกี่ยวกับกระดูกร้าวจากความเครียดและการฉีกขาดของเส้นเอ็นในไดโนเสาร์เทอโรพอดและผลกระทบต่อพฤติกรรมของพวกมัน เนื่องจากกระดูกร้าวจากความเครียดเกิดจากการบาดเจ็บซ้ำๆ มากกว่าเหตุการณ์เดียว จึงมีแนวโน้มที่จะเกิดจากพฤติกรรมปกติมากกว่าการบาดเจ็บประเภทอื่นๆ นักวิจัยพบรอยโรคที่คล้ายกับที่เกิดจากกระดูกร้าวจากความเครียดบนกรงเล็บมือของไดโนเสาร์ดรอเมโอซอริเด ซึ่งเป็นหนึ่งในสองรอยโรคกรงเล็บดังกล่าวที่ค้นพบในระหว่างการศึกษา กระดูกร้าวจากความเครียดในมือมีความสำคัญทางพฤติกรรมเป็นพิเศษเมื่อเทียบกับที่พบในเท้า เนื่องจากกระดูกร้าวจากความเครียดในเท้าสามารถเกิดขึ้นได้ในขณะวิ่งหรือระหว่างการอพยพ ในทางตรงกันข้าม การบาดเจ็บที่มือมีแนวโน้มที่จะเกิดขึ้นในขณะที่สัมผัสกับเหยื่อที่กำลังดิ้นรน[ 87 ]
การว่ายน้ำ

อย่างน้อยหนึ่งกลุ่มของไดโนเสาร์ดรอเมโอซอริเด คือHalszkaraptorinaeซึ่งสมาชิกคือ halszkaraptorines มีแนวโน้มที่จะมีความเชี่ยวชาญเฉพาะด้านสำหรับพฤติกรรมในน้ำหรือกึ่งน้ำโดยมีการพัฒนาสัดส่วนของแขนขา รูปร่างของฟัน และโครงกระดูกซี่โครงที่คล้ายกับนกดำน้ำ[ 48 ] [ 88 ] [ 89 ]
มีการเสนอพฤติกรรมการตกปลาสำหรับunenlagiines ซึ่งรวมถึงการเปรียบเทียบกับ spinosauridsกึ่งน้ำ[ 90 ]แต่ยังไม่มีการกล่าวถึงกลไกการขับเคลื่อนในน้ำแต่อย่างใด
การสืบพันธุ์

ในปี 2549 Grellet-Tinner และ Makovicky รายงานไข่ที่เกี่ยวข้องกับตัวอย่างของDeinonychusไข่ดังกล่าวมีความคล้ายคลึงกับไข่ของโอวิแรปทอริด และผู้เขียนตีความว่าความเกี่ยวข้องนี้อาจบ่งชี้ถึงการฟักไข่ [ 91 ]การศึกษาที่ตีพิมพ์ในเดือนพฤศจิกายน 2561 โดย Norell, Yang และ Wiemann และคณะ ระบุว่าDeinonychusวางไข่สีฟ้า ซึ่งน่าจะเพื่อพรางตัวและสร้างรังแบบเปิด ไดโนเสาร์ดรอเมโอซอริดอื่นๆ อาจทำเช่นเดียวกัน และมีทฤษฎีว่าพวกมันและไดโนเสาร์มานิแรปทอแรนอื่นๆ อาจเป็นจุดเริ่มต้นของการวางไข่สีและสร้างรังแบบเปิดเช่นเดียวกับนกหลายชนิดในปัจจุบัน[ 92 ] [ 93 ] [ 94 ]
ในวัฒนธรรมสมัยนิยม
เวโลซิแรปเตอร์ ซึ่งเป็นไดโนเสาร์ในกลุ่ม ดรอเม โอซอริ เด ได้รับความสนใจอย่างมากหลังจากที่มันปรากฏตัวอย่างโดดเด่นในภาพยนตร์เรื่องจูราสสิกพาร์คของสตีเวน สปีลเบิร์กในปี 1993อย่างไรก็ตามขนาดของเวในภาพยนตร์นั้นใหญ่กว่าสมาชิกที่ใหญ่ที่สุดในสกุลนั้นมาก โรเบิร์ต บักเกอร์เล่าว่าสปีลเบิร์กผิดหวังกับขนาดของเวโลซิแรปเตอร์และจึงเพิ่มขนาดให้ใหญ่ขึ้น [ 95 ]เกรกอรี เอส. พอลในหนังสือ Predatory Dinosaurs of the World ปี 1988 ของเขา ยังพิจารณาว่าไดโนนิคัส แอนติร์รอปัสเป็นสายพันธุ์หนึ่งของเวโลซิแรปเตอร์และจึงตั้งชื่อสายพันธุ์ใหม่ว่าเวโลซิแรปเตอร์ แอนติร์รอปัส [ 39 ] ความคิดเห็นทางอนุกรมวิธานนี้ไม่ได้รับการยอมรับอย่างกว้างขวาง [ 10 ] [ 96 ] [ 97 ]
ลำดับเวลาของสกุลไดโนเสาร์ดรอเมโอซอริเดีย

ดูเพิ่มเติม
ลิงก์ภายนอก
- ไดโนเสาร์วงศ์ Dromaeosauridae: แรปเตอร์!จากพิพิธภัณฑ์บรรพชีวินวิทยา มหาวิทยาลัยแคลิฟอร์เนีย เบิร์กลีย์
- ไดโนเสาร์ – ฉบับสมบูรณ์และฟรีทางออนไลน์ของหนังสือ "ไดโนเสาร์" ที่เขียนโดย WD Matthew (อ้างอิงในบทความนี้ว่าเป็นผู้เขียนวงศ์ Dromaeosauridae) อดีตภัณฑารักษ์ด้านบรรพชีวินวิทยาของสัตว์มีกระดูกสันหลังที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติอเมริกันในนิวยอร์ก ตีพิมพ์ครั้งแรกในปี 1915
- ไดโนเสาร์วงศ์ Dromaeosauridae : ข้อมูลอ้างอิงเกี่ยวกับโลกไดโนเสาร์ พร้อมคำอธิบายเชิงลึกและภาพของไดโนเสาร์วงศ์ Dromaeosauridae หลายชนิด
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ดรอเมโอซอริเด
Dromaeosauridae ( / ˌ d r ɒ m i . ə ˈ s ɔːr ɪ d iː / ) เป็นวงศ์ของไดโนเสาร์เทอโรพอดโคเอลูโรซอเรียน ที่มีขน พวกมันโดยทั่วไปเป็น สัตว์กินเนื้อที่มีขนขนาดเล็กถึงขนาดกลางที่เฟื่องฟูใน.
การวินิจฉัยทางเทคนิค
Dromaeosaurids มีลักษณะเฉพาะดังต่อไปนี้: กระดูกหน้าผาก รูปตัว T สั้น ที่สร้าง ขอบเขต ด้านหน้า ของ ช่อง supratemporal fenestra ; ชั้นวางที่ยื่นออกมาทางด้านท้ายและด้านข้างของ กระดูก squamosal ; กระบวนการด้านข้างของ กระดูก quadrate ที่สัมผัสกับกระดูก...
ขนาดและโครงสร้างโดยทั่วไป
ไดโนเสาร์กลุ่มดรอเมโอซอริเดเป็นไดโนเสาร์ขนาดเล็กถึงขนาดกลาง มีความยาวตั้งแต่ 1.5–2.07 เมตร (4.9–6.
เท้า
เช่นเดียวกับเทโรพอดอื่นๆ ดรอเมโอซอริเดเป็นสัตว์สองขา กล่าวคือ พวกมันเดินด้วยขาหลัง อย่างไรก็ตาม ในขณะที่เทโรพอดส่วนใหญ่เดินโดยใช้สามนิ้วสัมผัสพื้น ร่องรอยเท้าฟอสซิลยืนยันว่า กลุ่ม พาราเวียนยุค แรกหลาย กลุ่ม รวมถึงดรอเมโอซอริเด...