อ่าน 5 นาที
อีไอเอฟ2
ปัจจัยเริ่มต้นการสังเคราะห์โปรตีนในยูคาริโอต 2 ( eIF2 ) เป็น ปัจจัยเริ่มต้น การสังเคราะห์ โปรตีนในยูคาริโอต จำเป็นสำหรับกระบวนการ เริ่มต้น การแปล รหัส ในยูคาริโอตส่วนใหญ่ eIF2...
อีไอเอฟ2
ปัจจัยเริ่มต้นการสังเคราะห์โปรตีนในยูคาริโอต 2 ( eIF2 ) เป็นปัจจัยเริ่มต้น การสังเคราะห์ โปรตีนในยูคาริโอต จำเป็นสำหรับกระบวนการ เริ่มต้น การแปลรหัส ในยูคาริโอตส่วนใหญ่ eIF2 ทำหน้าที่เชื่อมโยงtRNA iMetกับไรโบโซมใน ลักษณะที่ขึ้นอยู่กับ GTP eIF2 เป็นเฮเทอโรไตรเมอร์ที่ประกอบด้วยหน่วยย่อยอัลฟา (เรียกอีกอย่างว่าหน่วยย่อยที่ 1, EIF2S1), หน่วย ย่อย เบตา (หน่วยย่อยที่ 2, EIF2S2) และ หน่วย ย่อยแกมมา (หน่วยย่อยที่ 3, EIF2S3)
เมื่อขั้นตอนการเริ่มต้นเสร็จสมบูรณ์ eIF2 จะถูกปล่อยออกจากไรโบโซมโดยยังคงจับอยู่กับGDPในรูปของสารประกอบเชิงซ้อนแบบไบนารีที่ไม่ทำงาน เพื่อที่จะเข้าร่วมในการเริ่มต้นการแปลรหัสรอบต่อไป GDP นี้จะต้องถูกแทนที่ด้วย GTP
การทำงาน

eIF2 เป็นปัจจัยสำคัญในการสังเคราะห์โปรตีนที่สร้างสารประกอบเชิงซ้อนสามส่วน (TC) กับGTP และ tRNAตัวเริ่มต้นMet - i Metหลังจากสร้าง TC แล้ว มันจะจับกับหน่วยย่อยไรโบโซม 40Sเพื่อสร้างสารประกอบเชิงซ้อนก่อนเริ่มต้น 43S (43S PIC) เชื่อกันว่าการประกอบ 43S PIC ถูกกระตุ้นโดยปัจจัยเริ่มต้นeIF1 , eIF1Aและ สารประกอบเชิงซ้อน eIF3ตาม การทดลอง ในหลอดทดลองจากนั้น 43S PIC จะจับกับmRNAที่ถูกคลายเกลียวโดย สารประกอบเชิงซ้อน eIF4F ก่อนหน้านี้ 43S PIC และโปรตีน eIF4F จะสร้างสารประกอบเชิงซ้อน 48S ใหม่บน mRNA ซึ่งจะเริ่มค้นหาโคดอนเริ่มต้น (AUG) ตาม mRNA เมื่อเบสของโคดอน AUG จับคู่กับ Met-tRNA แล้วeIF5 (ซึ่งเป็นโปรตีนกระตุ้น GTPaseหรือ GAP) จะถูกดึงเข้าสู่คอมเพล็กซ์และกระตุ้นให้ eIF2 ไฮโดรไลซ์ GTP ของมัน ซึ่งจะทำให้ eIF2-GDP ถูกปล่อยออกจากคอมเพล็กซ์ 48S นี้ และการแปลจะเริ่มต้นขึ้นหลังจากดึง ซับยูนิตไรโบโซม 60Sและการก่อตัวของ คอมเพล็กซ์เริ่มต้น 80Sในที่สุด ด้วยความช่วยเหลือของปัจจัยแลกเปลี่ยนนิวคลีโอไทด์กัวนีน (GEF) eIF2B [ 1 ] GDP ใน eIF2 จะถูกแลกเปลี่ยนเป็น GTP และคอมเพล็กซ์สามส่วน จะก่อตัวขึ้นใหม่เพื่อเริ่มต้นการแปลรอบใหม่ [ 2 ] [ 3 ] [ 4 ]
โครงสร้าง
eIF2 เป็นเฮเทอโรไตรเมอร์ที่มีมวลโมเลกุลรวม 126 กิโลดาลตันประกอบด้วยหน่วยย่อย 3 หน่วย ได้แก่α (หน่วยย่อยที่ 1), β (หน่วยย่อยที่ 2) และγ (หน่วยย่อยที่ 3) ลำดับของหน่วยย่อยทั้งสามมีความคล้ายคลึงกันสูง (ความเหมือนของกรดอะมิโนแบบคู่ระหว่างโปรตีนของ Homo sapiensและSaccharomyces cerevisiaeอยู่ในช่วง 47 ถึง 72% )
| หน่วยย่อย | อัลฟ่า | เบต้า | แกมมา |
|---|---|---|---|
| น้ำหนักโมเลกุล / กิโลดาลตัน | 36 | 38 | 52 |
| ความคล้ายคลึงกัน | eIF2-alpha family IPR011488 | eIF2-beta / eIF5 family IPR002735 | กลุ่มปัจจัยการยืดตัวที่จับกับ GTP (และอื่นๆ) P41091 |
| ปฏิสัมพันธ์ | การจับกันของ eIF5, eIF2B และ RNA | การจับกันของ GTP และ RNA |
หน่วยย่อยอัลฟาประกอบด้วยเป้าหมายหลักสำหรับการฟอสโฟรีเลชันคือซีรีนที่ตำแหน่ง 51 นอกจากนี้ยังประกอบด้วยโดเมนโมทีฟ S1 ซึ่งเป็นตำแหน่งที่อาจจับกับ RNA ได้ ดังนั้น หน่วยย่อยอัลฟาจึงถือได้ว่าเป็นหน่วยย่อยควบคุมของไตรเมอร์
หน่วยย่อยเบตาประกอบด้วยตำแหน่งฟอสโฟรีเลชันหลายตำแหน่ง (กรดอะมิโน 2, 13, 67, 218) สิ่งสำคัญที่ควรพิจารณาคือ ยังมี กลุ่ม ไลซีน สาม กลุ่มในโดเมนปลายด้าน N (NTD) ซึ่งมีความสำคัญต่อการโต้ตอบกับ eIF2B นอกจากนี้ ลำดับของโปรตีนยังประกอบด้วยโมทีฟนิ้วสังกะสีซึ่งแสดงให้เห็นว่ามีบทบาทในการสร้างทั้งคอมเพล็กซ์สามส่วนและคอมเพล็กซ์เริ่มต้นก่อน 43S นอกจากนี้ยังมีลำดับการจับกับนิวคลีโอไทด์กัวนีนสองลำดับ ซึ่งยังไม่พบว่ามีส่วนเกี่ยวข้องกับการควบคุมกิจกรรมของ eIF2 เชื่อกันว่าหน่วยย่อยเบตายังโต้ตอบกับทั้ง tRNA และ mRNA ด้วย
หน่วยย่อย γประกอบด้วยไซต์จับนิวคลีโอไทด์กัวนีน 3 ไซต์ และเป็นที่ทราบกันว่าเป็นไซต์เชื่อมต่อหลักสำหรับ GTP/GDP นอกจากนี้ยังมีโพรงจับ tRNA ซึ่งแสดงให้เห็นโดยการวิเคราะห์ผลึกด้วยรังสีเอกซ์มอทีฟซิงค์นัคเคิลสามารถจับไอออน Zn 2+ ได้หนึ่งไอออน [ 4 ] [ 6 ] [ 7 ]เกี่ยวข้องกับปัจจัยการยืดตัวบางอย่าง เช่นEF- Tu [ 8 ]
ระเบียบข้อบังคับ
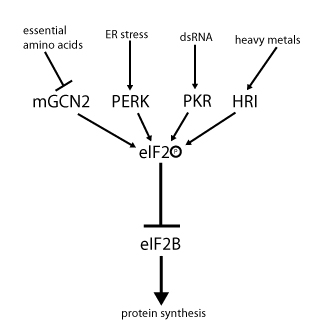
กิจกรรมของ eIF2 ถูกควบคุมโดยกลไกที่เกี่ยวข้องกับการแลกเปลี่ยนนิวคลีโอไทด์กัวนีนและการฟอสโฟรีเลชัน การฟอสโฟรีเลชันเกิดขึ้นที่ซับยูนิต α ซึ่งเป็นเป้าหมายของเซรินไคเนส จำนวนหนึ่ง ที่ฟอสโฟรีเลตเซริน 51 ไคเนสเหล่านั้นทำงานอันเป็นผลมาจากความเครียด เช่น การขาดกรดอะมิโน ( GCN2 ) ความเครียดของ ER ( PERK ) การมีอยู่ของ dsRNA ( PKR ) การขาดฮีม ( HRI ) หรืออินเตอร์เฟรอน [ 10 ] เมื่อฟอสโฟรีเลตแล้ว eIF2 จะแสดงความสัมพันธ์ที่เพิ่มขึ้นกับ eIF2B ซึ่งเป็น GEF ของมัน อย่างไรก็ตาม eIF2B สามารถแลกเปลี่ยน GDP กับ GTP ได้ก็ต่อเมื่อ eIF2 อยู่ในสถานะที่ไม่ได้รับการฟอสโฟรีเลตเท่านั้น แต่ eIF2 ที่ได้รับการฟอสโฟรีเลตแล้ว เนื่องจากมีการจับที่แข็งแรงกว่า จึงทำหน้าที่เป็นตัวยับยั้ง GEF ของตัวเอง (eIF2B) เนื่องจากความเข้มข้นของ eIF2B ในเซลล์ต่ำกว่า eIF2 มาก แม้แต่ eIF2 ที่ถูกฟอสโฟรีเลตเพียงเล็กน้อยก็สามารถยับยั้งการทำงานของ eIF2B ได้อย่างสมบูรณ์โดยการกักเก็บไว้ หากไม่มี GEF แล้ว eIF2 ก็ไม่สามารถกลับคืนสู่สถานะที่ทำงานได้ (จับกับ GTP) อีกต่อไป ผลที่ตามมาคือการแปลรหัสจะหยุดชะงักลง เนื่องจากการเริ่มต้นจะไม่สามารถเกิดขึ้นได้หากไม่มีสารประกอบเชิงซ้อนสามส่วนที่พร้อมใช้งาน นอกจากนี้ ความเข้มข้นต่ำของสารประกอบเชิงซ้อนสามส่วนยังช่วยให้เกิดการแสดงออกของ GCN4 (สภาวะอดอาหาร) ซึ่งส่งผลให้มีการกระตุ้นยีนสังเคราะห์กรดอะมิโนเพิ่มขึ้น[ 2 ] [ 3 ] [ 4 ] [ 9 ] [ 11 ]
โรค
เนื่องจาก eIF2 มีความสำคัญต่อการเริ่มต้นการแปลรหัสส่วนใหญ่ และการสังเคราะห์โปรตีน ดังนั้นความบกพร่องใน eIF2 จึงมักเป็นอันตรายถึงชีวิต โปรตีนนี้มีการอนุรักษ์สูงในสิ่งมีชีวิตที่มีวิวัฒนาการห่างไกลกัน ซึ่งบ่งชี้ว่าการกลายพันธุ์มีผลกระทบอย่างมากต่อความอยู่รอดของเซลล์ ดังนั้นจึงไม่พบโรคใด ๆ ที่เกี่ยวข้องโดยตรงกับการกลายพันธุ์ใน eIF2 อย่างไรก็ตาม มีหลายโรคที่เกิดจากการลดระดับของ eIF2 ผ่านไคเนสต้นน้ำ ตัวอย่างเช่น พบความเข้มข้นที่เพิ่มขึ้นของ PKR ที่ทำงานอยู่และ eIF2 ที่ไม่ทำงาน (ฟอสโฟรีเลต) ในผู้ป่วยที่เป็นโรคทางระบบประสาทเสื่อม เช่น โรค อัลไซเม อร์โรคพาร์กินสันและ โรค ฮันติงตันนอกจากนี้ยังมีตัวอย่างโรคที่ได้รับการพิสูจน์แล้วหนึ่งโรคที่เกี่ยวข้องกับ GEF eIF2B การกลายพันธุ์ในหน่วยย่อยทั้งห้าของ eIF2B เกี่ยวข้องกับโรค Vanishing White Matter (VWM) ซึ่งเป็นโรคทางพันธุกรรมที่ทำให้เนื้อเยื่อสีขาวในสมองเสื่อมและหายไป[ 12 ] [ 13 ]ยังไม่เป็นที่เข้าใจอย่างถ่องแท้ว่าเหตุใดจึงมีเพียงเซลล์สมองเท่านั้นที่ดูเหมือนจะได้รับผลกระทบจากความบกพร่องเหล่านี้ ระดับโปรตีนควบคุมที่ไม่เสถียรที่ลดลงอาจมีบทบาทในการพัฒนาของโรคที่กล่าวถึง[ 4 ] [ 14 ]
ดูเพิ่มเติม
- ปัจจัยเริ่มต้นของยูคาริโอต
- ไคเนสของ eIF2
- อีไอเอฟ2เอ
- อีไอเอฟ2ดี
- โรคเม็ดเลือดขาวในสมองที่มีการสูญเสียเนื้อเยื่อสีขาว
ลิงก์ภายนอก
- EIF-2 ที่ หัวข้อเรื่องทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- การเริ่มต้นการแปลรหัสที่ขึ้นอยู่กับแคปจาก Nature Reviews Microbiology เป็นภาพที่ดีและภาพรวมเกี่ยวกับหน้าที่ของปัจจัยเริ่มต้น
การควบคุมการสังเคราะห์โปรตีนในยูคาริโอต กลไกการทำงานของ eRF ซึ่งเป็นปัจจัยรีไซเคิล eIF-2 จากเรติคิวโลไซต์ของกระต่ายที่เกี่ยวข้องกับการแลกเปลี่ยน GDP/GTP Salimans M, Goumans H, Amesz H, Benne R, Voorma HO. Eur J Biochem. 1984 พฤศจิกายน
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ อีไอเอฟ2
ปัจจัยเริ่มต้นการสังเคราะห์โปรตีนในยูคาริโอต 2 ( eIF2 ) เป็น ปัจจัยเริ่มต้น การสังเคราะห์ โปรตีนในยูคาริโอต จำเป็นสำหรับกระบวนการ เริ่มต้น การแปล รหัส ในยูคาริโอตส่วนใหญ่ eIF2...
การทำงาน
eIF2 เป็นปัจจัยสำคัญในการสังเคราะห์โปรตีนที่สร้างสารประกอบเชิงซ้อนสามส่วน (TC) กับ GTP และ tRNA ตัวเริ่มต้น Met - i Met หลังจากสร้าง TC แล้ว มันจะจับกับ หน่วยย่อยไรโบโซม 40S เพื่อสร้าง สารประกอบเชิงซ้อนก่อนเริ่มต้น 43S (43S PIC) เชื่อกันว่าการประกอบ 43S PIC...
โครงสร้าง
eIF2 เป็นเฮเทอโรไตรเมอร์ที่มีมวลโมเลกุลรวม 126 กิโลดาลตัน ประกอบด้วยหน่วยย่อย 3 หน่วย ได้แก่ α (หน่วยย่อยที่ 1), β (หน่วยย่อยที่ 2) และ γ (หน่วยย่อยที่ 3) ลำดับของหน่วยย่อยทั้งสามมีความคล้ายคลึงกันสูง (ความเหมือนของกรดอะมิโนแบบคู่ระหว่างโปรตีนของ Homo sapiens...
ระเบียบข้อบังคับ
กิจกรรมของ eIF2 ถูกควบคุมโดยกลไกที่เกี่ยวข้องกับการแลกเปลี่ยนนิวคลีโอไทด์กัวนีนและการฟอสโฟรีเลชัน การฟอสโฟรีเลชันเกิดขึ้นที่ซับยูนิต α ซึ่งเป็นเป้าหมายของ เซรินไคเนส จำนวนหนึ่ง ที่ฟอสโฟรีเลต เซริน 51 ไคเนสเหล่านั้นทำงานอันเป็นผลมาจากความเครียด เช่น...