อ่าน 9 นาที
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging Theory หรือ OFT ) เป็น แบบจำลอง ทางนิเวศวิทยาเชิงพฤติกรรม ที่ช่วยทำนายพฤติกรรมของสัตว์เมื่อค้นหาอาหาร...
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด

ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging TheoryหรือOFT ) เป็น แบบจำลอง ทางนิเวศวิทยาเชิงพฤติกรรมที่ช่วยทำนายพฤติกรรมของสัตว์เมื่อค้นหาอาหาร แม้ว่าการหาอาหารจะให้พลังงานแก่สัตว์ แต่การค้นหาและจับอาหารนั้นต้องใช้ทั้งพลังงานและเวลา เพื่อให้ได้ประโยชน์สูงสุด สัตว์จะเลือกใช้ กลยุทธ์ การหาอาหารที่ให้ประโยชน์ (พลังงาน) มากที่สุดโดยใช้ต้นทุนน้อยที่สุด เพื่อเพิ่มพลังงานสุทธิที่ได้รับให้สูงสุด OFT ช่วยทำนายกลยุทธ์ที่ดีที่สุดที่สัตว์สามารถใช้เพื่อให้บรรลุเป้าหมายนี้ได้
OFT เป็นการประยุกต์ใช้แบบจำลองความเหมาะสม เชิงนิเวศวิทยา ทฤษฎีนี้ตั้งสมมติฐานว่ารูปแบบการหาอาหารที่ได้เปรียบทางเศรษฐกิจมากที่สุดจะถูกเลือกในสายพันธุ์ผ่านการคัดเลือกโดยธรรมชาติ [ 1 ] เมื่อใช้ OFT ในการจำลองพฤติกรรมการหาอาหาร สิ่งมีชีวิตจะถูกกล่าวว่ากำลังเพิ่มค่าตัวแปรที่เรียกว่าสกุลเงิน ให้สูงสุด เช่น อาหารมากที่สุดต่อหน่วยเวลา นอกจากนี้ข้อจำกัดของสิ่งแวดล้อมเป็นตัวแปรอื่นที่ต้องพิจารณา ข้อจำกัดถูกกำหนดให้เป็นปัจจัยที่สามารถจำกัดความสามารถของผู้หาอาหารในการเพิ่มสกุลเงินให้สูงสุดกฎการตัดสินใจที่เหมาะสมที่สุดหรือกลยุทธ์การหาอาหารที่ดีที่สุดของสิ่งมีชีวิต ถูกกำหนดให้เป็นการตัดสินใจที่เพิ่มสกุลเงินให้สูงสุดภายใต้ข้อจำกัดของสิ่งแวดล้อม การระบุถึงกฎการตัดสินใจที่เหมาะสมที่สุดเป็นเป้าหมายหลักของ OFT [ 2 ]ความเชื่อมโยงระหว่าง OFT และวิวัฒนาการทางชีวภาพได้รับความสนใจในช่วงหลายทศวรรษที่ผ่านมา การศึกษาเกี่ยวกับพฤติกรรมการหาอาหารที่เหมาะสมที่สุดในระดับประชากรได้ใช้แบบจำลองพลวัตการเกิด-ตายเชิงวิวัฒนาการ ในขณะที่แบบจำลองเหล่านี้ยืนยันการมีอยู่ของฟังก์ชันวัตถุประสงค์ เช่น "สกุลเงิน" ในบางสถานการณ์ แบบจำลองเหล่านี้ยังก่อให้เกิดคำถามเกี่ยวกับการประยุกต์ใช้ในข้อจำกัดอื่นๆ เช่น ปฏิสัมพันธ์ของประชากรจำนวนมาก[ 3 ]
การสร้างแบบจำลองการหาอาหารที่เหมาะสมที่สุด
แบบจำลองการหาอาหารที่เหมาะสมจะสร้างการคาดการณ์เชิงปริมาณว่าสัตว์จะเพิ่มความเหมาะสมสูงสุดได้อย่างไรในขณะที่พวกมันหาอาหาร กระบวนการสร้างแบบจำลองเกี่ยวข้องกับการระบุสกุลเงิน ข้อจำกัด และกฎการตัดสินใจที่เหมาะสมสำหรับผู้หาอาหาร[ 2 ] [ 4 ]
สกุลเงินถูกกำหนดให้เป็นหน่วยที่สัตว์ปรับให้เหมาะสมที่สุด นอกจากนี้ยังเป็นสมมติฐานเกี่ยวกับต้นทุนและผลประโยชน์ที่สัตว์นั้นต้องแบกรับ[ 5 ]ตัวอย่างเช่น สัตว์หาอาหารบางชนิดได้รับพลังงานจากอาหาร แต่ต้องเสียค่าใช้จ่ายในการค้นหาอาหาร เวลาและพลังงานที่ใช้ในการค้นหาอาจถูกนำไปใช้ในกิจกรรมอื่น ๆ เช่น การหาคู่หรือการปกป้องลูกอ่อน การเพิ่มผลประโยชน์สูงสุดด้วยต้นทุนที่ต่ำที่สุดจึงเป็นประโยชน์สูงสุดต่อสัตว์ ดังนั้น สกุลเงินในสถานการณ์นี้จึงสามารถกำหนดได้ว่าเป็นพลังงานสุทธิที่ได้รับต่อหน่วยเวลา[ 2 ]อย่างไรก็ตาม สำหรับสัตว์หาอาหารชนิดอื่น เวลาที่ใช้ในการย่อยอาหารหลังจากกินอาจเป็นต้นทุนที่สำคัญกว่าเวลาและพลังงานที่ใช้ในการค้นหาอาหาร ในกรณีนี้ สกุลเงินสามารถกำหนดได้ว่าเป็นพลังงานสุทธิที่ได้รับต่อเวลา การย่อยอาหาร แทนที่จะเป็นพลังงานสุทธิที่ได้รับต่อหน่วยเวลา[ 6 ]ยิ่งไปกว่านั้น ผลประโยชน์และต้นทุนอาจขึ้นอยู่กับชุมชนของสัตว์หา อาหารด้วย ตัวอย่างเช่น สัตว์หาอาหารที่อาศัยอยู่ในรังมักจะหาอาหารในลักษณะที่จะเพิ่มประสิทธิภาพให้กับอาณานิคมมากกว่าตัวมันเอง[ 5 ]โดยการระบุสกุลเงิน เราสามารถสร้างสมมติฐานเกี่ยวกับผลประโยชน์และต้นทุนที่สำคัญสำหรับสัตว์หาอาหารนั้นๆ ได้
ข้อจำกัดคือสมมติฐานเกี่ยวกับข้อจำกัดที่กำหนดไว้สำหรับสัตว์[ 5 ]ข้อจำกัดเหล่านี้อาจเกิดจากลักษณะของสภาพแวดล้อมหรือสรีรวิทยาของสัตว์ และอาจจำกัดประสิทธิภาพในการหาอาหารของพวกมัน เวลาที่สัตว์ใช้ในการเดินทางจากรังไปยังแหล่งหาอาหารเป็นตัวอย่างของข้อจำกัด จำนวนอาหารสูงสุดที่สัตว์สามารถนำกลับไปยังรังได้ก็เป็นอีกตัวอย่างหนึ่งของข้อจำกัด นอกจากนี้ยังอาจมีข้อจำกัดด้านการรับรู้ของสัตว์ เช่น ข้อจำกัดในการเรียนรู้และความจำ[ 2 ]ยิ่งสามารถระบุข้อจำกัดในระบบที่กำหนดได้มากเท่าใด แบบจำลองก็จะมีพลังในการทำนายมากขึ้นเท่านั้น[ 5 ]

จากสมมติฐานเกี่ยวกับสกุลเงินและข้อจำกัดกฎการตัดสินใจที่เหมาะสมที่สุดคือการคาดการณ์ของแบบจำลองว่ากลยุทธ์การหาอาหารที่ดีที่สุดของสัตว์ควรเป็นอย่างไร[ 2 ]ตัวอย่างที่เป็นไปได้ของกฎการตัดสินใจที่เหมาะสมที่สุดอาจเป็นจำนวนอาหารที่เหมาะสมที่สุดที่สัตว์ควรนำกลับไปยังรัง หรือขนาดที่เหมาะสมที่สุดของอาหารที่สัตว์ควรกิน รูปที่ 1 แสดงตัวอย่างวิธีการกำหนดกฎการตัดสินใจที่เหมาะสมที่สุดจาก แบบ จำลองกราฟิก[ 7 ]เส้นโค้งแสดงถึงพลังงานที่ได้รับต่อต้นทุน (E) สำหรับการใช้กลยุทธ์การหาอาหาร x พลังงานที่ได้รับต่อต้นทุนคือสกุลเงินที่กำลังได้รับการปรับให้เหมาะสม ข้อจำกัดของระบบกำหนดรูปร่างของเส้นโค้งนี้ กฎการตัดสินใจที่เหมาะสมที่สุด (x*) คือกลยุทธ์ที่สกุลเงิน พลังงานที่ได้รับต่อต้นทุน มีค่ามากที่สุด แบบจำลองการหาอาหารที่เหมาะสมที่สุดอาจดูแตกต่างกันมากและซับซ้อนมาก ขึ้นอยู่กับลักษณะของสกุลเงินและจำนวนข้อจำกัดที่พิจารณา อย่างไรก็ตาม หลักการทั่วไปของสกุลเงิน ข้อจำกัด และกฎการตัดสินใจที่เหมาะสมที่สุดยังคงเหมือนเดิมสำหรับทุกแบบจำลอง
เพื่อทดสอบแบบจำลอง เราสามารถเปรียบเทียบกลยุทธ์ที่คาดการณ์ไว้กับพฤติกรรมการหาอาหารจริงของสัตว์ได้ หากแบบจำลองเข้ากับข้อมูลที่สังเกตได้ดี แสดงว่าสมมติฐานเกี่ยวกับสกุลเงินและข้อจำกัดได้รับการสนับสนุน หากแบบจำลองไม่เข้ากับข้อมูลได้ดี ก็เป็นไปได้ว่าสกุลเงินหรือข้อจำกัดเฉพาะอย่างใดอย่างหนึ่งถูกระบุไม่ถูกต้อง[ 5 ]
ระบบการกินอาหารที่แตกต่างกันและประเภทของสัตว์ผู้ล่า
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging Theory หรือ OFT) สามารถนำไปประยุกต์ใช้ได้อย่างกว้างขวางกับระบบการกินอาหารในอาณาจักรสัตว์ ภายใต้ทฤษฎี OFT สิ่งมีชีวิต ใดๆก็ตามสามารถมองได้ว่าเป็นผู้ล่าที่หาอาหารจากเหยื่อ สิ่งมีชีวิตต่างๆ จัดอยู่ในกลุ่มผู้ล่าหลายประเภท และแต่ละประเภทก็มีกลยุทธ์ การหาอาหารและ การล่าเหยื่อ ที่แตกต่างกัน
- สัตว์นักล่าที่แท้จริงจะโจมตีเหยื่อจำนวนมากตลอดชีวิต พวกมันฆ่าเหยื่อทันทีหรือหลังจากโจมตีไม่นาน พวกมันอาจกินเหยื่อทั้งหมดหรือเพียงบางส่วน สัตว์นักล่าที่แท้จริง ได้แก่ เสือ สิงโตวาฬฉลามและมด[ 8 ]
- สัตว์กินพืชจะกินเหยื่อเพียงบางส่วนเท่านั้น พวกมันทำร้ายเหยื่อ แต่แทบจะไม่ฆ่าเหยื่อเลย สัตว์กินพืชได้แก่ละมั่งวัวและยุง
- ปรสิตเช่นเดียวกับสัตว์กินพืช กินเพียงบางส่วนของเหยื่อ (โฮสต์) แต่ไม่ค่อยกินทั้งตัว พวกมันใช้ชีวิตทั้งหมดหรือส่วนใหญ่ในวงจรชีวิตอาศัยอยู่ใน/บนโฮสต์เพียงตัวเดียว ความสัมพันธ์ที่ใกล้ชิดเช่นนี้พบได้ทั่วไปในพยาธิพยาธิใบไม้ในตับและปรสิตพืช เช่นโรคใบไหม้ของมันฝรั่ง
- ปรสิตส่วนใหญ่เป็นตัวต่อ (อันดับ Hymenoptera ) และแมลงวันบางชนิด (อันดับ Diptera ) พวกมันวางไข่ไว้ภายในตัวอ่อนของสัตว์ขาปล้อง ชนิดอื่น ไข่ จะฟักออกมาและกินโฮสต์จากภายใน ทำให้โฮสต์ตาย ความสัมพันธ์ระหว่างผู้ล่าและโฮสต์ที่ผิดปกตินี้พบได้ประมาณ 10% ของแมลงทั้งหมด [ 9 ]ไวรัสหลายชนิดที่โจมตีสิ่งมีชีวิตเซลล์เดียว (เช่นแบคทีริโอเฟจ ) ก็เป็นปรสิตเช่นกัน พวกมันขยายพันธุ์ภายในโฮสต์ตัวเดียวซึ่งจะถูกฆ่าตายอย่างหลีกเลี่ยงไม่ได้จากความสัมพันธ์นี้
การปรับกลยุทธ์การหาอาหารและการล่าเหยื่อที่แตกต่างกันเหล่านี้ให้เหมาะสมที่สุด สามารถอธิบายได้ด้วยทฤษฎีการหาอาหารที่เหมาะสมที่สุด ในแต่ละกรณี จะมีต้นทุน ผลประโยชน์ และข้อจำกัดที่กำหนดกฎการตัดสินใจที่เหมาะสมที่สุดที่ผู้ล่าควรปฏิบัติตามในที่สุด
แบบจำลองอาหารที่เหมาะสม
ทฤษฎีการหาอาหารที่เหมาะสมที่สุดแบบคลาสสิกแบบหนึ่งคือแบบจำลองอาหารที่เหมาะสมที่สุดซึ่งเรียกอีกอย่างว่าแบบจำลองการเลือกเหยื่อหรือแบบจำลองสถานการณ์ ในแบบจำลองนี้ ผู้ล่าจะพบกับเหยื่อที่แตกต่างกันและตัดสินใจว่าจะกินสิ่งที่ตนมีอยู่หรือค้นหาเหยื่อที่ให้ผลกำไรมากกว่า แบบจำลองนี้ทำนายว่าผู้หาอาหารควรเพิกเฉยต่อเหยื่อที่ให้ผลกำไรต่ำเมื่อมีเหยื่อที่ให้ผลกำไรมากกว่าอยู่มากมาย[ 10 ]
ผลกำไรของเหยื่อขึ้นอยู่กับตัวแปรทางนิเวศวิทยาหลายประการEคือปริมาณพลังงาน (แคลอรี) ที่เหยื่อมอบให้แก่ผู้ล่า เวลาในการจัดการ ( h ) คือระยะเวลาที่ผู้ล่าใช้ในการจัดการอาหาร เริ่มตั้งแต่เวลาที่ผู้ล่าพบเหยื่อจนถึงเวลาที่กินเหยื่อ ผลกำไรของเหยื่อจึงถูกกำหนดเป็นE/hนอกจากนี้ เวลาในการค้นหา ( S ) คือระยะเวลาที่ผู้ล่าใช้ในการค้นหาเหยื่อ ซึ่งขึ้นอยู่กับความอุดมสมบูรณ์ของอาหารและความง่ายในการค้นหา[ 2 ]ในแบบจำลองนี้ สกุลเงินคือปริมาณพลังงานที่ได้รับต่อหน่วยเวลา และข้อจำกัดรวมถึงค่าจริงของE , hและSรวมถึงข้อเท็จจริงที่ว่าเหยื่อจะถูกพบตามลำดับ
รูปแบบการเลือกเหยื่อระหว่างเหยื่อขนาดใหญ่และขนาดเล็ก
โดยใช้ตัวแปรเหล่านี้ แบบจำลองอาหารที่เหมาะสมที่สุดสามารถทำนายได้ว่าผู้ล่าจะเลือกเหยื่อสองประเภทอย่างไร ได้แก่เหยื่อ ขนาดใหญ่ 1ที่มีค่าพลังงานE 1และเวลาในการจัดการh 1 และ เหยื่อขนาดเล็ก2ที่มีค่าพลังงานE 2และเวลาในการจัดการh 2เพื่อให้ได้อัตราการได้รับพลังงานโดยรวมสูงสุด ผู้ล่าต้องพิจารณาถึงความคุ้มค่าของเหยื่อทั้งสองประเภท หากสมมติว่าเหยื่อขนาดใหญ่1มีความคุ้มค่ามากกว่าเหยื่อขนาดเล็ก2ดังนั้นE 1 /h 1 > E 2 /h 2ดังนั้น หากผู้ล่าพบเหยื่อ1มันควรเลือกกินเสมอเพราะมีความคุ้มค่าสูงกว่า มันไม่ควรเสียเวลาไปหาเหยื่อ2อย่างไรก็ตาม หากสัตว์พบเหยื่อ2มันควรปฏิเสธเพื่อหาเหยื่อ1 ที่มีความคุ้มค่ามากกว่า เว้นแต่ว่าเวลาที่ใช้ในการหาเหยื่อ1นั้นนานและมีค่าใช้จ่ายสูงเกินไปจนไม่คุ้มค่า ดังนั้น สัตว์ควรจะกินเหยื่อ2ก็ต่อเมื่อE 2 /h 2 > E 1 /(h 1 +S 1 )โดยที่S 1คือเวลาที่ใช้ในการค้นหาเหยื่อ1เนื่องจากการเลือกกินเหยื่อ1 เป็นสิ่งที่พึงปรารถนาเสมอ การเลือกกินเหยื่อ1จึงไม่ขึ้นอยู่กับความอุดมสมบูรณ์ของเหยื่อ2แต่เนื่องจากความยาวของS 1 (กล่าวคือ ความยากในการหาเหยื่อ 1) ขึ้นอยู่กับความหนาแน่นของเหยื่อ1ดังนั้นการเลือกกินเหยื่อ2 จึงขึ้นอยู่กับความอุดมสมบูรณ์ของเหยื่อ 1 [ 5 ]
อาหารทั่วไปและอาหารเฉพาะทาง
แบบจำลองอาหารที่เหมาะสมที่สุดยังทำนายว่าสัตว์ประเภทต่างๆ ควรเลือกกินอาหารที่แตกต่างกันไปตามความแปรผันของเวลาในการค้นหา แนวคิดนี้เป็นการขยายแบบจำลองการเลือกเหยื่อที่ได้กล่าวถึงข้างต้น สมการE 2 /h 2 > E 1 /(h 1 +S 1 )สามารถจัดเรียงใหม่ได้เป็น: S 1 > [(E 1 h 2 )/E 2 ] – h 1รูปแบบที่จัดเรียงใหม่นี้ให้ค่าเกณฑ์ว่าS 1ต้องยาวเท่าใดเพื่อให้สัตว์เลือกกินทั้งเหยื่อ1และเหยื่อ2 [ 5 ] สัตว์ที่มีS 1ถึงค่าเกณฑ์จะถูกกำหนดให้เป็นสัตว์กินทุกอย่างในธรรมชาติ สัตว์กินทุกอย่างจะรวมเหยื่อหลากหลายชนิดไว้ในอาหารของพวกมัน[ 11 ]ตัวอย่างของสัตว์กินทุกอย่างคือหนู ซึ่งกินเมล็ดพืช ธัญพืช และถั่วหลากหลายชนิด[ 12 ] ในทางตรงกันข้าม ผู้ล่าที่มี S 1ค่อนข้างสั้นยังคงได้เปรียบกว่าหากเลือกกินเฉพาะเหยื่อ1เท่านั้น สัตว์ประเภทนี้ถูกกำหนดให้เป็นผู้เชี่ยวชาญและมีอาหารเฉพาะเจาะจงมากในธรรมชาติ[ 11 ]ตัวอย่างของผู้เชี่ยวชาญคือโคอาลาซึ่งกินเฉพาะใบยูคาลิปตัส เท่านั้น [ 13 ]โดยทั่วไป สัตว์ต่าง ๆ ในสี่กลุ่มหน้าที่ของนักล่าจะแสดงกลยุทธ์ที่หลากหลายตั้งแต่การเป็นผู้เชี่ยวชาญทั่วไปไปจนถึงการเป็นผู้เชี่ยวชาญเฉพาะด้าน นอกจากนี้ เนื่องจากทางเลือกในการกินเหยื่อ 2 ขึ้นอยู่กับความอุดมสมบูรณ์ของเหยื่อ 1 (ดังที่ได้กล่าวไว้ก่อนหน้านี้) หากเหยื่อ 1 ขาดแคลนมากจน S1 ถึงเกณฑ์ สัตว์นั้นควรเปลี่ยนจากการกินเหยื่อ 1 เพียงอย่างเดียวไปเป็นการกินทั้งเหยื่อ 1 และเหยื่อ 2 [ 5 ]กล่าวอีกนัยหนึ่ง หากอาหารในอาหารของผู้เชี่ยวชาญขาดแคลนมาก ผู้เชี่ยวชาญอาจเปลี่ยนไปเป็นผู้เชี่ยวชาญทั่วไปได้ในบางครั้ง
เส้นโค้งการตอบสนองเชิงฟังก์ชัน
ดังที่กล่าวไว้ก่อนหน้านี้ ระยะเวลาที่ใช้ในการค้นหาเหยื่อขึ้นอยู่กับความหนาแน่นของเหยื่อเส้นโค้งการตอบสนองเชิงฟังก์ชันแสดงอัตราการจับเหยื่อเป็นฟังก์ชันของความหนาแน่นของอาหาร และสามารถใช้ร่วมกับทฤษฎีอาหารที่เหมาะสมที่สุดเพื่อทำนายพฤติกรรมการหาอาหารของผู้ล่าได้ มีเส้นโค้งการตอบสนองเชิงฟังก์ชันที่แตกต่างกันสามประเภท[ 14 ]
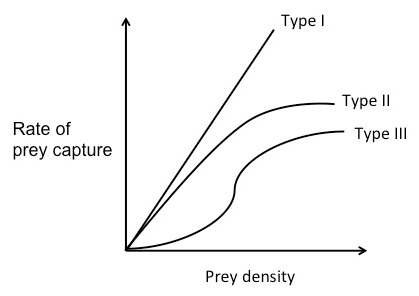
สำหรับ เส้นโค้งการตอบสนองเชิงฟังก์ชัน ประเภท Iอัตราการจับเหยื่อจะเพิ่มขึ้นเชิงเส้นตามความหนาแน่นของอาหาร ที่ความหนาแน่นของเหยื่อต่ำ เวลาในการค้นหาจะนาน เนื่องจากผู้ล่าใช้เวลาส่วนใหญ่ในการค้นหา มันจึงกินเหยื่อทุกตัวที่พบ เมื่อความหนาแน่นของเหยื่อเพิ่มขึ้น ผู้ล่าจะสามารถจับเหยื่อได้เร็วขึ้นเรื่อยๆ จนถึงจุดหนึ่ง อัตราการจับเหยื่อจะสูงมากจนผู้ล่าไม่จำเป็นต้องกินเหยื่อทุกตัวที่พบ หลังจากจุดนี้ ผู้ล่าควรเลือกเฉพาะเหยื่อที่มี E/h สูงที่สุดเท่านั้น[ 15 ]
สำหรับ เส้นโค้งการตอบสนองเชิงฟังก์ชัน ประเภท IIอัตราการจับเหยื่อจะเร่งตัวขึ้นในเชิงลบเมื่อความหนาแน่นของอาหารเพิ่มขึ้น[ 14 ]ทั้งนี้เนื่องจากถือว่าผู้ล่าถูกจำกัดด้วยความสามารถในการแปรรูปอาหาร กล่าวอีกนัยหนึ่งคือ เมื่อความหนาแน่นของอาหารเพิ่มขึ้น เวลาในการจัดการก็จะเพิ่มขึ้น ในช่วงเริ่มต้นของเส้นโค้ง อัตราการจับเหยื่อจะเพิ่มขึ้นเกือบเป็นเส้นตรงตามความหนาแน่นของเหยื่อ และแทบไม่มีเวลาในการจัดการเลย เมื่อความหนาแน่นของเหยื่อเพิ่มขึ้น ผู้ล่าจะใช้เวลาน้อยลงเรื่อยๆ ในการค้นหาเหยื่อและใช้เวลามากขึ้นเรื่อยๆ ในการจัดการเหยื่อ อัตราการจับเหยื่อจึงเพิ่มขึ้นน้อยลงเรื่อยๆ จนกระทั่งถึงจุดคงที่ในที่สุด จำนวนเหยื่อที่มากจะ "ท่วม" ผู้ล่าโดยพื้นฐาน[ 15 ]
เส้นโค้งการตอบสนองเชิงฟังก์ชัน ประเภทIIIเป็นเส้นโค้งซิกมอยด์อัตราการจับเหยื่อจะเพิ่มขึ้นในตอนแรกตามความหนาแน่นของเหยื่อในอัตราเร่งที่เป็นบวก แต่จากนั้นที่ความหนาแน่นสูงจะเปลี่ยนเป็นรูปแบบเร่งที่เป็นลบ คล้ายกับเส้นโค้งประเภท II [ 14 ]ที่ความหนาแน่นของเหยื่อสูง (ด้านบนของเส้นโค้ง) เหยื่อแต่ละตัวจะถูกจับได้เกือบจะทันที ผู้ล่าสามารถเลือกได้และไม่กินทุกตัวที่พบ ดังนั้น สมมติว่ามีเหยื่อสองประเภทที่มีความสามารถในการทำกำไรต่างกันและมีจำนวนมากทั้งคู่ ผู้ล่าจะเลือกเหยื่อที่มีE/h สูงกว่า อย่างไรก็ตาม ที่ความหนาแน่นของเหยื่อต่ำ (ด้านล่างของเส้นโค้ง) อัตราการจับเหยื่อจะเพิ่มขึ้นเร็วกว่าเชิงเส้น ซึ่งหมายความว่าเมื่อผู้ล่ากินอาหารและเหยื่อประเภทที่มีE/h สูงกว่า มีจำนวนน้อยลง ผู้ล่าจะเริ่มเปลี่ยนความชอบไปเป็นเหยื่อประเภทที่มีE/h ต่ำกว่า เนื่องจากประเภทนั้นจะมีจำนวนมากกว่า ปรากฏการณ์นี้เรียกว่า การ เปลี่ยนเหยื่อ[ 14 ]
ปฏิสัมพันธ์ระหว่างผู้ล่าและเหยื่อ
วิวัฒนาการร่วมกันระหว่างผู้ล่าและเหยื่อมักทำให้ผู้ล่าไม่สามารถกินเหยื่อบางชนิดได้ เนื่องจากกลไกป้องกันตัวจากผู้ล่าหลายอย่างทำให้ต้องใช้เวลานานในการจัดการ[ 16 ]ตัวอย่างเช่นขนของเม่นความน่ากินและการย่อยได้ของกบลูกดอกพิษการพรางตัวและพฤติกรรมการหลีกเลี่ยงผู้ล่าอื่นๆ นอกจากนี้ เนื่องจากสารพิษอาจมีอยู่ในเหยื่อหลายชนิด ผู้ล่าจึงกินอาหารที่หลากหลายเพื่อป้องกันไม่ให้สารพิษชนิดใดชนิดหนึ่งมีปริมาณถึงระดับที่เป็นอันตราย ดังนั้น แนวทางที่เน้นเฉพาะการรับพลังงานอาจไม่สามารถอธิบายพฤติกรรมการหาอาหารของสัตว์ในสถานการณ์เหล่านี้ได้อย่างครบถ้วน
ทฤษฎีมูลค่าส่วนเพิ่มและการหาอาหารที่เหมาะสมที่สุด

ทฤษฎีบทมูลค่าส่วนเพิ่มเป็นแบบจำลองความเหมาะสมประเภทหนึ่งที่มักนำมาใช้กับการหาอาหารที่เหมาะสมที่สุด ทฤษฎีบทนี้ใช้เพื่ออธิบายสถานการณ์ที่สิ่งมีชีวิตที่กำลังค้นหาอาหารในพื้นที่หนึ่งต้องตัดสินใจว่าเมื่อใดจึงจะคุ้มค่าทางเศรษฐกิจที่จะออกจากพื้นที่นั้น ในขณะที่สัตว์อยู่ในพื้นที่นั้น มันจะประสบกับกฎของผลตอบแทนที่ลดลงซึ่งหมายความว่ามันจะหาเหยื่อได้ยากขึ้นเรื่อยๆ เมื่อเวลาผ่านไป นี่อาจเป็นเพราะเหยื่อกำลังลดน้อยลง เหยื่อเริ่มหลบหลีกและจับได้ยากขึ้น หรือผู้ล่าเริ่มเดินตัดเส้นทางของตัวเองมากขึ้นในขณะที่ค้นหา[ 5 ]กฎของผลตอบแทนที่ลดลงนี้สามารถแสดงได้เป็นเส้นโค้งของพลังงานที่ได้รับต่อเวลาที่ใช้ในพื้นที่ (รูปที่ 3) เส้นโค้งเริ่มต้นด้วยความชันสูงและค่อยๆ ราบเรียบลงเมื่อหาเหยื่อได้ยากขึ้น ต้นทุนที่สำคัญอีกประการหนึ่งที่ต้องพิจารณาคือเวลาในการเดินทางระหว่างพื้นที่ต่างๆ และแหล่งทำรัง สัตว์จะสูญเสียเวลาในการหาอาหารในขณะที่เดินทางและใช้พลังงานในการเคลื่อนที่[ 2 ]
ในแบบจำลองนี้ สกุลเงินที่ถูกปรับให้เหมาะสมที่สุดมักจะเป็นพลังงานสุทธิที่ได้รับต่อหน่วยเวลา ข้อจำกัดคือเวลาในการเดินทางและรูปร่างของเส้นโค้งผลตอบแทนที่ลดลง ในทางกราฟิก สกุลเงิน (พลังงานสุทธิที่ได้รับต่อหน่วยเวลา) จะกำหนดโดยความชันของเส้นทแยงมุมที่เริ่มต้นที่จุดเริ่มต้นของเวลาในการเดินทางและตัดกับเส้นโค้งผลตอบแทนที่ลดลง (รูปที่ 3) เพื่อให้ได้สกุลเงินสูงสุด เราต้องการเส้นที่มีความชันมากที่สุดที่ยังคงสัมผัสกับเส้นโค้ง (เส้นสัมผัส) จุดที่เส้นนี้สัมผัสกับเส้นโค้งจะให้กฎการตัดสินใจที่เหมาะสมที่สุดเกี่ยวกับปริมาณเวลาที่สัตว์ควรใช้ในพื้นที่ก่อนที่จะจากไป
ตัวอย่างของแบบจำลองการหาอาหารที่เหมาะสมที่สุดในสัตว์

การหาอาหารที่เหมาะสมที่สุดของนกนางแอ่นทะเล
การกิน หอยแมลงภู่ของนกนางแอ่นเป็นตัวอย่างหนึ่งของการใช้แบบจำลองอาหารที่เหมาะสมที่สุด นกนางแอ่นหากินหอยแมลงภู่และแกะเปลือกด้วยปากของมัน ข้อจำกัดของนกเหล่านี้คือลักษณะของขนาดหอยแมลงภู่ที่แตกต่างกัน ในขณะที่หอยแมลงภู่ขนาดใหญ่ให้พลังงานมากกว่าหอยแมลงภู่ขนาดเล็ก แต่หอยแมลงภู่ขนาดใหญ่แกะยากกว่าเนื่องจากเปลือกหนากว่า ซึ่งหมายความว่าในขณะที่หอยแมลงภู่ขนาดใหญ่มีปริมาณพลังงาน ( E ) สูงกว่า แต่ก็ต้องใช้เวลาในการจัดการ ( h ) นานกว่า ความสามารถใน การทำกำไรของหอยแมลงภู่แต่ละตัวคำนวณได้จากE/hนกนางแอ่นต้องตัดสินใจว่าหอยแมลงภู่ขนาดใดจะให้สารอาหารเพียงพอที่จะคุ้มค่ากับต้นทุนและพลังงานที่ต้องใช้ในการแกะ[ 2 ]ในการศึกษาของพวกเขา Meire และ Ervynck พยายามสร้างแบบจำลองการตัดสินใจนี้โดยการสร้างกราฟความสามารถในการทำกำไรสัมพัทธ์ของหอยแมลงภู่ขนาดต่างๆ พวกเขาได้เส้นโค้งรูปทรงระฆัง ซึ่งบ่งชี้ว่าหอยแมลงภู่ขนาดปานกลางมีกำไรมากที่สุด อย่างไรก็ตาม พวกเขาพบว่าหากนกนางแอ่นทะเลปฏิเสธหอยแมลงภู่ขนาดเล็กมากเกินไป เวลาที่ใช้ในการค้นหาหอยแมลงภู่ที่เหมาะสมตัวต่อไปจะเพิ่มขึ้นอย่างมาก การสังเกตนี้ทำให้เส้นโค้งระฆังของพวกเขาเลื่อนไปทางขวา (รูปที่ 4) อย่างไรก็ตาม ในขณะที่แบบจำลองนี้ทำนายว่านกนางแอ่นทะเลควรชอบหอยแมลงภู่ขนาด 50–55 มม. ข้อมูลที่สังเกตได้แสดงให้เห็นว่านกนางแอ่นทะเลชอบหอยแมลงภู่ขนาด 30–45 มม. Meire และ Ervynk จึงตระหนักว่าความชอบขนาดของหอยแมลงภู่ไม่ได้ขึ้นอยู่กับผลกำไรของเหยื่อเท่านั้น แต่ยังขึ้นอยู่กับความหนาแน่นของเหยื่อด้วย หลังจากที่ได้พิจารณาเรื่องนี้แล้ว พวกเขาก็พบว่ามีความสอดคล้องกันที่ดีระหว่างการทำนายของแบบจำลองกับข้อมูลที่สังเกตได้[ 17 ]
การหาอาหารที่เหมาะสมที่สุดของนกสตาร์ลิง
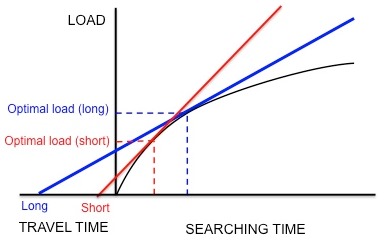
พฤติกรรมการหาอาหารของนกสตาร์ลิงยุโรป ( Sturnus vulgaris ) เป็นตัวอย่างหนึ่งของการใช้ทฤษฎีคุณค่าส่วนเพิ่ม (marginal value theorem) ในการจำลองการหาอาหารที่เหมาะสมที่สุด นกสตาร์ลิงจะออกจากรังและเดินทางไปยังแหล่งอาหารเพื่อค้นหาตัวอ่อนของแมลงเต่าทอง(leatherjackets)เพื่อนำกลับมาให้ลูกนก นกสตาร์ลิงต้องกำหนดจำนวนเหยื่อที่เหมาะสมที่สุดที่จะนำกลับมาในแต่ละครั้ง (กล่าวคือ ขนาดของน้ำหนักบรรทุกที่เหมาะสมที่สุด) ในขณะที่นกสตาร์ลิงหาอาหารอยู่ในแหล่งอาหาร พวกมันจะประสบกับผลตอบแทนที่ลดลง: นกสตาร์ลิงสามารถคาบตัวอ่อนของแมลงเต่าทองไว้ในปากได้เพียงจำนวนจำกัด ดังนั้นความเร็วในการที่พ่อแม่นกจะคาบตัวอ่อนกลับมาจึงลดลงตามจำนวนตัวอ่อนที่มันมีอยู่ในปากแล้ว ดังนั้นข้อจำกัดจึงเป็นรูปร่างของเส้นโค้งผลตอบแทนที่ลดลงและเวลาในการเดินทาง (เวลาที่ใช้ในการเดินทางไปกลับจากรังไปยังแหล่งอาหารและกลับมา) นอกจากนี้ สกุลเงินที่ตั้งสมมติฐานไว้คือพลังงานสุทธิที่ได้รับต่อหน่วยเวลา[ 5 ]เมื่อใช้สกุลเงินนี้และข้อจำกัดต่างๆ จะสามารถคาดการณ์ภาระที่เหมาะสมที่สุดได้โดยการลากเส้นสัมผัสกับเส้นโค้งผลตอบแทนที่ลดลง ดังที่ได้กล่าวไว้ก่อนหน้านี้ (รูปที่ 3)
Kacelnik และคณะต้องการตรวจสอบว่าสายพันธุ์นี้เพิ่มประสิทธิภาพการได้รับพลังงานสุทธิต่อหน่วยเวลาตามสมมติฐานหรือไม่[ 18 ]พวกเขาออกแบบการทดลองโดยฝึกนกสตาร์ลิงให้เก็บหนอนแมลงจากเครื่องให้อาหารเทียมในระยะทางที่แตกต่างกันจากรัง นักวิจัยสร้างเส้นโค้งผลตอบแทนที่ลดลงคงที่สำหรับนกโดยการปล่อยหนอนแมลงในช่วงเวลาที่ยาวขึ้นเรื่อยๆ นกยังคงเก็บหนอนแมลงต่อไปเมื่อมีการนำเสนอ จนกระทั่งถึงปริมาณที่ "เหมาะสม" แล้วบินกลับบ้าน ดังที่แสดงในรูปที่ 5 หากนกสตาร์ลิงเพิ่มประสิทธิภาพการได้รับพลังงานสุทธิต่อหน่วยเวลา เวลาเดินทางสั้นๆ จะทำนายปริมาณที่เหมาะสมที่น้อย และเวลาเดินทางที่ยาวจะทำนายปริมาณที่เหมาะสมที่มากขึ้น สอดคล้องกับการคาดการณ์เหล่านี้ Kacelnik พบว่ายิ่งระยะทางระหว่างรังและเครื่องให้อาหารเทียมยาวขึ้นเท่าใด ขนาดของปริมาณก็จะยิ่งใหญ่ขึ้นเท่านั้น นอกจากนี้ ขนาดของปริมาณที่สังเกตได้ยังสอดคล้องกับการคาดการณ์ของแบบจำลองในเชิงปริมาณอย่างใกล้ชิดอีกด้วย แบบจำลองอื่นๆ ที่ใช้สกุลเงินต่างกัน เช่น พลังงานที่ได้รับต่อพลังงานที่ใช้ไป (เช่นประสิทธิภาพการใช้พลังงาน ) ไม่สามารถทำนายขนาดโหลดที่สังเกตได้อย่างแม่นยำ ดังนั้น Kacelnik จึงสรุปว่านกสตาร์ลิงจะเพิ่มพลังงานสุทธิที่ได้รับต่อหน่วยเวลาให้สูงสุด ข้อสรุปนี้ไม่ได้รับการพิสูจน์ว่าผิดในการทดลองในภายหลัง[ 19 ] [ 20 ]
การหาอาหารที่เหมาะสมที่สุดในผึ้ง
ผึ้งงานเป็นอีกตัวอย่างหนึ่งของการใช้ทฤษฎีคุณค่าส่วนเพิ่มในการจำลองพฤติกรรมการหาอาหารที่เหมาะสมที่สุด ผึ้งจะหาอาหารจากดอกไม้ดอกหนึ่งไปยังอีกดอกหนึ่งเพื่อเก็บน้ำหวานกลับไปที่รัง แม้ว่าสถานการณ์นี้จะคล้ายกับนกสตาร์ลิง แต่ข้อจำกัดและมูลค่าที่แท้จริงนั้นแตกต่างกันสำหรับผึ้ง
ผึ้งจะไม่ประสบกับผลตอบแทนที่ลดลงเนื่องจากการหมดไปของน้ำหวานหรือลักษณะอื่นใดของดอกไม้เอง ปริมาณน้ำหวานทั้งหมดที่หาได้จะเพิ่มขึ้นเป็นเส้นตรงตามเวลาที่ใช้ในบริเวณนั้น อย่างไรก็ตาม น้ำหนักของน้ำหวานจะเพิ่มต้นทุนอย่างมากต่อการบินของผึ้งระหว่างดอกไม้และการเดินทางกลับรัง Wolf และ Schmid-Hempel ได้แสดงให้เห็นโดยการทดลองวางน้ำหนักที่แตกต่างกันไว้บนหลังของผึ้งว่า ต้นทุนของน้ำหวานที่มีน้ำหนักมากนั้นสูงมากจนทำให้อายุขัยของผึ้งสั้นลง[ 21 ]ยิ่งอายุขัยของผึ้งงานสั้นลงเท่าใด เวลาโดยรวมที่มันสามารถช่วยรังได้ก็จะยิ่งน้อยลงเท่านั้น ดังนั้นจึงมีเส้นโค้งของผลตอบแทนที่ลดลงสำหรับผลผลิตสุทธิของพลังงานที่รังได้รับเมื่อผึ้งเก็บน้ำหวานมากขึ้นในระหว่างการเดินทางแต่ละครั้ง[ 5 ]
ต้นทุนของน้ำหวานที่มีน้ำหนักมากยังส่งผลกระทบต่อสกุลเงินที่ผึ้งใช้ด้วย ต่างจากนกสตาร์ลิงในตัวอย่างก่อนหน้านี้ ผึ้งจะเพิ่มประสิทธิภาพการใช้พลังงาน (พลังงานที่ได้รับต่อพลังงานที่ใช้ไป) ให้สูงสุด มากกว่าอัตราสุทธิของการได้รับพลังงาน (พลังงานสุทธิที่ได้รับต่อเวลา) เนื่องจากภาระที่เหมาะสมที่คาดการณ์โดยการเพิ่มอัตราสุทธิของการได้รับพลังงานให้สูงสุดนั้นหนักเกินไปสำหรับผึ้งและทำให้อายุขัยของพวกมันสั้นลง ลดผลผลิตโดยรวมของรังลง ดังที่ได้อธิบายไว้ก่อนหน้านี้ โดยการเพิ่มประสิทธิภาพการใช้พลังงานให้สูงสุด ผึ้งสามารถหลีกเลี่ยงการใช้พลังงานมากเกินไปต่อการเดินทางแต่ละครั้งและสามารถมีชีวิตอยู่ได้นานพอที่จะเพิ่มผลผลิตตลอดช่วงชีวิตของพวกมันให้สูงสุดสำหรับรัง[ 5 ]ในเอกสารอื่น Schmid-Hempel แสดงให้เห็นว่าความสัมพันธ์ที่สังเกตได้ระหว่างขนาดของภาระและเวลาบินมีความสัมพันธ์ที่ดีกับการคาดการณ์โดยอิงจากการเพิ่มประสิทธิภาพการใช้พลังงานให้สูงสุด แต่มีความสัมพันธ์ที่แย่มากกับการคาดการณ์โดยอิงจากการเพิ่มอัตราสุทธิของการได้รับพลังงานให้สูงสุด[ 22 ]
การหาอาหารที่เหมาะสมที่สุดในปลาวงศ์ Centrarchidae
ลักษณะการเลือกเหยื่อของปลาสองชนิดในวงศ์ Centrarchidae ( ปลาแครปปี้ขาวและปลาบลูจิลล์ ) ได้ถูกนำเสนอเป็นแบบจำลองที่รวมกลยุทธ์การหาอาหารที่เหมาะสมที่สุดโดย Manatunge และ Asaeda [ 23 ]ขอบเขตการมองเห็นของปลาที่กำลังหาอาหารซึ่งแสดงโดยระยะการตอบสนองได้รับการวิเคราะห์อย่างละเอียดเพื่อประเมินจำนวนการพบเหยื่อต่อการค้นหาแต่ละครั้ง ระยะการตอบสนองที่คาดการณ์ไว้ถูกนำมาเปรียบเทียบกับข้อมูลจากการทดลอง ต้นทุนพลังงานที่เกี่ยวข้องกับพฤติกรรมการหาอาหารของปลาถูกคำนวณโดยอิงจากลำดับเหตุการณ์ที่เกิดขึ้นสำหรับเหยื่อแต่ละตัวที่ถูกกิน การเปรียบเทียบความอุดมสมบูรณ์สัมพัทธ์ของชนิดเหยื่อและขนาดในกระเพาะกับสภาพแวดล้อมของทะเลสาบแสดงให้เห็นว่าทั้งปลาแครปปี้ขาวและปลาบลูจิลล์ (ความยาว < 100 มม.) เลือกเหยื่อโดยใช้กลยุทธ์การเพิ่มประสิทธิภาพพลังงานอย่างมาก ในกรณีส่วนใหญ่ ปลาจะเลือกเฉพาะDaphnia ขนาดใหญ่ โดยไม่สนใจเหยื่อที่หลบหลีกได้ ( Cyclops , diaptomids ) และcladocera ขนาดเล็ก การเลือกกินเหยื่อนี้เป็นผลมาจากการที่ปลาหลีกเลี่ยงเหยื่อที่มีความสามารถในการหลบหนีสูง แม้ว่าเหยื่อเหล่านั้นจะมีพลังงานสูงก็ตาม และได้นำความสามารถนี้มาปรับใช้ให้เกิดการเลือกกินเหยื่ออย่างเหมาะสมผ่านอัตราความสำเร็จในการจับเหยื่อ การพิจารณาเรื่องพลังงานและระบบการมองเห็นนอกเหนือจากความสามารถของปลาในการจับเหยื่อแล้ว ยังเป็นปัจจัยหลักที่กำหนดการเลือกกินเหยื่อของปลาบลูจิลและปลาไวท์แครปปี้ขนาดใหญ่ที่ยังอยู่ในช่วงกินแพลงก์ตอน
คำวิจารณ์และข้อจำกัด
แม้ว่างานวิจัยหลายชิ้น เช่น งานวิจัยที่ยกตัวอย่างข้างต้น จะให้การสนับสนุนเชิงปริมาณแก่ทฤษฎีการหาอาหารอย่างเหมาะสมและแสดงให้เห็นถึงประโยชน์ของทฤษฎีนี้ แต่แบบจำลองดังกล่าวก็ได้รับการวิพากษ์วิจารณ์เกี่ยวกับความถูกต้องและข้อจำกัดของมัน
ประการแรก ทฤษฎีการหาอาหารที่เหมาะสมที่สุดอาศัยสมมติฐานที่ว่าการคัดเลือกโดยธรรมชาติจะปรับกลยุทธ์การหาอาหารของสิ่งมีชีวิตให้เหมาะสมที่สุด อย่างไรก็ตาม การคัดเลือกโดยธรรมชาติไม่ใช่พลังที่ทรงอำนาจทุกอย่างที่สร้างการออกแบบที่สมบูรณ์แบบ แต่เป็นกระบวนการคัดเลือกแบบพาสซีฟสำหรับลักษณะทางพันธุกรรมที่เพิ่มความสำเร็จในการสืบพันธุ์ ของสิ่งมีชีวิต เนื่องจากพันธุศาสตร์เกี่ยวข้องกับปฏิสัมพันธ์ระหว่างตำแหน่งการรวมตัวใหม่และความซับซ้อนอื่นๆ จึงไม่มีการรับประกันว่าการคัดเลือกโดยธรรมชาติจะสามารถปรับพารามิเตอร์พฤติกรรมเฉพาะให้เหมาะสมที่สุดได้[ 24 ]
นอกจากนี้ OFT ยังถือว่าพฤติกรรมการหาอาหารสามารถถูกกำหนดรูปแบบได้อย่างอิสระโดยการคัดเลือกโดยธรรมชาติ เนื่องจากพฤติกรรมเหล่านี้เป็นอิสระจากกิจกรรมอื่นๆ ของสิ่งมีชีวิต[ 24 ]อย่างไรก็ตาม เนื่องจากสิ่งมีชีวิตเป็นระบบที่บูรณาการ ไม่ใช่กลุ่มชิ้นส่วนเชิงกล จึงไม่ใช่เช่นนั้นเสมอไป ตัวอย่างเช่น ความจำเป็นในการหลีกเลี่ยงผู้ล่าอาจจำกัดให้ผู้หาอาหารกินอาหารน้อยกว่าอัตราที่เหมาะสม ดังนั้น พฤติกรรมการหาอาหารของสิ่งมีชีวิตอาจไม่ได้รับการปรับให้เหมาะสมตามที่ OFT คาดการณ์ไว้ เนื่องจากไม่ได้เป็นอิสระจากพฤติกรรมอื่นๆ[ 25 ]
ข้อจำกัดอีกประการหนึ่งของ OFT คือขาดความแม่นยำในทางปฏิบัติ ในทางทฤษฎี แบบจำลองการหาอาหารที่เหมาะสมที่สุดจะให้การคาดการณ์เชิงปริมาณที่เฉพาะเจาะจงแก่ผู้วิจัยเกี่ยวกับกฎการตัดสินใจที่เหมาะสมที่สุดของผู้ล่าโดยอิงจากสมมติฐานเกี่ยวกับสกุลเงินและข้อจำกัดของระบบ อย่างไรก็ตาม ในความเป็นจริง เป็นเรื่องยากที่จะกำหนดแนวคิดพื้นฐาน เช่น ประเภทของเหยื่อ อัตราการพบเจอ หรือแม้แต่พื้นที่ตามที่ผู้หาอาหารรับรู้[ 24 ]ดังนั้น แม้ว่าตัวแปรของ OFT อาจดูสอดคล้องกันในทางทฤษฎี แต่ในทางปฏิบัติ ตัวแปรเหล่านั้นอาจเป็นไปโดยพลการและยากต่อการวัด
นอกจากนี้ แม้ว่าหลักการของ OFT คือการเพิ่มความเหมาะสมของสิ่งมีชีวิตให้สูงสุด แต่การศึกษาหลายชิ้นแสดงให้เห็นเพียงความสัมพันธ์ระหว่างพฤติกรรมการหาอาหารที่สังเกตได้และที่คาดการณ์ไว้ และหยุดอยู่เพียงแค่การทดสอบว่าพฤติกรรมของสัตว์จะเพิ่มความเหมาะสมในการสืบพันธุ์จริงหรือไม่ เป็นไปได้ว่าในบางกรณี อาจไม่มีความสัมพันธ์ระหว่างผลตอบแทนจากการหาอาหารและความสำเร็จในการสืบพันธุ์เลย[ 24 ]หากไม่คำนึงถึงความเป็นไปได้นี้ การศึกษาหลายชิ้นที่ใช้ OFT จึงยังคงไม่สมบูรณ์และไม่สามารถกล่าวถึงและทดสอบประเด็นหลักของทฤษฎีได้
หนึ่งในคำวิจารณ์ที่สำคัญที่สุดของ OFT คืออาจไม่สามารถทดสอบได้อย่างแท้จริง ปัญหานี้เกิดขึ้นเมื่อใดก็ตามที่มีความคลาดเคลื่อนระหว่างการคาดการณ์ของแบบจำลองกับการสังเกตจริง เป็นเรื่องยากที่จะบอกได้ว่าแบบจำลองนั้นผิดพลาดโดยพื้นฐานหรือไม่ หรือว่าตัวแปรเฉพาะเจาะจงนั้นถูกระบุอย่างไม่ถูกต้องหรือถูกละเลยไป เนื่องจากเป็นไปได้ที่จะเพิ่มการปรับเปลี่ยนที่สมเหตุสมผลอย่างไม่สิ้นสุดให้กับแบบจำลอง แบบจำลองของความเหมาะสมที่สุดจึงอาจไม่เคยถูกปฏิเสธ[ 24 ]สิ่งนี้สร้างปัญหาให้นักวิจัยสร้างแบบจำลองของตนให้เข้ากับการสังเกต แทนที่จะทดสอบสมมติฐานเกี่ยวกับพฤติกรรมการหาอาหารของสัตว์อย่างเข้มงวด
ในทางโบราณคดี
ทฤษฎีการหาอาหารที่เหมาะสมที่สุดถูกนำมาใช้เพื่อทำนายพฤติกรรมของสัตว์เมื่อค้นหาอาหาร แต่ยังสามารถใช้กับมนุษย์ (โดยเฉพาะนักล่าและผู้เก็บเกี่ยว) ได้อีกด้วย อาหารให้พลังงานแต่ก็ต้องใช้พลังงานในการหามาเช่นกัน กลยุทธ์การหาอาหารต้องให้ประโยชน์สูงสุดด้วยต้นทุนที่ต่ำที่สุด ซึ่งเป็นความสมดุลระหว่างคุณค่าทางโภชนาการและพลังงานที่ต้องการ พลังงานเป็นหน่วยวัดของทฤษฎีการหาอาหารที่เหมาะสมที่สุด เพราะเป็นองค์ประกอบสำคัญสำหรับสิ่งมีชีวิต แต่ก็เป็นจุดอ่อนของทฤษฎีการหาอาหารที่เหมาะสมที่สุดในด้านโบราณคดีเช่นกัน[ 26 ]ทฤษฎีการหาอาหารที่เหมาะสมที่สุดถือว่าพฤติกรรมได้รับอิทธิพลจากองค์ประกอบทางพันธุกรรมในระดับหนึ่ง อย่างไรก็ตาม การยอมรับสิ่งนี้อาจเป็นเรื่องยากในกรณีของสัตว์ที่ซับซ้อนซึ่งมีความยืดหยุ่นทางพฤติกรรมสูง[ 27 ]
พฤติกรรมของมนุษย์ไม่สามารถคาดเดาได้เสมอไปเมื่อใช้สมมติฐานของทฤษฎีการหาอาหารที่เหมาะสมที่สุด – นักล่าและผู้เก็บเกี่ยวอาจเลือกเกมที่ไม่เป็นประโยชน์ต่อพลังงาน แต่เป็นประโยชน์ต่อความต้องการอื่นๆ เพื่อจุดประสงค์ทางพิธีกรรมหรือการจัดงานเลี้ยง[ 26 ]เป้าหมายและค่านิยมของมนุษย์มักเปลี่ยนแปลงได้เนื่องจากความยืดหยุ่นทางพฤติกรรม การเปลี่ยนแปลงเหล่านี้อาจเกิดขึ้นในช่วงระยะเวลานาน ในช่วงฤดูกาล หรือระหว่างการล่าสัตว์ ดังนั้นทฤษฎีการหาอาหารที่เหมาะสมที่สุดจึงต้องมีความซับซ้อนมากขึ้นและนำเสนอเป้าหมายและข้อจำกัดเพิ่มเติมเพื่อให้สอดคล้องกับกระบวนการตัดสินใจที่ซับซ้อนซึ่งมนุษย์ใช้[ 28 ]นอกจากนี้ กระบวนการตัดสินใจยังอาจได้รับอิทธิพลจากประสบการณ์ระยะยาวและระยะสั้น และประสบการณ์เหล่านี้ก็สามารถส่งผลต่อผู้อื่นได้อีกด้วย[ 28 ]
แต่ทฤษฎีการหาอาหารที่เหมาะสมที่สุดได้ช่วยให้โบราณคดีสร้างการตีความรูปแบบใหม่ในบันทึกทางโบราณคดีและคิดเกี่ยวกับพฤติกรรมของมนุษย์ในรายละเอียดมากขึ้น[ 26 ]ทฤษฎีการหาอาหารที่เหมาะสมที่สุดยังช่วยให้โบราณคดีเข้าใจต้นทุนและผลประโยชน์ของทรัพยากรต่างๆ ได้ดียิ่งขึ้น[ 26 ]
อ่านเพิ่มเติม
- ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging Theory) โดย แบร์รี ซิเนอร์โว (1997) วิชา "นิเวศวิทยาเชิงพฤติกรรม 2013" ภาควิชานิเวศวิทยาและชีววิทยาวิวัฒนาการ มหาวิทยาลัยแคลิฟอร์เนียเซาท์แคโรไลนา – ส่วนนี้ของวิชาที่ UCSC จะพิจารณาถึง OFT และ 'สมมติฐานการปรับตัว' ('การลองผิดลองถูกแบบมีทิศทาง สัญชาตญาณ') พร้อมกับหัวข้อเพิ่มเติม เช่น "ขนาดเหยื่อ" "ระยะเวลาการอาศัยอยู่ในแหล่งอาหาร" "คุณภาพของแหล่งอาหารและคู่แข่ง" "กลยุทธ์การค้นหา" "พฤติกรรมหลีกเลี่ยงความเสี่ยง" และวิธีการหาอาหารภายใต้ "ข้อจำกัดของอาหาร" ดูเพิ่มเติม: ขึ้นไปหนึ่งระดับสำหรับส่วนหลักของวิชา ซึ่งมีไฟล์ PDF ให้ดาวน์โหลด (เนื่องจากรูปภาพในหน้านั้นดูเหมือนจะเสียอยู่ในขณะนี้) ไฟล์ PDF จากลิงก์ด้านบนมีความยาว 26 หน้า (พร้อมรูปภาพ)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ทฤษฎีการหาอาหารที่เหมาะสมที่สุด
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging Theory หรือ OFT ) เป็น แบบจำลอง ทางนิเวศวิทยาเชิงพฤติกรรม ที่ช่วยทำนายพฤติกรรมของสัตว์เมื่อค้นหาอาหาร...
การสร้างแบบจำลองการหาอาหารที่เหมาะสมที่สุด
แบบจำลองการหาอาหารที่เหมาะสมจะสร้างการคาดการณ์เชิงปริมาณว่าสัตว์จะเพิ่มความเหมาะสมสูงสุดได้อย่างไรในขณะที่พวกมันหาอาหาร กระบวนการสร้างแบบจำลองเกี่ยวข้องกับการระบุสกุลเงิน ข้อจำกัด และกฎการตัดสินใจที่เหมาะสมสำหรับผู้หาอาหาร [ 2 ] [ 4 ]
ระบบการกินอาหารที่แตกต่างกันและประเภทของสัตว์ผู้ล่า
ทฤษฎีการหาอาหารที่เหมาะสมที่สุด (Optimal Foraging Theory หรือ OFT) สามารถนำไปประยุกต์ใช้ได้อย่างกว้างขวางกับระบบการกินอาหารในอาณาจักรสัตว์ ภายใต้ทฤษฎี OFT สิ่งมีชีวิต ใดๆ ก็ตามสามารถมองได้ว่าเป็นผู้ล่าที่หาอาหารจากเหยื่อ สิ่งมีชีวิตต่างๆ...
แบบจำลองอาหารที่เหมาะสม
ทฤษฎีการหาอาหารที่เหมาะสมที่สุดแบบคลาสสิกแบบหนึ่งคือ แบบจำลองอาหารที่เหมาะสมที่สุด ซึ่งเรียกอีกอย่างว่าแบบจำลองการเลือกเหยื่อหรือแบบจำลองสถานการณ์ ในแบบจำลองนี้...