อ่าน 8 นาที
การถ่ายทอดยีนในแนวนอนในวิวัฒนาการ
การถ่ายทอดยีนในแนวนอน ( HGT ) หมายถึงการถ่ายทอดยีนระหว่างกิ่งก้านที่อยู่ห่างไกลกันบนแผนภูมิวิวัฒนาการในทางวิวัฒนาการ...
การถ่ายทอดยีนในแนวนอนในวิวัฒนาการ
การถ่ายทอดยีนในแนวนอน ( HGT ) หมายถึงการถ่ายทอดยีนระหว่างกิ่งก้านที่อยู่ห่างไกลกันบนแผนภูมิวิวัฒนาการในทางวิวัฒนาการ การถ่ายทอดยีนในแนวนอนสามารถทำให้ข้อมูลที่จำเป็นในการสร้างแผนภูมิวิวัฒนาการของสิ่งมี ชีวิต และความสัมพันธ์ระหว่างสิ่งมีชีวิตต่างๆ เกิดความสับสนได้
นอกจากนี้ HGT ยังช่วยให้นักวิทยาศาสตร์สามารถสร้างและกำหนดอายุของแผนภูมิวิวัฒนาการของสิ่งมีชีวิตได้ เนื่องจากสามารถใช้การถ่ายทอดยีนเป็นเครื่องหมายทางวิวัฒนาการ หรือเป็นหลักฐานแสดงถึงความร่วมสมัยของสิ่งมีชีวิตผู้ให้และผู้รับ และเป็นร่องรอยของความหลากหลายทางชีวภาพที่สูญพันธุ์ไปแล้ว
การถ่ายทอดยีนแนวนอน (HGT) เกิดขึ้นน้อยมาก – ในระดับสิ่งมีชีวิตแต่ละตัว โอกาสที่จะเกิดเหตุการณ์เช่นนี้ขึ้นนั้นแทบเป็นไปไม่ได้เลย อย่างไรก็ตาม ในระดับที่ใหญ่กว่านั้น คือประวัติศาสตร์วิวัฒนาการ เหตุการณ์เหล่านี้เกิดขึ้นอย่างสม่ำเสมอ ในด้านหนึ่ง สิ่งนี้บังคับให้นักชีววิทยาต้องละทิ้งการใช้ยีนแต่ละตัวเป็นตัวบ่งชี้ที่ดีสำหรับประวัติศาสตร์ของสิ่งมีชีวิต ในอีกด้านหนึ่ง นี่เป็นแหล่งข้อมูลขนาดใหญ่ที่ยังไม่ได้ถูกนำมาใช้ประโยชน์อย่างเต็มที่เกี่ยวกับอดีต
สามขอบเขตของชีวิต
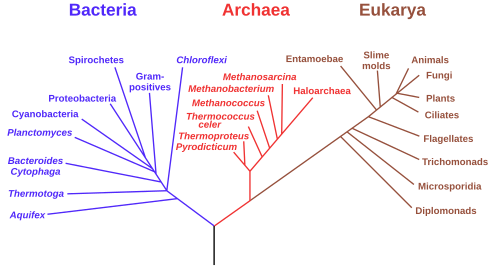
นักจุลชีววิทยาได้ศึกษาอย่างละเอียด เกี่ยวกับ สาขาหลักสามสาขาแรกของแผนภูมิวิวัฒนาการของสิ่งมีชีวิตเนื่องจากสิ่งมีชีวิตกลุ่มแรกคือจุลินทรีย์ นักจุลชีววิทยา (นำโดยคาร์ล โวเอส ) ได้เสนอคำว่า"โดเมน"สำหรับสาขาหลักสามสาขาของแผนภูมิวิวัฒนาการนี้ โดยที่โดเมนเป็นคำศัพท์ทางวิวัฒนาการที่มีความหมายคล้ายกับ " อาณาจักร " ทางชีววิทยา ในการสร้างแผนภูมิวิวัฒนาการนี้ขึ้นใหม่ ลำดับยีนที่เข้ารหัสหน่วยย่อยขนาดเล็กของไรโบโซมอาร์เอ็นเอ (SSU rRNA, 16s rRNA ) ได้พิสูจน์แล้วว่ามีประโยชน์ และแผนภูมิวิวัฒนาการ (ดังแสดงในภาพ) อาศัยข้อมูลจากยีนเพียงยีนเดียวนี้เป็นอย่างมาก
โดเมนของสิ่งมีชีวิตทั้งสามนี้แสดงถึงสายวิวัฒนาการหลักของสิ่งมีชีวิตเซลล์เดียวในยุคแรกเริ่ม และปัจจุบันประกอบด้วยแบคทีเรีย อา ร์เคีย (สิ่งมีชีวิตเซลล์เดียวที่มีลักษณะภายนอกคล้ายแบคทีเรีย) และยูคาริโอตา ยูคาริโอตาประกอบด้วยเฉพาะสิ่งมีชีวิตที่มีนิวเคลียสที่ชัดเจน เช่นราโปรติสต์และสิ่งมีชีวิตทั้งหมดในอาณาจักรพืชและสัตว์ (ดูภาพประกอบ)
ยีนที่ใช้บ่อยที่สุดในการสร้างความสัมพันธ์ทางวิวัฒนาการในจุลินทรีย์คือยีน ribosomal RNA หน่วยย่อยขนาดเล็ก เนื่องจากลำดับของมันมีแนวโน้มที่จะได้รับการอนุรักษ์ไว้ในสมาชิกที่มีระยะห่างทางวิวัฒนาการใกล้เคียงกัน แต่ก็มีความแปรปรวนมากพอที่จะวัดความแตกต่างได้ [ 1 ] การใช้ SSU rRNAเป็นตัววัดระยะห่างทางวิวัฒนาการได้รับการริเริ่มโดยCarl Woeseเมื่อเขาสร้าง "แผนภูมิวิวัฒนาการ" สมัยใหม่เป็นครั้งแรก และผลลัพธ์ของเขานำไปสู่การเสนอArchaeaเป็นโดเมนที่สามของสิ่งมีชีวิตอย่างไรก็ตาม เมื่อเร็วๆ นี้มีการโต้แย้งว่ายีน SSU rRNA สามารถถ่ายโอนในแนวนอนได้เช่นกัน[ 2 ] แม้ว่าสิ่งนี้อาจเกิดขึ้นได้ยาก แต่ความเป็นไปได้นี้กำลังบังคับให้มีการตรวจสอบความถูกต้องของแผนภูมิวิวัฒนาการที่อิงตาม SSU rRNA
การค้นพบ HGT ที่แพร่หลายในจุลินทรีย์เมื่อเร็ว ๆ นี้ และการตรวจพบการเคลื่อนย้ายในแนวนอนของยีนแม้กระทั่งยีนสำหรับหน่วยย่อยขนาดเล็กของ RNA ไรโบโซม ทำให้เหล่านักชีววิทยาต้องตั้งคำถามถึงความถูกต้องของกิ่งก้านแรก ๆ ในแผนภูมิวิวัฒนาการ และแม้กระทั่งตั้งคำถามถึงความถูกต้องของแผนภูมิวิวัฒนาการในฐานะแบบจำลองที่มีประโยชน์ของวิวัฒนาการในยุคแรก[ 3 ]ในความเป็นจริง วิวัฒนาการในยุคแรกถือว่าเกิดขึ้นจากชุมชนของโปรเจโนตซึ่งสามารถแลกเปลี่ยนโมเลกุลขนาดใหญ่ได้เมื่อ HGT เป็นมาตรฐาน การถ่ายโอนยีนในแนวนอนนี้เกิดขึ้นนอกเหนือจากเกณฑ์ของดาร์วินหลังจากที่พันธุกรรมหรือการถ่ายโอนยีนในแนวดิ่งได้รับการสถาปนาขึ้น[ 4 ] [ 5 ] "การเปรียบเทียบลำดับชี้ให้เห็นถึงการถ่ายโอนยีน ในแนวนอนเมื่อเร็ว ๆ นี้ ใน หลาย สายพันธุ์รวมถึงข้ามขอบเขตของ "โดเมน" ทางวิวัฒนาการดังนั้นการกำหนดประวัติทางวิวัฒนาการของสายพันธุ์จึงไม่สามารถทำได้อย่างแน่ชัดโดยการกำหนดแผนภูมิวิวัฒนาการสำหรับยีนเดี่ยว" [ 6 ]ดังนั้น HGT จึงเป็นปัจจัยรบกวน ที่อาจเกิดขึ้น ในการอนุมานแผนภูมิวิวัฒนาการจากลำดับของยีนเดียวตัวอย่างเช่น หากแบคทีเรียสองชนิดที่มีความสัมพันธ์ห่างไกลกันได้แลกเปลี่ยนยีนกันแผนภูมิวิวัฒนาการที่รวมสายพันธุ์เหล่านั้นจะแสดงให้เห็นว่าพวกมันมีความสัมพันธ์ใกล้ชิดกัน แม้ว่ายีนอื่นๆ ส่วนใหญ่จะแตกต่างกันอย่างมากก็ตาม ด้วยเหตุนี้ การใช้ข้อมูลอื่นๆ เพื่ออนุมานแผนภูมิวิวัฒนาการจึงมีความสำคัญ เช่น การมีอยู่หรือไม่มีอยู่ของยีน หรือโดยทั่วไปแล้ว การรวมยีนที่หลากหลายที่สุดเท่าที่จะเป็นไปได้สำหรับการวิเคราะห์[ 6 ]
เชื่อกันว่ามีการถ่ายโอนยีนแนวนอน (HGT) เกิดขึ้นก่อนหน้านี้บรรพบุรุษร่วมสากลแรก (FUCA) ซึ่งเป็นบรรพบุรุษที่เก่าแก่ที่สุดของบรรพบุรุษร่วมสุดท้ายของสิ่งมีชีวิตทั้งหมด (LUCA) เชื่อกันว่ามีลูกหลานอื่น ๆ ที่มีสายเลือดของตนเอง[ 7 ] [ 8 ]สายเลือดพี่น้องของ LUCA ที่สูญพันธุ์ไปแล้วซึ่งสืบเชื้อสายมาจาก FUCA เชื่อกันว่าได้ถ่ายโอนยีนบางส่วนในแนวนอนไปยังจีโนมของลูกหลานยุคแรกของ LUCA [ 8 ]
เลือกใช้คำอุปมา: ต้นไม้, ตาข่าย, ใยแมงมุม หรือ วงแหวน
ในบทความUprooting the Tree of Life ของเขา W. Ford Doolittleได้กล่าวถึงบรรพบุรุษร่วมสากลสุดท้าย – รากของต้นไม้แห่งชีวิต – และปัญหาของแนวคิดนั้นที่เกิดจาก HGT [ 9 ]เขาอธิบายว่าจุลินทรีย์Archaeoglobus fulgidusเป็นสิ่งผิดปกติเมื่อเทียบกับ ต้นไม้ ทางวิวัฒนาการโดยอิงจากรหัสของเอนไซม์HMGCoA reductase – สิ่งมีชีวิตนี้เป็นอาร์เคียนอย่างแน่นอน โดยมีไขมันเซลล์และกลไกการถอดรหัสทั้งหมดที่คาดหวังจากอาร์เคียน แต่ยีน HMGCoA ของมันมีต้นกำเนิดมาจากแบคทีเรีย ในบทความ Doolittle กล่าวว่าในขณะที่ปัจจุบันเป็นที่ยอมรับกันอย่างกว้างขวางว่าไมโท คอนเดรีย ในยูคาริโอตมาจากเซลล์อัลฟา-โปรตีโอแบคทีเรีย และคลอโรพลาสต์ มาจาก ไซยาโนแบคทีเรียที่ถูกกลืนกิน[ 9 ]
"...จึงไม่ปลอดภัยอีกต่อไปที่จะสันนิษฐานว่านั่นเป็นการถ่ายทอดยีนในแนวนอนเพียงอย่างเดียวที่เกิดขึ้นหลังจากยูคาริโอตกลุ่มแรกถือกำเนิดขึ้น เฉพาะในยูคาริโอตหลายเซลล์ในยุคหลังเท่านั้นที่เราทราบถึงข้อจำกัดที่ชัดเจนเกี่ยวกับการแลกเปลี่ยนยีนในแนวนอน เช่น การเกิดขึ้นของเซลล์สืบพันธุ์ ที่แยกออกจากกัน (และได้รับการปกป้อง) ...
หากไม่มีการถ่ายโอนยีนด้านข้างเลย ต้นไม้ของยีนแต่ละต้นจะมีโทโพโลยีเดียวกัน (ลำดับการแตกแขนงเดียวกัน) และยีนบรรพบุรุษที่รากของแต่ละต้นไม้จะมีอยู่ในบรรพบุรุษร่วมสากลสุดท้าย ซึ่งเป็นเซลล์โบราณเพียงเซลล์เดียว แต่การถ่ายโอนที่กว้างขวางหมายความว่าไม่ใช่เช่นนั้น ต้นไม้ของยีนจะแตกต่างกัน (แม้ว่าหลายต้นจะมีบริเวณที่มีโทโพโลยีคล้ายกัน) และจะไม่มีเซลล์เดียวที่สามารถเรียกว่าบรรพบุรุษร่วมสากลสุดท้ายได้..." [ 9 ]
ดูลิตเติลเสนอว่าบรรพบุรุษร่วมสากลไม่น่าจะเป็นสิ่งมีชีวิตชนิดใดชนิดหนึ่งโดยเฉพาะ แต่ต้องเป็นกลุ่มเซลล์ดั้งเดิมที่หลากหลายและวิวัฒนาการร่วมกัน เซลล์ยุคแรกเหล่านี้แต่ละเซลล์มีจำนวนยีนค่อนข้างน้อย แตกต่างกันในหลายด้าน และแลกเปลี่ยนยีนกันอย่างอิสระ ในที่สุด จากเซลล์ที่หลากหลายเหล่านี้ก็กำเนิดเป็นสามโดเมนของสิ่งมีชีวิตอย่างที่เรารู้จักในปัจจุบัน ได้แก่แบคทีเรีย อา ร์เคียและยูคาริโอตโดเมนเหล่านี้มีความแตกต่างกันอย่างเห็นได้ชัดในปัจจุบัน เนื่องจากการถ่ายโอนยีนส่วนใหญ่ที่ยังคงเกิดขึ้นนั้นเกิดขึ้นภายในโดเมนเหล่านี้ มากกว่าที่จะเกิดขึ้นระหว่างโดเมน นักชีววิทยาปีเตอร์ โกการ์เทนได้เสริมข้อโต้แย้งเหล่านี้ และเสนอว่าอุปมาเรื่องต้นไม้ไม่เหมาะสมกับข้อมูลจากการวิจัยจีโนมล่าสุด และนักชีววิทยาควรใช้ "อุปมาเรื่องโมเสกเพื่ออธิบายประวัติศาสตร์ที่แตกต่างกันที่รวมกันอยู่ในจีโนมแต่ละตัว และใช้อุปมาเรื่องตาข่ายเพื่อแสดงภาพการแลกเปลี่ยนที่หลากหลายและผลกระทบร่วมกันของ HGT ในหมู่จุลินทรีย์" [ 10 ]
การขจัดความไม่แน่นอนด้วยพันธุศาสตร์เชิงวิวัฒนาการ
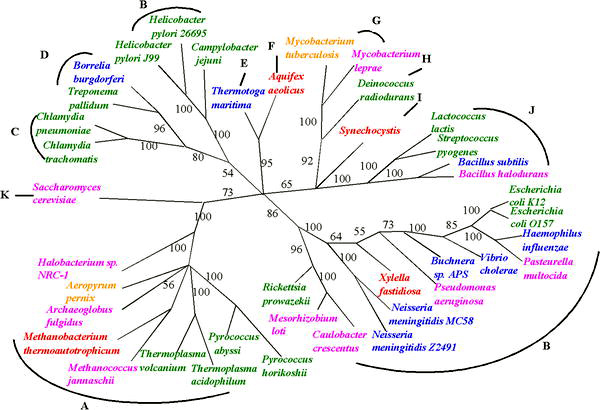
แม้จะมีความไม่แน่นอนในการสร้างแผนภูมิวิวัฒนาการย้อนกลับไปถึงจุดเริ่มต้นของสิ่งมีชีวิต แต่ก็มีความคืบหน้าในการสร้างแผนภูมิวิวัฒนาการของสิ่งมีชีวิตขึ้นมาใหม่ท่ามกลางความไม่แน่นอนที่เกิดจาก HGT ความไม่แน่นอนของแผนภูมิวิวัฒนาการที่อนุมานขึ้นจากยีนเพียงยีนเดียวสามารถแก้ไขได้โดยใช้ยีนทั่วไปหลายยีนหรือแม้แต่หลักฐานจากจีโนมทั้งหมด[ 12 ]แนวทางหนึ่งดังกล่าว ซึ่งบางครั้งเรียกว่า 'การพิมพ์แบบหลายตำแหน่ง' ได้ถูกนำมาใช้เพื่ออนุมานแผนภูมิวิวัฒนาการสำหรับสิ่งมีชีวิตที่แลกเปลี่ยนยีนกัน เช่น แบคทีเรียที่ทำให้เกิดโรคเยื่อหุ้มสมองอักเสบ[ 13 ]
โจนาธาน ไอเซนและแคลร์ เฟรเซอร์ได้ชี้ให้เห็นว่า:
“ในการสร้างแผนภูมิวิวัฒนาการของสิ่งมีชีวิต การวิเคราะห์จีโนมทั้งหมดได้เริ่มเข้ามาเสริม และในบางกรณีก็ปรับปรุงการศึกษาที่เคยทำมาก่อนโดยใช้ยีนเพียงหนึ่งหรือสองยีน ตัวอย่างเช่น การศึกษาจีโนมแบคทีเรียที่สมบูรณ์เมื่อเร็ว ๆ นี้ชี้ให้เห็นว่า สายพันธุ์ ที่ทนความร้อนสูงไม่ได้แตกแขนงออกไปลึก หากเป็นเช่นนั้นจริง ก็จะทำให้เกิดข้อสงสัยเกี่ยวกับแนวคิดที่ว่าสิ่งมีชีวิตรูปแบบแรกเป็นสิ่งมีชีวิตที่ทนความร้อน การวิเคราะห์จีโนมของปรสิตยูคาริโอตEncephalitozoon cuniculiสนับสนุนข้อเสนอแนะที่ว่ากลุ่มMicrosporidiaไม่ใช่โปรติสต์ที่แตกแขนงออกไปลึก แต่เป็นสมาชิกของอาณาจักรเชื้อรา การวิเคราะห์จีโนมยังสามารถช่วยแก้ไขความสัมพันธ์ภายในสายพันธุ์ได้ เช่น โดยการให้เครื่องหมายทางพันธุกรรมใหม่สำหรับการศึกษาพันธุศาสตร์ประชากรในแบคทีเรียที่ก่อให้เกิดโรคแอนแทรกซ์หรือวัณโรค ในการศึกษาทั้งหมดนี้ ข้อมูลเพิ่มเติมที่ได้จากลำดับจีโนมที่สมบูรณ์จะช่วยให้สามารถแยกสัญญาณทางวิวัฒนาการออกจากสัญญาณรบกวนได้ นี่ไม่ได้หมายความว่าแผนภูมิวิวัฒนาการของสิ่งมีชีวิตได้รับการแก้ไขแล้ว – เราเพิ่งสุ่มตัวอย่างจีโนมเพียงเล็กน้อยเท่านั้น” และหลายกลุ่มยังไม่ได้รับผลกระทบ” [ 14 ]
แนวทางเหล่านี้ทำให้สามารถประมาณความถี่สัมพัทธ์ของ HGT ได้ ค่าที่ค่อนข้างต่ำที่สังเกตได้บ่งชี้ว่า 'ต้นไม้' ยังคงเป็นอุปมาที่ถูกต้องสำหรับวิวัฒนาการ แต่ต้นไม้นั้นประดับประดาไปด้วย 'ใยแมงมุม' ของยีนที่ถ่ายโอนในแนวนอน นี่คือข้อสรุปหลักของการศึกษาในปี 2548 เกี่ยวกับลำดับจีโนมจุลินทรีย์ที่สมบูรณ์มากกว่า 40 ลำดับโดย Fan Ge, Li-San Wang และJunhyong Kimพวกเขาประมาณความถี่ของเหตุการณ์ HGT ไว้ที่ประมาณ 2% ของยีนหลักต่อจีโนม[ 15 ]แนวทางจีโนมทั้งหมดที่คล้ายกันในการประเมินวิวัฒนาการยังช่วยให้มีความก้าวหน้าในการระบุเหตุการณ์ในช่วงแรก ๆ ในต้นไม้แห่งชีวิต เช่น ข้อเสนอที่ว่ายูคาริโอตเกิดขึ้นจากการรวมกันของจีโนมโปรคาริโอตที่สมบูรณ์แต่แตกต่างกันมากสองจีโนม: หนึ่งจากแบคทีเรียและอีกหนึ่งจากเซลล์อาร์เคีย[ 3 ]
สมมติฐานการหลอมรวมของสิ่งมีชีวิตดังกล่าวสำหรับต้นกำเนิดของเซลล์ที่มีนิวเคลียสที่ซับซ้อนได้รับการเสนอโดยLynn Margulisโดยใช้เหตุผลที่แตกต่างกันอย่างสิ้นเชิงเกี่ยวกับภาวะพึ่งพาอาศัยกันระหว่างแบคทีเรียและอาร์เคียที่เกิดขึ้นในกลุ่มจุลินทรีย์โบราณ[ 16 ]
ใช้ในการศึกษาด้านวิวัฒนาการ
แม้ว่า HGT มักถูกมองว่าเป็นความท้าทายสำหรับการสร้างแผนภูมิวิวัฒนาการของสิ่งมีชีวิต แต่ในอีกมุมมองหนึ่ง มันกลับให้ข้อมูลที่มีค่าเพิ่มเติมสำหรับการสร้างแผนภูมิวิวัฒนาการดังกล่าว
ประการแรก สำหรับสิ่งมีชีวิตผู้รับ HGT เป็นการกลายพันธุ์ของ DNAเช่นเดียวกับการกลายพันธุ์อื่นๆ และด้วยเหตุนี้จึงสามารถสร้างแบบจำลองและนำไปใช้ในการสร้างแผนภูมิและรากของต้นไม้ได้[ 17 ]
ประการที่สอง จำเป็นที่ผู้รับการได้มาซึ่งยีนโดย HGT จะต้องมีชีวิตอยู่ในช่วงเวลาเดียวกันกับผู้ให้ หรือในช่วงเวลาต่อมา[ 18 ]ผลที่ตามมาคือมีข้อมูลเกี่ยวกับช่วงเวลาของการกระจายตัวใน HGT [ 19 ]สิ่งนี้ยิ่งน่าทึ่งมากขึ้นไปอีก เนื่องจากแหล่งข้อมูลหลักที่ใช้ในการกำหนดอายุในโลกของสิ่งมีชีวิต ซึ่งก็คือบันทึกฟอสซิลนั้น ไม่มีอยู่จริงในบริเวณที่มี HGT มากมาย นั่นก็คือโลกของจุลินทรีย์
ประการที่สาม จะให้ข้อมูลเกี่ยวกับความหลากหลายทางชีวภาพที่สูญพันธุ์ เนื่องจากการถ่ายโอนมีแนวโน้มที่จะเกิดขึ้นจากสายพันธุ์ที่สูญพันธุ์[ 20 ]
ดูเพิ่มเติม
- ชีววิทยาการพัฒนาเชิงวิวัฒนาการ
- การถ่ายทอดยีนในแนวนอน
- การอนุมานการถ่ายทอดยีนในแนวนอน
- แผนภูมิวิวัฒนาการของสิ่งมีชีวิต (ชีววิทยา)
อ่านเพิ่มเติม
- Gogarten, JP; Townsend, JP (2005). "การถ่ายโอนยีนแนวนอน นวัตกรรมจีโนม และวิวัฒนาการ" Nature Reviews. Microbiology . 3 (9): 679– 87. doi : 10.1038/nrmicro1204 . PMID 16138096 . S2CID 8092665 .บทความหนึ่งในวารสารNature Reviews Microbiology ฉบับเต็ม ซึ่งส่วนใหญ่กล่าวถึงการถ่ายทอดยีนแนวนอน (HGT)
- Daubin, V; Moran, NA; Ochman, H (2003). "Phylogenetics and the cohesion of bacterial genomes". Science . 301 ( 5634): 829– 32. Bibcode : 2003Sci...301..829D . doi : 10.1126/science.1086568 . PMID 12907801. S2CID 11268678 .
- Eisen, JA; Fraser, CM (2003) . "มุมมองฟิโลจีโนมิกส์: จุดตัดระหว่างวิวัฒนาการและจีโนมิกส์". Science . 300 (5626): 1706–7 . Bibcode : 2003Sci...300.1706E . doi : 10.1126/science.1086292 . PMID 12805538. S2CID 42394233 .
- Ge, F; Wang, LS; Kim, J (2005). "ใยแมงมุมแห่งชีวิตที่เปิดเผยโดยการประมาณการการถ่ายโอนยีนแนวนอนในระดับจีโนม" PLOS Biology 3 ( 10): e316. doi : 10.1371/journal.pbio.0030316 . PMC 1233574 . PMID 16122348 .

- เฮนซ์ เอสอาร์; ฮูสัน ดีเอช; อ๊ะ AF; นีเซลท์-สตรูเว, เค; ชูสเตอร์, เซาท์แคโรไลนา (2005) "สายวิวัฒนาการโปรคาริโอตทั้งจีโนม " ชีวสารสนเทศศาสตร์ . 21 (10): 2329– 35. ดอย : 10.1093/bioinformatics/bth324 . PMID15166018 .
- Lerat, E; Daubin, V; Ochman, H; Moran, NA (2005). "ต้นกำเนิดวิวัฒนาการของชุดยีนในแบคทีเรีย" . PLOS Biology . 3 (5): e130. doi : 10.1371/journal.pbio.0030130 . PMC 1073693 . PMID 15799709 .
- Urwin, R; Maiden, MC (2003). "การพิมพ์ลำดับหลายตำแหน่ง: เครื่องมือสำหรับระบาดวิทยาระดับโลก" Trends Microbiol . 11 (10): 479– 87. doi : 10.1016/j.tim.2003.08.006 . PMID 14557031 .
- Adoutte, A; Balavoine, G; Lartillot, N; Lespinet, O; Prud'homme, B; De Rosa, R (2000). "วิวัฒนาการของสัตว์แบบใหม่: ความน่าเชื่อถือและนัยยะ" . Proc Natl Acad Sci USA . 97 (9): 4453– 6. Bibcode : 2000PNAS...97.4453A . doi : 10.1073/pnas.97.9.4453 . PMC 34321 . PMID 10781043 .
- Delsuc, F; Brinkmann, H; Philippe, H (2005). "Phylogenomics and the reconstruction of the tree of life". Nat Rev Genet . 6 (5): 361– 75. CiteSeerX 10.1.1.333.1615 . doi : 10.1038/nrg1603 . PMID 15861208 . S2CID 16379422 .
- Steenkamp, ET; Wright, J; Baldauf, SL (2006). "ต้นกำเนิดของสัตว์และเชื้อราจากโปรติสต์" . Mol Biol Evol . 23 (1): 93– 106. doi : 10.1093/molbev/msj011 . PMID 16151185 .
- Thomas, CM; Nielsen, KM. (2005). "กลไกและอุปสรรคต่อการถ่ายทอดยีนในแนวนอนระหว่างแบคทีเรีย" Nat Rev Microbiol . 3 (9): 711– 21. doi : 10.1038/nrmicro1234 . PMID 16138099. S2CID 1231127 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การถ่ายทอดยีนในแนวนอนในวิวัฒนาการ
การถ่ายทอดยีนในแนวนอน ( HGT ) หมายถึงการถ่ายทอดยีนระหว่างกิ่งก้านที่อยู่ห่างไกลกันบนแผนภูมิวิวัฒนาการในทางวิวัฒนาการ...
สามขอบเขตของชีวิต
นักจุลชีววิทยา ได้ศึกษาอย่างละเอียด เกี่ยวกับ สาขาหลักสามสาขาแรกของ แผนภูมิวิวัฒนาการของสิ่งมีชีวิต เนื่องจากสิ่งมีชีวิตกลุ่มแรกคือจุลินทรีย์ นักจุลชีววิทยา (นำโดย คาร์ล โวเอส ) ได้เสนอคำว่า "โดเมน" สำหรับสาขาหลักสามสาขาของแผนภูมิวิวัฒนาการนี้ โดยที่ โดเมน...
เลือกใช้คำอุปมา: ต้นไม้, ตาข่าย, ใยแมงมุม หรือ วงแหวน
ในบทความ Uprooting the Tree of Life ของเขา W. Ford Doolittle ได้กล่าวถึง บรรพบุรุษร่วมสากลสุดท้าย – รากของต้นไม้แห่งชีวิต – และปัญหาของแนวคิดนั้นที่เกิดจาก HGT [ 9 ] เขาอธิบายว่าจุลินทรีย์ Archaeoglobus fulgidus เป็นสิ่งผิดปกติเมื่อเทียบกับ ต้นไม้...
การขจัดความไม่แน่นอนด้วยพันธุศาสตร์เชิงวิวัฒนาการ
แม้จะมีความไม่แน่นอนในการสร้างแผนภูมิวิวัฒนาการย้อนกลับไปถึงจุดเริ่มต้นของสิ่งมีชีวิต แต่ก็มีความคืบหน้าในการสร้างแผนภูมิวิวัฒนาการของสิ่งมีชีวิตขึ้นมาใหม่ท่ามกลางความไม่แน่นอนที่เกิดจาก HGT...