อ่าน 10 นาที
การทำซ้ำแบบกลับด้าน
ลำดับ นิวคลีโอไทด์ แบบกลับหัว (หรือ IR ) คือลำดับนิว คลีโอไทด์ สายเดี่ยวที่ตามด้วย ลำดับนิวคลีโอ ไทด์ผกผันที่อยู่ถัด ไป [ 1 ]...
การทำซ้ำแบบกลับด้าน
ลำดับ นิวคลีโอไทด์ แบบกลับหัว (หรือIR ) คือลำดับนิว คลีโอไทด์สายเดี่ยวที่ตามด้วยลำดับนิวคลีโอ ไทด์ผกผันที่อยู่ถัด ไป[ 1 ]ลำดับนิวคลีโอไทด์ที่คั่นระหว่างลำดับเริ่มต้นกับลำดับนิวคลีโอไทด์ผกผันสามารถมีความยาวได้ทุกระยะ รวมถึงศูนย์ ตัวอย่างเช่น5'---TTACGnnnnnn CGTAA---3'เป็นลำดับนิวคลีโอไทด์แบบกลับหัว เมื่อความยาวของส่วนที่คั่นเป็นศูนย์ ลำดับที่ได้จะเป็นลำดับพาลินโดรม[ 2 ]
ทั้งลำดับนิวคลีโอไทด์แบบกลับหัวและแบบตรงต่างก็เป็นลำดับนิวคลีโอไทด์ที่เกิดขึ้นซ้ำๆลำดับ DNA ที่ซ้ำกัน เหล่านี้ มักมีตั้งแต่คู่ของนิวคลีโอไทด์ไปจนถึงยีน ทั้งยีน ในขณะที่ความใกล้ชิดของลำดับที่ซ้ำกันนั้นแตกต่างกันไปตั้งแต่แบบกระจายตัวอย่างกว้างขวางไปจนถึงแบบเรียงต่อกันอย่างง่าย[ 3 ]ลำดับที่ซ้ำกันแบบเรียงต่อกันสั้นๆอาจมีอยู่เพียงไม่กี่สำเนาในบริเวณเล็กๆ ไปจนถึงหลายพันสำเนาที่กระจายอยู่ทั่วจีโนมของยูคาริโอตส่วน ใหญ่ [ 4 ] ลำดับที่ซ้ำกันที่มี เบสประมาณ 10–100 คู่ เรียก ว่า มินิ แซทเทลไลต์ ในขณะที่ลำดับที่ซ้ำกันที่สั้นกว่าซึ่งส่วนใหญ่มีเบส 2–4 คู่เรียกว่าไมโครแซทเทลไลต์ [ 5 ] ลำดับที่ซ้ำกันที่พบได้บ่อยที่สุด ได้แก่ ลำดับที่ซ้ำกันของไดนิวคลีโอไทด์ ซึ่งมีเบส ACบนสาย DNA หนึ่งสาย และ GT บนสายที่เสริมกัน[ 3 ]องค์ประกอบบางส่วนของจีโนมที่มีลำดับเฉพาะทำหน้าที่เป็นเอ็กซอนอินทรอนและ DNA ควบคุม[ 6 ]แม้ว่าตำแหน่งที่คุ้นเคยที่สุดของลำดับซ้ำคือเซนโทรเมียร์และเทโลเมียร์ [ 6 ] แต่ลำดับซ้ำส่วนใหญ่ในจีโนมก็พบอยู่ในดีเอ็นเอที่ไม่เข้ารหัส[ 5 ]
ลำดับซ้ำแบบกลับหัวมีหน้าที่ทางชีววิทยาที่สำคัญหลายประการ พวกมันกำหนดขอบเขตในทรานสโพซอน และบ่งชี้บริเวณที่สามารถ จับคู่เบสแบบเสริมกันได้(บริเวณภายในลำดับเดียวที่สามารถจับคู่เบสกันได้) คุณสมบัติเหล่านี้มีบทบาทสำคัญในความไม่เสถียรของจีโนม[ 7 ]และมีส่วนช่วยไม่เพียงแต่ต่อ วิวัฒนาการ ของเซลล์และความหลากหลายทางพันธุกรรม[ 8 ]แต่ยังรวมถึงการกลายพันธุ์และโรคด้วย[ 9 ] เพื่อศึกษาผลกระทบเหล่านี้อย่างละเอียด จึงมีการพัฒนาโปรแกรมและฐานข้อมูลจำนวนมากเพื่อช่วยในการค้นหาและระบุลำดับซ้ำแบบกลับหัวในจีโนมต่างๆ
ทำความเข้าใจเกี่ยวกับลำดับซ้ำแบบกลับด้าน
ตัวอย่างของการทำซ้ำแบบกลับด้าน
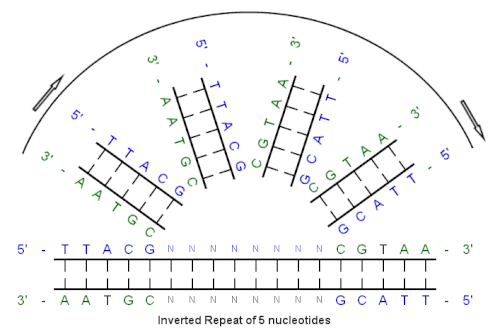
เริ่มจากลำดับเริ่มต้นนี้: 5'-TTACG-3'
ส่วนเติมเต็มที่เกิดจากการจับคู่เบสคือ: 3'-AATGC-5'
ส่วนเติมเต็มแบบกลับด้านคือ: 5'-CGTAA-3'
และลำดับการทำซ้ำแบบกลับด้านคือ: 5'---TTACGnnnnnn CGTAA---3'
"nnnnnn" แทนจำนวนนิวคลีโอไทด์ที่คั่นอยู่ใดๆ ก็ได้
เทียบกับการทำซ้ำโดยตรง
การทำซ้ำโดยตรงเกิดขึ้นเมื่อลำดับถูกทำซ้ำด้วยรูปแบบเดียวกันที่ปลายน้ำ[ 1 ] ไม่มีการผกผันและไม่มีส่วนเติมเต็มย้อนกลับที่เกี่ยวข้องกับการทำซ้ำโดยตรง ลำดับนิวคลีโอไทด์ที่เขียนด้วยตัวหนาแสดงถึงลำดับที่ทำซ้ำ อาจมีหรือไม่มีนิวคลีโอไทด์คั่นกลางก็ได้
- 5´ TTACG nnnnnnTTACG 3´
- 3´ AATGC nnnnnnAATGC 5´
ในทางภาษาศาสตร์ การซ้ำคำโดยตรงโดยทั่วไปเทียบได้กับการสัมผัสคล้องจอง เช่น "time on ad ime "
เทียบกับการทำซ้ำแบบคู่ขนาน
ลำดับนิวคลีโอไทด์ที่ซ้ำกันโดยตรงโดยไม่มีนิวคลีโอไทด์คั่นระหว่างลำดับเริ่มต้นและสำเนาถัดไปเรียกว่า ลำดับซ้ำแบบ แทนเดม (Tandem repeat ) ลำดับนิวคลีโอไทด์ที่เขียนด้วยตัวหนาแสดงถึงลำดับที่ซ้ำกัน
- 5´ TTACG TTACG 3´
- 3´ AATGC AATGC 5´
ในทางภาษาศาสตร์ การพูดซ้ำแบบคู่โดยทั่วไปเทียบได้กับการพูดติดอ่าง หรือการพูดซ้ำคำโดยเจตนา เช่นในคำว่า "ลาก่อน"
เทียบกับ พาลินโดรม
ลำดับการทำซ้ำแบบกลับด้านที่ไม่มีนิวคลีโอไทด์คั่นระหว่างลำดับเริ่มต้นและส่วนเติมเต็มแบบย้อนกลับ ที่อยู่ถัดไป เรียกว่าพาลินโดรม [ 1 ] ตัวอย่าง : ขั้นตอนที่ 1: เริ่มต้นด้วยการทำซ้ำแบบกลับด้าน: 5' TTACGnnnnnnCGTAA 3' ขั้นตอนที่ 2: ลบนิวคลีโอไทด์คั่น: 5' TTACGCGTAA 3' ลำดับที่ได้นี้เป็นพาลินโดรมเพราะเป็นส่วนเติมเต็มแบบย้อนกลับของตัวมันเอง[ 1 ]
- ลำดับทดสอบ5' TTACGCGTAA 3' (จากขั้นตอนที่ 2 โดยตัดนิวคลีโอไทด์ที่คั่นกลางออก)
- 3' AATGCGCATT 5' ส่วนเติมเต็มของลำดับทดสอบ
- 5' TTACGCGTAA 3' ลำดับกลับด้าน (reverse complement) นี่คือลำดับเดียวกับลำดับทดสอบข้างต้น ดังนั้นจึงเป็นพาลินโดรม (palindrome)
ลักษณะทางชีวภาพและการทำงาน
สภาวะที่เอื้อต่อการสังเคราะห์
ลำดับซ้ำที่หลากหลายทั่วทั้งจีโนมได้มาจากองค์ประกอบที่เคลื่อนย้ายได้ซึ่งในปัจจุบันเข้าใจกันว่าสามารถ "กระโดด" ไปยังตำแหน่งจีโนมต่างๆ ได้โดยไม่ถ่ายโอนสำเนาเดิม[ 10 ]การเคลื่อนย้ายลำดับเดียวกันซ้ำๆ กันหลายชั่วอายุคนทำให้มั่นใจได้ว่าลำดับเหล่านั้นจะมีอยู่มากมายทั่วทั้งจีโนม[ 10 ]การรวมตัวกันอย่างจำกัดของลำดับระหว่างองค์ประกอบลำดับที่แตกต่างกันสององค์ประกอบที่เรียกว่าการรวมตัวกันเฉพาะไซต์แบบอนุรักษ์ (CSSR) ส่งผลให้เกิดการผกผันของส่วน DNA โดยขึ้นอยู่กับการจัดเรียงของลำดับการจดจำการรวมตัวกันบน DNA ผู้ให้และ DNA ผู้รับ[ 10 ]อีกครั้ง การวางแนวของไซต์การรวมตัวกันสองไซต์ภายในโมเลกุล DNA ผู้ให้เมื่อเทียบกับความไม่สมมาตรของลำดับการตัด DNA ที่คั่นกลาง ซึ่งเรียกว่าบริเวณครอสโอเวอร์ มีความสำคัญต่อการก่อตัวของลำดับซ้ำแบบผกผันหรือลำดับซ้ำแบบตรง[ 10 ]ดังนั้น การรวมตัวกันที่เกิดขึ้นที่ไซต์แบบผกผันคู่หนึ่งจะผกผันลำดับ DNA ระหว่างไซต์ทั้งสอง[ 10 ]พบว่าโครโมโซมที่มีความเสถียรสูงมากมีจำนวนการทำซ้ำแบบกลับด้านน้อยกว่าการทำซ้ำแบบตรง ซึ่งบ่งชี้ถึงความสัมพันธ์ระหว่างความเสถียรของโครโมโซมกับจำนวนการทำซ้ำ[ 11 ]
ภูมิภาคที่การปรากฏตัวเป็นสิ่งจำเป็น
มีการสังเกตพบการทำซ้ำแบบกลับหัวที่ปลายในดีเอ็นเอของทรานสโพซอนยูคาริโอตต่างๆ แม้ว่าแหล่งที่มาของพวกมันจะยังไม่เป็นที่ทราบแน่ชัดก็ตาม[ 12 ]การทำซ้ำแบบกลับหัวส่วนใหญ่พบที่จุดเริ่มต้นของการจำลองแบบของเซลล์สิ่งมีชีวิตและออร์แกเนลล์ต่างๆ ตั้งแต่พลาสมิดของฟาจ ไมโตคอนเดรีย และไวรัสยูคาริโอต ไปจนถึงเซลล์ของสัตว์เลี้ยงลูกด้วยนม[ 13 ]จุดเริ่มต้นของการจำลองแบบของฟาจ G4 และฟาจอื่นๆ ที่เกี่ยวข้องประกอบด้วยส่วนของเบสนิวคลีโอไทด์เกือบ 139 เบส ซึ่งรวมถึงลำดับการทำซ้ำแบบกลับหัวสามลำดับที่จำเป็นสำหรับการเริ่มต้นการจำลองแบบ[ 13 ]
ในจีโนม
ในระดับหนึ่ง ส่วนของนิวคลีโอไทด์ที่ซ้ำกันมักพบเห็นได้บ่อยในส่วนประกอบของ DNA ที่หายาก[ 14 ]การซ้ำหลักสามแบบที่พบได้มากในโครงสร้าง DNA เฉพาะ ได้แก่ การซ้ำแบบกลับด้านของโฮโมพิวรีน-โฮโมไพริมิดีนที่แม่นยำอย่างใกล้ชิด ซึ่งเรียกอีกอย่างว่า H พาลินโดรม ซึ่งเป็นปรากฏการณ์ทั่วไปในโครงสร้างเกลียวสามชั้น H ที่อาจประกอบด้วยไตรแอดนิวคลีโอไทด์ TAT หรือ CGC ส่วนอื่นๆ อาจอธิบายได้ว่าเป็นการซ้ำแบบกลับด้านที่ยาวซึ่งมีแนวโน้มที่จะสร้างแฮร์พินและรูปกากบาท และสุดท้ายคือการซ้ำแบบเรียงต่อกันโดยตรง ซึ่งมักพบในโครงสร้างที่อธิบายว่าเป็นลูปแบบเลื่อน รูปกากบาท และ Z-DNA แบบมือซ้าย[ 14 ]
พบได้ทั่วไปในสิ่งมีชีวิตหลายชนิด
การศึกษาในอดีตชี้ให้เห็นว่าการทำซ้ำเป็นลักษณะทั่วไปของยูคาริโอตซึ่งแตกต่างจากโปรคาริโอตและอาร์เคีย [ 14 ] รายงานอื่นๆ ชี้ให้เห็นว่าถึงแม้จะมีองค์ประกอบการทำซ้ำที่ค่อนข้างน้อยในจีโนมของโปรคาริโอต แต่ก็ยังมีการทำซ้ำขนาดใหญ่หลายร้อยหรือหลายพันครั้ง[ 15 ] การวิเคราะห์จีโนมในปัจจุบันดูเหมือนจะชี้ให้เห็นถึงการมีอยู่ของการทำซ้ำแบบผกผันที่สมบูรณ์แบบจำนวนมากในจีโนมของโปรคาริโอตหลายๆ ชนิด เมื่อเทียบกับจีโนมของยูคาริโอต[ 16 ]

สำหรับการหาปริมาณและการเปรียบเทียบการทำซ้ำแบบกลับด้านระหว่างหลายสปีชีส์ โดยเฉพาะในอาร์เคีย โปรดดู[ 17 ]
การทำซ้ำแบบกลับด้านในปมเทียม
ปมเทียม (Pseudoknots)เป็นโครงสร้างโมทีฟทั่วไปที่พบใน RNA เกิดจากการเรียงซ้อนกันของก้านและห่วงโดยที่ก้านของโครงสร้างหนึ่งเกิดจากห่วงของอีกโครงสร้างหนึ่ง ปมเทียมมีโครงสร้าง การพับหลายแบบ และมีความยาวห่วงที่แตกต่างกันมาก ทำให้เป็นกลุ่มที่มีโครงสร้างหลากหลาย[ 18 ]
ลำดับซ้ำแบบกลับด้านเป็นองค์ประกอบสำคัญของปมเทียม ดังที่เห็นได้จากภาพประกอบของปมเทียมที่เกิดขึ้นตามธรรมชาติซึ่งพบในองค์ประกอบ RNA ของเทโลเมอเรสของ มนุษย์ [ 19 ]โครงสร้างนี้เกี่ยวข้องกับลำดับซ้ำแบบกลับด้านที่แตกต่างกันสี่ชุด ชุดที่ 1 และ 2 เป็นลำต้นของก้านห่วง A และเป็นส่วนหนึ่งของห่วงสำหรับก้านห่วง B ในทำนองเดียวกัน ชุดที่ 3 และ 4 เป็นลำต้นสำหรับก้านห่วง B และเป็นส่วนหนึ่งของห่วงสำหรับก้านห่วง A
ปมเทียมมีบทบาทหลายอย่างในทางชีววิทยา ปมเทียมของเทโลเมอเรสในภาพประกอบมีความสำคัญต่อกิจกรรมของเอนไซม์นั้น[ 19 ] ไรโบไซม์ของไวรัสตับอักเสบเดลต้า (HDV)พับตัวเป็นโครงสร้างปมเทียมคู่และตัดจีโนมวงกลมของตัวเองเพื่อสร้าง RNA ที่มีความยาวเท่ากับจีโนมเดียว ปมเทียมยังมีบทบาทในการเลื่อนเฟรมของไรโบโซม ตามโปรแกรม ที่พบในไวรัสบางชนิดและจำเป็นในการจำลองแบบของเรโทรไวรัส[ 18 ]
ในริโบสวิตช์
ลำดับซ้ำแบบกลับหัวมีบทบาทสำคัญในไรโบสวิตช์ซึ่งเป็นองค์ประกอบควบคุม RNA ที่ควบคุมการแสดงออกของยีนที่สร้าง mRNA ซึ่งเป็นส่วนหนึ่งขององค์ประกอบเหล่านั้น[ 10 ] ตัวอย่างที่ง่ายขึ้นของ ไรโบสวิตช์ ฟลาวินโมโนนิวคลีโอไทด์ (FMN) แสดงอยู่ในภาพประกอบ ไรโบสวิตช์นี้มีอยู่ใน ทรานสคริปต์ mRNAและมี โครงสร้าง ก้านและห่วง หลายอัน อยู่เหนือบริเวณการเข้ารหัสอย่างไรก็ตาม มีเพียงก้านและห่วงที่สำคัญเท่านั้นที่แสดงในภาพประกอบ ซึ่งถูกทำให้ง่ายขึ้นอย่างมากเพื่อช่วยแสดงบทบาทของลำดับซ้ำแบบกลับหัว มีลำดับซ้ำแบบกลับหัวหลายอันในไรโบสวิตช์นี้ ดังที่ระบุไว้ในสีเขียว (พื้นหลังสีเหลือง) และสีน้ำเงิน (พื้นหลังสีส้ม)

ในกรณีที่ไม่มี FMN โครงสร้าง Anti-termination จะเป็นโครงสร้าง ที่เหมาะสมที่สุด สำหรับ mRNA transcript โดยจะถูกสร้างขึ้นโดยการจับคู่เบสของบริเวณซ้ำแบบกลับด้านที่วงกลมสีแดงไว้ เมื่อมี FMN อยู่ FMN อาจจับกับลูปและป้องกันการก่อตัวของโครงสร้าง Anti-termination ซึ่งจะทำให้ชุดซ้ำแบบกลับด้านสองชุดที่แตกต่างกันสามารถจับคู่เบสและสร้างโครงสร้าง Termination ได้[ 20 ] ก้านลูปที่ปลาย 3' เป็นตัวยุติการถอดรหัสเนื่องจากลำดับที่อยู่ถัดจากนั้นเป็นสายของยูราซิล (U) หากก้านลูปนี้ก่อตัวขึ้น (เนื่องจากการมีอยู่ของ FMN) เมื่อสาย RNA ที่กำลังเติบโตโผล่ออกมาจาก คอมเพล็กซ์ RNA polymeraseมันจะสร้างแรงตึงโครงสร้างมากพอที่จะทำให้สาย RNA แยกตัวออกและยุติการถอดรหัส การแยกตัวเกิดขึ้นได้ง่ายเนื่องจากการจับคู่เบสระหว่าง U ใน RNA และ A ในสายแม่แบบเป็นการจับคู่เบสที่อ่อนแอที่สุด[ 10 ] ดังนั้นที่ระดับความเข้มข้นที่สูงขึ้น FMN จะควบคุมการถอดรหัสของตัวเองโดยการเพิ่มการสร้างโครงสร้างการยุติ
การกลายพันธุ์และโรค
ลำดับซ้ำแบบกลับหัวมักถูกอธิบายว่าเป็น "จุดร้อน" ของความไม่เสถียรของจีโนมยูคาริโอตและโปรคาริโอต[ 7 ]ลำดับซ้ำแบบกลับหัวที่ยาวถือว่ามีอิทธิพลอย่างมากต่อความเสถียรของจีโนมของสิ่งมีชีวิตต่างๆ[ 21 ]ตัวอย่างเช่นในE. coliซึ่งลำดับจีโนมที่มีลำดับซ้ำแบบกลับหัวที่ยาวนั้นแทบจะไม่ถูกจำลอง แต่กลับถูกลบออกอย่างรวดเร็ว[ 21 ]ในทางกลับกัน ลำดับซ้ำแบบกลับหัวที่ยาวที่พบในยีสต์ส่งเสริมการรวมตัวกันใหม่ภายในโครโมโซมเดียวกันและโครโมโซมที่อยู่ติดกันอย่างมาก ส่งผลให้อัตราการลบออกสูงมากเช่นกัน[ 21 ]สุดท้ายนี้ ยังพบอัตราการลบออกและการรวมตัวกันใหม่ที่สูงมากในบริเวณโครโมโซมของสัตว์เลี้ยงลูกด้วยนมที่มีลำดับซ้ำแบบกลับหัว[ 21 ]ความแตกต่างที่รายงานในความเสถียรของจีโนมของสิ่งมีชีวิตที่เกี่ยวข้องกันนั้นมักเป็นข้อบ่งชี้ถึงความแตกต่างในลำดับซ้ำแบบกลับหัว[ 11 ]ความไม่เสถียรเกิดจากแนวโน้มของลำดับซ้ำแบบกลับหัวที่จะพับเป็นโครงสร้าง DNA คล้ายกิ๊บติดผมหรือกากบาท โครงสร้างพิเศษเหล่านี้สามารถขัดขวางหรือทำให้การจำลอง DNA และกิจกรรมทางจีโนมอื่นๆ สับสนได้[ 7 ]ดังนั้น ลำดับซ้ำแบบกลับหัวจึงนำไปสู่การกำหนดค่าพิเศษทั้งในRNAและDNAซึ่งในที่สุดอาจทำให้เกิดการกลายพันธุ์และโรคได้[ 9 ]

ภาพประกอบแสดงให้เห็นการทำซ้ำแบบกลับด้านที่กำลังเกิดการอัดรีดแบบกากบาท DNA ในบริเวณของการทำซ้ำแบบกลับด้านจะคลายตัวออกแล้วรวมตัวกันใหม่ ก่อให้เกิดจุดเชื่อมต่อสี่ทางที่มี โครงสร้าง ก้านห่วง สองอัน โครงสร้างแบบกากบาทเกิดขึ้นเนื่องจากลำดับการทำซ้ำแบบกลับด้านจับคู่กันเองบนสายของตัวเอง[ 22 ]
กากบาทที่ยื่นออกมาสามารถนำไปสู่การกลายพันธุ์แบบเฟรมชิฟต์ได้เมื่อลำดับ DNA มีการทำซ้ำแบบกลับหัวในรูปแบบของพาลินโดรมรวมกับบริเวณที่มีการทำซ้ำโดยตรงที่ด้านใดด้านหนึ่ง ในระหว่างการถอดรหัสการลื่นไถลและการแยกตัวบางส่วนของพอลิเมอเรสออกจากสายแม่แบบสามารถนำไปสู่การกลายพันธุ์ ทั้งแบบ การลบและการแทรกได้[ 9 ] การลบเกิดขึ้นเมื่อส่วนหนึ่งของ สายแม่แบบ ที่คลายเกลียวก่อตัวเป็นก้านห่วงซึ่งจะถูก "ข้าม" โดยกลไกการถอดรหัส การแทรกเกิดขึ้นเมื่อก้านห่วงก่อตัวขึ้นในส่วนที่แยกตัวของสายที่เกิดขึ้นใหม่ (สังเคราะห์ขึ้นใหม่) ทำให้ส่วนหนึ่งของสายแม่แบบถูกถอดรหัสสองครั้ง[ 9 ]
ภาวะขาดแอนติทรอมบินเนื่องจากการกลายพันธุ์แบบจุด
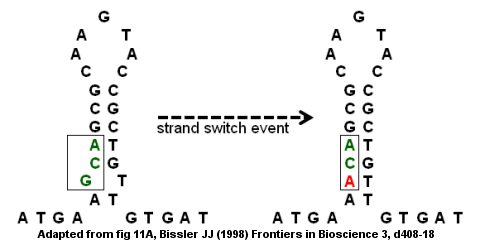
ลำดับซ้ำแบบกลับหัวที่ไม่สมบูรณ์อาจนำไปสู่การกลายพันธุ์ผ่านการสลับสายภายในและระหว่างสาย[ 9 ] บริเวณการเข้ารหัสของ ยีนแอนติทรอมบิน III เป็นตัวอย่างของลำดับซ้ำแบบกลับหัวที่ไม่สมบูรณ์ดังแสดงในรูปทางด้านขวา โครงสร้าง ก้านห่วงเกิดขึ้นโดยมีส่วนนูนที่ด้านล่างเนื่องจาก G และ T ไม่จับคู่กัน เหตุการณ์การสลับสายอาจส่งผลให้ G (ในส่วนนูน) ถูกแทนที่ด้วย A ซึ่งจะขจัด "ความไม่สมบูรณ์" ในลำดับซ้ำแบบกลับหัวและให้โครงสร้างก้านห่วงที่แข็งแรงขึ้น อย่างไรก็ตาม การแทนที่ยังสร้างการกลายพันธุ์แบบจุดที่เปลี่ยนรหัส GCA เป็น ACA หากเหตุการณ์การสลับสายตามมาด้วยการจำลองแบบ DNA รอบที่สอง การกลายพันธุ์อาจคงที่ในจีโนมและนำไปสู่โรค โดยเฉพาะอย่างยิ่งการกลายพันธุ์แบบมิสเซนส์จะนำไปสู่ยีนที่บกพร่องและการขาดแอนติทรอมบิน ซึ่งอาจส่งผลให้เกิดภาวะลิ่มเลือดอุดตันในหลอดเลือดดำ (ลิ่มเลือดภายในหลอดเลือดดำ) [ 9 ]

โรคกระดูกเปราะจากการกลายพันธุ์แบบเฟรมชิฟต์
การกลายพันธุ์ใน ยีน คอลลาเจนสามารถนำไปสู่โรค กระดูกเปราะ (Osteogenesis Imperfecta ) ซึ่งมีลักษณะเฉพาะคือกระดูกเปราะ[ 9 ]ในภาพประกอบ ก้านห่วงที่เกิดจากการทำซ้ำแบบกลับด้านที่ไม่สมบูรณ์จะกลายพันธุ์ด้วยการแทรกนิวคลีโอไทด์ไทมีน (T) อันเป็นผลมาจากการสลับระหว่างสายหรือภายในสาย การเพิ่ม T จะสร้างการ จับ คู่เบส "ที่ตรงกัน" กับอะดีนีน (A) ซึ่งก่อนหน้านี้เป็น "ส่วนนูน" ทางด้านซ้ายของก้าน แม้ว่าการเพิ่มนี้จะทำให้ก้านแข็งแรงขึ้นและทำให้การทำซ้ำแบบกลับด้านสมบูรณ์ขึ้น แต่ก็ยังสร้างการกลายพันธุ์แบบเลื่อนเฟรมในลำดับนิวคลีโอไทด์ซึ่งเปลี่ยนแปลงเฟรมการอ่านและจะส่งผลให้มีการแสดงออกของยีนที่ไม่ถูกต้อง[ 9 ]
โปรแกรมและฐานข้อมูล
รายการต่อไปนี้ให้ข้อมูลและลิงก์ภายนอกไปยังโปรแกรมและฐานข้อมูลต่างๆ สำหรับลำดับซ้ำแบบกลับด้าน:
- non-B DBฐานข้อมูลสำหรับคำอธิบายประกอบแบบบูรณาการและการวิเคราะห์โมทีฟการสร้าง DNA ที่ไม่ใช่ B [ 23 ] ฐานข้อมูลนี้จัดทำโดยศูนย์การคำนวณชีวการแพทย์ขั้นสูง (ABCC) ที่ห้องปฏิบัติการวิจัยมะเร็งแห่งชาติเฟรเดอริค (FNLCR) ฐานข้อมูลนี้ครอบคลุม โครงสร้าง A-DNAและZ-DNAหรือที่รู้จักกันในชื่อ "DNA ที่ไม่ใช่ B" เนื่องจากไม่ใช่ รูปแบบ B-DNA ที่พบได้ทั่วไป ของ เกลียวคู่ Watson-Crick แบบมือขวา"DNA ที่ไม่ใช่ B" เหล่านี้รวมถึง Z-DNA แบบมือซ้าย, รูปทรงกากบาท , ไตรเพล็กซ์, เตตระเพล็กซ์ และโครงสร้างแฮร์พิน[ 23 ]สามารถทำการค้นหาได้กับ "ประเภทการทำซ้ำ" ที่หลากหลาย (รวมถึงการทำซ้ำแบบกลับด้าน) และกับหลายสายพันธุ์
- ฐานข้อมูลลำดับซ้ำแบบกลับด้าน เก็บถาวรเมื่อวันที่ 1 กันยายน 2020 ที่Wayback Machineมหาวิทยาลัยบอสตัน ฐานข้อมูลนี้เป็นแอปพลิเคชันบนเว็บที่อนุญาตให้สอบถามและวิเคราะห์ลำดับซ้ำที่เก็บไว้ในโครงการฐานข้อมูลสาธารณะ นักวิทยาศาสตร์ยังสามารถวิเคราะห์ลำดับของตนเองด้วยอัลกอริทึม Inverted Repeats Finder ได้อีกด้วย[ 24 ]
- P-MITE: ฐานข้อมูล Plant MITE — ฐานข้อมูล Miniature Inverted-repeat Transposable Elements (MITEs) นี้ ประกอบด้วยลำดับจาก จีโนมของพืชสามารถค้นหาหรือดาวน์โหลดลำดับจากฐานข้อมูลได้[ 25 ]
- EMBOSSคือ "ชุดซอฟต์แวร์เปิดชีววิทยาโมเลกุลของยุโรป" ซึ่งทำงานบน ระบบ ปฏิบัติการ UNIXและระบบปฏิบัติการที่คล้าย UNIX [ 26 ]เอกสารและไฟล์ซอร์สโค้ดของโปรแกรมมีให้ใช้งานบนเว็บไซต์ EMBOSSแอปพลิเคชันที่เกี่ยวข้องกับการทำซ้ำแบบกลับด้านโดยเฉพาะมีดังต่อไปนี้:
- EMBOSS einverted : ค้นหาการทำซ้ำแบบกลับด้านในลำดับนิวคลีโอไทด์สามารถตั้งค่าเกณฑ์เพื่อจำกัดขอบเขตของการค้นหาได้[ 26 ]
- EMBOSS พาลินโดรม : ค้นหาพาลินโดรมเช่น บริเวณ ก้านห่วงในลำดับนิวคลีโอไทด์ โปรแกรมจะค้นหาลำดับที่มีส่วนของความไม่ตรงกันและช่องว่างที่อาจสอดคล้องกับส่วนที่โป่งออกมาในก้านห่วง[ 26 ]
ลิงก์ภายนอก
- Inverted+Repeat+Sequence ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การทำซ้ำแบบกลับด้าน
ลำดับ นิวคลีโอไทด์ แบบกลับหัว (หรือ IR ) คือลำดับนิว คลีโอไทด์ สายเดี่ยวที่ตามด้วย ลำดับนิวคลีโอ ไทด์ผกผันที่อยู่ถัด ไป [ 1 ]...
เทียบกับการทำซ้ำโดยตรง
การทำซ้ำโดยตรงเกิดขึ้นเมื่อลำดับถูกทำซ้ำด้วยรูปแบบเดียวกันที่ปลายน้ำ [ 1 ] ไม่มีการผกผันและไม่มีส่วนเติมเต็มย้อนกลับที่เกี่ยวข้องกับการทำซ้ำโดยตรง ลำดับนิวคลีโอไทด์ที่เขียนด้วยตัวหนาแสดงถึงลำดับที่ทำซ้ำ อาจมีหรือไม่มีนิวคลีโอไทด์คั่นกลางก็ได้
เทียบกับการทำซ้ำแบบคู่ขนาน
ลำดับนิวคลีโอไทด์ที่ซ้ำกันโดยตรงโดย ไม่มี นิวคลีโอไทด์คั่นระหว่างลำดับเริ่มต้นและสำเนาถัดไปเรียกว่า ลำดับซ้ำแบบ แทนเดม (Tandem repeat ) ลำดับนิวคลีโอไทด์ที่เขียนด้วยตัวหนาแสดงถึงลำดับที่ซ้ำกัน
เทียบกับ พาลินโดรม
ลำดับการทำซ้ำแบบกลับด้านที่ ไม่มี นิวคลีโอไทด์คั่นระหว่างลำดับเริ่มต้นและ ส่วนเติมเต็มแบบย้อนกลับ ที่อยู่ถัดไป เรียกว่า พาลินโดรม [ 1 ] ตัวอย่าง : ขั้นตอนที่ 1: เริ่มต้นด้วยการทำซ้ำแบบกลับด้าน: 5' TTACGnnnnnnCGTAA 3' ขั้นตอนที่ 2: ลบนิวคลีโอไทด์คั่น: 5'...