อ่าน 7 นาที
โพลีคริกอส
โพลีคริกอส (จากภาษากรีก "poly" - มาก และ "krikos" - วงแหวนหรือวงกลม) เป็นหนึ่งในสกุลของวงศ์ Polykrikaceae ซึ่งรวมถึงไดโนแฟลเจลเลตที่สร้างอาณานิคมเทียมแบบไม่มีเปลือกหุ้ม โพ...
โพลีคริกอส
| โพลีคริกอส | |
|---|---|
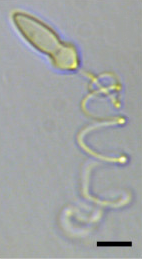 | |
| ภาพถ่ายจุลทรรศน์แสงของPolykrikos kofoidiiแสดงให้เห็นเนมาโตซิสต์ ที่ถูกขับออกมา แถบมาตราส่วน = 10 µm [ 1 ] | |
| การจำแนกทางวิทยาศาสตร์ | |
| โดเมน: | ยูคาริโอตา |
| กลุ่มสายพันธุ์ : | ซาร์ |
| กลุ่มสายพันธุ์ : | อัลวีโอลาตา |
| แผนก: | ไดโนแฟลเจลลาต้า |
| ระดับ: | ไดโนไฟซีเอ |
| คำสั่ง: | ยิมโนดินาเลส |
| ตระกูล: | โพลีคริกซี |
| ประเภท: | Polykrikos Bütschli, 1873 |

โพลีคริกอส (จากภาษากรีก "poly" - มาก และ "krikos" - วงแหวนหรือวงกลม) เป็นหนึ่งในสกุลของวงศ์ Polykrikaceaeซึ่งรวมถึงไดโนแฟลเจลเลตที่สร้างอาณานิคมเทียมแบบไม่มีเปลือกหุ้ม โพ ลีคริกอสมีลักษณะเด่นคือกลไกการยิงที่ซับซ้อน [ 2 ]ซึ่งเรียกว่าคอมเพล็กซ์เนมาโตซิสต์-เทนิโอซิสต์ ซึ่งช่วยให้สายพันธุ์ต่างๆ สามารถล่าเหยื่อได้หลากหลายชนิดพบว่าโพลีคริกอส ช่วยควบคุมการแพร่กระจายของสาหร่ายเนื่องจากพวกมันกินไดโนแฟลเจลเลตที่เป็นพิษ [ 3 ] [ 4 ]อย่างไรก็ตาม ยังมีข้อมูลบางส่วนที่ระบุว่าโพลีคริกอสเป็นพิษต่อปลา [ 5 ]
ประวัติศาสตร์แห่งความรู้
Polykrikosถูกพบเห็นครั้งแรกในปี พ.ศ. 2411 โดย Uljanin และถูกเข้าใจผิดว่าเป็นตัวอ่อนของหนอนแบนชนิดหนึ่ง ในปี พ.ศ. 2416 Butschili ได้ตรวจสอบตัวอย่างอีกครั้งและสรุปว่าเซลล์ดังกล่าวเป็นซีลิเอตที่ผิดปกติ และต่อมาในปี พ.ศ. 2424 Bergh ได้ชี้แจงความสัมพันธ์ ของ Polykrikos กับได โนแฟลเจลเลต [ 6 ]
สัณฐานวิทยา
โพลีคริกอสคือกลุ่มของซูออยด์ (หน่วยย่อยของสิ่งมีชีวิตแบบอาณานิคม) ที่ทำหน้าที่ต่างๆ ของเซลล์ทั้งเซลล์พร้อมกัน
โพลีคริกอสทุกชนิดมี: 1) ร่องตามยาวโค้งเล็กน้อย (ซัลคัส) ที่ทอดยาวไปจนถึงส่วนท้ายของสิ่งมีชีวิต 2) อะโครเบสรูปห่วง ซึ่งเป็นส่วนขยายด้านหน้าจากซัลคัส 3) ร่องตามขวาง (ซิงกูลัม) ที่มีการเคลื่อนที่ 4) คอมเพล็กซ์เทนิโอซิสต์-เนมาโตซิสต์ 5) จำนวนนิวเคลียสน้อยกว่าจำนวนซูออยด์สองหรือสี่เท่า และ 6) ความสามารถในการแยกตัวออกเป็นพсевโดโคโลนีที่มีซูออยด์น้อยลงและมีนิวเคลียสเพียงหนึ่งเดียว[ 7 ]
ลักษณะเด่นที่สุดของสกุลนี้คือการก่อตัวของกลุ่มเซลล์เทียมที่มีนิวเคลียสหลายอันซึ่งประกอบด้วยซูออยด์จำนวนคู่ซูออยด์ แต่ละตัว มีแฟลเจลลาคู่หนึ่ง (แฟลเจลลาตามขวางและตามยาว) และมีร่องตามขวางของตัวเองที่เรียกว่าซิงกูลัม แต่ร่องตามยาวของซูออยด์ที่เรียกว่าซัลซีจะรวมกัน[ 8 ]แฟลเจลลาตามขวางมีส่วนยื่นด้านข้างที่เรียกว่ามาสติโกนีมและเส้นใยลายที่พบได้ทั่วไปในไดโนแฟลเจลเลตอื่นๆ[ 9 ]บ่อยครั้งที่โพลีคริคอสมีจำนวนนิวเคลียสครึ่งหนึ่งของจำนวนซูออยด์ และซูออยด์แต่ละคู่จะใช้นิวเคลียสร่วมกัน ภายในกลุ่มนี้มีความแปรผันบ้างในออร์แกเนลล์ที่พบ แต่มีการสังเกตพบไตรโคซิสต์ เนมาโตซิสต์ เทนิโอซิสต์ มิวโคซิสต์ และพลาสติดในสมาชิกต่างๆ ภายในกลุ่มนี้[ 8 ]
ไซโตพลาซึมของPolykrikosมีลักษณะเฉพาะคือมี โครงข่าย เอนโดพลาสมิกเรติคูลัม แบบหยาบจำนวนมาก คอมเพล็กซ์กอลจิ และแวคิวโอลPolykrikosเป็นที่รู้จักกันดีว่าผลิตออร์แกเนลล์แบบขับออกได้ ซึ่งก็คือ เอ็กซ์ทรูโซม หนึ่งในนั้นคือเนมาโตซิสต์ที่เกิดขึ้นในซูออยด์ เอ็กซ์ทรูโซมอีกอันหนึ่งที่พบในสิ่งมีชีวิตคือเทนิโอซิสต์รูปแท่ง ซึ่งตั้งอยู่ด้านปลายของเนมาโตซิสต์ และก่อนหน้านี้เคยเข้าใจผิดว่าเป็นสารตั้งต้นของเนมาโตซิสต์[ 10 ]ออร์แกเนลล์เหล่านี้รวมกันเป็นคอมเพล็กซ์เทนิโอซิสต์-เนมาโตซิสต์ ซึ่งคิดว่าเป็นซินอะโพมอร์ฟี ที่ดีที่สุด สำหรับกลุ่มPolykrikos [ 8 ]แวคิวโอลที่ได้มาจากกอลจิเป็นของออร์แกเนลล์ทั้งสองชนิดร่วมกัน และจัดหาโมเลกุลที่จำเป็นสำหรับการเจริญเติบโตให้แก่แต่ละชนิด พร้อมทั้งมีส่วนร่วมในการสร้างข้อต่อ NTC [ 10 ]ออร์แกเนลล์เหล่านี้ตั้งอยู่ใกล้กัน แต่ตั้งอยู่ภายในเยื่อหุ้มที่แตกต่างกัน และถูกคั่นด้วยทางเดินที่เรียกว่า "ชูต" เนมาโตซิสต์เป็นออร์แกเนลล์ขนาดใหญ่กว่าและอยู่ด้านหลังของเทนิโอซิสต์ งานวิจัยล่าสุดบางชิ้น[ 3 ]แสดงให้เห็นว่าการทำงานของออร์แกเนลล์ทั้งสองนั้นเชื่อมโยงกัน โดยเทนิโอซิสต์จะยึดเกาะกับเหยื่อ ตามด้วยการปล่อยเนมาโตซิสต์เพื่อเจาะเหยื่อ และสุดท้ายคือการดึงเหยื่อกลับมาโดยใช้เส้นใยที่อยู่ปลายเนมาโตซิสต์ใกล้กับถุงด้านหลัง ท่อที่ฝังอยู่ภายในเนมาโตซิสต์จะปล่อยไปยังเหยื่อและสันนิษฐานว่าใช้สำหรับการเจาะเหยื่อ
นอกจากนี้ Gavelis et al. [ 3 ]ได้ตรวจสอบสัณฐานวิทยาและกลไกการยิงของ NTC อย่างละเอียด ซึ่งแสดงให้เห็นว่าแตกต่างจากสัตว์กลุ่ม Cnidaria อย่างสิ้นเชิง แสดงให้เห็นว่าเนมาโตซิสต์ได้วิวัฒนาการอย่างอิสระในไดโนแฟลเจลเลตเซลล์เดียว แคปซูลท่อขดที่ห่อหุ้มนั้น ต่างจากในสัตว์กลุ่ม Cnidaria ตรงที่มันถูกปิดผนึก ซึ่งบังคับให้สไตเลตเมื่อยิงต้องเจาะแคปซูลจากภายในก่อนเพื่อปลดปล่อยเส้นใย จากนั้นจึงเจาะเหยื่อ เมื่อท่อผ่านหัวฉีด มันจะเปิดฝาปิดและคลายตัวออก การยิงในเนมาโตซิสต์ของสัตว์กลุ่ม Cnidaria เกิดจากการสังเคราะห์สารขับเคลื่อนออสโมติกโพลีแกมมากลูตาเมตซินเทส PgsAA ในขณะที่ในPolykrikosเชื่อว่าเกิดขึ้นเนื่องจากแรงดันที่เหนี่ยวนำอันเป็นผลมาจากการหดตัวของเส้นใยแคปซูลในผนังแคปซูล[ 3 ] นิวเคลียสมีลักษณะเฉพาะคือคอร์เทกซ์เส้นใยสองชั้นที่อยู่ใต้เยื่อหุ้มนิวเคลียส ที่ยื่นออกมา คอร์เทกซ์ถูกสันนิษฐานว่าให้ความแข็งแรงและรูปร่างแก่นิวเคลียส ในขณะที่การยื่นออกมาของนิวเคลียสเชื่อว่าจะเพิ่มพื้นที่แลกเปลี่ยนระหว่างนิวเคลียสและไซโตพลาสซึมที่บริเวณรูพรุนของคอร์เทกซ์[ 9 ]
ในPolykrikosริบบิ้นเส้นใยที่กำหนดไว้อย่างดีมีส่วนเกี่ยวข้องกับการเชื่อมต่อระหว่างนิวเคลียสและแฟลเจลลา และการยึดกับอุปกรณ์แฟลเจลลาอาจทำหน้าที่ในการวางแนวของนิวเคลียสที่สัมพันธ์กับแฟลเจลลาในระหว่างกระบวนการเคลื่อนที่ ไมโทซิส และการแบ่งเซลล์[ 9 ]
พลาสติด
มีความหลากหลายในการรับสารอาหารในส ปีชีส์ของ Polykrikosเนื่องจากบางชนิดพึ่งพาการสังเคราะห์แสงเพียงอย่างเดียว บางชนิดเป็นมิกโซโทรฟ (mixothroph) ในขณะที่บางชนิดเป็นเฮเทอโรโทรฟ (heterotroph) อย่างสมบูรณ์ ทำให้Polykrikosเป็นกลุ่มที่มีประโยชน์ในการศึกษาการวิวัฒนาการของออร์แกเนลล์Polykrikos geminatumและP. hartmanii ซึ่งเป็นโพลีคริคอยด์ที่แตกแขนงออกมาในช่วงต้น มีพลาสติดสามเยื่อหุ้มพร้อมไทลาคอยด์เรียงซ้อนกันสามชั้น ซึ่งบ่งชี้ถึงพลาสติดชนิดเพอริดินินทุติยภูมิที่พบได้ทั่วไปในไดโนแฟลเจลเลต อย่างไรก็ตามP. lebouriaeซึ่งเป็นมิกโซโทรฟ ซึ่งอยู่ในกลุ่มโพลีคริคอยด์เฮเทอโรโทรฟ มีพลาสติดที่ผิดปกติจากไดโนแฟลเจลเลต มันมีเยื่อหุ้มสองชั้นและมีไทลาคอยด์เรียงซ้อนกันสองชั้นซึ่งพบในไดอะตอมและแฮปโทไฟต์ อย่างไรก็ตาม การวิเคราะห์ข้อมูลโมเลกุลโดย Gavelis et al. [ 11 ]ได้แสดงให้เห็นว่าP. lebouriaeมีพลาสติดชนิดเพอริดินินซึ่งน่าจะได้รับมาจากโพลีคริคอยด์บรรพบุรุษ การวิเคราะห์ทรานสคริปโตมิกส์แสดงให้เห็นเหตุการณ์การสูญเสียหลายครั้งในโพลีคริคอยด์ซึ่งอาจอธิบายได้จากมุมมองด้านพลังงานและข้อจำกัดทางสรีรวิทยา การเพิ่มขนาดของโพลีคริคอสอย่างค่อยเป็นค่อยไปทำให้สิ่งมีชีวิตเหล่านี้เป็นผู้ล่าที่ยอดเยี่ยม แต่การเพิ่มขนาดของเซลล์เดี่ยวเป็นที่ทราบกันดีว่าส่งผลให้คลอโรพลาสต์ถูกบังแสงมากขึ้น และอัตราส่วนพื้นที่ผิวต่อปริมาตรลดลง ส่งผลให้ประสิทธิภาพการสังเคราะห์แสงลดลง การมี NTC ที่เฉพาะเจาะจงและขนาดเซลล์ที่ใหญ่อาจกระตุ้นให้เกิดการสูญเสียการสังเคราะห์แสงหลายครั้ง[ 11 ]
วงจรชีวิตของP. kofoidii
สำหรับสกุลPolykrikosมีข้อมูลโดยละเอียดเกี่ยวกับการสืบพันธุ์ของชนิดต้นแบบ (holotype) P. kofoidii ซึ่งวงจรชีวิตคล้ายกับวงจรของไดโนแฟลเจลเลตทั่วไป โดยเซลล์สืบพันธุ์จะสร้างแกมีตที่รวมตัวกันเพื่อสร้างไซโกตแบบดิพลอยด์ (2n) ที่สามารถเกิดซีสต์ได้ แต่ลักษณะแบบพсевโดโคโลเนีย ลทำให้ การพัฒนาของ Polykrikosมีลักษณะเฉพาะหลายประการ[ 12 ]
เมื่อสิ่งมีชีวิตได้รับอาหารอย่างเพียงพอ พวกมันจะปรากฏเป็นกลุ่มเซลล์เทียมที่มี 4 ซูออยด์และ 2 นิวเคลียส และในระหว่างการสืบพันธุ์แบบไม่อาศัยเพศ จำนวนซูออยด์จะเพิ่มขึ้นเป็นสองเท่า ตามด้วยการแบ่งนิวเคลียส ทำให้เกิดระยะ 8 ซูออยด์และ 4 นิวเคลียส จากนั้นจึงแบ่งตัวแบบไบนารีตามขวางต่อไปเป็น โพลีคริ คอส 2 ตัว ที่มี 4 ซูออยด์และ 2 นิวเคลียส การสร้างเซลล์สืบพันธุ์มีความพิเศษ เนื่องจากกลุ่มเซลล์เทียมสร้างเซลล์สืบพันธุ์ 4 เซลล์ที่มีขนาดและรูปร่างแตกต่างจากเซลล์สืบพันธุ์แบบไม่อาศัยเพศ รูปแบบการสืบพันธุ์แบบไม่อาศัยเพศจะเพิ่มจำนวนซูออยด์เป็นสองเท่าและต่อมาแบ่งออกเป็นเซลล์สืบพันธุ์ 4 เซลล์ ในรูปแบบ 2 ซูออยด์และ 1 นิวเคลียส เซลล์สืบพันธุ์สองเซลล์จะจับคู่กันโดยใช้ด้านท้องและรวมตัวกันเป็นพลาโนไซโกต สำหรับP. kofoidiiพบโครงสร้างรูปนิ้วมือสองอันในเซลล์สืบพันธุ์ ซึ่งคาดว่าเกี่ยวข้องกับการสัมผัสและการรวมตัวของเซลล์สืบพันธุ์ แต่จำเป็นต้องมีข้อมูลเพิ่มเติมเพื่อยืนยันเรื่องนี้ เซลล์สืบพันธุ์ที่หลอมรวมกันทางด้านท้องต้องอาศัยการจัดเรียงใหม่ที่ซับซ้อนของแฟลเจลลาทั้งแปดเส้นและการก่อตัวของร่องและส่วนโค้ง พลาโนไซโกตแบบ 4 ซูออยด์มีนิวเคลียสเพียงหนึ่งเดียวและมีเส้นทางการพัฒนาสองเส้นทางขึ้นอยู่กับปริมาณอาหาร ภายใต้สภาวะอดอาหาร พลาโนไซโกตจะแตกตัวเป็น 2 ซูออยด์สองตัว โดยตัวหนึ่งขาดนิวเคลียส แต่ไม่ได้มีการตรวจสอบชะตากรรมต่อไป ภายใต้สภาวะการเพาะเลี้ยง สิ่งมีชีวิตส่วนใหญ่จะผ่านกระบวนการไมโอซิสและเข้าสู่ระยะเจริญเติบโตโดยตรง มีพลาโนไซโกตเพียงไม่กี่ตัวเท่านั้นที่ผ่านระยะซีสต์พักตัว ระยะซีสต์คงอยู่เป็นเวลา 1 เดือน ซึ่งถือว่าเป็นช่วงเวลาที่ค่อนข้างสั้นเมื่อเทียบกับไดโนแฟลเจลเลตชนิดอื่น ๆ ซึ่งมีระยะพักตัวที่จำเป็นอาจนานถึง 6 เดือน ระยะเวลาของการสร้างซีสต์มีความสัมพันธ์กับกลยุทธ์การหาอาหารเชิงนิเวศ ระยะพักตัวที่สั้นอาจช่วยให้การเปลี่ยนผ่านระหว่างระยะต่าง ๆ ของวงจรชีวิตเป็นไปอย่างรวดเร็ว ซึ่งอาจเป็นประโยชน์ต่อสิ่งมีชีวิตที่กินสิ่งมีชีวิตอื่นเป็นอาหารในกรณีที่ปริมาณอาหารผันผวน อย่างไรก็ตาม ความเป็นไปได้ของการส่งสัญญาณทางเคมีที่เกี่ยวข้องกับการฟักตัวของซีสต์ในPolykrikosยังคงต้องได้รับการตรวจสอบต่อไป
การงอกของเซลล์ซูออยด์เดี่ยวที่โผล่ออกมาจากซีสต์นั้นมีการพัฒนาที่ไม่เหมือนใคร ซึ่งไม่เคยมีการบันทึกไว้สำหรับไดโนแฟลเจลเลตที่ดำรงชีวิตอิสระใดๆ มาก่อน สัณฐานวิทยาของมันเปลี่ยนจาก 1 ซูออยด์ 1 นิวเคลียส ไปเป็น 2 ซูออยด์ 1 นิวเคลียส และ 4 ซูออยด์ 1 นิวเคลียส ไปสู่ระยะ 4 ซูออยด์ 2 นิวเคลียส ข้อมูลนี้ทำให้เกิดคำถามว่ารูปแบบการฟักตัวดังกล่าวอาจสะท้อนถึงวิวัฒนาการของพсевโดโคโลนีของโพลี คริกอสหรือไม่ [ 12 ]
ถิ่นที่อยู่และระบบนิเวศ (แหล่งที่อยู่อาศัย, อาหาร)
โพลีคริกอสทุกชนิดที่รู้จักพบได้ในสภาพแวดล้อมทางทะเล ส่วนใหญ่เป็นแพลงก์ตอน ยกเว้นโพลีคริกอส เลบูเรียและโพลีคริกอส เฮิร์ดมาเนที่พบในแหล่งที่อยู่อาศัยใต้ทะเล[ 7 ] นอกจากนี้ยังมีความหลากหลายในระบบนิเวศการกินอาหาร เนื่องจากบางชนิดมีพลาสติดและสามารถใช้การสังเคราะห์แสงเพื่อรับสารอาหาร แต่มักจะเป็นมิกโซโทรฟ ( P. barnegatensis, P. lebouriae, P. tanit, P. hartmannii ) มีเพียงชนิดเดียวที่ทราบว่าเป็นออโตโทรฟโดยเฉพาะ ( P. geminatum ) ในขณะที่บางชนิดไม่มีพลาสติดและเป็นเฮเทอโรโทรฟโดยสมบูรณ์ ( P. grassei, P. herdmanae, P. kofoidiiและP. schwartzii ) [ 13 ] [ 8 ]
Polykrikos hartmanniiเป็นไดโนแฟลเจลเลตที่สังเคราะห์แสงได้ และมีรายงานพบในน่านน้ำของแคนาดา[ 14 ]สหรัฐอเมริกา เม็กซิโก จีน อินเดีย ญี่ปุ่น และเกาหลี[ 5 ] [ 13 ] P. hartmaniiกินสาหร่ายโดยการกลืนกินหลังจากยึดเหยื่อโดยใช้คอมเพล็กซ์เนมาโตซิสต์-เทนิโอซิสต์ (ต่อมาเรียกว่า NTC) [ 13 ] Tang et al. [ 5 ]สังเกตเห็นการแพร่กระจายของP. hartmannii ที่ทำให้ปลาไซ ปริโนดอน วาริเอเกตส์ (sheepshead minnows) วัยอ่อนตาย 100% ภายใน 24 ชั่วโมง ซึ่งบ่งชี้ว่าP. hartmanniiเป็นสาหร่ายที่เป็นพิษต่อปลาและเป็นอันตรายP.hartmanniiกินไดโนแฟลเจลเลตที่สร้างสายโซ่Cochlodinium polykrikoidesและGymnodinium catenatumซึ่งเป็นที่ทราบกันดีว่าทำให้ปลาตายได้ ดังนั้นจึงเชื่อว่าP. hartmannii มีเอนไซม์ที่ล้างพิษที่ผลิตโดยไดโนแฟลเจลเลตที่เป็นเหยื่อเหล่านี้ [ 13 ]นอกจากนี้ การเปรียบเทียบการกินอาหารของPolykrikos สาม ชนิดเผยให้เห็นว่าชนิดต่างๆ มีความแตกต่างกันในความชอบเหยื่อ และบางชนิดมีความเชี่ยวชาญมากกว่าชนิดอื่นๆ เช่น การล่าเหยื่อของ P. hartmaniiมีความหลากหลายน้อยกว่า (กินเหยื่อ 2 ชนิด) เมื่อเทียบกับP. kofoidiiและP. lebouriaeซึ่งกินสาหร่าย 14 ชนิดที่แตกต่างกัน[ 13 ]
การล่าเหยื่อโดยPolykrikos ที่เป็นเฮเทอโรโทรฟิก กลายเป็นหัวข้อที่น่าสนใจอย่างมาก เนื่องจากสิ่งมีชีวิตบางชนิดกินไดโนแฟลเจลเลตที่ทำให้เกิดการระบาดของสารพิษ มีรายงานผลกระทบจากการล่าเหยื่อสูงของPolykrikos schwartzii Butschili ต่อไดโนแฟลเจลเลตที่เป็นพิษAlexandrium tamarense (Lebour) Balech ในอาร์เจนตินา ในขณะที่Polykrikos kofoidii Chatton ควบคุมGymnodium catenatum Graham ในน่านน้ำชายฝั่งของโปรตุเกสและญี่ปุ่น[ 4 ] G. catenatumเป็นหนึ่งในสายพันธุ์ที่ทำให้เกิดพิษจากหอยที่เป็นอัมพาต (PSP) และพบได้ในน่านน้ำของออสเตรเลีย ญี่ปุ่น เม็กซิโก และสเปน[ 15 ] การสังเกตพฤติกรรมการกินอาหารชี้ให้เห็นว่าP. kofoidiiเริ่มแรกแสดงพฤติกรรมการว่ายน้ำวนเป็นวงกลมในระยะใกล้กับเหยื่อ ตามด้วยการปล่อยเนมาโตซิสต์ ดึงเหยื่อเข้าไปในร่างกายผ่านร่องด้านหลัง และสุดท้ายก็กลืนกินเหยื่อ[ 16 ] PSP ก่อให้เกิดความกังวลด้านสังคม เศรษฐกิจ และสิ่งแวดล้อม เนื่องจากส่งผลกระทบต่อสุขภาพของทั้งสัตว์เลี้ยงลูกด้วยนมในทะเลและมนุษย์ และกลไกการควบคุมประชากรสาหร่ายขนาดเล็กที่เป็นพิษโดยP.kofoidiiอาจมีความสำคัญในการตรวจสอบสิ่งแวดล้อมและการกำจัดอันตรายต่อสุขภาพ
ความสำคัญในทางปฏิบัติ
การล่าสาหร่ายขนาดเล็กที่เป็นพิษโดยไดโนแฟลเจลเลตแบบเฮเทอโรโทรฟิกเป็นหนึ่งในปัจจัยที่ควบคุมการแพร่กระจายของสาหร่าย โพลีคริโคเป็นที่รู้จักกันดีว่าสามารถควบคุมประชากรของได โนแฟลเจลเลต เช่น Alexandrium tamarense [ 3 ] [ 4 ] และG. catenatum [ 17 ]ซึ่งเป็นตัวการสำคัญในการแพร่กระจายของสาหร่ายที่เป็นพิษ โพลีคริโคแบบเฮเทอโรโทรฟิกเหล่านี้ไม่เพียงแต่จะลดระดับความเป็นพิษที่เกิดจากเหยื่อในห่วงโซ่อาหารทางทะเลเท่านั้น แต่ยังสามารถหยุดการแพร่กระจายของสาหร่ายที่เป็นพิษและสามารถนำมาใช้ในการบำบัดทางชีวภาพได้[ 17 ]ดังนั้น การลดความเป็นพิษของน้ำอาจช่วยควบคุมความสมดุลของห่วงโซ่อาหารทางทะเลและลดอัตราการตายของปลา สัตว์เลี้ยงลูกด้วยนมในทะเล และนกทะเล การศึกษาเพิ่มเติมเกี่ยวกับกลไกทางโมเลกุลของการล้างพิษโดยโพลีคริโคอาจเป็นประโยชน์ในด้านชีวการแพทย์และการเฝ้าระวังสิ่งแวดล้อม
อย่างไรก็ตามโพลิคริคอส บางชนิด ก่อให้เกิดความเสี่ยงต่อสุขภาพของปลาบางชนิด ในขณะที่โพลิคริคอสที่ควบคุมการแพร่กระจายมักถูกล่าโดยสัตว์ไม่มีกระดูกสันหลังในทะเล เช่นแอมฟิพอดซึ่งจะนำสารพิษกลับเข้าสู่ห่วงโซ่อาหาร[ 17 ]การเฝ้าระวังประชากรโพลิคริคอสบางชนิดและการตรวจสอบพลวัตของสารพิษภายในร่างกายของสัตว์กินพืชอาจช่วยให้เข้าใจห่วงโซ่อาหารของแพลงก์ตอนได้ดีขึ้น ประเมินระดับความเป็นพิษในระบบนิเวศ และเสนอแนวทางการกำจัดสารพิษที่เป็นไปได้
รายชื่อสายพันธุ์
ปัจจุบันมีการยอมรับทางอนุกรมวิธานของ 10 ชนิดดังต่อไปนี้: [ 18 ]
- Polykrikos barnegatensis GWMartin,
- Polykrikos geminatus (F.Schütt) DXQiu และ Senjie Lin
- Polykrikos grassei Lecal,
- โพลีคริคอส ฮาร์ทมันนี ดับเบิลยู.ซิมเมอร์มันน์,
- Polykrikos herdmaniae Hoppenrath & Leander,
- Polykrikos kofoidii Chatton,
- Polykrikos lebouriae Herdman,
- Polykrikos schwartzii Bütschli,
- Polykrikos tanit Reñé,
- Polykrikos tentaculatus O.Wetzel
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ โพลีคริกอส
โพลีคริกอส (จากภาษากรีก "poly" - มาก และ "krikos" - วงแหวนหรือวงกลม) เป็นหนึ่งในสกุลของวงศ์ Polykrikaceae ซึ่งรวมถึงไดโนแฟลเจลเลตที่สร้างอาณานิคมเทียมแบบไม่มีเปลือกหุ้ม โพ...
ประวัติศาสตร์แห่งความรู้
Polykrikos ถูกพบเห็นครั้งแรกในปี พ.ศ. 2411 โดย Uljanin และถูกเข้าใจผิดว่าเป็นตัวอ่อนของหนอนแบนชนิดหนึ่ง ในปี พ.ศ. 2416 Butschili ได้ตรวจสอบตัวอย่างอีกครั้งและสรุปว่าเซลล์ดังกล่าวเป็นซีลิเอตที่ผิดปกติ และต่อมาในปี พ.ศ.
สัณฐานวิทยา
โพลีคริกอส คือกลุ่มของซูออยด์ (หน่วยย่อยของสิ่งมีชีวิตแบบอาณานิคม) ที่ทำหน้าที่ต่างๆ ของเซลล์ทั้งเซลล์พร้อมกัน
พลาสติด
มีความหลากหลายในการรับสารอาหารในส ปีชีส์ของ Polykrikos เนื่องจากบางชนิดพึ่งพาการสังเคราะห์แสงเพียงอย่างเดียว บางชนิดเป็นมิกโซโทรฟ (mixothroph) ในขณะที่บางชนิดเป็นเฮเทอโรโทรฟ (heterotroph) อย่างสมบูรณ์ ทำให้ Polykrikos...