อ่าน 5 นาที
การเมทิลเลชันของโปรตีน
การเติมหมู่เมทิลให้กับโปรตีน เป็นรูปแบบหนึ่งของ การดัดแปลงหลังการแปลรหัส ซึ่งมีลักษณะเป็นการเพิ่ม หมู่เมทิล ให้กับ โปรตีน กระบวนการ นี้สามารถเกิดขึ้นได้ที่โซ่ข้างที่มีไนโตรเจนของ...
การเมทิลเลชันของโปรตีน
การเติมหมู่เมทิลให้กับโปรตีนเป็นรูปแบบหนึ่งของการดัดแปลงหลังการแปลรหัสซึ่งมีลักษณะเป็นการเพิ่มหมู่เมทิลให้กับโปรตีน กระบวนการ นี้สามารถเกิดขึ้นได้ที่โซ่ข้างที่มีไนโตรเจนของอาร์จินีนและไลซีน [ 1 ] [ 2 ]แต่ยังเกิดขึ้นที่ปลายอะมิโนและคาร์บอกซีของโปรตีนหลายชนิดด้วย ในทางชีววิทยาเอนไซม์เมทิลทรานส เฟอเร สจะเร่งปฏิกิริยาการเติมหมู่เมทิล โดยส่วนใหญ่จะถูกกระตุ้นด้วยS-adenosylmethionine การเติมหมู่เมทิลให้กับโปรตีนได้รับการศึกษามากที่สุดในฮิสโตนซึ่งการถ่ายโอนหมู่เมทิลจาก S-adenosylmethionine จะถูกเร่งปฏิกิริยาโดยเอนไซม์ฮิสโตนเมทิลทรานสเฟอเรส ฮิ สโตนที่ถูกเติมหมู่เมทิลที่สารตกค้างบางชนิดสามารถทำหน้าที่ทางเอพิเจเนติกส์เพื่อยับยั้งหรือกระตุ้นการแสดงออกของยีนได้[ 3 ] [ 4 ]
การเติมหมู่เมทิลโดยสารตั้งต้น
โปรตีนสามารถถูกเติมหมู่เมทิลได้หลายตำแหน่ง สำหรับการเติมหมู่เมทิลบางประเภท เช่น การเติมหมู่เมทิลที่ปลาย N-terminus และการเติมหมู่เมทิลที่พรีนิลซิสเทอีน จำเป็นต้องมีการประมวลผลเพิ่มเติม ในขณะที่การเติมหมู่เมทิลประเภทอื่น ๆ เช่น การเติมหมู่เมทิลที่อาร์จินีนและการเติมหมู่เมทิลที่ไลซีน ไม่จำเป็นต้องมีการประมวลผลล่วงหน้า
อาร์จินีน

อาร์จินีนสามารถถูกเมทิลเลชันได้หนึ่งครั้ง ( เมทิลอาร์จินีน ) หรือสองครั้ง (ไดเมทิลเลตอาร์จินีน) การเมทิลเลชันของหมู่อาร์จินีนถูกเร่งปฏิกิริยาโดยโปรตีนอาร์จินีนเมทิลทรานสเฟอเรส (PRMT) สามประเภทที่แตกต่างกัน: PRMT ประเภท I (PRMT1, PRMT2, PRMT3, PRMT4, PRMT6 และ PRMT8) จะติดหมู่เมทิลสองหมู่เข้ากับอะตอมไนโตรเจนปลายสุดเพียงอะตอมเดียว ทำให้เกิดไดเมทิลอาร์จินีนแบบไม่สมมาตร (N G,N G-ไดเมทิลอาร์จินีน) ในทางตรงกันข้าม PRMT ประเภท II (PRMT5 และ PRMT9) จะเร่งปฏิกิริยาการสร้างไดเมทิลอาร์จินีนแบบสมมาตรโดยมีหมู่เมทิลหนึ่งหมู่บนอะตอมไนโตรเจนปลายสุดแต่ละอะตอม (N G,N' G-ไดเมทิลอาร์จินีนแบบสมมาตร) ทั้ง PRMT ประเภท I และ II สร้างสารตัวกลาง N G-โมโนเมทิลอาร์จินีน; PRMT7 ซึ่งเป็น PRMT ประเภท III เพียงตัวเดียวที่รู้จัก สร้างเฉพาะอาร์จินีนที่ถูกเมทิลเลชันเพียงอะตอมเดียว[ 5 ]
การเติมหมู่เมทิลให้กับอาร์จินีนมักเกิดขึ้นที่บริเวณที่มีไกลซีนและอาร์จินีนมาก ซึ่งเรียกว่า "โมทีฟ GAR" [ 6 ]ซึ่งน่าจะเป็นผลมาจากความยืดหยุ่นที่เพิ่มขึ้นของบริเวณเหล่านี้ ทำให้สามารถแทรกอาร์จินีนเข้าไปในบริเวณออกฤทธิ์ ของ PRMT ได้ อย่างไรก็ตาม PRMT ที่มีลำดับคอนเซนซัสที่ไม่ใช่ GAR ก็มีอยู่เช่นกัน[ 5 ] PRMT มีอยู่ในนิวเคลียสและไซโตพลาสซึม ในการโต้ตอบของโปรตีนกับกรดนิวคลีอิก หมู่ของอาร์จินีนเป็นตัวให้ พันธะไฮโดรเจนที่สำคัญสำหรับโครงสร้างฟอสเฟต โปรตีนที่มีการเติมหมู่เมทิลให้กับอาร์จินีนจำนวนมากพบว่ามีปฏิสัมพันธ์กับ DNA หรือ RNA [ 6 ] [ 7 ]
เอนไซม์ที่อำนวยความสะดวกใน การอะเซทิเลชัน ของฮิสโตนรวมถึงฮิสโตนเอง สามารถถูกเมทิลเลชันด้วยอาร์จินีนได้ การเมทิลเลชันของอาร์จินีนส่งผลต่อปฏิสัมพันธ์ระหว่างโปรตีน และมีส่วนเกี่ยวข้องในกระบวนการต่างๆ ของเซลล์ รวมถึงการลำเลียงโปรตีน การส่งสัญญาณ และการควบคุมการถอดรหัส[ 6 ]ในด้านเอพิเจเนติกส์ การเมทิลเลชันของอาร์จินีนในฮิสโตน H3 และ H4 เกี่ยวข้องกับโครงสร้างโครมาตินที่เข้าถึงได้ง่ายขึ้น และส่งผลให้ระดับการถอดรหัสสูงขึ้น การมีอยู่ของอาร์จินีนดีเมทิเลสที่สามารถย้อนกลับการเมทิลเลชันของอาร์จินีนได้นั้นยังเป็นที่ถกเถียงกันอยู่[ 5 ]
ไลซีน
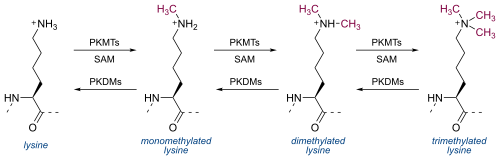
ไลซีนสามารถถูกเมทิลเลชันได้หนึ่งครั้ง สองครั้ง หรือสามครั้งโดยไลซีนเมทิลทรานสเฟอเรส (PKMTs) [ 8 ]ไลซีนเมทิลทรานสเฟอเรสส่วนใหญ่มีโดเมน SET ที่ได้รับการอนุรักษ์ทางวิวัฒนาการ ซึ่งมี กิจกรรมเมทิลทรานสเฟอเรสที่ขึ้นอยู่กับ S-adenosylmethionineแต่มีโครงสร้างที่แตกต่างจากโปรตีนที่จับกับ S-adenosylmethionine อื่นๆ การเมทิลเลชันของไลซีนมีบทบาทสำคัญในวิธีที่ฮิสโตนโต้ตอบกับโปรตีน[ 9 ]การเมทิลเลชันของไลซีนสามารถย้อนกลับได้โดยไลซีนดีเมทิลเลส (PKDMs) [ 8 ]
โปรตีนที่มีโดเมน SET ต่างกันจะมีคุณสมบัติเฉพาะในการจับกับสารตั้งต้นที่แตกต่างกัน ตัวอย่างเช่น SET1, SET7 และ MLL จะเติมหมู่เมทิลที่ไลซีน 4 ของฮิสโตน H3 ในขณะที่ Suv39h1, ESET และ G9a จะเติมหมู่เมทิลที่ไลซีน 9 ของฮิสโตน H3 โดยเฉพาะ การเติมหมู่เมทิลที่ไลซีน 4 และไลซีน 9 นั้นเกิดขึ้นแยกกัน และผลทางด้านเอพิเจเนติกส์ของการเติมหมู่เมทิลเฉพาะตำแหน่งนั้นตรงกันข้ามกันอย่างสิ้นเชิง กล่าวคือ การเติมหมู่เมทิลที่ไลซีน 4 สัมพันธ์กับสถานะการถอดรหัสที่ทำงานอยู่ ในขณะที่การเติมหมู่เมทิลที่ไลซีน 9 เกี่ยวข้องกับการยับยั้งการถอดรหัสและเฮเทอโรโครมาติน ไลซีนเรซิดิวอื่นๆ บนฮิสโตน H3และฮิสโตน H4ก็เป็นตำแหน่งสำคัญของการเติมหมู่เมทิลโดยเอนไซม์ที่มีโดเมน SET จำเพาะเช่นกัน แม้ว่าฮิสโตนจะเป็นเป้าหมายหลักของไลซีนเมทิลทรานสเฟอเรส แต่โปรตีนในเซลล์อื่นๆ ก็มีสารตกค้าง N-เมทิลไลซีนอยู่ด้วย รวมถึงปัจจัยการยืดตัว 1A และโปรตีนตรวจจับแคลเซียมแคลโมดูลิน[ 9 ]
การเติมหมู่เมทิลที่ปลาย N
โปรตีนยูคาริโอตจำนวนมากได้รับการดัดแปลงหลังการแปลรหัสที่ปลาย N ของพวกมัน รูปแบบทั่วไปของการดัดแปลงปลาย N คือการเติมหมู่เมทิลที่ปลาย N (Nt-methylation) โดยเมทิลทรานสเฟอเรสที่ปลาย N (NTMTs) โปรตีนที่มีโมทีฟคอนเซนซัส H 2 N-X-Pro-Lys- (โดยที่ X สามารถเป็น Ala, Pro หรือ Ser) หลังจากการกำจัดเมไทโอนีนเริ่มต้น (iMet) สามารถได้รับการเติมหมู่เมทิลที่ปลาย N ด้วยหมู่ α-อะมิโน[ 10 ]การเติมหมู่เมทิลหนึ่งหมู่ อาจมีผลเล็กน้อยต่อความเป็นนิวคลีโอฟิลและความเป็นเบสของไนโตรเจน α-อะมิโน ในขณะที่การเติมหมู่เมทิลสามหมู่ (หรือการเติมหมู่เมทิลสองหมู่ในกรณีของโพรลีน) จะส่งผลให้ความเป็นนิวคลีโอฟิลหายไปและมีประจุบวกถาวรบนหมู่อะมิโนที่ปลาย N แม้ว่าจากมุมมองทางชีวเคมีการดีเมทิลเลชันของเอมีนจะเป็นไปได้ แต่การเมทิลเลชัน Nt ถือว่าไม่สามารถย้อนกลับได้ เนื่องจากยังไม่มีการอธิบายเอนไซม์ดีเมทิลเลสที่ปลาย N จนถึงปัจจุบัน[ 10 ]พบว่า ฮิสโตนชนิดCENP-AและCENP-B มีการเมทิลเลชัน Nt ในร่างกาย [ 10 ]
พรีนิลซิสเทอีน
โปรตีน ยูคาริโอตที่มีปลาย C ที่ลงท้ายด้วยโมทีฟ CAAX มักจะได้รับการดัดแปลงหลังการแปลรหัสหลายขั้นตอน การประมวลผลส่วนหาง CAAX เกิดขึ้นในสามขั้นตอน: ขั้นแรกตัวยึดไขมันพรีนิลจะถูกติดเข้ากับซิสเทอีนผ่าน พันธะ ไทโอเอสเทอร์ จากนั้นจะเกิดเอนโดโปรตีโอไลซิสเพื่อกำจัดกรดอะมิโนสามตัวสุดท้ายของโปรตีนเพื่อเปิดเผยหมู่ α-COOH ของพรีนิลซิสเทอีน สุดท้าย หมู่พรีนิลซิสเทอีนที่เปิดเผยจะถูกเมทิลเลชัน ความสำคัญของการดัดแปลงนี้สามารถเห็นได้จากการรบกวนเป้าหมายของเมทิลทรานสเฟอเรสสำหรับโปรตีน CAAX ของหนู ซึ่งการสูญเสียไอโซพรีนิลซิสเทอีนคาร์บอกซิลเมทิลทรานสเฟอเรสส่งผลให้เกิดการตายในช่วงกลางของการตั้งครรภ์[ 11 ]
หน้าที่ทางชีวภาพของการเติมหมู่เมทิลที่พรีนิลซิสเทอีนคือการอำนวยความสะดวกในการกำหนดเป้าหมายของโปรตีน CAAX ไปยังพื้นผิวเมมเบรนภายในเซลล์ พรีนิลซิสเทอีนสามารถถูกกำจัดหมู่เมทิลได้ และปฏิกิริยาย้อนกลับนี้ถูกเร่งปฏิกิริยาโดยไอโซพรีนิลซิสเทอีนคาร์บอกซิลเมทิลเอสเทอเรส โปรตีนที่มีกล่อง CAAX ที่ได้รับการเติมหมู่เมทิลที่พรีนิลซิสเทอีน ได้แก่Rasโปรตีนที่จับกับ GTP นิวเคลียร์ลามินและโปรตีนไคเนส บางชนิด โปรตีนเหล่านี้จำนวนมากมีส่วนร่วมในการส่งสัญญาณของเซลล์ และพวกมันใช้การเติมหมู่เมทิลที่พรีนิลซิสเทอีนเพื่อรวมพวกมันไว้บนพื้นผิวไซโตโซลของเยื่อหุ้มพลาสมาซึ่งเป็นที่ที่พวกมันทำงานได้[ 11 ]
การเติมหมู่เมทิลที่ปลาย C-terminus สามารถเพิ่มความหลากหลายทางเคมีของโปรตีนได้[ 12 ]และเป็นที่ทราบกันดีว่ามีผลอย่างมากต่อการทำงานของโปรตีน[ 1 ]
โปรตีนฟอสฟาเทส 2
ในเซลล์ยูคาริโอติกฟอสฟาเทสทำหน้าที่เร่งปฏิกิริยาการกำจัดหมู่ฟอสเฟตออกจากไทโรซีน เซรีน และทรีโอนีนฟอสโฟโปรตีน หน่วยย่อยเร่งปฏิกิริยาของเซรีน/ทรีโอนีนฟอสฟาเทสหลัก เช่นโปรตีนฟอสฟาเทส 2จะถูกดัดแปลงด้วยพันธะโควาเลนต์โดยการเติมหมู่เมทิลที่ปลาย C-terminus แบบย้อนกลับได้เพื่อสร้างลิวซีนคาร์บอกซีเมทิลเอสเทอร์ ซึ่งแตกต่างจากการเติมหมู่เมทิลที่โมทีฟ CAAX ตรงที่ไม่จำเป็นต้องมีการประมวลผลที่ปลาย C-terminus เพื่ออำนวยความสะดวกในการเติมหมู่เมทิล เหตุการณ์การเติมหมู่เมทิลที่ปลาย C-terminus นี้จะควบคุมการดึงดูดโปรตีนควบคุมเข้าสู่คอมเพล็กซ์ผ่านการกระตุ้นปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน ดังนั้นจึงควบคุมกิจกรรมของคอมเพล็กซ์เซรีน-ทรีโอนีนฟอสฟาเทสทางอ้อม[ 13 ]การเติมหมู่เมทิลจะถูกเร่งปฏิกิริยาโดยโปรตีนฟอสฟาเทสเมทิลทรานสเฟอเรสที่มีเอกลักษณ์เฉพาะ หมู่เมทิลจะถูกกำจัดออกโดยโปรตีนฟอสฟาเทสเมทิลเอสเทอเรสที่เฉพาะเจาะจง เอนไซม์สองชนิดที่ตรงข้ามกันนี้ทำให้การเติมหมู่เมทิลของเซริน-ทรีโอนีนฟอสฟาเทสเป็นกระบวนการแบบไดนามิกเพื่อตอบสนองต่อสิ่งเร้า[ 13 ]
แอล-ไอโซแอสปาร์ทิล
โปรตีนที่เสียหายจะสะสมไอโซแอสปาร์ทิลซึ่งทำให้โปรตีนไม่เสถียร สูญเสียการทำงานทางชีวภาพ และกระตุ้นการตอบสนองของระบบภูมิคุ้มกัน การสลายตัวของหมู่แอล-แอสปาร์ทิลที่เกิดขึ้นเองตามอายุ ส่งผลให้เกิดสารตัวกลางซัคซินิ มิดิล ซึ่งเป็นอนุมูล ซัคซินิมิด สารนี้จะถูกไฮโดรไลซ์เองโดยธรรมชาติกลับไปเป็นแอล-แอสปาร์ทิล หรือในปฏิกิริยาที่เกิดขึ้นได้ง่ายกว่า จะกลายเป็นแอล-ไอโซแอสปาร์ทิลที่ผิดปกติ มีวิถีทางที่ขึ้นอยู่กับเมทิลทรานสเฟอเรสสำหรับการเปลี่ยนแอล-ไอโซแอสปาร์ทิลกลับไปเป็นแอล-แอสปาร์ทิล เพื่อป้องกันการสะสมของแอล-ไอโซแอสปาร์ทิล หมู่ดังกล่าวจะถูกเมทิลเลชันโดยโปรตีนแอล-ไอโซแอสปาร์ทิลเมทิลทรานสเฟอเรสซึ่งเร่งปฏิกิริยาการสร้างเมทิลเอสเทอร์ ซึ่งจะถูกเปลี่ยนกลับไปเป็นสารตัวกลางซัคซินิมิดิลอีกครั้ง[ 14 ] การกลายพันธุ์ที่ทำให้เกิดการสูญเสียและการได้รับฟังก์ชันได้เผยให้เห็นถึงความสำคัญทางชีววิทยาของ L-isoaspartyl O-methyltransferase ในกระบวนการที่เกี่ยวข้องกับอายุ: หนูที่ขาดเอนไซม์จะตายตั้งแต่อายุยังน้อยด้วยโรคลมชักร้ายแรง ในขณะที่แมลงวันที่ได้รับการดัดแปลงพันธุกรรมให้มีการแสดงออกของเอนไซม์นี้มากเกินไปจะมีอายุขัยเพิ่มขึ้นมากกว่า 30% [ 14 ]
ผลกระทบทางกายภาพ
ธีมทั่วไปของโปรตีนที่มีการเติมหมู่เมทิล เช่นเดียวกับโปรตีนที่มีการเติมหมู่ฟอสเฟต คือบทบาทของการดัดแปลงนี้ในการควบคุมปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนการเติมหมู่เมทิลที่อาร์จินีนของโปรตีนสามารถยับยั้งหรือส่งเสริมปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนได้ ขึ้นอยู่กับชนิดของการเติมหมู่เมทิล การเติมหมู่เมทิลแบบไม่สมมาตรที่หมู่อาร์จินีนซึ่งอยู่ใกล้กับโมทีฟที่อุดมด้วยโพรลีนสามารถยับยั้งการจับกับโดเมน SH3ได้[ 15 ] ใน ทางตรงกันข้าม จะพบผลตรงกันข้ามกับปฏิสัมพันธ์ระหว่างโปรตีนการอยู่รอดของเซลล์ประสาทสั่งการและโปรตีน snRNP SmD1, SmD3 และ SmB/B' ซึ่งการจับกันจะถูกส่งเสริมโดยการเติมหมู่เมทิลแบบสมมาตรที่หมู่อาร์จินีนในโปรตีน snRNP [ 16 ]
ตัวอย่างที่มีลักษณะเฉพาะของการโต้ตอบระหว่างโปรตีนที่ขึ้นอยู่กับการเมทิลเลชันนั้นเกี่ยวข้องกับการเมทิลเลชันแบบเลือกของไลซีน 9 โดยSUV39H1 บนส่วนปลาย N ของฮิสโตน H3 [ 9 ]การไดเมทิลเลชันและไตรเมทิลเลชันของไลซีนตกค้างนี้ช่วยอำนวยความสะดวกในการจับของโปรตีนเฮเทอโรโครมาติน 1 (HP1) เนื่องจาก HP1 และ Suv39h1 โต้ตอบกัน จึงคิดว่าการจับของ HP1 กับฮิสโตน H3 ยังคงอยู่และอาจทำให้แพร่กระจายไปตามโครมาตินได้ โปรตีน HP1 มีโครโมโดเมนซึ่งรับผิดชอบต่อการโต้ตอบที่ขึ้นอยู่กับเมทิลระหว่างมันกับไลซีน 9 ของฮิสโตน H3 เป็นไปได้ว่าโปรตีนที่มีโครโมโดเมนเพิ่มเติมจะจับกับไซต์เดียวกันกับ HP1 และตำแหน่งไลซีนที่ถูกเมทิลเลชันอื่นๆ บนฮิสโตน H3 และฮิสโตน H4 [ 13 ]
การเมทิลเลชันของโปรตีนปลาย C ควบคุมการประกอบของโปรตีนฟอสฟาเทส การเมทิลเลชันของ ซับยูนิตเร่งปฏิกิริยาของ โปรตีนฟอสฟาเทส 2Aช่วยเพิ่มการจับของซับยูนิตควบคุม B และอำนวยความสะดวกในการประกอบโฮโลเอนไซม์[ 13 ]
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การเมทิลเลชันของโปรตีน
การเติมหมู่เมทิลให้กับโปรตีน เป็นรูปแบบหนึ่งของ การดัดแปลงหลังการแปลรหัส ซึ่งมีลักษณะเป็นการเพิ่ม หมู่เมทิล ให้กับ โปรตีน กระบวนการ นี้สามารถเกิดขึ้นได้ที่โซ่ข้างที่มีไนโตรเจนของ...
การเติมหมู่เมทิลโดยสารตั้งต้น
โปรตีนสามารถถูกเติมหมู่เมทิลได้หลายตำแหน่ง สำหรับการเติมหมู่เมทิลบางประเภท เช่น การเติมหมู่เมทิลที่ปลาย N-terminus และการเติมหมู่เมทิลที่พรีนิลซิสเทอีน จำเป็นต้องมีการประมวลผลเพิ่มเติม ในขณะที่การเติมหมู่เมทิลประเภทอื่น ๆ เช่น...
อาร์จินีน
อาร์จินีนสามารถถูกเมทิลเลชันได้หนึ่งครั้ง ( เมทิลอาร์จินีน ) หรือสองครั้ง (ไดเมทิลเลตอาร์จินีน) การเมทิลเลชันของหมู่อาร์จินีนถูกเร่งปฏิกิริยาโดยโปรตีนอาร์จินีนเมทิลทรานสเฟอเรส (PRMT) สามประเภทที่แตกต่างกัน: PRMT ประเภท I (PRMT1, PRMT2, PRMT3, PRMT4, PRMT6 และ...
ไลซีน
ไลซีนสามารถถูกเมทิลเลชันได้หนึ่งครั้ง สองครั้ง หรือสามครั้งโดยไลซีนเมทิลทรานสเฟอเรส (PKMTs) [ 8 ] ไลซีนเมทิลทรานสเฟอเรสส่วนใหญ่มี โดเมน SET ที่ได้รับการอนุรักษ์ทางวิวัฒนาการ ซึ่งมี กิจกรรมเมทิลทรานสเฟอเรสที่ขึ้นอยู่กับ S-adenosylmethionine...