อ่าน 9 นาที
สไปโซโซม
สไปลโซโซมเป็นคอมเพล็กซ์ไรโบนิวคลีโอโปรตีน (RNP) ขนาดใหญ่ที่พบได้เป็นหลักภายในนิวเคลียสของเซลล์ยูคาริโอติก สไปลโซโซมประกอบขึ้นจากอาร์เอ็นเอนิวเคลียร์ขนาดเล็ก ( snRNA )...
สไปโซโซม
สไปลโซโซมเป็นคอมเพล็กซ์ไรโบนิวคลีโอโปรตีน (RNP) ขนาดใหญ่ที่พบได้เป็นหลักภายในนิวเคลียสของเซลล์ยูคาริโอติก สไปลโซโซมประกอบขึ้นจากอาร์เอ็นเอนิวเคลียร์ขนาดเล็ก ( snRNA ) และโปรตีนจำนวนมาก โมเลกุลของอาร์เอ็นเอนิวเคลียร์ขนาดเล็ก (snRNA) จะจับกับโปรตีนเฉพาะเพื่อสร้างคอมเพล็กซ์ไรโบนิวคลีโอโปรตีนนิวเคลียร์ขนาดเล็ก (snRNP, ออกเสียงว่า "snurps") ซึ่งจะรวมกับ snRNP อื่นๆ เพื่อสร้างคอมเพล็กซ์ไรโบนิวคลีโอโปรตีนขนาดใหญ่ที่เรียกว่าสไปลโซโซม สไปลโซโซมจะกำจัดอินทรอนออกจาก พรี-mRNA ที่ถอดรหัส ซึ่งเป็นทราน สคริปต์หลักชนิดหนึ่งกระบวนการนี้โดยทั่วไปเรียกว่าการตัดต่อ [ 1 ] เปรียบเทียบ ได้กับบรรณาธิการภาพยนตร์ที่ตัดเนื้อหาที่ไม่เกี่ยวข้องหรือไม่ถูกต้อง (เทียบเท่ากับอินทรอน ) ออกจากภาพยนตร์ต้นฉบับอย่างเลือกสรร และส่งเวอร์ชันที่แก้ไขแล้วไปยังผู้กำกับเพื่อตัดต่อขั้นสุดท้าย
อย่างไรก็ตาม ในบางครั้ง RNA ภายในอินทรอนจะทำหน้าที่เป็นไรโบไซม์โดยทำการตัดต่อตัวเองโดยไม่ต้องใช้สไปโซโซมหรือเอนไซม์ใดๆ
ประวัติศาสตร์
ในปี พ.ศ. 2520 ผลงานของ ห้องปฏิบัติการ SharpและRobertsเปิดเผยว่ายีนของสิ่งมีชีวิตชั้นสูงนั้น "แยกออก" หรือมีอยู่ในหลายส่วนที่แตกต่างกันตามโมเลกุล DNA [ 2 ] [ 3 ] บริเวณที่เข้ารหัสของยีนถูกคั่นด้วยDNA ที่ไม่เข้ารหัสซึ่งไม่ได้เกี่ยวข้องกับการแสดงออกของโปรตีน โครงสร้างยีนที่แยกออกนี้พบได้เมื่อ mRNA ของอะดีโนไวรัสถูกไฮบริดกับชิ้นส่วนที่ถูกตัดด้วยเอนโดนิวคลีเอสของ DNA ไวรัสแบบสายเดี่ยว[ 2 ] พบว่า mRNA ของไฮบริด mRNA-DNA มี หาง 5'และ3'ของบริเวณที่ไม่มีพันธะไฮโดรเจน เมื่อใช้ชิ้นส่วน DNA ไวรัสขนาดใหญ่ขึ้น จะสังเกตเห็นโครงสร้างแบบแยกสาขาของ DNA ที่เป็นห่วงเมื่อไฮบริดกับ mRNA ของไวรัส พบว่าบริเวณที่เป็นห่วง ซึ่งก็คืออินทรอนถูกตัดออกจาก mRNA ต้นแบบในกระบวนการที่ Sharp ตั้งชื่อว่า "การต่อเชื่อม" (splicing) ต่อมาพบว่าโครงสร้างยีนแบบแยกส่วนนั้นพบได้ทั่วไปในยีน ของสิ่ง มีชีวิต ยูคาริโอตส่วนใหญ่ ฟิลิปป์ ชาร์ปและริชาร์ด เจ. โรเบิร์ตส์ได้รับรางวัลโนเบลสาขาการแพทย์ในปี 1998จากการค้นพบอินทรอนและกระบวนการสไปลซิง
องค์ประกอบ
สไปโซโซมแต่ละอันประกอบด้วยRNA นิวเคลียร์ขนาดเล็ก (snRNA) ห้า อันและปัจจัยโปรตีนที่เกี่ยวข้องหลายชนิด เมื่อ RNA ขนาดเล็กเหล่านี้รวมกับปัจจัยโปรตีน พวกมันจะสร้างสารประกอบ RNA-โปรตีนที่เรียกว่าsnRNP ( small nuclear ribonucleo proteinsออกเสียง ว่า " snurps ") snRNA ที่ประกอบเป็นสไปโซโซมหลักมีชื่อว่าU1 , U2 , U4 , U5และU6ซึ่งตั้งชื่อเช่นนี้เพราะอุดมไปด้วยยูริดีนและมีส่วนร่วมในปฏิกิริยา RNA-RNA และ RNA-โปรตีนหลายอย่าง[ 1 ]
การประกอบสไปโซโซมเกิดขึ้นที่พรีเอ็มอาร์เอ็นเอ แต่ละตัว (หรือที่เรียกว่าอาร์เอ็นเอนิวเคลียร์แบบไม่เป็นเนื้อเดียวกัน หรือ hn-RNA) ณ จุดเชื่อมต่อระหว่างเอ็กซอนและอินตรอนแต่ละจุด อินตรอนของพรีเอ็มอาร์เอ็นเอมีลำดับเบสจำเพาะที่ถูกจดจำและนำมาใช้ในระหว่างการประกอบสไปโซโซม ซึ่งได้แก่ ตำแหน่งเชื่อมต่อปลาย 5', ลำดับเบสจุดแยกแขนง, ส่วนโพลีไพริมิดีน และตำแหน่งเชื่อมต่อปลาย 3' สไปโซโซมทำหน้าที่เร่งปฏิกิริยาการกำจัดอินตรอนและการเชื่อมต่อเอ็กซอนที่อยู่ข้างเคียง
โดยทั่วไป อินทรอนจะมีลำดับนิวคลีโอไทด์ GU ที่ตำแหน่งปลาย 5' และ AG ที่ตำแหน่งปลาย 3' ตำแหน่งปลาย 3' สามารถกำหนดได้ละเอียดขึ้นด้วยโพลีไพริมิดีนที่มีความยาวแปรผันได้ เรียกว่า โพลีไพริมิดีนแทร็ก (PPT) ซึ่งทำหน้าที่สองอย่างคือ ดึงดูดปัจจัยต่างๆ มายังตำแหน่งปลาย 3' และอาจดึงดูดปัจจัยต่างๆ มายังลำดับจุดแยกสาขา (BPS) ด้วย BPS ประกอบด้วยอะดีโนซีน ที่คงสภาพ ซึ่งจำเป็นสำหรับขั้นตอนแรกของการตัดต่อ
โปรตีนหลายชนิดแสดงโมทีฟการจับสังกะสี ซึ่งเน้นย้ำถึงความสำคัญของสังกะสีในกลไกการตัดต่อ[ 4 ] [ 5 ] [ 6 ]การสร้างโครงสร้างระดับโมเลกุลครั้งแรกของคอมเพล็กซ์ไตรนิวคลีโอโปรตีนนิวเคลียร์ขนาดเล็กสามตัว U4/U6.U5 (tri-snRNP) ได้รับการรายงานในปี 2016 [ 7 ]

Cryo-EM ถูกนำมาใช้อย่างกว้างขวางโดย Shi et al. เพื่ออธิบายโครงสร้างระดับใกล้อะตอม/อะตอมของสไปโซโซมทั้งในยีสต์[ 9 ]และมนุษย์[ 10 ]โครงสร้างโมเลกุลของสไปโซโซมที่ความละเอียดระดับใกล้อะตอมแสดงให้เห็นว่าส่วนประกอบ Spp42 ของ U5 snRNP สร้างโครงสร้างหลักและยึดศูนย์กลางเร่งปฏิกิริยาในยีสต์ โครงสร้างอะตอมของสไปโซโซมของมนุษย์แสดงให้เห็นว่าส่วนประกอบขั้นตอนที่ II Slu7 มีโครงสร้างที่ยืดออก พร้อมที่จะเลือกไซต์การตัดต่อ 3' โลหะทั้งห้าชนิด (กำหนดให้เป็น Mg2+) ในคอมเพล็กซ์ของยีสต์ยังคงอยู่ในคอมเพล็กซ์ของมนุษย์
การตัดต่อทางเลือก
การต่อเชื่อมแบบทางเลือก (การรวมตัวกันใหม่ของเอ็กซอน ที่แตกต่างกัน ) เป็นแหล่งสำคัญของความหลากหลายทางพันธุกรรมในยูคาริโอต ตัวแปรการต่อเชื่อมถูกนำมาใช้เพื่ออธิบายจำนวนยีน ที่เข้ารหัสโปรตีนที่ค่อนข้างน้อย ในจีโนมมนุษย์ ซึ่งปัจจุบันคาดว่ามีประมาณ 20,000 ยีน ยีนDscam ของแมลงหวี่ตัวหนึ่งถูกคาดการณ์ว่ามีการต่อเชื่อมแบบทางเลือกเป็นmRNA ที่แตกต่างกันถึง 38,000 ชนิด โดยสมมติว่าเอ็กซอนทั้งหมดสามารถต่อเชื่อมได้อย่างอิสระจากกันและกัน[ 11 ]
ตำแหน่งของการเชื่อมต่อ
ปัจจัยการตัดต่อ pre-mRNA เดิมทีพบว่ามีความเข้มข้นอยู่ในนิวเคลียร์บอดี้ที่เรียกว่านิวเคลียร์สเปคเคิล[ 12 ]เดิมทีมีการตั้งสมมติฐานว่านิวเคลียร์สเปคเคิลเป็นได้ทั้งไซต์ของการตัดต่อ mRNA หรือไซต์เก็บสะสมปัจจัยการตัดต่อ mRNA ปัจจุบันเป็นที่เข้าใจกันว่านิวเคลียร์สเปคเคิลช่วยเพิ่มความเข้มข้นของปัจจัยการตัดต่อให้อยู่ใกล้กับยีนที่อยู่ใกล้เคียง ยีนที่อยู่ห่างจากสเปคเคิลยังคงสามารถถอดรหัสและตัดต่อได้ แต่การตัดต่อจะมีประสิทธิภาพน้อยกว่าเมื่อเทียบกับยีนที่อยู่ใกล้กับสเปคเคิล[ 13 ]การตัดต่อ RNAเป็นปฏิกิริยาทางชีวเคมี และเช่นเดียวกับปฏิกิริยาทางชีวเคมีทั้งหมด อัตราการเกิดปฏิกิริยาขึ้นอยู่กับความเข้มข้นของเอนไซม์และสารตั้งต้น ในกรณีนี้ เอนไซม์คือสไปโซโซม และสารตั้งต้นคือ pre-mRNA การเปลี่ยนแปลงความเข้มข้นของสไปโซโซมและ pre-mRNA ตามความใกล้เคียงกับนิวเคลียร์สเปคเคิล เซลล์อาจสามารถควบคุมประสิทธิภาพของการตัดต่อได้[ 13 ]
การประกอบ
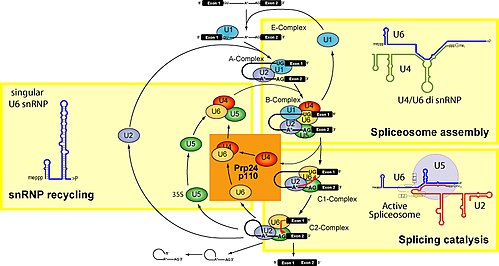
แบบจำลองสำหรับการก่อตัวของตำแหน่งออกฤทธิ์ของสไปโซโซมเกี่ยวข้องกับการประกอบอนุภาค snRNP ที่แยกจากกันอย่างเป็นระเบียบและเป็นขั้นตอนบนพื้นผิวพรี-mRNA การจดจำพรี-mRNA ครั้งแรกเกี่ยวข้องกับการจับกันของ U1 snRNP กับตำแหน่งสไปซ์ปลาย 5' ของพรี-mRNA และปัจจัยอื่นๆ ที่ไม่เกี่ยวข้องกับ snRNP เพื่อสร้างคอมเพล็กซ์ที่กำหนด หรือคอมเพล็กซ์ต้น (E) ในสัตว์เลี้ยงลูกด้วยนม[ 14 ] [ 15 ] คอมเพล็กซ์ที่กำหนดเป็นคอมเพล็กซ์ที่ไม่ขึ้นกับ ATP ซึ่งกำหนดพรี-mRNA ให้กับเส้นทางการสไปซ์[ 16 ] U2 snRNP จะถูกดึงดูดไปยังบริเวณกิ่งก้านผ่านปฏิสัมพันธ์กับส่วนประกอบของคอมเพล็กซ์ E คือU2AF (ปัจจัยเสริมของ U2 snRNP) และอาจรวมถึง U1 snRNP ด้วย ในปฏิกิริยาที่ขึ้นอยู่กับ ATP นั้น U2 snRNP จะเชื่อมโยงอย่างแน่นหนากับลำดับจุดแตกแขนง (BPS) เพื่อสร้างคอมเพล็กซ์ A คู่เกลียวที่เกิดขึ้นระหว่าง U2 snRNP และบริเวณแตกแขนงของ pre-mRNA จะยื่นอะดีโนซีนของแขนงออกมา ทำให้ระบุว่าเป็นนิวคลีโอไฟล์สำหรับการถ่ายโอนเอสเทอร์ครั้งแรก[ 17 ]
การมีอยู่ของ สารตกค้าง pseudouridineใน U2 snRNA ซึ่งอยู่เกือบตรงข้ามกับตำแหน่งกิ่ง ส่งผลให้โครงสร้างของ RNA-RNA duplex เปลี่ยนแปลงไปเมื่อ U2 snRNP จับกับ RNA-RNA โดยเฉพาะอย่างยิ่ง โครงสร้างของ duplex ที่เปลี่ยนแปลงไปซึ่งเกิดจาก pseudouridine ทำให้ 2' OH ของ adenosine ที่โป่งออกมาอยู่ในตำแหน่งที่เหมาะสมสำหรับขั้นตอนแรกของการตัดต่อ[ 18 ] U4/U5/U6 tri-snRNP (ดูรูปที่ 1) จะถูกดึงดูดไปยัง spliceosome ที่กำลังประกอบเพื่อสร้าง complex B และหลังจากมีการจัดเรียงใหม่หลายครั้ง complex C จะถูกกระตุ้นเพื่อเร่งปฏิกิริยา[ 19 ] [ 20 ]ยังไม่ชัดเจนว่า tri-snRNP ถูกดึงดูดไปยัง complex A ได้อย่างไร แต่กระบวนการนี้อาจเกิดขึ้นผ่านปฏิกิริยาระหว่างโปรตีนกับโปรตีนและ/หรือปฏิกิริยาการจับคู่เบสระหว่าง U2 snRNA และ U6 snRNA
U5 snRNP โต้ตอบกับลำดับที่ไซต์สไปลซ์ 5' และ 3' ผ่านลูปคงที่ของ U5 snRNA [ 21 ]และส่วนประกอบโปรตีน U5 โต้ตอบกับบริเวณไซต์สไปลซ์ 3' [ 22 ]
เมื่อมีการคัดเลือก tri-snRNP การจัดเรียง RNA-RNA ใหม่หลายครั้งจะเกิดขึ้นก่อนขั้นตอนการเร่งปฏิกิริยาครั้งแรก และการจัดเรียงใหม่เพิ่มเติมจะเกิดขึ้นในสไปโซโซมที่ทำงานเร่งปฏิกิริยา ปฏิสัมพันธ์ RNA-RNA หลายอย่างเป็นแบบแยกจากกันโดยสิ้นเชิง อย่างไรก็ตาม ยังไม่ทราบว่าอะไรเป็นตัวกระตุ้นปฏิสัมพันธ์เหล่านี้ หรือลำดับของการจัดเรียงใหม่เหล่านี้ การจัดเรียงใหม่ครั้งแรกน่าจะเป็นการแทนที่U1 snRNPจากไซต์สไปซ์ 5' และการก่อตัวของปฏิสัมพันธ์ U6 snRNA เป็นที่ทราบกันว่า U1 snRNP เชื่อมโยงกับสไปโซโซมที่ก่อตัวอย่างสมบูรณ์เพียงเล็กน้อย[ 23 ]และ U1 snRNP ยับยั้งการก่อตัวของปฏิสัมพันธ์ U6-ไซต์สไปซ์ 5' บนแบบจำลองของสารตั้งต้นโอลิโกนิวคลีโอไทด์ที่มีเอ็กซอน 5' สั้นและไซต์สไปซ์ 5' [ 24 ] การจับของ U2 snRNP กับลำดับจุดแตกแขนง (BPS) เป็นตัวอย่างหนึ่งของปฏิสัมพันธ์ RNA-RNA ที่แทนที่ปฏิสัมพันธ์โปรตีน-RNA เมื่อมีการดึง U2 snRNP เข้ามา โปรตีน SF1 ที่ทำหน้าที่จับกับกิ่งในคอมเพล็กซ์ที่กำหนดทิศทางจะถูกแทนที่ เนื่องจากตำแหน่งการจับของ U2 snRNA และ SF1 เป็นเหตุการณ์ที่ไม่สามารถเกิดขึ้นพร้อมกันได้
ภายใน U2 snRNA มีการจัดเรียงใหม่แบบแยกกันเองอื่นๆ ที่เกิดขึ้นระหว่างคอนฟอร์เมชันที่แข่งขันกัน ตัวอย่างเช่น ในรูปแบบที่ใช้งานอยู่ ก้านห่วง IIa เป็นที่นิยม ในรูปแบบที่ไม่ทำงาน ปฏิสัมพันธ์แบบแยกกันเองระหว่างห่วงและลำดับปลายน้ำจะเด่นกว่า[ 20 ]ยังไม่ชัดเจนว่า U4 ถูกแทนที่ออกจาก U6 snRNA ได้อย่างไร แม้ว่า RNA จะมีส่วนเกี่ยวข้องกับการประกอบสไปโซโซม และอาจทำหน้าที่คลายเกลียว U4/U6 และส่งเสริมการก่อตัวของปฏิสัมพันธ์ U2/U6 snRNA ปฏิสัมพันธ์ของก้านห่วง I และ II ของ U4/U6 จะแยกออกจากกัน และบริเวณก้านห่วง II ที่เป็นอิสระของ U6 จะพับตัวเองเพื่อสร้างก้านห่วงภายในโมเลกุล และ U4 ไม่จำเป็นอีกต่อไปในการประกอบสไปโซโซมต่อไป บริเวณก้านห่วงอิสระ I ของ U6 จับคู่กับ snRNA ของ U2 ก่อให้เกิดโครงสร้างเกลียว I ของ U2/U6 อย่างไรก็ตาม โครงสร้างเกลียว I นั้นไม่สามารถจับคู่กับครึ่ง 3' ของบริเวณก้านห่วง 5' ภายในของ snRNA ของ U2 ได้
สไปโซโซมขนาดเล็ก
ยูคาริโอตบางชนิดมีสไปโซโซมตัวที่สอง ซึ่งเรียกว่าสไปโซโซมรอง[ 25 ] กลุ่ม snRNA ที่มีปริมาณน้อยกว่า ได้แก่U11 , U12 , U4atacและU6atacร่วมกับ U5 เป็นหน่วยย่อยของสไปโซโซมรองที่ตัดต่ออินทรอน pre-mRNA ชนิดหายาก ซึ่งเรียกว่าชนิด U12 สไปโซโซมรองตั้งอยู่ในนิวเคลียสเช่นเดียวกับสไปโซโซมหลัก[ 26 ]แม้ว่าจะมีข้อยกเว้นในเซลล์เฉพาะบางชนิด เช่น เกล็ดเลือดที่ไม่มีนิวเคลียส[ 27 ]และเดนโดรพลาสม์ ( ไซโตพลาสม์ของ เดนไดรต์ ) ของเซลล์ประสาท[ 28 ]
อ่านเพิ่มเติม
- Butcher SE (2011). "บทที่ 8. สไปโซโซมและไอออนโลหะ". ใน Sigel A, Sigel H, Sigel RK (บรรณาธิการ). บทบาทเชิงโครงสร้างและเร่งปฏิกิริยาของไอออนโลหะใน RNA . ไอออนโลหะในวิทยาศาสตร์ชีวภาพ. เล่มที่ 9. RSC Publishing. หน้า 235–251 . doi : 10.1039/9781849732512-00235 . ISBN 978-1-84973-094-5PMID 22010274
- Nilsen TW (ธันวาคม 2003). "สไปโซโซม: เครื่องจักรโมเลกุลขนาดใหญ่ที่ซับซ้อนที่สุดในเซลล์หรือไม่?" BioEssays . 25 (12): 1147– 9. doi : 10.1002/bies.10394 . PMID 14635248 .
ลิงก์ภายนอก
- โครงสร้างระดับมหภาคสามมิติของสไปโซโซมจากฐานข้อมูล EM (EMDB)
- Spliceosomes ใน หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ สไปโซโซม
สไปลโซโซมเป็นคอมเพล็กซ์ไรโบนิวคลีโอโปรตีน (RNP) ขนาดใหญ่ที่พบได้เป็นหลักภายในนิวเคลียสของเซลล์ยูคาริโอติก สไปลโซโซมประกอบขึ้นจากอาร์เอ็นเอนิวเคลียร์ขนาดเล็ก ( snRNA )...
ประวัติศาสตร์
ในปี พ.ศ. 2520 ผลงานของ ห้องปฏิบัติการ Sharp และ Roberts เปิดเผยว่ายีนของสิ่งมีชีวิตชั้นสูงนั้น "แยกออก" หรือมีอยู่ในหลายส่วนที่แตกต่างกันตามโมเลกุล DNA [ 2 ] [ 3 ] บริเวณที่เข้ารหัสของยีนถูกคั่นด้วย DNA ที่ไม่เข้ารหัส...
องค์ประกอบ
สไปโซโซมแต่ละอันประกอบด้วย RNA นิวเคลียร์ขนาดเล็ก (snRNA) ห้า อัน และปัจจัยโปรตีนที่เกี่ยวข้องหลายชนิด เมื่อ RNA ขนาดเล็กเหล่านี้รวมกับปัจจัยโปรตีน พวกมันจะสร้างสารประกอบ RNA-โปรตีนที่เรียกว่า snRNP ( small nuclear ribonucleo proteins ออกเสียง ว่า " snurps ")...
การตัดต่อทางเลือก
การต่อเชื่อมแบบทางเลือก (การรวมตัวกันใหม่ของ เอ็กซอน ที่แตกต่างกัน ) เป็นแหล่งสำคัญของ ความหลากหลายทางพันธุกรรม ในยูคาริ โอต ตัวแปรการต่อเชื่อม ถูกนำมาใช้เพื่ออธิบายจำนวน ยีน ที่เข้ารหัสโปรตีนที่ค่อนข้างน้อย ใน จีโนมมนุษย์ ซึ่งปัจจุบันคาดว่ามีประมาณ 20,000...