อ่าน 11 นาที
เทโนโมดูลิน
เทโนโมดู ลิ นหรือเรียกอีกอย่างว่าเทนดินไมโอดูลินTnmdหรือTeM เป็นโปรตีนที่เข้ารหัสโดยยีน TNMD (Tnmd) และถูกค้นพบโดยอิสระโดย Brandau และ Shukunami ในปี 2001...
เทโนโมดูลิน
| ทีเอ็นเอ็มดี | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | TNMD , BRICD4, CHM1L, TEM, ทีโนโมดูลิน | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 300459 ; เอ็มจีไอ : 1929885 ; โฮโมโลยีน : 11152 ; GeneCards : TNMD ; OMA : TNMD - ออร์โธล็อก | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




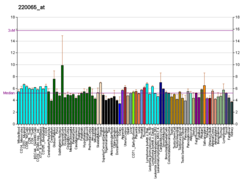
เทโนโมดู ลิ นหรือเรียกอีกอย่างว่าเทนดินไมโอดูลินTnmdหรือTeM [ 5 ]เป็นโปรตีนที่เข้ารหัสโดยยีน TNMD (Tnmd) และถูกค้นพบโดยอิสระโดย Brandau และ Shukunami ในปี 2001 ในฐานะยีนที่มีความคล้ายคลึงสูงกับคอนโดรโมดูลิน-1 (Chm1) ที่รู้จักกันอยู่แล้ว [ 6 ] [ 7 ]เป็นยีนบ่งชี้เฉพาะของเอ็นที่ทราบกันว่ามีความสำคัญต่อการเจริญเติบโตของเอ็น โดยมีนัยสำคัญต่อเซลล์ต้นกำเนิด/เซลล์บรรพบุรุษของเอ็น (TSPCs) ตลอดจนการควบคุมการเคลื่อนย้ายของเซลล์ บุผนังหลอดเลือด ในเอ็นยึดหัวใจและในแบบจำลองเนื้องอกทดลอง มีการแสดงออกสูงในเอ็นซึ่งอธิบายถึงเหตุผลเบื้องหลังชื่อและการจัดตั้งเป็นยีนบ่งชี้สำหรับสายพันธุ์เอ็นและเอ็นยึด[ 8 ]
โครงสร้างของยีนและโปรตีน
TNMD จัดอยู่ในกลุ่ม โปรตีนไกลโคโปรตีนทรานส์เมมเบรนชนิด II กลุ่มใหม่ยีนนี้ตั้งอยู่บนโครโมโซม Xและมีทรานสคริปต์ประมาณ 1.4 กิโลเบส และโปรตีนที่คาดการณ์ไว้ประกอบด้วยกรดอะมิโน 317 ตัว[ 6 ] [ 7 ]ยีนนี้ประกอบด้วยเอ็กซอน 7 ส่วน เอ็กซอนที่สองเข้ารหัสโดเมนทรานส์เมมเบรน (ตำแหน่งกรดอะมิโน 31-49) และไม่มีเปปไทด์สัญญาณ TNMD มีลำดับการจดจำโปรตีเอสที่คาดการณ์ไว้ (Arg-Xxx-Xxx-Arg) ที่ระบุตำแหน่ง 233-236 [ 9 ] [ 10 ] [ 11 ]แตกต่างจากคอนโดรโมดูลิน-1 TNMD ไม่มีสัญญาณการประมวลผลสำหรับ โปรตีเอส ฟูรินส่วนนอกเซลล์ ก่อนถึงตำแหน่งการตัดที่คาดการณ์ไว้ มีโดเมนนอกเซลล์ BRICHOS ซึ่งพบในโปรตีนอื่นๆ ที่ไม่เกี่ยวข้องอีกหลายชนิด โดเมนนี้ประกอบด้วยลำดับโฮโมล็อกของกรดอะมิโนประมาณ 100 ตัว ซึ่งมีกรดอะมิโนซิสเทอีนที่อนุรักษ์ไว้คู่หนึ่ง มีการเสนอแนะว่า BRICHOS มีส่วนร่วมในกระบวนการหลังการแปลโปรตีน อย่างไรก็ตาม หน้าที่ที่แท้จริงยังคงไม่ชัดเจน[ 12 ] TNMD มี ไซต์ N-ไกลโคซิเลชัน สอง ไซต์ที่ตำแหน่ง 94 และ 180 [ 7 ]การวิเคราะห์โปรตีนในตาและเอ็นยึดปริทันต์เผยให้เห็นโปรตีน TNMD ที่มีความยาวเต็มเป็นแถบคู่ขนาด 40 และ 45 kDa [ 9 ] [ 13 ]ได้รับการพิสูจน์ทางทดลองแล้วว่าแถบขนาด 45 kDa สอดคล้องกับ TNMD ที่มีไกลโคซิเลชัน ในขณะที่แถบขนาด 40 kDa คือ TNMD ที่ไม่มีไกลโคซิเลชัน[ 13 ]เอ็กซอนสุดท้ายของยีน TNMD เข้ารหัสโดเมนที่อุดมด้วยซิสเทอีนที่ปลาย C ซึ่งได้รับการอนุรักษ์ไว้ ซึ่งเป็นส่วนประกอบของโปรตีนที่มีความคล้ายคลึงกับคอนโดรโมดูลิน-I มากที่สุด (ความคล้ายคลึง 77% / ความเหมือน 66%) [ 7 ]โดเมนนี้ประกอบด้วยหางไฮโดรโฟบิกที่ปลาย C ซึ่งมีสารตกค้างซิสเทอีน 8 ตัวที่สร้างสะพานไดซัลไฟด์ 4 อัน ซึ่งได้รับการอนุรักษ์ไว้อย่างดีในสัตว์มีกระดูกสันหลังหลายชนิด[ 9 ] [ 14 ]โครงสร้างวงจรขนาดเล็กที่เกิดจากสะพานไดซัลไฟด์ Cys280-Cys292 เพียงอันเดียวใน TNMD ได้รับการแสดงให้เห็นว่ามีฟังก์ชันต้านการสร้างหลอดเลือดใหม่[ 15 ]ในขณะที่สะพานไดซัลไฟด์อีก 3 อันนั้นคาดว่าจะยึดโครงสร้างวงจรนี้และหางไฮโดรโฟบิกที่ปลาย C ให้แยกออกจากกันเพื่อหลีกเลี่ยงการก่อตัวของสารรวมตัวภายในโมเลกุล[ 15 ]ในเนื้อเยื่อเอ็นบางชนิด เช่นเอ็นร้อยหวายและ chordae tendineae cordis ส่วนปลาย C ที่ถูกตัดออกขนาด 16 kDa ของ TNMD ถูกตรวจพบในเมทริกซ์นอก เซลล์ที่เป็นคอลลา เจน[ 16 ] [ 17 ]
รูปแบบการแสดงออก
TNMD มีการแสดงออกในระดับสูงทั้งในระดับสารส่งสัญญาณและโปรตีนในเอ็นและเส้นเอ็น แต่ก็พบได้ในเนื้อเยื่ออื่นๆ ด้วยเช่นกัน
- ในการพัฒนาเอ็น สามารถตรวจพบสัญญาณแรกได้เร็วที่สุดที่ E9.5 [ 7 ]แต่มีการเพิ่มขึ้นตั้งแต่ E14.5 เป็นต้นไป ซึ่งบ่งบอกถึงระยะที่แตกต่างกันของเซลล์ต้นกำเนิดเอ็น[ 18 ]
- เอ็นยึดปริทันต์ของหนูแสดงให้เห็นการแสดงออกของโปรตีนเทโนโมดูลินในช่วง 3 และ 4 สัปดาห์หลังคลอด ซึ่งเป็นช่วงเวลาที่สอดคล้องกับระยะการงอกและระยะหลังการงอกของฟันกรามเมื่อฟันเริ่มใช้งานได้[ 13 ]
- เนื้อเยื่อเอ็นอื่นๆ ที่ทราบกันว่ามีการแสดงออกของ Tnmd ได้แก่ ไดอะแฟรม[ 7 ]และเอ็นยึดหัวใจ[ 17 ]
- กล้ามเนื้อมาสเซเตอร์ถูกแบ่งเป็นส่วนๆ ด้วยโครงสร้างแบบแผ่น ซึ่งแสดงให้เห็นว่ามีการเพิ่มระดับ mRNA ของ Tnmd ในตัวอ่อนของหนูระหว่าง E12.5 ถึง E17.5 ซึ่งลดลงอีกหลังคลอด[ 19 ]เอพิไมเซียมของกล้ามเนื้อโครงร่างก็มี TNMD เป็นบวกเช่นกัน[ 6 ] [ 7 ]
- ตรวจพบ mRNA ของ Tnmd ในดวงตา โดยเฉพาะใน sclerocornea, เอ็นของกล้ามเนื้อนอกลูกตา และ ชั้น เซลล์แกงลีออนของจอประสาทตา , เซลล์เส้นใยเลนส์, เซลล์ชั้นนิวเคลียสชั้นใน และเยื่อบุผิวเม็ดสี[ 20 ]
- ตรวจพบ mRNA ของ Tnmd ในผิวหนังของหนูที่ E15.5 และในเนื้อเยื่อไขมันใต้ผิวหนังและเซลล์ไขมันของมนุษย์[ 21 ]
- การตรวจด้วยวิธีไฮบริดไดเซชันในแหล่งกำเนิดเผยให้เห็นการแสดงออกของ Tnmd ในส่วนต่างๆ ของสมองหนู โตเต็มวัย เช่นเดนเตตไจรัสบริเวณ CA ของฮิปโปแคมปัสเซลล์ประสาทในนิวเคลียสของสมองใหญ่สมองน้อยเซลล์พูร์คินเจและเซลล์ประสาทในนิวเคลียสของสมองน้อย[ 7 ]
- กระดูกอ่อนข้อต่อขากรรไกรของหนูมี mRNA ของ Tnmd เป็นบวกในสัปดาห์ที่ 1 และลดลงหลังจาก 5 สัปดาห์[ 22 ]
เส้นทางการส่งสัญญาณที่คาดการณ์ไว้
เส้นทางการส่งสัญญาณที่คาดการณ์ไว้ของ TNMD ส่วนใหญ่ยังไม่เป็นที่รู้จักเนื่องจากพันธมิตรการจับโดยตรงที่ยังไม่ได้ รับการระบุ แบบจำลอง หนูน็อคเอาท์ หลายแบบ ที่มีฟีโนไทป์ของเอ็นช่วยให้เข้าใจว่าปัจจัยต้นน้ำหรือเส้นทางใดส่งผลต่อการแสดงออกของ Tnmd ในทำนองเดียวกัน การสร้างแบบจำลองหนูน็อคเอาท์ Tnmd ช่วยให้สามารถเสนอแนะตัวกระตุ้นปลายน้ำที่เป็นไปได้ การศึกษาด้านล่างส่วนใหญ่แสดงให้เห็นความสัมพันธ์ระหว่างการแสดงออกหรือการทำงานของ Tnmd กับยีนอื่น ๆ และไม่ใช่การเชื่อมโยงโดยตรงในลำดับการส่งสัญญาณทั่วไป เกี่ยวกับตัวควบคุมต้นน้ำของการแสดงออกของ Tnmd คำอธิบายของ สายพันธุ์หนูน็อคเอาท์ scleraxis (Scx) ชี้ให้เห็นว่า Scx สามารถขับเคลื่อนการถอดรหัส Tnmd ได้โดยตรง เนื่องจากการลบ Scx นำไปสู่การกำจัดการแสดงออกของ Tnmd อย่างสมบูรณ์[ 23 ]การแสดงออกเกินของ scleraxis ในเทโนไซต์ที่เพาะเลี้ยง[ 8 ]หรือในเซลล์ต้นกำเนิดมีเซนไคม์ทำให้การแสดงออกของ Tnmd เพิ่มขึ้นอย่างมีนัยสำคัญ[ 24 ]การลบไมโอสแตตินในหนูส่งผลให้ระดับ mRNA ของ Scx และ Tnmd ลดลงพร้อมกัน[ 25 ]ในขณะที่การกระตุ้นไฟโบรบลาสต์ด้วยไมโอสแตตินทำให้มีการเพิ่มระดับขึ้น ซึ่งแสดงให้เห็นว่าไมโอสแตตินเป็นปัจจัยต้นน้ำในเส้นทาง Tnmd ปัจจัยการถอดรหัส Egr1/2 สามารถกระตุ้นการแสดงออกของยีน Scx และคอลลาเจน I ได้[ 26 ]ดังนั้นจึงน่าสนใจที่จะตรวจสอบว่า Egr1 หรือ 2 สามารถส่งผลต่อการแสดงออกของ Tnmd ได้หรือไม่ การไม่มีอยู่ของยีน Mohawk (Mkx) ส่งผลให้การแสดงออกของ Tnmd ลดลงอย่างมีนัยสำคัญ เช่นเดียวกับคอลลาเจน I และไฟโบรโมดูลิน[ 27 ]การสูญเสีย Tnmd อย่างมีนัยสำคัญนั้นสังเกตได้ในหนูที่ขาด Mkx ที่ E16.5 ในขณะที่การแสดงออกของ Scx ไม่เปลี่ยนแปลง [22] ซึ่งแสดงให้เห็นว่า Mkx สามารถส่งผลต่อการแสดงออกของ Tnmd ได้โดยตรงเช่นกัน การกระตุ้นวิถีการส่งสัญญาณ Wnt/β-catenin ในเซลล์ต้นกำเนิดที่ได้จากไขกระดูกส่งผลให้มีการเพิ่มการแสดงออกของ Tnmd การแสดงออกของ Scx และ Mkx ไม่ได้รับผลกระทบ ซึ่งบ่งชี้ว่าการส่งสัญญาณ Wnt/β-catenin ทำงานโดยอิสระจากปัจจัยการถอดรหัสเหล่านี้[ 28 ] ในส่วนของปัจจัยปลายน้ำ แบบจำลองหนูที่ขาด Tnmd ชี้ให้เห็นถึงความสัมพันธ์กับคอลลาเจน Iโดยอิงจากการสร้างเส้นใยคอลลาเจนที่ผิดปกติซึ่งส่งผลให้เส้นใยหนาขึ้นอย่างผิดปกติ[ 16 ]ความหนาแน่นของเซลล์และการแพร่กระจายที่ลดลงในเอ็นที่กลายพันธุ์[ 16 ]รวมถึงการต่ออายุตัวเองที่ลดลงและการแก่ก่อนวัยของเซลล์ต้นกำเนิด/เซลล์บรรพบุรุษของเอ็นที่ขาด Tnmd นั้นเชื่อมโยงกับการลดลงของการแสดงออกของเครื่องหมายการแพร่กระจาย Cyclin D1 และการเพิ่มขึ้นของการแสดงออกของเครื่องหมายการแก่ชรา p53 [ 29 ]การศึกษาวิเคราะห์การฉีกขาดของเอ็นยึดหัวใจของมนุษย์เผยให้เห็นการสูญเสียการแสดงออกของ Tnmd ในบริเวณที่ได้รับผลกระทบควบคู่กับการเพิ่มขึ้นของการแสดงออกของ VEGF-A และ MMP1, 2 และ 13 [ 17 ]
หน้าที่และความสัมพันธ์กับโรค
ในช่วงทศวรรษที่ผ่านมา มีความก้าวหน้าครั้งสำคัญในการทำความเข้าใจบทบาทของ TNMD ในเส้นเอ็นและเนื้อเยื่อและเซลล์อื่นๆ อย่างไรก็ตาม หน้าที่ที่แท้จริงของ TNMD นั้นแตกต่างกันไปตามชนิดของเซลล์และเนื้อเยื่อ และส่วนใหญ่ยังคงไม่ได้รับการไขปริศนาอย่างสมบูรณ์ นอกจากนี้ ยังไม่เป็นที่แน่ชัดว่า TNMD มีส่วนเกี่ยวข้องกับพยาธิสภาพของโรคที่เกี่ยวข้องบางชนิดอย่างไร
- ในเอ็นนั้นพิสูจน์แล้วว่ามีหน้าที่ที่เป็นประโยชน์ต่อการบำรุงรักษาเนื้อเยื่อ เนื่องจากการสูญเสียส่งผลให้เอ็นเสื่อมสภาพก่อนวัยอันควร ซึ่งมีลักษณะเฉพาะคือการสร้างเส้นใยคอลลาเจนที่ผิดปกติ และความหนาแน่นและการแพร่กระจายของเซลล์ลดลง[ 16 ] Tnmd มีผลดีต่อเซลล์ต้นกำเนิด/เซลล์บรรพบุรุษที่ได้จากเอ็น โดยสนับสนุนการสร้างใหม่ด้วยตนเองและป้องกันการเสื่อมสภาพ ซึ่งการกระทำดังกล่าวมีเพียงโดเมนที่อุดมด้วยซิสเทอีนที่ปลาย C เท่านั้นก็เพียงพอแล้ว[ 29 ]การศึกษาครั้งแรกเกี่ยวกับการแสดงออกของ Tnmd ในระหว่างการรักษาเอ็นชี้ให้เห็นถึงบทบาทที่ขึ้นอยู่กับเวลา ซึ่งจำเป็นต้องมีการอธิบายเพิ่มเติม[ 30 ] [ 31 ]
- ในเอ็นยึดปริทันต์ที่ทำหน้าที่เชื่อมต่อฟันกับกระดูกขากรรไกร Tnmd มีส่วนช่วยในการยึดเกาะของไฟโบรบลาสต์ อย่างเหมาะสม [ 13 ]
- ในโครงสร้างเอ็น chordae tendineae cordis ซึ่งเชื่อมต่อกล้ามเนื้อ papillaryกับลิ้นหัวใจ atrioventricularในหัวใจ การขาด Tnmd ในบริเวณนั้นจะนำไปสู่การเพิ่มขึ้นของ การสร้าง หลอดเลือดใหม่ การผลิต VEGF-Aและการกระตุ้น MMPs ตามมาด้วยการฉีกขาดของ cordis ซึ่งอาจทำให้เกิดภาวะลิ้นหัวใจไมทรัลรั่วและโรคลิ้นหัวใจ[ 17 ] [ 32 ]
- ในส่วนที่เกี่ยวกับการทำงานต้านการสร้างหลอดเลือดของ Tnmd ในร่างกาย ไม่พบความผิดปกติที่สำคัญในการสร้างและความหนาแน่นของหลอดเลือดในระหว่างการพัฒนาเอ็นและเรตินาในแบบจำลองหนูน็อคเอาท์[ 16 ]ผลการค้นพบหลังนี้ยังเป็นที่ถกเถียงกันอยู่ เพราะการศึกษาด้วยเทโนโมดูลินแบบรีคอมบิแนนท์แสดงให้เห็นผลการทำลายหลอดเลือดในเรตินาเมื่อฉีดเข้าไปในร่างกายส่วนวุ้นตาในร่างกาย[ 33 ]
- ในแบบจำลองเนื้องอกนอกตำแหน่งในร่างกาย การแสดงออกของ TNMD ที่ถูกเหนี่ยวนำใน เซลล์ มะเร็งผิวหนัง ของหนู ส่งผลให้การเจริญเติบโตของเนื้องอกถูกยับยั้งเนื่องจากความหนาแน่นของหลอดเลือดลดลง[ 34 ]
- การถ่ายทอด TNMD ในเซลล์เยื่อบุผนังหลอดเลือดดำของจอประสาทตาและสายสะดือของมนุษย์ส่งผลให้การเพิ่มจำนวนเซลล์หรือการเคลื่อนย้ายลดลงตามลำดับ[ 34 ]
- งานวิจัยหลายชิ้นเกี่ยวกับการจำแนกลักษณะเซลล์หลังจากการแสดงออกของยีนมากเกินไป การกระตุ้นด้วยปัจจัยการเจริญเติบโตหรือความเครียดทางกล การวิศวกรรมเนื้อเยื่อ และการประเมินวัสดุชีวภาพ ใช้การแสดงออกของยีน Tnmd เป็นเครื่องหมายบ่งชี้สายพันธุ์เซลล์เอ็นและเอ็นยึดกระดูก
- การวิจัยที่ดำเนินการในระดับจีโนมโดยใช้โพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยวได้นำเสนอความสัมพันธ์ที่น่าสนใจระหว่าง Tnmd และโรคต่างๆ ได้แก่โรคอ้วน [ 35 ]โรคเบาหวานประเภท 2 [ 35 ]กลุ่มอาการเมตาบอลิก[ 36 ]โรคอัลไซเมอร์[ 37 ]และโรคจอประสาทตาเสื่อมตามอายุ[ 38 ]ยังไม่เป็นที่ทราบแน่ชัดว่า SNP เหล่านี้ส่งผลต่อการถอดรหัส การตัดต่อ หรือลำดับกรดอะมิโนของโปรตีน Tnmd อย่างไร
- ความสัมพันธ์ที่แข็งแกร่งระหว่างการแสดงออกของ mRNA ของ Tnmd และความก้าวหน้าของโรคต่างๆ เช่น โรคอ้วน[ 21 ] [ 39 ]กลุ่มอาการเมตาบอลิก[ 40 ]และโรคผิวหนังอักเสบในเด็ก[ 41 ]ได้รับการแสดงให้เห็นแล้ว โดยทั่วไป ในทุกกรณีเหล่านี้ ระดับเทโนโมดูลินที่สูงขึ้นจะสอดคล้องกับภาวะโรคที่รุนแรงขึ้น
หมายเหตุ
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เทโนโมดูลิน
เทโนโมดู ลิ นหรือเรียกอีกอย่างว่าเทนดินไมโอดูลินTnmdหรือTeM เป็นโปรตีนที่เข้ารหัสโดยยีน TNMD (Tnmd) และถูกค้นพบโดยอิสระโดย Brandau และ Shukunami ในปี 2001...
โครงสร้างของยีนและโปรตีน
TNMD จัดอยู่ในกลุ่ม โปรตีนไกลโคโปรตีน ทรานส์เมมเบรนชนิด II กลุ่มใหม่ยีนนี้ตั้งอยู่บน โครโมโซม X และมีทรานสคริปต์ประมาณ 1.
รูปแบบการแสดงออก
TNMD มีการแสดงออกในระดับสูงทั้งในระดับสารส่งสัญญาณและโปรตีนในเอ็นและเส้นเอ็น แต่ก็พบได้ในเนื้อเยื่ออื่นๆ ด้วยเช่นกัน
เส้นทางการส่งสัญญาณที่คาดการณ์ไว้
เส้นทางการส่งสัญญาณที่คาดการณ์ไว้ของ TNMD ส่วนใหญ่ยังไม่เป็นที่รู้จักเนื่องจากพันธมิตรการจับโดยตรงที่ยังไม่ได้ รับการระบุ แบบจำลอง หนูน็อคเอาท์ หลายแบบ ที่มีฟีโนไทป์ของเอ็นช่วยให้เข้าใจว่าปัจจัยต้นน้ำหรือเส้นทางใดส่งผลต่อการแสดงออกของ Tnmd ในทำนองเดียวกัน...