อ่าน 11 นาที
ระบบการหลั่งของแบคทีเรีย
ระบบการหลั่งของแบคทีเรียเป็นโปรตีนเชิงซ้อนที่มีอยู่บนเยื่อหุ้มเซลล์ของแบคทีเรียเพื่อหลั่งสารต่างๆ โดยเฉพาะอย่างยิ่ง เป็นกลไกของเซลล์ที่แบคทีเรียก่อโรคใช้ในการหลั่งปัจจัยก่อโรค...
ระบบการหลั่งของแบคทีเรีย

ระบบการหลั่งของแบคทีเรียเป็นโปรตีนเชิงซ้อนที่มีอยู่บนเยื่อหุ้มเซลล์ของแบคทีเรียเพื่อหลั่งสารต่างๆ โดยเฉพาะอย่างยิ่ง เป็นกลไกของเซลล์ที่แบคทีเรียก่อโรคใช้ในการหลั่งปัจจัยก่อโรค (ส่วนใหญ่เป็นโปรตีน) เพื่อบุกรุกเซลล์เจ้าบ้าน สามารถจำแนกประเภทได้ตามโครงสร้าง องค์ประกอบ และกิจกรรมที่เฉพาะเจาะจง โดยทั่วไป โปรตีนสามารถหลั่งได้ผ่านกระบวนการสองแบบ กระบวนการหนึ่งเป็นกลไกแบบขั้นตอนเดียว ซึ่งโปรตีนจากไซโตพลาสซึมของแบคทีเรียจะถูกขนส่งและส่งตรงผ่านเยื่อหุ้มเซลล์เข้าไปในเซลล์เจ้าบ้าน อีกกระบวนการหนึ่งเกี่ยวข้องกับกิจกรรมสองขั้นตอน ซึ่งโปรตีนจะถูกขนส่งออกจากเยื่อหุ้มเซลล์ชั้นในก่อน จากนั้นสะสมในเพริพลาสซึมและสุดท้ายผ่านเยื่อหุ้มเซลล์ชั้นนอกเข้าไปในเซลล์เจ้าบ้าน[ 2 ]
ความแตกต่างที่สำคัญเหล่านี้สามารถแยกแยะได้ระหว่างแบคทีเรียแกรมลบแบบไดเดอร์ม และแบคทีเรียแกรมบวกแบบโมโนเดอร์มแต่การจำแนกประเภทนี้ไม่ได้ชัดเจนและสมบูรณ์แต่อย่างใด มีอย่างน้อยแปดประเภทที่เฉพาะเจาะจงสำหรับแบคทีเรียแกรมลบ สี่ประเภทสำหรับแบคทีเรียแกรมบวก ในขณะที่สองประเภทเป็นแบบทั่วไปสำหรับทั้งสองกลุ่ม[ 3 ]นอกจากนี้ ยังมีความแตกต่างที่เห็นได้ชัดระหว่างแบคทีเรียไดเดอร์มที่มีลิโปโพลีแซคคาไรด์บนเยื่อหุ้มชั้นนอก (ไดเดอร์ม-LPS) และแบคทีเรียที่มีกรดไมโคลิก (ไดเดอร์ม-ไมโคเลต) [ 4 ]
เส้นทางการส่งออก
เส้นทางการส่งออกมีหน้าที่ในการข้ามเยื่อหุ้มเซลล์ชั้นในในไดเดิร์ม และเป็นเยื่อหุ้มเซลล์เพียงแห่งเดียวในโมโนเดิร์ม[ 4 ]
ระบบรักษาความปลอดภัย
การหลั่งทั่วไป (Sec) เกี่ยวข้องกับการหลั่งโปรตีนที่ยังไม่พับตัวซึ่งในตอนแรกจะคงอยู่ภายในเซลล์ ในแบคทีเรียแกรมลบ โปรตีนที่หลั่งออกมาจะถูกส่งไปยังเยื่อหุ้มเซลล์ชั้นในหรือเพริพลาสม์ แต่ในแบคทีเรียแกรมบวก โปรตีนสามารถคงอยู่ในเซลล์หรือส่วนใหญ่จะถูกขนส่งออกจากแบคทีเรียโดยใช้ระบบการหลั่งอื่นๆ ในบรรดาแบคทีเรียแกรมลบEscherichia coli , Vibrio cholerae , Klebsiella pneumoniaeและYersinia enterocoliticaใช้ระบบ Sec ส่วนStaphylococcus aureusและListeria monocytogenesเป็นแบคทีเรียแกรมบวกที่ใช้ระบบ Sec [ 5 ]
ระบบSecใช้เส้นทางการหลั่งสองเส้นทางที่แตกต่างกัน ได้แก่ เส้นทาง SecA และ เส้นทาง อนุภาคการรับรู้สัญญาณ (SRP) SecA เป็นโปรตีนมอเตอร์ ATPase และมีโปรตีนที่เกี่ยวข้องหลายชนิด ได้แก่ SecD, SecE, SecF, SegG, SecM และ SecY SRP เป็นไรโบนิวคลีโอโปรตีน (โปรตีน-RNA คอมเพล็กซ์) ที่จดจำและกำหนดเป้าหมายโปรตีนเฉพาะไปยังเอนโดพลาสมิกเรติคูลัมในยูคาริโอตและไปยังเยื่อหุ้มเซลล์ในโปรคาริโอต เส้นทางทั้งสองต้องการโมเลกุลชาเปอโรนที่แตกต่างกัน และในที่สุดก็ใช้ช่องทางการขนส่งโปรตีน SecYEG สำหรับการขนส่งโปรตีนข้ามเยื่อหุ้มเซลล์ชั้นใน[ 6 ]ในเส้นทางที่ขึ้นอยู่กับ SecA นั้น SecB ทำหน้าที่เป็นชาเปอโรน โดยรักษาโปรตีนที่สังเคราะห์ขึ้นใหม่ให้อยู่ในสถานะที่คลี่ออก และส่งไปยัง SecA ที่จับกับทรานสโลคอน จากนั้นโปรตีนก่อนการหลั่งจะถูกหลั่งไปยังเพริพลาสม์ผ่านทรานสโลคอน SecYEG ในขณะที่ในเส้นทาง SRP นั้น SRP จะดึงดูดคอมเพล็กซ์ไรโบโซม-สายโซ่เกิดใหม่ (RNC) ไปยังเยื่อหุ้มเซลล์ในระหว่างการสังเคราะห์โปรตีน ในE. coliโปรตีนเยื่อหุ้มชั้นในส่วนใหญ่จะถูกกำหนดเป้าหมายโดยเส้นทาง SRP และโปรตีนเยื่อหุ้มชั้นนอกจะถูกกำหนดเป้าหมายโดยเส้นทาง SecA [ 7 ]อย่างไรก็ตาม การศึกษา โปรไฟล์ไรโบโซม แบบเลือกสรรเมื่อเร็ว ๆ นี้ ชี้ให้เห็นว่าโปรตีนเยื่อหุ้มชั้นในที่มีลูปเพริพลาสมิกขนาดใหญ่จะถูกกำหนดเป้าหมายโดยเส้นทาง SecA [ 8 ]
SecA หรือเส้นทางหลังการแปล
โปรตีนถูกสังเคราะห์ในไรโบโซมโดยกระบวนการเพิ่มกรดอะมิโนทีละตัว เรียกว่าการแปลในเส้นทาง SecA ปัจจัยกระตุ้นชาเปอโรน (TF) จะจับกับลำดับสัญญาณ N-terminal ที่เปิดเผยของสายเปปไทด์ก่อน เมื่อการยืดสายเปปไทด์ดำเนินต่อไป TF จะถูกแทนที่ด้วย SecB SecB จะรักษาสภาพของเปปไทด์ให้อยู่ในสถานะคลายตัวโดยเฉพาะ และช่วยในการจับของ SecA จากนั้นคอมเพล็กซ์สามารถจับกับ SecYEG ซึ่ง SecA จะถูกกระตุ้นโดยการจับกับ ATP ด้วยพลังงานจาก ATP SecA จะผลักโปรตีนผ่านช่อง secYEG คอมเพล็กซ์ SecD/F ยังช่วยในการดึงโปรตีนจากอีกด้านหนึ่งของเยื่อหุ้มเซลล์ด้วย[ 9 ]
นอกจากนี้ ยังมีการเสนอแนะว่าเส้นทาง SecA มีกลไกการกำหนดเป้าหมายแบบร่วมการแปล ซึ่งหมายความว่าโพลีเปปไทด์จะถูกกำหนดเป้าหมายโดยตรงโดย SecA ในระหว่างการสังเคราะห์[ 10 ]
เส้นทาง SRP
ในเส้นทางนี้ SRP จะแข่งขันกับ TF และจับกับลำดับสัญญาณ N-terminal โปรตีนจากเยื่อหุ้มชั้นในจะหยุดกระบวนการยืดสายโซ่ จากนั้น SRP จะจับกับตัวรับเยื่อหุ้มเซลล์ FtsY คอมเพล็กซ์สายโซ่เปปไทด์-SRP-FtsY จะถูกขนส่งไปยัง SecY ซึ่งการยืดสายโซ่เปปไทด์จะดำเนินต่อไป[ 11 ]
ระบบแทท
เส้นทางการขนส่งทวินอาร์จินีน (เส้นทาง Tat) คล้ายกับ Sec ในกระบวนการหลั่งโปรตีน อย่างไรก็ตาม มันส่งโปรตีนเฉพาะในสถานะพับ (ตติยภูมิ) เท่านั้น แบคทีเรียทุกชนิดใช้เส้นทางนี้ เช่นเดียวกับอาร์เคีย และคลอโรพลาสต์และไมโทคอนเดรียของพืช[ 12 ]ในแบคทีเรีย ระบบ Tat จะส่งออกโปรตีนจากไซโตพลาสซึมผ่านเยื่อหุ้มเซลล์ชั้นใน ในขณะที่ในคลอโรพลาสต์ ระบบนี้จะอยู่ใน เยื่อ ไทลา คอยด์ ซึ่งช่วยในการนำเข้าโปรตีนจากสโตรมา[ 13 ]โปรตีน Tat มีความแปรผันสูงในแบคทีเรียต่าง ๆ และถูกจัดประเภทเป็นสามประเภทหลัก ได้แก่ TatA, TatB และ TatC ตัวอย่างเช่น ในขณะที่มีโปรตีน Tat ที่ใช้งานได้เพียงสองชนิดในBacillus subtilis [ 14 ] แต่ ใน Streptomyces coelicolorอาจมีมากกว่าร้อยชนิด[ 15 ]เปปไทด์สัญญาณที่สามารถจดจำโปรตีน Tat ได้นั้นมีลักษณะเฉพาะด้วยโมทีฟที่เป็นฉันทามติ Ser/Thr-Arg-Arg-X-Phe-Leu-Lys (โดยที่ X สามารถเป็นกรดอะมิโนขั้วใดก็ได้) ชื่อการเคลื่อนย้ายอาร์จินีนคู่ (twin arginine translocation) มาจากอาร์จินีนสองตัวที่ต่อเนื่องกัน การแทนที่อาร์จินีนตัวใดตัวหนึ่งจะนำไปสู่การชะลอตัวหรือความล้มเหลวของการหลั่ง[ 16 ]
เส้นทาง Wss/Esx
เส้นทาง Wss/Esx ( ระบบ ESAT-6 ) บางครั้งเรียกว่าระบบการหลั่งประเภท VII (T7SS) แม้ว่าจะเป็นเส้นทางการส่งออกก็ตาม[ 4 ]มีอยู่ในแบคทีเรียแกรมบวก (ในรูป WSS) และ สายพันธุ์ Mycobacterium (ในรูป Esx ใน diderm-mycolates ทั้งหมด) เช่นM. tuberculosis , M. bovis , S. coelicolorและS. aureusนอกจากนี้ยังเรียกว่าระบบ T7b ในB. subtilisและS. aureusประกอบด้วยส่วนประกอบพื้นฐานสองส่วน ได้แก่ ATPase เฮกซาเมอร์ที่ยึดติดกับเยื่อหุ้มเซลล์ซึ่งเป็นสมาชิกของตระกูลโปรตีนFtsK /SpoIIIE [ 17 ]และโปรตีนที่เกี่ยวข้องกับ EsxA/EsxB ใดๆ เช่น EsaA, EsaD, EsxB, EsxD รวมถึงระบบ Ess (EssA, EssB และ EsxC ที่พบในS. aureus ) [ 18 ] EsxA และ EsxB อยู่ในกลุ่ม โปรตีน WXG100 ขนาดใหญ่ ที่สร้างโครงสร้างเกลียวคู่แบบแฮร์พิน
ในS. aureusนั้น T7SS จะหลั่งสารพิษขนาดใหญ่ที่เรียกว่า EsaD ซึ่งเป็นสมาชิกของ เอนไซม์ นิวคลีเอส EsaD จะถูกทำให้ไม่เป็นอันตราย (ดีท็อกซ์) ในระหว่างการสังเคราะห์ทางชีวภาพด้วยความช่วยเหลือของสารต้านพิษ EsaG ที่เป็นคู่กัน จากนั้นคอมเพล็กซ์ EsaD-EsaG จะจับกับ EsaE ส่วนของ EsaE จะจับกับ EssC ซึ่งเป็นเอนไซม์ ATPase ของคอมเพล็กซ์ T7SS ในระหว่างการหลั่ง EsaG จะยังคงอยู่ในไซโตพลาสซึม และมีเพียง EsaD และ EsaE เท่านั้นที่ถูกหลั่งออกมาพร้อมกัน แต่ในบางสายพันธุ์ของS. aureusนั้น EsaD จะไม่ถูกผลิตขึ้น แต่จะมีการสร้างโปรตีนที่คล้าย EsaG สองสำเนาแทน ซึ่งอาจอธิบายการเกิดขึ้นของ T7SS ในสายพันธุ์ที่ไม่ก่อโรค เช่นB. subtilisและS. coelicolor [ 19 ]
ระบบการหลั่ง
ระบบการหลั่งมีหน้าที่ในการข้ามเยื่อหุ้มเซลล์ชั้นนอกหรือเยื่อหุ้มทั้งสองชั้นในไดเดิร์ม ระบบการตั้งชื่อในปัจจุบันใช้กับไดเดิร์ม-LPS เท่านั้น เนื่องจากไม่มีข้อมูลว่าแบคทีเรียไดเดิร์ม-ไมโคเลตใช้อะไรในการข้ามเยื่อหุ้มชั้นนอก[ 4 ]
ประเภทที่ 1

ระบบการหลั่งแบบ Type I (T1SS หรือ TOSS) พบในแบคทีเรียแกรมลบ ระบบนี้อาศัยการทำงานของโปรตีนช่วยพับ (chaperone protein) โดยใช้โปรตีน Hly และ Tol ระบบจะทำงานเมื่อลำดับสัญญาณ HlyA จับกับ HlyB บนเยื่อหุ้มเซลล์ ลำดับสัญญาณนี้เป็นตัวขนส่งแบบ ABCคอมเพล็กซ์ HlyAB จะกระตุ้น HlyD ซึ่งจะคลายตัวและเคลื่อนไปยังเยื่อหุ้มเซลล์ชั้นนอก สัญญาณสุดท้ายจะถูกจดจำโดย TolC ในเยื่อหุ้มเซลล์ชั้นใน HlyA จะถูกหลั่งออกจากเยื่อหุ้มเซลล์ชั้นนอกผ่านช่องโปรตีนที่มีลักษณะคล้ายอุโมงค์
T1SS ขนส่งโมเลกุลต่างๆ รวมถึงไอออน คาร์โบไฮเดรต ยา และโปรตีน โมเลกุลที่หลั่งออกมามีขนาดแตกต่างกัน ตั้งแต่ เปปไทด์โคลิซิน V ของ E. coli ขนาดเล็ก ซึ่งมีขนาด 10 kDa ไปจนถึง โปรตีนยึดเกาะเซลล์ LapA ของ Pseudomonas fluorescensซึ่งมีขนาด 520 kDa [ 20 ]ในบรรดาโมเลกุลที่รู้จักกันดีที่สุด ได้แก่สารพิษ RTXและเอนไซม์ไลเปส
ประเภท II

ระบบการหลั่งแบบ Type II (T2SS) ขึ้นอยู่กับระบบ Sec หรือ Tat สำหรับการหลั่งเริ่มต้นภายในเซลล์แบคทีเรีย จากเพริพลาสม์ โปรตีนจะถูกหลั่งออกมาจากซีเครตินของเยื่อหุ้มเซลล์ชั้นนอก ซีเครตินเป็นคอมเพล็กซ์ของโปรตีนที่สร้างรูพรุนแบบมัลติเมอร์ (12–14 หน่วยย่อย) ซีเครตินได้รับการสนับสนุนจากโปรตีนเยื่อหุ้มเซลล์ชั้นในและชั้นนอกอื่นๆ อีก 10–15 ชนิดเพื่อประกอบเป็นอุปกรณ์การหลั่งที่สมบูรณ์[ 21 ]
ประเภท III

ระบบการหลั่งแบบ Type III (T3SS หรือ TTSS) มีโครงสร้างคล้ายคลึงและเกี่ยวข้องกับฐานของแฟลเจลลาของแบคทีเรียพบได้ในแบคทีเรียแกรมลบที่มีความรุนแรงสูงบางชนิด เช่นSalmonella , Shigella , YersiniaและVibrioโดยใช้ในการฉีดโปรตีนที่เป็นพิษเข้าไปในเซลล์ยูคาริโอต โครงสร้างของ T3SS มักถูกอธิบายว่าเป็นอินเจกติโซมหรืออุปกรณ์คล้ายเข็ม/กระบอกฉีดยา ค้นพบในYersinia pestisพบว่า T3SS สามารถฉีดสารพิษโดยตรงจากไซโตพลาสซึมของแบคทีเรียเข้าไปในไซโตพลาสซึมของเซลล์โฮสต์ได้[ 22 ]
ประเภท IV

ระบบการหลั่งแบบ Type IV (T4SS หรือ TFSS) เกี่ยวข้องกับ ระบบ การถ่ายทอดยีนของแบคทีเรียซึ่งแบคทีเรียต่างชนิดกันสามารถแลกเปลี่ยน DNA ของกันได้ แบคทีเรียที่เข้าร่วมอาจเป็นแบคทีเรียแกรมลบชนิดเดียวกันหรือต่างชนิดกันก็ได้ ระบบนี้สามารถขนส่งโปรตีนเดี่ยว รวมถึงโปรตีน-โปรตีน และ DNA-โปรตีนเชิงซ้อน การหลั่งจะถูกถ่ายโอนโดยตรงจากเซลล์ผู้รับผ่านเยื่อหุ้มเซลล์Agrobacterium tumefaciensซึ่งเป็นแบคทีเรียที่ค้นพบระบบนี้เป็นครั้งแรก ใช้ระบบนี้ในการส่งส่วน T-DNA ของพลาสมิด Ti เข้าสู่เซลล์พืช ส่งผลให้เกิดปุ่มราก (เนื้องอก) ขึ้นHelicobacter pyloriใช้ระบบนี้ในการส่งCagAเข้าสู่เซลล์เยื่อบุผิวของกระเพาะอาหาร เพื่อกระตุ้นให้เกิดมะเร็งกระเพาะอาหาร[ 23 ] Bordetella pertussisซึ่งเป็นแบคทีเรียที่เป็นสาเหตุของโรคไอกรุน หลั่งสารพิษเพอร์ทัสซิสบางส่วนผ่าน T4SS Legionella pneumophilaที่ทำให้เกิดโรคเลจิโอเนลโลซิส (โรคเลจิโอเนลลา) มีระบบหลั่ง T4SS ที่เรียกว่า icm/dot ( ยีนการขนส่ง/การคูณภายในเซลล์ ที่ บกพร่อง) ซึ่งขนส่งโปรตีนแบคทีเรียหลายชนิดเข้าสู่โฮสต์ยูคาริโอต[ 24 ] เมื่อไม่นานมานี้ พบว่าเชื้อก่อโรคพืชXanthomonas citriใช้ระบบหลั่ง T4SS ในการหลั่งสารออกฤทธิ์ที่ทำให้แบคทีเรียชนิดอื่นตายได้ จึงทำให้ระบบนี้เป็นตัวกำหนดความเหมาะสมที่สำคัญในการแข่งขันระหว่างแบคทีเรียต่างสายพันธุ์[ 25 ] [ 26 ] ระบบหลั่งแบบ Type IVA ต้นแบบคือคอมเพล็กซ์ VirB ของAgrobacterium tumefaciens [ 27 ]
ประเภท V
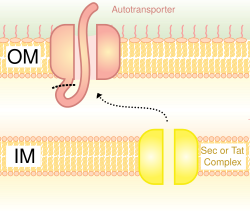
ระบบการหลั่งแบบ Type V (T5SS) แตกต่างจากระบบการหลั่งอื่นๆ ตรงที่มันหลั่งตัวเองและเกี่ยวข้องเฉพาะเยื่อหุ้มเซลล์ชั้นนอกเท่านั้น ในการที่โปรตีนที่หลั่งออกมาจะผ่านเยื่อหุ้มเซลล์ชั้นในได้ T5SS ต้องอาศัยระบบ Sec ซึ่งมีโดเมน β-barrel ที่แทรกเข้าไปในเยื่อหุ้มเซลล์ชั้นนอกและสร้างช่องทางที่สามารถขนส่งโปรตีนที่หลั่งออกมาพร้อมกับมันได้ ด้วยกิจกรรมนี้ พวกมันจึงถูกเรียกว่าระบบ autotransporter ด้วยเช่นกัน[ 28 ]เมื่อโปรตีนที่หลั่งออกมาถูกเปิดเผยสู่ภายนอก autotransporter จะถูกตัดออก (cleaved) ทำให้โปรตีนหลุดออกจากโดเมน β-barrel ตัวอย่างของ autotransporter คือadhesins ซึ่งเป็น autotransporter แบบไตรเมอร์[ 29 ]
ประเภท VI
ระบบการหลั่งแบบ Type VI (T6SS) ถูกค้นพบโดยทีมของJohn Mekalanosที่ Harvard Medical School ในปี 2006 จากVibrio choleraeและPseudomonas aeruginosa [ 30 ] [ 31 ] ระบบนี้ได้รับการยอมรับเมื่อการกลายพันธุ์ใน ยีน HcpและVrgG ของ Vibrio Cholerae ทำให้ความรุนแรงและความสามารถในการก่อโรคลดลง[ 32 ] [ 33 ]นอกเหนือจากบทบาทดั้งเดิมในฐานะปัจจัยก่อโรคแล้ว T6SS ยังมีส่วนเกี่ยวข้องในการป้องกันผู้ล่าแบบยูคาริโอตอย่างง่าย และในปฏิสัมพันธ์ระหว่างแบคทีเรียด้วยกัน[ 34 ] [ 35 ]ยีนสำหรับ T6SS ก่อตัวเป็นกลุ่มยีนที่ประกอบด้วยยีนมากกว่า 15 ยีน โดยยีน HcpและVgrGเป็นยีนที่พบได้ทั่วไปมากที่สุด ความคล้ายคลึงกันทางโครงสร้างของ T6SS กับหนามหางของฟาจ T4 บ่งชี้ว่ากระบวนการติดเชื้อนั้นคล้ายคลึงกับของฟาจ[ 36 ]
ประเภท VII

T7SS ของแบคทีเรีย diderm-LPS คือเส้นทางchaperone-usher [ 4 ]
ในแบคทีเรียไดเดอร์ม-ไมโคเลต ระบบการหลั่งนี้คือESAT- 6 [ 4 ]
ประเภทที่ 8
T8SS ของแบคทีเรีย diderm-LPS คือเส้นทางการตกตะกอนแบบนิวเคลียสภายนอกเซลล์[ 4 ] [ 37 ]

ประเภท IX
ระบบการหลั่งแบบ Type IX (T9SS) พบได้ทั่วไปใน แบคทีเรียสายพันธุ์ Fibrobacteres-Chlorobi-Bacteroidetesซึ่งสมาชิกในกลุ่มประกอบด้วยเยื่อหุ้มชั้นนอก ระบบนี้มีส่วนเกี่ยวข้องกับการเคลื่อนที่แบบเลื่อนชนิดหนึ่ง การกำหนดเป้าหมายปัจจัยก่อโรคบางชนิดไปยังพื้นผิวเซลล์อย่างเหมาะสม และการย่อยสลายสารประกอบของไบโอโพลีเมอร์ [ 39 ] T9SSยังเป็นที่รู้จักในชื่อการหลั่ง Por ( การสะสม พอร์ไฟรินบนพื้นผิวเซลล์) [ 4 ]ตามชื่อเชื้อก่อโรคในช่องปากPorphyromonas gingivalisมีการอธิบายส่วนประกอบโครงสร้างของระบบอย่างน้อยสิบหกส่วน รวมถึง PorU ซึ่งเป็นทรานส์เปปติเดสที่คัดแยกโปรตีนซึ่งจะกำจัดสัญญาณการคัดแยกที่ปลาย C ออกจากโปรตีนเป้าหมายและทำหน้าที่เป็นตัวกลางในการยึดเกาะกับลิโปโพลีแซคคาไรด์แทน
ประเภท XI
ระบบการหลั่งแบบ Type XI (T11SS) เป็นระบบการหลั่งของแบคทีเรียที่มีกระบวนการหลั่งสองขั้นตอน ขั้นแรกจะใช้ทรานสโลคอน Sec ในการขนส่งโปรตีนเฉพาะ ซึ่งส่วนใหญ่เป็นโฮโมล็อกของเฮโมฟิลิน เข้าสู่เพริพลาสม์ในสถานะที่คลายตัว จากนั้นโปรตีนเหล่านี้จะทำปฏิกิริยากับตัวขนส่งเยื่อหุ้มเซลล์ด้านนอกของ T11SS ซึ่งช่วยอำนวยความสะดวกในการหลั่งขั้นสุดท้าย[ 40 ]โฮโมล็อกของเฮโมฟิลินมีบทบาทสำคัญในการรับฮีมของแบคทีเรียโดยการจับฮีมจากเฮโมโปรตีนของโฮสต์และถ่ายโอนไปยังตัวรับที่ขึ้นอยู่กับ TonB เพื่อการดูดซึมธาตุเหล็ก ความจำเพาะของการหลั่งของ T11SS ได้รับอิทธิพลจากโดเมน β-barrel ปลาย C ของเฮโมฟิลิน ทำให้มั่นใจได้ว่าพวกมันจะถูกนำทางผ่านเส้นทางการหลั่งที่ถูกต้อง ด้วยการทำให้การรับธาตุเหล็กมีประสิทธิภาพ T11SS จึงมีส่วนช่วยในการอยู่รอดและการก่อโรคของแบคทีเรีย[ 41 ]
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ระบบการหลั่งของแบคทีเรีย
ระบบการหลั่งของแบคทีเรียเป็นโปรตีนเชิงซ้อนที่มีอยู่บนเยื่อหุ้มเซลล์ของแบคทีเรียเพื่อหลั่งสารต่างๆ โดยเฉพาะอย่างยิ่ง เป็นกลไกของเซลล์ที่แบคทีเรียก่อโรคใช้ในการหลั่งปัจจัยก่อโรค...
เส้นทางการส่งออก
เส้นทางการส่งออกมีหน้าที่ในการข้ามเยื่อหุ้มเซลล์ชั้นในในไดเดิร์ม และเป็นเยื่อหุ้มเซลล์เพียงแห่งเดียวในโมโนเดิร์ม [ 4 ]
ระบบรักษาความปลอดภัย
การหลั่งทั่วไป (Sec) เกี่ยวข้องกับการหลั่งโปรตีนที่ยังไม่พับตัวซึ่งในตอนแรกจะคงอยู่ภายในเซลล์ ในแบคทีเรียแกรมลบ โปรตีนที่หลั่งออกมาจะถูกส่งไปยังเยื่อหุ้มเซลล์ชั้นในหรือเพริพลาสม์ แต่ในแบคทีเรียแกรมบวก...
ระบบแทท
เส้นทาง การขนส่งทวินอาร์จินีน (เส้นทาง Tat) คล้ายกับ Sec ในกระบวนการหลั่งโปรตีน อย่างไรก็ตาม มันส่งโปรตีนเฉพาะในสถานะพับ (ตติยภูมิ) เท่านั้น แบคทีเรียทุกชนิดใช้เส้นทางนี้ เช่นเดียวกับอาร์เคีย และคลอโรพลาสต์และไมโทคอนเดรียของพืช [ 12 ] ในแบคทีเรีย ระบบ Tat...