อ่าน 8 นาที
xDNA
xDNA (หรือที่รู้จักกันในชื่อ expanded DNA หรือ benzo-homologated DNA ) เป็น ระบบ นิวคลีโอไทด์ ที่มีขนาดขยายใหญ่ขึ้น ซึ่ง สังเคราะห์ขึ้นจากการรวมกันของ วงแหวนเบนซีน และเบสธรรมชาติ...
xDNA
 |
| อะดีนีนที่ถูกทำให้เป็นโฮโมโลจเบนโซ |
|---|
 |
| ไทมีนที่ถูกดัดแปลงด้วยเบนโซ |
 |
| เบนโซ-โฮโมโลเกตไซโตซีน |
 |
| กัวนีนที่ถูกทำให้เป็นโฮโมโลจด้วยเบนโซ |
xDNA (หรือที่รู้จักกันในชื่อexpanded DNAหรือbenzo-homologated DNA ) เป็น ระบบ นิวคลีโอไทด์ ที่มีขนาดขยายใหญ่ขึ้น ซึ่ง สังเคราะห์ขึ้นจากการรวมกันของวงแหวนเบนซีนและเบสธรรมชาติ 4 ชนิด ได้แก่อะดีนีนกั ว นีนไซโตซีนและไทมีน [ 1 ] การขยายขนาดนี้ทำให้เกิดตัวอักษร 8 ตัว ซึ่งมีความจุในการจัดเก็บข้อมูล มากกว่าตัวอักษร 4 ตัว ของ DNAธรรมชาติ(มักเรียกว่า B-DNA ในเอกสาร) [ 2 ]เช่นเดียวกับการจับคู่เบส ปกติ A จับคู่กับ xT, C จับคู่กับ xG, G จับคู่กับ xC และ T จับคู่กับ xA ดังนั้น เกลียวคู่จึงกว้างกว่าเกลียวคู่ธรรมชาติ 2.4 Å [ 3 ] [ 4 ]แม้ว่าจะมีโครงสร้างคล้ายกับ B-DNA แต่ xDNA มีคุณสมบัติการดูดซับ การเรืองแสง และการเรียงซ้อนที่เป็นเอกลักษณ์[ 5 ] [ 6 ] [ 7 ]
เบนโซ-โฮโมโลเกตอะดีนีน ถูกสังเคราะห์ขึ้นครั้งแรกโดยกลุ่มของเนลสัน เจ. เลียวนาร์ด ในฐานะโพรบเอนไซม์ ต่อมา กลุ่มของเอริค ที. คูล ได้สังเคราะห์ เบส ขยายอีกสามชนิดที่เหลือ จนเสร็จสมบูรณ์ ตามมาด้วย yDNA ("wide" DNA) ซึ่งเป็นระบบนิวคลีโอไทด์เบนโซ-โฮโมโลเกตอีกระบบหนึ่ง และ xxDNA และ yyDNA แนฟโท -โฮโมโลเกต xDNA มีความเสถียรมากกว่า DNA ทั่วไปเมื่อได้รับอุณหภูมิสูง และถึงแม้ว่าจะมีสาย xDNA, yDNA, xxDNA และ yyDNA ที่สมบูรณ์อยู่แล้ว แต่ในปัจจุบันยังยากที่จะสังเคราะห์และบำรุงรักษา การทดลองกับ xDNA ให้ข้อมูลเชิงลึกใหม่เกี่ยวกับพฤติกรรมของ B-DNA ตามธรรมชาติ เบสขยาย xA, xC, xG และ xT มีคุณสมบัติเรืองแสง ตามธรรมชาติ และสายเดี่ยวที่ประกอบด้วยเบสขยายเท่านั้นสามารถจดจำและจับกับสายเดี่ยวของ DNA ตามธรรมชาติได้ ทำให้เป็นเครื่องมือที่มีประโยชน์สำหรับการศึกษาระบบชีวภาพ[ 3 ] [ 8 ] xDNA มักเกิดขึ้นจากการจับคู่เบสระหว่างนิวคลีโอเบส ธรรมชาติและนิวคลีโอเบสที่ขยาย อย่างไรก็ตาม นิวคลีโอเบส x ก็สามารถจับคู่กันได้เช่นกัน[ 5 ]งานวิจัยในปัจจุบันสนับสนุน xDNA ในฐานะระบบการเข้ารหัสทางพันธุกรรมที่ใช้งานได้ในอนาคตอันใกล้[ 4 ]
ต้นกำเนิด
นิวคลี โอไทด์ตัวแรกที่ถูกขยายคือพิวรีนอะดีนีน เนลสัน เจ. เลียวนาร์ดและเพื่อนร่วมงานได้สังเคราะห์นิวคลีโอไทด์ x ดั้งเดิมนี้ ซึ่งเรียกว่า "อะดีนีนที่ขยาย" xA ถูกใช้เป็นโพรบในการตรวจสอบตำแหน่งที่ใช้งานของเอนไซม์ที่ขึ้นอยู่กับATPโดยเฉพาะอย่างยิ่งการดัดแปลงที่ ซับสเตรต สามารถรับได้ในขณะที่ยังคงทำงานได้[ 8 ] [ 9 ]เกือบสองทศวรรษต่อมา เบสอีกสามตัวได้รับการขยายและรวมเข้ากับเกลียวคู่ ได้สำเร็จในภายหลัง โดยเอริค ที. คูลและเพื่อนร่วมงาน เป้าหมายของพวกเขาคือการสร้างระบบพันธุกรรมสังเคราะห์ที่เลียนแบบและเหนือกว่าการทำงานของระบบพันธุกรรมตามธรรมชาติ[ 10 ]และเพื่อขยายการใช้งานของDNAทั้งในเซลล์ที่มีชีวิตและในชีวเคมี เชิงทดลอง เมื่อสร้างชุดเบสที่ขยายแล้ว เป้าหมายก็เปลี่ยนไปเป็นการระบุหรือพัฒนาเอนไซม์การจำลองแบบที่ถูกต้องและเพิ่มประสิทธิภาพตัวอักษร DNA ที่ขยายต่อไป[ 8 ]
สังเคราะห์
ในพิวรีนที่ถูกเบนโซโฮโมโลเกต (xA และ xG) วงแหวนเบนซีนจะเชื่อมต่อกับเบสที่มีไนโตรเจนผ่านพันธะไนโตรเจน-คาร์บอน (NC) ไพริมิดีนที่ถูกเบนโซโฮโมโลเกตจะเกิดขึ้นผ่านพันธะคาร์บอน-คาร์บอน (CC) ระหว่างเบสและเบนซีน[ 3 ]จนถึงปัจจุบัน นิวคลีโอเบส x ได้ถูกเพิ่มเข้าไปในสาย DNA โดยใช้ อนุพันธ์ ฟอสโฟราไมไดต์ เนื่องจากพอ ลิเมอ เร สแบบดั้งเดิม ไม่ประสบความสำเร็จในการสังเคราะห์สาย xDNA นิวคลีโอไทด์ x เป็นตัวเลือกที่ไม่ดีในฐานะสารตั้งต้นสำหรับพอลิเมอเรส B-DNA เนื่องจากขนาดของมันรบกวนการจับที่โดเมนเร่ง ปฏิกิริยา ความพยายามในการใช้ เอนไซม์ ที่ไม่ขึ้นกับแม่แบบประสบความสำเร็จ เนื่องจากมีข้อจำกัดทางเรขาคณิตที่ลดลงสำหรับสารตั้งต้น เทอร์มินัลดีออกซีนิวคลีโอไทด์ทรานสเฟอเรส (TdT) เคยถูกนำมาใช้ก่อนหน้านี้เพื่อสังเคราะห์สายของเบสที่เชื่อมต่อกับฟลูออโรฟอร์ การใช้TdT ทำให้สามารถรวม โมโนเมอร์ได้มากถึง 30 ตัว เพื่อสร้างเกลียวคู่ของ xDNA อย่างไรก็ตาม xDNA แบบโอลิโกเมอร์ นี้ ดูเหมือนจะยับยั้งการขยายตัวเกินความยาวนี้เนื่องจากพันธะไฮโดรเจนที่มากเกินไป เพื่อลดการยับยั้ง xDNA สามารถไฮบริดเป็นเกลียวปกติได้[ 7 ] [ 11 ]
การจำลองแบบ
เพื่อให้ xDNA สามารถใช้เป็นโครงสร้างทดแทนสำหรับการจัดเก็บข้อมูลได้ จำเป็นต้องมีกลไกการจำลองแบบที่เชื่อถือได้ การวิจัยเกี่ยวกับการจำลองแบบ xDNA โดยใช้Klenow fragmentจากDNA polymerase Iแสดงให้เห็นว่าคู่เบสตามธรรมชาติจะถูกเพิ่มเข้ามาอย่างเลือกสรรในกรณีของการแทรกนิวคลีโอไทด์เดี่ยว อย่างไรก็ตามDNA polymerase IV (Dpo4) สามารถใช้ xDNA สำหรับการแทรกประเภทนี้ได้อย่างประสบความสำเร็จด้วยความแม่นยำสูง ทำให้เป็นตัวเลือกที่น่าสนใจสำหรับการวิจัยในอนาคตในการขยายการจำลองแบบของ xDNA [ 4 ]ความไวต่อความไม่ตรงกันของ xDNA คล้ายกับของB- DNA [ 2 ]
โครงสร้าง
 |  |  | 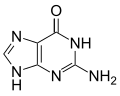 |
| อะดีนีน | ไทมีน | ไซโตซีน | กัวนีน |
 |  | 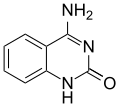 |  |
| ขนาดที่ขยาย xA | xT ที่ขยายขนาด | xC ขยายขนาด | xG ที่ขยายขนาด |
เช่นเดียวกับเบสตามธรรมชาติ นิวคลีโอไทด์ x จะประกอบกันเป็นโครงสร้างคู่ที่คล้ายกับ B-DNA [ 4 ]เดิมที xDNA ถูกสังเคราะห์โดยการรวมวงแหวนเบนซีนเข้ากับเบสที่มีไนโตรเจน อย่างไรก็ตาม เบสที่ขยายออกไปอื่นๆ ก็สามารถรวมไทโอฟีนและเบนโซ[b]ไทโอฟีนได้เช่นกัน xDNA และ yDNA ใช้วงแหวนเบนซีนเพื่อขยายเบส จึงเรียกว่า "เบนโซโฮโมโลเกต" นิวคลีโอเบสที่ขยายออกไปอีกรูปแบบหนึ่งที่เรียกว่า yyDNA จะรวมแนฟทาลีนเข้ากับเบสและเรียกว่า "แนฟโทโฮโมโลเกต" xDNA มีความยาวเพิ่มขึ้น 3.2 Åและมีการบิดตัว 32° ซึ่งเล็กกว่า B-DNA อย่างมีนัยสำคัญ โดย B-DNA มีความยาวเพิ่มขึ้น 3.3 Åและมีการบิดตัว 34.2° [ 3 ]นิวคลีโอไทด์ของ xDNA สามารถเกิดขึ้นได้บนทั้งสองสาย—ไม่ว่าจะอยู่เดี่ยวๆ (เรียกว่า "DNA ที่ขยายสองเท่า" [ 8 ] ) หรือผสมกับเบสตามธรรมชาติ—หรือเฉพาะบนสายใดสายหนึ่งเท่านั้น คล้ายกับ B-DNA xDNA สามารถจดจำและจับกับ ลำดับDNAหรือRNAสายเดี่ยวที่เสริมกันได้[ 2 ]
ดูเพล็กซ์ที่เกิดจาก xDNA มีลักษณะคล้ายกับดูเพล็กซ์ตามธรรมชาติยกเว้นระยะห่างระหว่างโครงสร้างน้ำตาลฟอสเฟตสองอัน เกลียว xDNA มีจำนวนคู่เบสต่อรอบของเกลียวมากกว่า อันเป็นผลมาจากระยะห่างระหว่างนิวคลีโอไทด์ข้างเคียงที่ลดลง สเปกตรัม NMR รายงานว่าเกลียว xDNA เป็นแบบแอนติพาราเลลมือขวาและมีโครงสร้างแอนติรอบพันธะไกลโคไซด์โดยมีน้ำตาล C2'-เอนโดที่โค้งงอ[ 5 ] [ 11 ]เกลียวที่สร้างจาก xDNA มีแนวโน้มที่จะมีโครงสร้างแบบ B-helix มากกว่า A-helix [ 2 ]และมีความกว้างของร่องหลักเพิ่มขึ้น 6.5 Å (บริเวณที่โครงสร้างอยู่ห่างกันมากที่สุด) และความกว้างของร่องรองลดลง 5.5 Å (บริเวณที่โครงสร้างอยู่ใกล้กันมากที่สุด) เมื่อเทียบกับB- DNA การเปลี่ยนแปลงความกว้างของร่องส่งผลต่อความสามารถของ xDNA ในการเชื่อมโยงกับโปรตีนที่จับกับ DNA [ 12 ]แต่ตราบใดที่นิวคลีโอไทด์ ที่ขยายออกไป นั้นอยู่เฉพาะในสายเดียว ตำแหน่งการจดจำจะมีความคล้ายคลึงกับB-DNA มากพอ ที่จะอนุญาตให้ปัจจัยการถอดรหัสและโมเลกุลโพลีอะไม ด์ขนาดเล็ก จับกันได้ เกลียวผสมทำให้มีความเป็นไปได้ที่จะจดจำเบสที่ขยายออกไปทั้งสี่โดยใช้โมเลกุลที่จับกับ DNA อื่นๆ[ 11 ]
คุณสมบัติ
นิวคลีโอไทด์ที่ขยายและเกลียวโอลิโกเมอร์ของพวกมันมีคุณสมบัติหลายอย่างร่วมกับ คู่ B-DNA ตามธรรมชาติ รวมถึงความชอบในการจับคู่: AกับT , CกับG [ 11 ] ความแตกต่างต่างๆ ในคุณสมบัติทางเคมีระหว่าง xDNA และB-DNAสนับสนุนสมมติฐานที่ว่าวงแหวนเบนซีนที่ขยายนิวคลีโอเบส x นั้นไม่ได้เฉื่อยทางเคมี[ 5 ] xDNA มีคุณสมบัติไม่ชอบน้ำมากกว่าB-DNA [ 7 ]และยังมีช่องว่าง HOMO–LUMO ที่เล็กกว่า (ระยะห่างระหว่างออร์บิทัลโมเลกุลที่ถูกครอบครองสูงสุดและออร์บิทัลโมเลกุลที่ว่างต่ำสุด) อันเป็นผลมาจากการอิ่มตัว ที่เปลี่ยนแปลง ไป[ 3 ] xDNA มีอุณหภูมิหลอมเหลวสูงกว่าB-DNA (เดคาเมอร์ผสมของ xA และ T มีอุณหภูมิหลอมเหลว 55.6 °C ซึ่งสูงกว่าเดคาเมอร์เดียวกันของ A และ T ถึง 34.3 °C [ 11 ] ) และแสดงพฤติกรรมการหลอมเหลวแบบ "ทั้งหมดหรือไม่มีเลย" [ 2 ]
การกำหนดค่า
ภายใต้สภาวะในห้องปฏิบัติการ xDNA จะวางตัวในรูปแบบsynน่าเสียดายที่การวางตัวแบบนี้ไม่ได้ทำให้หน้าสัมผัสของนิวคลีโอไทด์ xDNA หันไปทางสายข้างเคียงเพื่อการจับกัน ซึ่งหมายความว่าต้องใช้มาตรการเพิ่มเติมเพื่อเปลี่ยนแปลงโครงสร้างของ xDNA ก่อนที่จะพยายามสร้างเกลียว อย่างไรก็ตาม การวางตัว แบบ antiและsyn นั้น แทบจะเหมือนกันในเชิงพลังงานในเบสที่ขยายออก[ 9 ]ความชอบในการวางตัวนี้พบได้เป็นหลักในไพริมิดีนและพิวรีนแสดงความชอบในการวางตัวน้อยที่สุด[ 5 ]
การซ้อนที่ได้รับการปรับปรุง
การเรียงตัวของนิวคลีโอไทด์ในเกลียวคู่เป็นปัจจัยสำคัญที่กำหนดความเสถียรของเกลียว ด้วยพื้นที่ผิว ที่เพิ่มขึ้น และไฮโดรเจนที่พร้อมสำหรับการสร้างพันธะ ศักยภาพในการเรียงตัวของนิวคลีโอเบสจะเพิ่มขึ้นเมื่อมีการเพิ่มตัวคั่นเบนซีน การเพิ่มระยะห่างระหว่าง เบสไนโตรเจนและโครงสร้างน้ำตาล-ฟอสเฟตจะทำให้พลังงานการเรียงตัวของเกลียวมีความแปรปรวนน้อยลงและมีความเสถียรมากขึ้น พลังงานสำหรับคู่ของนิวคลีโอเบสตามธรรมชาติจะแตกต่างกันไปตั้งแต่ 18 ถึง 52 kJ/mol ความแปรปรวนนี้มีเพียง 14–40 kJ/mol สำหรับ xDNA [ 8 ]
เนื่องจากการทับซ้อนที่เพิ่มขึ้นระหว่างสายDNA ที่ขยาย และสายข้างเคียง ทำให้เกิดปฏิสัมพันธ์ระหว่างสายมากขึ้นในเกลียวที่ขยายและผสม ส่งผลให้ความเสถียรของเกลียวเพิ่มขึ้นอย่างมีนัยสำคัญ xDNA มีความสามารถในการเรียงซ้อนที่เพิ่มขึ้นอันเป็นผลมาจากการเปลี่ยนแปลงในพันธะไฮโดรเจน ระหว่างสายและภายในสาย ที่เกิดขึ้นจากการเพิ่ม ตัวคั่นเบน ซีนแต่การขยายเบสไม่ได้เปลี่ยนแปลงการมีส่วนร่วมของไฮโดรเจนต่อความเสถียรของคู่เกลียว ความสามารถในการเรียงซ้อนเหล่านี้ถูกใช้ประโยชน์โดยเกลียวที่ประกอบด้วยทั้ง xDNA และB-DNAเพื่อเพิ่มประสิทธิภาพความแข็งแรงของเกลียว การเรียงซ้อนที่เพิ่มขึ้นนั้นเห็นได้ชัดเจนที่สุดในสายที่ประกอบด้วยAและ xA และTและ xT เท่านั้น เนื่องจากT -xA มีปฏิสัมพันธ์การเรียงซ้อนที่แข็งแรงกว่า T - A [ 3 ]
พลังงานที่ได้จากไพริมิดีนมีค่าตั้งแต่ 30 ถึง 49 kJ/mol ส่วนพิวรีนมีค่าระหว่าง 40-58 kJ/mol การแทนที่นิวคลีโอไทด์หนึ่งตัวในเกลียวคู่ด้วยนิวคลีโอไทด์ที่ขยายขนาด จะทำให้ความแข็งแรงของปฏิกิริยาการเรียงซ้อนเพิ่มขึ้น 50% การขยายขนาดนิวคลีโอไทด์ทั้งสองตัวจะทำให้ความแข็งแรงของการเรียงซ้อนเพิ่มขึ้น 90% แม้ว่า xG จะมีผลเสียโดยรวมต่อความแข็งแรงของการจับยึดของเกลียว แต่เบสที่ขยายขนาดอีกสามตัวมีผลดีมากกว่า การเปลี่ยนแปลงพลังงานที่เกิดจากการขยายเบสส่วนใหญ่ขึ้นอยู่กับการหมุนของพันธะ รอบ จุดศูนย์กลาง มวล ของนิวคลีโอเบสและปฏิกิริยาการเรียงซ้อนของจุดศูนย์กลางมวลจะช่วยเพิ่มศักยภาพการเรียงซ้อนของเกลียว[ 5 ]เนื่องจากเบสที่ขยายขนาดทำให้เกลียวกว้างขึ้น จึงมีความเสถียรทางความร้อนมากขึ้นและมีอุณหภูมิหลอมเหลวสูงขึ้น[ 7 ]
การดูดซึม
การเพิ่ม ตัวเชื่อมเบน ซีนในนิวคลีโอเบส x ส่งผลต่อสเปกตรัม การ ดูดกลืนแสง ของเบส ทฤษฎีฟังก์ชันความหนาแน่นแบบขึ้นอยู่กับเวลา (TDDFT) ที่นำมาใช้กับ xDNA เผยให้เห็นว่าองค์ประกอบเบนซีนของออร์บิทัลโมเลกุลที่มีพลังงานสูงสุด ( HOMO ) ในเบส x ทำให้จุดเริ่มต้นของการดูดกลืนแสงเกิดขึ้นเร็วกว่าเบส ตามธรรมชาติ ลักษณะพิเศษอีกอย่างหนึ่งของสเปกตรัมการดูดกลืนแสงของ xDNA คือ เอ็กไซเมอร์ ของ xA ที่เลื่อนไปทางสีแดงในช่วงความถี่ต่ำ ในแง่ของลายนิ้วมือการเรียงซ้อน พบว่ามีภาวะไฮโปโครมิซิตี้ที่เด่นชัดกว่าในคู่เบส xA- T ที่ อยู่ติดกัน
ผลกระทบของการดูดซับที่เปลี่ยนแปลงไปของ xDNA รวมถึงการประยุกต์ใช้ในเทคโนโลยีนาโนอิเล็กทรอนิกส์และนาโนเทคโนโลยีชีวภาพระยะห่างที่ลดลงระหว่างนิวคลีโอไทด์ x ทำให้เกลียวแข็งขึ้น ดังนั้นจึงไม่ได้รับผลกระทบจากพื้นผิวอิเล็กโทรดและ แรงของ อนุภาคนาโน ที่มีฟังก์ชันได้ง่ายนัก การเปลี่ยนแปลงอื่นๆ ของ นิวคลีโอไทด์ตามธรรมชาติที่ส่งผลให้สเปกตรัมการดูดซับแตกต่างกันจะขยายขอบเขตการใช้งานเหล่านี้ในอนาคต[ 6 ]
การเรืองแสง
คุณสมบัติเฉพาะอย่างหนึ่งของ xDNA คือการเรืองแสง โดยธรรมชาติ เบสธรรมชาติสามารถจับกับฟลูออโรฟอร์ ได้โดยตรง เพื่อใช้ในไมโครอาร์เรย์การผสมแบบin situและ การวิเคราะห์ โพลีมอร์ฟิซึมอย่างไรก็ตาม เบสธรรมชาติที่เรืองแสงเหล่านี้มักจะล้มเหลวเนื่องจากการดับแสงด้วยตัวเองซึ่งทำให้ความเข้มของการเรืองแสงลดลงและลดความสามารถในการใช้งานเป็นแท็ก DNA ที่มองเห็นได้ปฏิสัมพันธ์ของไพน์ระหว่างวงแหวนในนิวคลีโอเบส x ส่งผลให้เกิดการเรืองแสง โดยธรรมชาติ ในช่วงสีม่วง-น้ำเงิน โดยมีค่าStokes shiftระหว่าง 50 ถึง 80 นาโนเมตร นอกจากนี้ยังมีผลผลิตควอนตัมอยู่ในช่วง 0.3–0.6 xC มีการปล่อยแสงเรืองแสงมากที่สุด[ 10 ] [ 7 ]
ฐานขยายอื่นๆ
หลังจากมีการสร้างและวิจัย xDNA จนประสบความสำเร็จ ก็มีการศึกษาค้นคว้าเกี่ยวกับนิวคลีโอไทด์ที่ขยายตัวในรูปแบบอื่นๆ เพิ่มเติม yDNA เป็นระบบนิวคลีโอไทด์ที่สองที่คล้ายคลึงกัน โดยใช้วงแหวนเบนซีนในการขยายฐานธรรมชาติ ทั้งสี่ xxDNA และ yyDNA ใช้แนฟทาลีน ซึ่งเป็น โมเลกุลโพลีไซคลิกที่ประกอบด้วย วงแหวน ไฮโดรคาร์บอน สอง วง วงแหวนทั้งสองนี้ช่วยขยายฐานให้กว้างขึ้นไปอีก ทำให้คุณสมบัติทางเคมีของฐานเปลี่ยนแปลงไปอีก
yDNA

ความสำเร็จและนัยสำคัญของ xDNA กระตุ้นให้เกิดการวิจัยเพื่อตรวจสอบปัจจัยอื่นๆ ที่อาจเปลี่ยนแปลงคุณสมบัติทางเคมีของB-DNA และสร้างระบบใหม่สำหรับการจัดเก็บข้อมูลที่มีการใช้งานที่กว้างขึ้น yDNA ก็ใช้ แหวนเบนซีนเช่นเดียวกับ xDNA โดยมีความแตกต่างเพียงอย่างเดียวคือตำแหน่งการเพิ่มแหวนอะโรมาติกตำแหน่งของ แหวน เบนซีนเปลี่ยนโครงสร้างที่ต้องการของเกลียวที่ขยายออก การเปลี่ยนแปลงโครงสร้างทำให้ yDNA มีความคล้ายคลึงกับB-DNA มากขึ้น ในแง่ของการวางตัวโดยการเปลี่ยนแปลงพันธะไฮโดรเจนระหว่างสาย ความเสถียรขึ้นอยู่กับการหมุนของเบสรอบจุดเชื่อมต่อระหว่างเบสและน้ำตาลของโครงสร้างหลัก การเปลี่ยนแปลงความชอบของ yDNA สำหรับการวางตัวนี้ทำให้มีความเสถียรโดยรวมมากกว่า xDNA ตำแหน่งของ ตัวคั่นเบน ซีนยังส่งผลต่อรูปทรงเรขาคณิตของร่องเบส ทำให้เกิดการเปลี่ยนแปลงปฏิสัมพันธ์ระหว่างเบสข้างเคียง คู่เบสระหว่างนิวคลีโอไทด์ y และนิวคลีโอไทด์ตามธรรมชาติเป็นระนาบ ไม่บิดเล็กน้อยเหมือนใน xDNA วิธีนี้ช่วยลดการยกตัวของเกลียว ให้ น้อยลงไปอีก เมื่อเทียบกับวิธี xDNA

แม้ว่า xDNA และ yDNA จะมีความคล้ายคลึงกันในคุณสมบัติส่วนใหญ่ รวมถึงปฏิกิริยาการเรียงซ้อนที่เพิ่มขึ้น แต่ yDNA แสดงความสามารถในการจดจำความไม่ตรงกันได้ดีกว่า ไพริมิดีน y แสดงปฏิกิริยาการเรียงซ้อนที่แข็งแรงกว่าไพริมิดีน x เล็กน้อย อันเป็นผลมาจากระยะห่างระหว่าง คาร์บอน อะโนเมอริก ทั้งสอง ซึ่งใน yDNA นั้นมีระยะห่างมากกว่าเล็กน้อย xDNA ยังคงมีปฏิกิริยาการเรียงซ้อนที่แข็งแรงกว่าในแบบจำลองเกลียว แต่การเพิ่มไพริมิดีน x หรือ y เข้าไปในเกลียวคู่ ตามธรรมชาติ จะเสริมความแข็งแรงของปฏิกิริยาภายในและระหว่างสาย ทำให้ความเสถียรของเกลียวโดยรวมเพิ่มขึ้น ในท้ายที่สุดแล้ว ว่าอันไหนมีปฏิกิริยาการเรียงซ้อนโดยรวมที่แข็งแรงที่สุดนั้นขึ้นอยู่กับลำดับของเบส xT และ yT จับกับAด้วยความแข็งแรงที่ใกล้เคียงกัน แต่พลังงานการเรียงซ้อนของ yC ที่จับกับGนั้นแข็งแรงกว่า xC ถึง 4 kJ/mol yDNA และเบสที่ขยายอื่นๆ เป็นส่วนหนึ่งของสาขาใหม่ที่ยังมีการศึกษาไม่มากนัก งานวิจัยชี้ให้เห็นว่าโครงสร้างที่เหมาะสมที่สุดยังคงต้องค้นพบ แต่การรู้ว่า ตำแหน่ง ของเบนซีนส่งผลต่อการวางแนวและโครงสร้างของนิวคลีโอเบสที่ขยายออกไปจะเพิ่มข้อมูลให้กับการออกแบบในอนาคต[ 8 ]
yyDNA และ xxDNA

นิวคลีโอเบส แบบขยายสองเท่า (หรือแบบแนฟโท-โฮโมโลเกต ) จะมีตัวเชื่อม แน ฟทาลีนแทนวงแหวนเบนซีนทำให้เบสมีความกว้างเพิ่มขึ้นเป็นสองเท่าด้วยโครงสร้างสองวงแหวน โครงสร้างเหล่านี้ (รู้จักกันในชื่อ xxDNA และ yyDNA) มีความกว้างกว่าเบสธรรมชาติ 4.8 Åและถูกสร้างขึ้นอีกครั้งจากการวิจัยของเลียวนาร์ดเกี่ยวกับการขยายตัวของอะดีนีนในเอนไซม์ที่ขึ้นอยู่กับATPในปี 1984 ไม่มีการตีพิมพ์เอกสารใดๆ เกี่ยวกับเบสแบบขยายสองเท่าเหล่านี้เป็นเวลากว่าสามทศวรรษ จนกระทั่งปี 2013 เมื่อ xxG ตัวแรกถูกสร้างขึ้นโดยชาร์มา ไลท์ และเวทมอร์ และรวมเข้ากับ xxA ในเกลียวธรรมชาติแม้ว่าจะมีการวิจัยเกี่ยวกับ xxDNA น้อยมาก แต่ ก็แสดงให้เห็นแล้วว่า พิวรีน xx ที่อยู่ใกล้เคียงจะเพิ่มพลังงานการเรียงซ้อนภายในสายได้มากถึง 119% (ตรงข้ามกับ 62% ในพิวรีน x) ปฏิสัมพันธ์ระหว่าง xx- purineและpyrimidineแสดงให้เห็นถึงการลดลงโดยรวมของพลังงานการเรียงซ้อน แต่ความเสถียรโดยรวมของคู่สายที่มี pyrimidine และ xx-purine เพิ่มขึ้น 22% ซึ่งมากกว่าสองเท่าของ pyrimidine และ x-purine [ 9 ]
การใช้งาน
xDNA มีการใช้งานมากมายในการวิจัยทางเคมีและชีววิทยา รวมถึงการขยายขอบเขตการใช้งานของDNA ตามธรรมชาติ เช่น การสร้างโครงสร้างค้ำยัน เพื่อสร้างโครงสร้างนาโนที่ประกอบตัวเองได้ จำเป็นต้องมีโครงสร้างค้ำยันเป็นเหมือนโครงตาข่ายเพื่อรองรับการเจริญเติบโต ในอดีตมีการใช้ DNA เป็นวิธีการเพื่อจุดประสงค์นี้ แต่โครงสร้างค้ำยันที่ขยายใหญ่ขึ้นทำให้สามารถสร้างโครงสร้างค้ำยันขนาดใหญ่ขึ้นสำหรับการประกอบตัวเองที่ซับซ้อนมากขึ้นได้[ 1 ] คุณสมบัติ การนำไฟฟ้าของ xDNA ยังทำให้มันเป็นตัวเลือกที่ดีเยี่ยมในฐานะสายไฟระดับโมเลกุลเนื่องจากปฏิกิริยา π-πช่วยให้มันนำไฟฟ้าได้อย่างมีประสิทธิภาพ[ 3 ]ตัวอักษร 8 ตัว ( A , T , C , G , xA, xT, xC, xG) ทำให้มันมีศักยภาพในการจัดเก็บ สถานะได้มากกว่า DNA ถึง 2 nเท่าต่อลำดับ โดยที่ nคือจำนวนเบสในลำดับ ตัวอย่างเช่น การรวมนิวคลีโอไทด์ 6 ตัวของกับB-DNAจะได้ลำดับที่เป็นไปได้ 4096 ลำดับ ในขณะที่การรวมนิวคลีโอไทด์จำนวนเท่ากันที่สร้างด้วย xDNA จะได้ลำดับที่เป็นไปได้ 262,144 ลำดับ นอกจากนี้ xDNA ยังสามารถใช้เป็นโพรบเรืองแสงที่ตำแหน่งออกฤทธิ์ ของเอนไซม์ ได้ เช่นเดียวกับการใช้งานดั้งเดิมโดย Leonard et al. [ 2 ]
xDNA ยังถูกนำไปใช้ในการศึกษาปฏิสัมพันธ์ระหว่างโปรตีนและ DNA ด้วย เนื่องจากคุณสมบัติการเรืองแสง ตามธรรมชาติของ xDNA ทำให้สามารถมองเห็นได้ง่ายทั้งในห้องปฏิบัติการและในสิ่งมีชีวิต [ 5 ] การสร้างและ การรวมตัวของ xDNA ทำได้ง่ายขึ้น และการจับกับลำดับ DNAและRNA ที่เสริมกัน ด้วยความสัมพันธ์สูงหมายความว่ามันไม่เพียงแต่ช่วยระบุตำแหน่งของลำดับเหล่านี้ที่ลอยอยู่รอบๆ เซลล์เท่านั้น แต่ยังรวมถึงเมื่อพวกมันมีปฏิสัมพันธ์กับโครงสร้างอื่นๆ ภายในเซลล์ด้วย[ 10 ] xDNA ยังมีศักยภาพในการประยุกต์ใช้ในการทดสอบที่ใช้TdTเนื่องจากอาจช่วยปรับปรุงรีพอร์เตอร์ และสามารถใช้เป็นแท็กความสัมพันธ์สำหรับพันธะระหว่างสายได้[ 7 ]
ดูเพิ่มเติม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ xDNA
xDNA (หรือที่รู้จักกันในชื่อ expanded DNA หรือ benzo-homologated DNA ) เป็น ระบบ นิวคลีโอไทด์ ที่มีขนาดขยายใหญ่ขึ้น ซึ่ง สังเคราะห์ขึ้นจากการรวมกันของ วงแหวนเบนซีน และเบสธรรมชาติ...
ต้นกำเนิด
นิวคลี โอไทด์ ตัวแรกที่ถูกขยายคือ พิวรีน อะดีนี น เนลสัน เจ. เลียวนาร์ดและเพื่อนร่วมงานได้สังเคราะห์นิวคลีโอไทด์ x ดั้งเดิมนี้ ซึ่งเรียกว่า "อะดีนีนที่ขยาย" xA ถูกใช้เป็นโพรบในการตรวจสอบ ตำแหน่งที่ใช้งาน ของ เอนไซม์ ที่ขึ้นอยู่กับ ATP...
สังเคราะห์
ในพิวรีนที่ถูกเบนโซโฮโมโลเกต (xA และ xG) วงแหวนเบนซีน จะเชื่อมต่อกับ เบสที่มีไนโตรเจน ผ่านพันธะไนโตรเจน-คาร์บอน (NC) ไพริมิดีนที่ถูกเบนโซโฮโมโลเกตจะเกิดขึ้นผ่านพันธะคาร์บอน-คาร์บอน (CC) ระหว่างเบสและเบนซีน [ 3 ] จนถึงปัจจุบัน นิวคลีโอเบส x...
การจำลองแบบ
เพื่อให้ xDNA สามารถใช้เป็นโครงสร้างทดแทนสำหรับการจัดเก็บข้อมูลได้ จำเป็นต้องมีกลไกการจำลองแบบที่เชื่อถือได้ การวิจัยเกี่ยวกับการจำลองแบบ xDNA โดยใช้ Klenow fragment จาก DNA polymerase I...