อ่าน 9 นาที
อาร์เอ็นเอไรโบโซม 5S
ไร โบโซมอลอาร์เอ็นเอ 5S ( 5S rRNA ) เป็น โมเลกุล ไรโบโซมอลอาร์เอ็นเอ ที่มีความยาวประมาณ 120 นิวคลีโอไทด์ และมีมวล 40 กิโลดาลตัน...
อาร์เอ็นเอไรโบโซม 5S
| อาร์เอ็นเอไรโบโซม 5S | |
|---|---|
 การคาดการณ์โครงสร้างทุติยภูมิและการอนุรักษ์ลำดับของ RNA ไรโบโซม 5S | |
| ตัวระบุ | |
| เครื่องหมาย | 5S_rRNA |
| อาร์แฟม | RF00001 CL00113 |
| ข้อมูลอื่นๆ | |
| ประเภทRNA | ยีน ; อาร์เอ็นเอ |
| โดเมน | ยูคาริโอต ; แบคทีเรีย ; อาร์เคีย |
| ไป | GO:0005840 GO:0003735 |
| ดังนั้น | SO:0000652 |
| โครงสร้างPDB | พีดีบี |
ไรโบโซมอลอาร์เอ็นเอ 5S ( 5S rRNA ) เป็น โมเลกุล ไรโบโซมอลอาร์เอ็นเอ ที่มีความยาวประมาณ 120 นิวคลีโอไทด์ และมีมวล 40 กิโลดาลตันเป็นส่วนประกอบโครงสร้างและหน้าที่ของหน่วยย่อยขนาดใหญ่ของไรโบโซมในสิ่งมีชีวิตทุกโดเมน ( แบคทีเรียอาร์เคียและยูคาริโอต ) ยกเว้นไรโบโซมไมโทคอนเดรียของเชื้อราและสัตว์ การกำหนด 5S หมายถึง ค่าสัมประสิทธิ์ การตกตะกอน ของโมเลกุล ในเครื่องอัลตราเซนตริฟิวจ์ ซึ่งวัดเป็นหน่วยสเวดเบิร์ก (S) [ 1 ]

การสังเคราะห์ทางชีวภาพ
ในโปรคาริโอต ยีน 5S rRNA มักจะอยู่ในโอเปรอน rRNA ถัดจาก rRNA หน่วยย่อยขนาดเล็กและขนาดใหญ่และถูกถอดรหัสร่วมกันเป็นสารตั้งต้นโพลีซิสโทรนิ ก[ 3 ]ลักษณะเฉพาะของจีโนมนิวเคลียร์ของยูคาริโอตคือการมีสำเนายีน 5S rRNA หลายชุด (5S rDNA) ที่รวมกลุ่มกันเป็นชุดซ้ำ โดยจำนวนสำเนาจะแตกต่างกันไปในแต่ละสปีชีส์[ 4 ] [ 5 ] 5S rRNA ของยูคาริโอตถูกสังเคราะห์โดยRNA polymerase IIIในขณะที่ rRNA อื่นๆ ของยูคาริโอตถูกตัดจากสารตั้งต้น 45S ที่ถอดรหัสโดยRNA polymerase Iในโอโอไซต์ของ Xenopusพบว่านิ้วที่ 4–7 ของปัจจัยการถอดรหัสแบบนิ้วสังกะสีเก้านิ้ว TFIIIA สามารถจับกับบริเวณตรงกลางของ 5S RNA ได้[ 6 ] [ 7 ]การจับกันระหว่าง 5S rRNA และ TFIIIA ทำหน้าที่ทั้งยับยั้งการถอดรหัสยีน 5S RNA เพิ่มเติม และทำให้ 5S RNA มีเสถียรภาพจนกว่าจะจำเป็นสำหรับการประกอบไรโบโซม[ 8 ]
โครงสร้าง
โครงสร้างทุติยภูมิของ 5S rRNA ประกอบด้วยเกลียวห้าเกลียว (ระบุด้วยเลขโรมัน I–V ) ห่วงสี่ห่วง (BE) และบานพับหนึ่งอัน (A) ซึ่งรวมกันเป็นโครงสร้างคล้ายตัว Y ห่วง C และ D เป็นแฮร์พินปลายสุด และห่วง B และ E เป็นแฮร์พินภายใน[ 4 ]จากการศึกษาทางวิวัฒนาการ เกลียว I และ III น่าจะเป็นบรรพบุรุษ[ 9 ]เกลียว III ประกอบด้วยอะดีโนซีนที่อนุรักษ์ไว้อย่างสูงสองตัว[ 10 ]เกลียว V ที่มีโครงสร้างแฮร์พิน เชื่อว่ามีปฏิสัมพันธ์กับ TFIIIA [ 4 ]
ตำแหน่งภายในไรโบโซม

การใช้เทคนิคทางโมเลกุลที่หลากหลาย รวมถึงกล้องจุลทรรศน์อิเล็กตรอนภูมิคุ้มกันกล้องจุลทรรศน์อิเล็กตรอนแช่แข็งการเชื่อมโยงข้ามทางเคมีระหว่าง โมเลกุล และผลึกศาสตร์รังสีเอกซ์ทำให้สามารถกำหนดตำแหน่งของ 5S rRNA ภายในหน่วยย่อยไรโบโซมขนาดใหญ่ได้อย่างแม่นยำ ในแบคทีเรียและอาร์เคียหน่วยย่อยไรโบโซมขนาดใหญ่ (LSU) นั้นประกอบด้วยโมเลกุล RNA สองโมเลกุล คือ 5S rRNA และ RNA ขนาดใหญ่อีกโมเลกุลหนึ่งที่เรียกว่า23S rRNAพร้อมด้วยโปรตีนที่เกี่ยวข้องจำนวนมาก[ 3 ] [ 12 ]
ในยูคาริโอต LSU ประกอบด้วย rRNA 5S, 5.8Sและ28Sและโปรตีนอีกมากมาย[ 13 ] [ 14 ] โครงสร้างของ LSU ใน 3 มิติแสดงให้เห็นพื้นผิวเรียบด้านหนึ่งและพื้นผิวตรงข้ามที่มีส่วนยื่น 3 ส่วน ได้แก่ ส่วนยื่น L1 ส่วนยื่นกลาง (CP) และก้าน L7/L12 ส่วนยื่น L1 และก้าน L7/L12 เรียงตัวอยู่ด้านข้างล้อมรอบ CP rRNA 5S ตั้งอยู่ใน CP และมีส่วนร่วมในการสร้างและโครงสร้างของส่วนยื่นนี้ ส่วนประกอบหลักอื่นๆ ของส่วนยื่นกลาง ได้แก่ rRNA 23S (หรือ 28S ในยูคาริโอต) และโปรตีนหลายชนิด ได้แก่ L5, L18, L25 และ L27 [ 15 ]
หน้าที่ของไรโบโซม
หน้าที่ที่แท้จริงของ 5S rRNA ยังไม่ชัดเจน ในEscherichia coliการลบยีน 5S rRNA จะลดอัตราการสังเคราะห์โปรตีนและมีผลเสียต่อสมรรถภาพของเซลล์มากกว่าการลบยีน rRNA อื่นๆ ( 16Sและ23S ) ในจำนวนสำเนาที่เทียบเท่ากัน [ 16 ] การศึกษาทางผลึกศาสตร์บ่งชี้ว่าโปรตีนที่จับกับ 5S rRNA และโปรตีนอื่นๆ ของส่วนยื่นตรงกลางของ LSU มีบทบาทในการจับกับ tRNA [ 15 ]นอกจากนี้ ความใกล้ชิดทางภูมิศาสตร์และทางกายภาพระหว่าง 5S rRNA และ 23S rRNA ซึ่งก่อตัวเป็นศูนย์เชื่อมโยงเปปทิดิลทรานสเฟอเรสและ GTPase บ่งชี้ว่า 5S rRNA ทำหน้าที่เป็นตัวกลางระหว่างศูนย์การทำงานทั้งสองของไรโบโซมโดยการสร้างสะพานเชื่อมระหว่างหน่วยย่อยและตำแหน่งการจับ tRNA ร่วมกับโปรตีนที่จับกับ 5S rRNA และส่วนประกอบอื่นๆ ของส่วนยื่นกลาง[ 15 ]
บทบาทในการประกอบไรโบโซม
ในยูคาริโอต ไรโบโซมในไซโตพลาสซึมประกอบขึ้นจาก rRNA สี่โมเลกุลและโปรตีนมากกว่า 80 โมเลกุล[ 14 ] [ 17 ] เมื่อถอดรหัสแล้ว ปลาย 3' ของ rRNA 5S จะถูกตัดแต่งให้มีความยาวที่สมบูรณ์ได้โดยโฮโมล็อกที่ทำหน้าที่ของRNase Tเท่านั้น เช่น Rex1p ในSaccharomyces cerevisiae [ 18 ] หน่วย ย่อยไรโบโซม 60Sและ40S จะถูกส่งออกจากนิวเคลียสไปยังไซโตพลาสซึม ซึ่งพวกมันจะรวมกันเพื่อสร้าง ไรโบโซม 80S ที่สมบูรณ์และ พร้อม สำหรับการแปลยังคงเป็นที่ถกเถียงกันอยู่ว่า 5S rRNA จะถูกรวมเข้ากับไรโบโซมเมื่อใด[ 4 ]แต่โดยทั่วไปเป็นที่ยอมรับกันว่า 5S rRNA จะถูกรวมเข้ากับอนุภาค 90S ซึ่งเป็นสารตั้งต้นของอนุภาค 60S โดยเป็นส่วนหนึ่งของคอมเพล็กซ์ RNP ขนาดเล็กที่ไม่ขึ้นกับไรโบโซมซึ่งเกิดจาก 5S rRNA และโปรตีนไรโบโซม L5 [ 17 ]
ปฏิสัมพันธ์กับโปรตีน
โปรตีนสำคัญหลายชนิดที่ทำปฏิกิริยากับ 5S rRNA มีรายชื่อดังต่อไปนี้
โปรตีนลา
ปฏิสัมพันธ์ของ 5S rRNA กับโปรตีน Laป้องกันไม่ให้ RNA ถูกย่อยสลายโดยเอ็กโซนิวคลีเอสในเซลล์[ 19 ] โปรตีน Laพบในนิวเคลียสในสิ่งมีชีวิตยูคาริโอตทั้งหมดและเชื่อมโยงกับ RNA หลายประเภทที่ถอดรหัสโดย RNA pol III โปรตีน La ทำปฏิกิริยากับ RNA เหล่านี้ (รวมถึง 5S rRNA) ผ่านทาง 3' oligo-uridine tract ช่วยให้ RNA มีความเสถียรและพับตัวได้[ 4 ] [ 20 ]
โปรตีน L5
ในเซลล์ยูคาริโอติก โปรตีนไรโบโซม L5 จะเชื่อมโยงและทำให้ 5S rRNA มีเสถียรภาพ โดยสร้างอนุภาคไรโบนิวคลีโอโปรตีน ก่อนไรโบโซม (RNP) ซึ่งพบได้ทั้งในไซโตโซลและนิวเคลียส การขาด L5 จะขัดขวางการขนส่ง 5S rRNA ไปยังนิวเคลียสและส่งผลให้การประกอบไรโบโซมลดลง[ 4 ]
โปรตีนไรโบโซมอื่นๆ
ในโปรคาริโอต 5S rRNA จะจับกับโปรตีนไรโบโซม L5, L18 และ L25 ในขณะที่ในยูคาริโอต 5S rRNA เป็นที่ทราบกันว่าจับกับโปรตีนไรโบโซม L5 เท่านั้น[ 21 ]ในT. brucei ซึ่งเป็นสาเหตุของโรคเหงาหลับ 5S rRNA จะมีปฏิสัมพันธ์กับโปรตีนที่จับกับ RNA ที่เกี่ยวข้องอย่างใกล้ชิดสองชนิด ได้แก่ P34 และ P37 ซึ่งการสูญเสียโปรตีนเหล่านี้ส่งผลให้ระดับ 5S rRNA โดยรวมลดลง[ 4 ]
การปรากฏตัวของไรโบโซมในออร์แกเนลล์
| จีโนมไมโทคอนเดรียที่เรียงลำดับใหม่ซึ่งเข้ารหัส 5S rRNA | |
|---|---|
| ตัวระบุ | |
| เครื่องหมาย | เอ็มทีเพอร์ม-5เอส |
| อาร์แฟม | RF02547 CL00113 |
| ข้อมูลอื่นๆ | |
| ประเภทRNA | ยีน ; อาร์เอ็นเอ |
| โดเมน | ยูคาริโอตา ; |
| ไป | GO:0005840 GO:0003735 |
| ดังนั้น | SO:0000652 |
| โครงสร้างPDB | พีดีบี |

กลไกการแปลของไมโตคอนเดรียและพลาสติด (ออร์แกเนลล์ที่มีต้นกำเนิดจากแบคทีเรียแบบเอนโดซิมไบโอติก) และแบคทีเรียที่เกี่ยวข้องมีลักษณะร่วมกันหลายอย่าง แต่ก็แสดงความแตกต่างที่ชัดเจนเช่นกัน จีโนมของออร์แกเนลล์เข้ารหัส SSU และ LSU rRNA โดยไม่มีข้อยกเว้น แต่การกระจายตัวของยีน 5S rRNA ( rrn5 ) นั้นไม่สม่ำเสมอมากrrn5สามารถระบุได้ง่ายและพบได้ทั่วไปในจีโนมของพลาสติดส่วนใหญ่ ในทางตรงกันข้ามrrn5 ของไมโตคอนเดรีย ในตอนแรกดูเหมือนจะจำกัดอยู่เฉพาะในพืชและโปรติสต์จำนวนเล็กน้อย[ 22 ] [ 23 ]นอกจากนี้ 5S rRNA ของออร์แกเนลล์ที่แตกต่างกันมากขึ้นจะถูกระบุได้เฉพาะด้วยแบบจำลองความแปรปรวนร่วมเฉพาะที่รวมข้อมูลเกี่ยวกับอคติขององค์ประกอบลำดับที่เด่นชัดและความแปรผันของโครงสร้าง[ 24 ]การวิเคราะห์นี้ระบุยีน 5S rRNA เพิ่มเติมไม่เพียงแต่ในจีโนมไมโทคอนเดรียของ สายพันธุ์ โปรติสต์ ส่วนใหญ่เท่านั้น แต่ยังรวมถึงในจีโนมของอะพิโคพลาสต์ บางชนิด (พลาสติดที่ไม่สังเคราะห์แสงของโปรโตซัวก่อโรค เช่นToxoplasma gondiiและEimeria tenella )
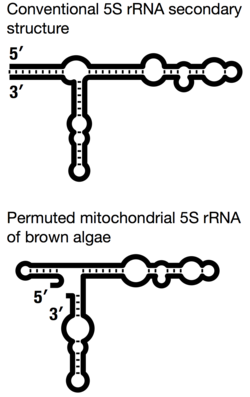
5S rRNA ของไมโทคอนเดรียในสเตรเมโนไพล์ ส่วนใหญ่ ประกอบด้วยโครงสร้างทุติยภูมิที่มีความหลากหลายมากที่สุด[ 24 ] 5S rRNA ของไมโทคอนเดรียที่สลับตำแหน่งในสาหร่ายสีน้ำตาลแสดงถึงกรณีที่ผิดปกติที่สุด โดยที่เกลียวปิด I ซึ่งปกติจะนำปลาย 5′ และ 3′ ของโมเลกุลมารวมกัน จะถูกแทนที่ด้วยแฮร์พิน (ปิด) ส่งผลให้เกิดจุดเชื่อมต่อสามทางแบบเปิด
หลักฐานปัจจุบันบ่งชี้ว่าดีเอ็นเอไมโทคอนเดรียของกลุ่มเพียงไม่กี่กลุ่ม โดยเฉพาะสัตว์เชื้อราอัล วี โอเลตและยูเกลโนโซแอนขาดจีนดังกล่าว[ 24 ]ส่วนนูนตรงกลางซึ่งปกติแล้วจะมี 5S rRNA และโปรตีนที่เกี่ยวข้องอยู่ (ดูรูปที่ 2 ) ได้รับการปรับเปลี่ยนในหลายวิธี ในไรโบโซมไมโทคอนเดรียของเชื้อรา 5S rRNA ถูกแทนที่ด้วยลำดับการขยายตัวของ LSU rRNA [ 25 ]ในคิเนโทพลาสติด (ยูเกลโนโซแอน) ส่วนนูนตรงกลางทำมาจากโปรตีนไรโบโซมไมโทคอนเดรียที่วิวัฒนาการขึ้นใหม่ทั้งหมด[ 26 ]สุดท้าย ไรโบโซมไมโทคอนเดรียของสัตว์ได้นำ tRNA ไมโทคอนเดรียเฉพาะ (Val ในสัตว์มีกระดูกสันหลัง) มาใช้แทน 5S rRNA ที่หายไป[ 27 ] [ 28 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
- หน้าเว็บสำหรับ RNA ไรโบโซม 5Sที่Rfam
- 5SData เก็บถาวรเมื่อ 2010-04-27 ที่Wayback Machine
- 5S+Ribosomal+RNA ใน หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- ตำแหน่งของยีน 5S_rRNAในมนุษย์ใน UCSC Genome Browser
- 5S rRNA ของ Halococcus morrhuae (อาร์คีแบคทีเรีย)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ อาร์เอ็นเอไรโบโซม 5S
ไร โบโซมอลอาร์เอ็นเอ 5S ( 5S rRNA ) เป็น โมเลกุล ไรโบโซมอลอาร์เอ็นเอ ที่มีความยาวประมาณ 120 นิวคลีโอไทด์ และมีมวล 40 กิโลดาลตัน...
การสังเคราะห์ทางชีวภาพ
ในโปรคาริโอต ยีน 5S rRNA มักจะอยู่ใน โอเปรอน rRNA ถัดจาก rRNA หน่วยย่อยขนาด เล็กและขนาดใหญ่และถูกถอดรหัสร่วมกันเป็น สารตั้งต้น โพลีซิสโทรนิ ก [ 3 ] ลักษณะเฉพาะของจีโนมนิวเคลียร์ของยูคาริโอตคือการมีสำเนายีน 5S rRNA หลายชุด (5S rDNA) ที่รวมกลุ่มกันเป็นชุดซ้ำ...
โครงสร้าง
โครงสร้าง ทุติยภูมิ ของ 5S rRNA ประกอบด้วยเกลียวห้าเกลียว (ระบุด้วย เลขโรมัน I–V ) ห่วงสี่ห่วง (BE) และบานพับหนึ่งอัน (A) ซึ่งรวมกันเป็นโครงสร้างคล้ายตัว Y ห่วง C และ D เป็นแฮร์พินปลายสุด และห่วง B และ E เป็นแฮร์พินภายใน [ 4 ] จากการศึกษาทางวิวัฒนาการ เกลียว...
ตำแหน่งภายในไรโบโซม
การใช้เทคนิคทางโมเลกุลที่หลากหลาย รวมถึง กล้องจุลทรรศน์อิเล็กตรอนภูมิคุ้มกัน กล้องจุลทรรศน์ อิเล็กตรอนแช่แข็ง การเชื่อมโยงข้ามทางเคมี ระหว่าง โมเลกุล และ ผลึกศาสตร์รังสีเอกซ์ ทำให้สามารถกำหนดตำแหน่งของ 5S rRNA ภายในหน่วยย่อยไรโบโซมขนาดใหญ่ได้อย่างแม่นยำ ใน...