อ่าน 13 นาที
ALOX12
ALOX12 ( EC 1.13.11.31 ) หรือที่รู้จักกันในชื่อ อาราคิโดเนต 12-ลิโปออกซิ เจเนส , 12-ลิโปออกซิเจเน ส , 12 S -ลิ โปออกซิเจเนส , 12-LOX และ 12 S -LOX เป็น เอนไซม์ ประเภท ลิโปออก...
ALOX12
| ALOX12 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | ALOX12 , 12-LOX, 12S-LOX, LOG12, อะราคิโดเนต 12-ลิโปออกซิเจเนส ชนิด 12S | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | OMIM : 152391 ; MGI : 87998 ; HomoloGene : 560 ; GeneCards : ALOX12 ; OMA : ALOX12 - orthologs | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




ALOX12 ( EC 1.13.11.31 ) หรือที่รู้จักกันในชื่ออาราคิโดเนต 12-ลิโปออกซิ เจเนส , 12-ลิโปออกซิเจเน ส , 12 S -ลิ โปออกซิเจเนส , 12-LOXและ12 S -LOXเป็นเอนไซม์ประเภทลิโปออก ซิเจเนส ซึ่งในมนุษย์ถูกเข้ารหัสโดยยีนALOX12ซึ่งตั้งอยู่ร่วมกับลิโปออกซิเจเนสอื่นๆ บนโครโมโซม 17p13.3 [ 5 ] [ 6 ] ALOX12 เป็นโปรตีนขนาด 75 กิโลดาลตันประกอบด้วยกรดอะมิโน 663 ตัว
การตั้งชื่อ
| อะราคิโดเนต 12-ลิโปออกซิเจเนส | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| ตัวระบุ | |||||||||
| หมายเลข EC | 1.13.11.31 | ||||||||
| หมายเลข CAS | 82391-43-3 | ||||||||
| ฐานข้อมูล | |||||||||
| อินท์เอ็นซ์ | มุมมองของ IntEnz | ||||||||
| เบรนด้า | เบรนด้าเข้าร่วม | ||||||||
| เอ็กซ์แพซี่ | มุมมองของ NiceZyme | ||||||||
| เคกก์ | รายการ KEGG | ||||||||
| เมตาไซค์ | วิถีการเผาผลาญ | ||||||||
| ไพรแอม | ประวัติโดยย่อ | ||||||||
| โครงสร้างPDB | RCSB PDB PDBe PDBsum | ||||||||
| ออนโทโลยีของยีน | อามิโก้ / ควิกโก้ | ||||||||
| |||||||||
ชื่อทางระบบอื่นๆของ ALOX12 ได้แก่ 12S-ลิโปออกซิเจเนส, 12-ลิโปออกซิเจเนสชนิดเกล็ดเลือด, อะราคิโดเนต:ออกซิเจน 12-ออกซิโดรีดักเทส, เดลต้า 12-ลิโปออกซิเจเนส, 12เดลต้า-ลิโปออกซิเจเนส และ C-12 ลิโปออกซิเจเนส ALOX12 ซึ่งมักเรียกว่า 12-ลิโปออกซิเจเนสชนิดเกล็ดเลือดนั้น แตกต่างจาก 12-ลิโปออกซิเจเนสชนิดเม็ดเลือดขาว ซึ่งพบในหนู หนูแรต วัว และหมู แต่ไม่พบในมนุษย์ ลิวโคไซต์-ไทป์ 12-ลิโปออกซิเจเนสในสัตว์เหล่านี้มีลำดับกรดอะมิโนที่เหมือนกัน 73-86% กับALOX15 ของมนุษย์ แต่มีความเหมือนกันเพียง 57-66% กับลิโปออกซิเจเนส-ไทป์ 12-เกล็ดเลือดของมนุษย์ และเช่นเดียวกับ ALOX15 จะเผาผลาญกรดอะราคิโดนิกเป็นหลักไปเป็น 15( S )-ไฮโดรเปอร์ออก ซี -5Z ,8Z , 11Z , 13E- กรดไอโคซาเตตราอีโนอิก (เช่น 15( S )-HpETE; ดู15-ไฮดรอกซีไอโคซาเตตราอีโนอิกแอซิด ) [ 7 ]ดังนั้น ลิวโคไซต์ 12-ลิโปออกซิเจเนสของสัตว์ฟันแทะจึงถือเป็นออร์โธล็อกของALOX15 และถูกกำหนดให้เป็นAlox15 [ 8 ]
ALOX12 และ ALOX15 ของมนุษย์ รวมถึงAlox12และAlox15 ชนิดเม็ดเลือดขาวของสัตว์ฟันแทะ มักถูกเรียกว่า 12/15-ลิโปออกซิเจเนส โดยพิจารณาจากความสามารถในการเผาผลาญกรดอะราคิโดนิกให้เป็นทั้ง 12( S )-HpETE และ 15( S )-HpETE และดำเนินการเผาผลาญแบบเดียวกันนี้กับกรดอะราคิโดนิกที่ถูกเอสเทอริฟายด์กับฟอสโฟลิ ปิดของเยื่อหุ้มเซลล์ ALOX15B ของมนุษย์สร้าง 15( S )-HpETE แต่ไม่สร้าง 12( S )-HpETE ดังนั้นจึงไม่ถือว่าเป็น 12/15-ลิโปออกซิเจเนส[ 9 ] การศึกษาบทบาทของ ALOX12 ในพยาธิสรีรวิทยาโดยใช้แบบจำลองหลักสำหรับการศึกษาการทำงานดังกล่าว เช่น หนูและหนูทดลอง มีความซับซ้อน เนื่องจากทั้งสองชนิดไม่มีลิโปออกซิเจเนส ที่สร้าง 12( S )-HETE เป็นหลักและดังนั้นจึงเทียบเท่ากับการเผาผลาญของ ALOX12 [ 7 ] [ 9 ]ตัวอย่างเช่น หน้าที่ที่อนุมานได้สำหรับAlox12ในหนูที่ขาดAlox12โดยใช้ วิธี การน็อคเอาท์อาจไม่บ่งชี้ถึงหน้าที่ที่คล้ายคลึงกันสำหรับALOX12ในมนุษย์เนื่องจากความแตกต่างในกิจกรรมการเผาผลาญของเอนไซม์ทั้งสองนี้ หน้าที่ของ ALOX12 ยังคลุมเครือมากขึ้นด้วย ALOX15 ของมนุษย์ซึ่งเผาผลาญกรดอะราคิโดนิกเป็นหลักไปเป็น 15( S )-HpETE แต่ยังสร้าง 12( S )-HpETE ในปริมาณที่น้อยกว่าแต่ก็ยังมีความสำคัญอยู่(ดูALOX15 )
ALOX12 ยังแตกต่างจากอะราคิโดเนต 12-ลิโปออกซิเจเนส ชนิด 12R (ALOX12B) ซึ่งเผาผลาญกรดอะราคิโดนิกไปเป็นสเตอริโอไอโซเมอร์R ของ 12( S )-HpETE นั่นคือ 12( R )-ไฮโดรเปอร์ออกซี-5Z , 8Z , 10E , 14Z- ไอโคซาเตตราอีโนอิกแอซิด (12( R )-HpETE) ซึ่งเป็นผลิตภัณฑ์ที่มีบทบาททางพยาธิสรีรวิทยาที่แตกต่างจาก 12( S )-HpETE อย่างมาก (ดูALOX12B )
การค้นพบ
ALOX12 ซึ่งเดิมเรียกว่า อะราคิโดเนต 12-ลิโปออกซิเจเนส ได้รับการระบุลักษณะครั้งแรกโดยผู้ได้รับรางวัลโนเบล เบงต์ ไอ. ซามูเอลสันและเพื่อนร่วมงานที่มีชื่อเสียงของเขา มัตส์ แฮมเบิร์ก ในปี 1974 โดยแสดงให้เห็นว่าเกล็ดเลือด ของมนุษย์ เผาผลาญกรดอะราคิโดนิก ไม่เพียงแต่โดยวิถี ไซโคลออกซิ เจเนส ที่รู้จักกันดีไปเป็นโปรสตาแกลนดินและกรด 12-ไฮดรอกซี เฮปตาเดคาไตรเอโนอิก แต่ยังโดยวิถีที่ไม่ขึ้นกับไซโคลออกซิเจเนสไปเป็นกรด 12( S )-ไฮโดรเปอร์ออกซี-5,8,10,14-ไอโคซาเตตราเอโนอิก กิจกรรมนี้เป็นกิจกรรมลิโปออกซิเจเนสของสัตว์เลี้ยงลูกด้วยนมตัวแรกที่ได้รับการระบุลักษณะ[ 10 ]ในปี 1975 กิจกรรมทางชีวภาพแรกถูกเชื่อมโยงกับเมตาโบไลต์นี้ในการศึกษาที่แสดงให้เห็นว่ามันจำลองการเคลื่อนที่ ของ นิวโทรฟิลของมนุษย์[ 11 ] ในช่วงหลายปีต่อมา ALOX12 ของมนุษย์ได้รับการทำให้บริสุทธิ์ มีลักษณะทางชีวเคมี และมี การโคลนยีนในระดับโมเลกุล[ 7 ] [ 12 ]
การกระจายตัวของเนื้อเยื่อ
โดยอาศัยการมีอยู่ของmRNA เป็นหลัก ALOX12 ของมนุษย์จะกระจายตัวอยู่เป็นส่วนใหญ่ในเกล็ดเลือดและเม็ดเลือดขาวและในระดับที่ต่ำกว่าในชั้นฐานของหนังกำพร้า (โดยเฉพาะในรอยโรคของโรคสะเก็ดเงิน ) เกาะลังเกอร์ฮันส์ภายในตับอ่อน และมะเร็งบางชนิด[ 13 ]
กิจกรรมของเอนไซม์
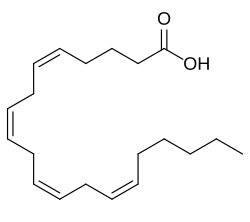
การควบคุมกิจกรรมของ ALOX12 ดูเหมือนจะขึ้นอยู่กับความพร้อมของ สารตั้งต้น กรดไขมันไม่อิ่มตัวเชิงซ้อน (PUFA) ซึ่งถูกปล่อยออกมาจากการจัดเก็บในฟอสโฟลิ ปิดของเยื่อหุ้มเซลล์ โดยการกระตุ้นเซลล์ เป็นหลัก [ 14 ]เอนไซม์มีส่วนร่วมในการเผาผลาญกรดอะราคิโดนิก โดยดำเนิน การปฏิกิริยาเคมีต่อไปนี้โดยที่ สาร ตั้งต้น ของมัน คือกรดอะราคิโดนิก (เรียกอีกอย่างว่าอะราคิโดเนต หรือทางเคมีคือ 5 Z ,8 Z ,11 Z ,14 Z -eicosatetraenoic acid) และ O 2 (เช่นออกซิเจน ) และผลิตภัณฑ์ ของมัน คือ 12 S -hydroperoxy- 5 Z ,8 Z ,10 E ,14 Z -eicosatetraenoic acid (เช่น 12 S -hydroperoxyeicosatetraenoic acid หรือ 12 S -HpETE): [ 10 ] [ 15 ]
- อะราคิโดเนต + O 2 → 12 S -ไฮโดรเปอร์ออกซี-5 Z ,8 Z ,10 E ,14 Z -กรดไอโคซาเตตราอีโนอิก
ในเซลล์ 12 S HpETE อาจถูกเมตาบอไลซ์ต่อไปโดย ALOX12 เอง โดยALOXE3หรืออาจเป็นการสังเคราะห์เฮปอกซิลินอื่นๆ ที่ยังไม่ได้รับการระบุอย่างสมบูรณ์ เพื่อสร้างเฮปอกซิลิน A3 (8 R/S -hydroxy-11,12-oxido-5 Z ,9 E ,14 Z -eicosatrienoic acid) และ B3 (10 R/S -hydroxy-11,12-oxido-5 Z ,8 Z ,14 Z -eicosatrienoic acid): [ 16 ] [ 17 ] [ 18 ]
- 12 S -ไฮโดรเปอร์ออกซี-5Z,8Z,10E,14Z-กรดไอโคซาเตตราอีโนอิก → 8 R/S -ไฮดรอกซี-11,12-ออกซิโด-5 Z ,9 E ,14 Z -กรดไอโคซาไตรอีโนอิก + 10 R/S -ไฮดรอกซี-11,12-ออกซิโด-5 Z ,8 Z ,14 Z - กรดไอโคซา ไตรอีโนอิก
เฮปอกซิลินสามารถส่งเสริม การตอบสนอง การอักเสบ บางอย่าง เพิ่มการรับรู้ความเจ็บปวด (เช่นภาวะไว ต่อการสัมผัส ) ควบคุมการไหลเวียนของเลือดในบริเวณต่างๆ และมีส่วนช่วยในการควบคุมความดันโลหิตในแบบจำลองสัตว์ (ดูเฮปอกซิลิน ) อย่างไรก็ตาม โดยทั่วไปแล้ว 12 S -HpETE จะถูกลดลงอย่างรวดเร็วเป็น ผลิตภัณฑ์ ไฮดรอกซิลโดยกิจกรรมของเอนไซม์เปอร์ออกซิเดสในเซลล์ทั่วไป ทำให้เกิด 12 S -ไฮดรอกซี-5 Z ,8 Z ,10 E ,14 Z -กรดไอโคซาเตตราอีโนอิก หรือ12-ไฮดรอกซีไอโคซาเตตราอีโนอิกหรือ 12 S -HETE: [ 19 ]
- 12 S -ไฮโดรเปอร์ออกซี-5( Z ),8( Z ),10( E ),14( Z )-กรดไอโคซาเตตราอีโนอิก → 12 S -ไฮดรอกซี-5( Z ),8( Z ),10( E ),14( Z )-กรดไอโคซาเตตราอีโนอิก
12S - HETE กระตุ้นการตอบสนองต่อการอักเสบ อาจมีส่วนเกี่ยวข้องกับการรับรู้ความรู้สึกคันที่ผิวหนัง และควบคุมการไหลเวียนของเลือดในบริเวณต่างๆ ในแบบจำลองสัตว์ นอกจากนี้ยังส่งเสริมพฤติกรรมร้ายแรงของเซลล์มะเร็งมนุษย์ที่เพาะเลี้ยง ตลอดจนการเจริญเติบโตของมะเร็งบางชนิดในแบบจำลองสัตว์ (ดู12-HETE ) ในขณะที่อะราคิโดเนตและ 12( S )-HETE เป็นสารตั้งต้นและผลิตภัณฑ์หลักของ ALOX12 ตามลำดับ เอนไซม์นี้ยังเผาผลาญ PUFA อื่นๆ ด้วย
มันเผาผลาญกรดไขมันโอเมก้า-3โดโคซาเฮกซาเอโนอิกแอซิด (DHA เช่น 4( Z ),7( Z ),10( Z ) ,13( Z ),16( Z ),19( Z ) -โดโคซาเฮกซาเอโนอิกแอซิด ไปเป็น 14( R )-ไฮโดรเปอร์ออกซี-4( Z ),8( Z ),10( Z ),12( E ),16( Z ),19( Z )-โดโคซาเฮกซาเอโนอิกแอซิด) (เช่น 17-ไฮโดรเปอร์ออกซี-DHA)
จากนั้น ALOX12 หรือเอนไซม์ประเภทอีพอกซิเดสที่ไม่ทราบชนิด อาจเผาผลาญสารตัวกลางนี้ให้กลายเป็นอีพอกไซด์ 13,14-epoxy-4( Z ),7 ( Z ),9( E ),11( E ) ,16( Z ),19( Z )-docosahexaenoic acid (เช่น 13,14-e-maresin)
สารนี้จะถูกเมตาบอไลซ์ต่อไปเป็น 7 R ,14 S -dihydroxy-4 Z ,8 E ,10 E ,12 Z ,16 Z ,19 Z -docosahexaenoic acid (เช่นMaresin 1) โดย เอนไซม์ประเภท อีพอกไซด์ไฮโดรเลส ที่ไม่ทราบ ชนิด:
- ดีเอชเอ → 17-ไฮโดรเพอรอกซี-ดีเอชเอ → 13,14-อี-มาเรซิน → มาเรซิน-1
Maresin 1 มีฤทธิ์หลายอย่างที่อาจขัดแย้งกับฤทธิ์ของ 12( S )-HETE และเฮปอกซิลินส์ โดยเป็นสมาชิกของกลุ่มเมตาบอไลต์ PUFA ที่เรียกว่าSpecialized pro-resolution mediators (SPMs) ซึ่งมีฤทธิ์ต้านการอักเสบ บรรเทาอาการปวด และฤทธิ์ป้องกันอื่นๆ[ 20 ] ALOX12 ยังออกฤทธิ์ต่อลิวโคไตรอีน A4 (LTA4) ในปฏิกิริยาสองเซลล์ที่เรียกว่าการเผาผลาญข้ามเซลล์: นิวโทรฟิลของมนุษย์เผาผลาญกรดอะราคิโดนิกเป็น 5,6-อีพอกไซด์ LTA4 และปล่อยสารตัวกลางนี้ไปยังนิวโทรฟิลที่อยู่ใกล้เคียงซึ่งเผาผลาญเป็นลิพอกซิน A4 (5 S ,6 R ,15 S -ไตร ไฮดรอกซี-7 E ,9 E ,11 Z ,13 Z -กรดไอโคซาเตตราอีโนอิก) และลิพอกซิน B4 (5 S ,14 R ,15 S -ไตรไฮดรอกซี-6 E ,8 Z ,10 E ,12 E -กรดไอโคซาเตตราอีโนอิก); ลิพอกซินทั้งสองเป็น SPM ที่มีกิจกรรมคล้าย SPM หลายอย่าง (ดูลิพอกซิน ) [ 21 ] ALOX12 อาจเผาผลาญ DHA ในปริมาณที่น้อยกว่าให้เป็นผลิตภัณฑ์รอง ได้แก่ 17-ไฮโดรเปอร์ออกซี-DHA, 11-ไฮโดรเปอร์ออกซี-DHA และ 8,14-ไดไฮดรอกซี-DHA [ 20 ] ALOX12 อาจเผาผลาญ 5( S )-HETE ให้เป็น 5 S ,12 S -ไดไฮดรอกซีไอโคซาเตตราอีโนอิกแอซิด (12,15-ไดHETE) และ 15 S -HETE ให้เป็น 14,15 S - ไดETE [ 14 ] แม้ว่าสารประกอบเหล่านี้จะยังไม่ได้รับการประเมินอย่างละเอียดถี่ถ้วนเกี่ยวกับฤทธิ์ทางชีวภาพ แต่ 17-ไฮโดรเปอร์ออกซี-HDHA และผลิตภัณฑ์ที่ลดลงซึ่งถูกแปลงอย่างรวดเร็วในเซลล์ คือ 17-ไฮดรอกซี-HDHA ได้แสดงให้เห็นว่าสามารถยับยั้งการเจริญเติบโตของเซลล์มะเร็งต่อมลูกหมากของมนุษย์ที่เพาะเลี้ยงโดยทำให้เซลล์เหล่านั้นเข้าสู่ภาวะอะพอพโทซิส[ 22 ]
การศึกษาในสัตว์
การศึกษาในสัตว์ฟันแทะที่ขาดหรือเกิดภาวะขาดเอนไซม์ลิโปออกซิเจเนสชนิดเม็ดเลือดขาว 12, Alox12 (ซึ่งมีความสัมพันธ์ใกล้ชิดกับ ALOX15 ในมนุษย์มากที่สุด) บ่งชี้ว่าเอนไซม์นี้มีส่วนเกี่ยวข้องใน: ก) การป้องกันการเกิดและภาวะแทรกซ้อนของโรคเบาหวานที่เกิดจากอาหารและ/หรือพันธุกรรม ความผิดปกติของเซลล์/เนื้อเยื่อไขมันและ โรค อ้วนข)การเกิด ภาวะ หลอดเลือด แดงแข็งและตับอักเสบจากไขมัน ค ) การควบคุมการหดตัว การขยายตัว ความดัน การปรับโครงสร้าง และการสร้างหลอดเลือด ใหม่ ค)การรักษาการทำงานของไต ระบบประสาท และสมองให้เป็นปกติ และง)การเกิดโรคอัลไซเมอร์ [ 8 ] [ 9 ] [ 23 ] ในการศึกษาเหล่านี้ มักไม่ชัดเจนว่าเมตาโบไลต์ใดของAlox12มีส่วนเกี่ยวข้อง
การศึกษาทางคลินิกก่อนการทดลอง
กลุ่มอาการเมตาบอลิก
กลุ่มอาการเมตาบอลิกคือการรวมตัวกันของภาวะทางการแพทย์อย่างน้อยสามในห้าอย่างต่อไปนี้: โรคอ้วนลงพุง (โรคอ้วนส่วนกลาง) ความดันโลหิตสูงระดับน้ำตาลในพลาสมาขณะอดอาหาร สูง (หรือโรคเบาหวาน อย่างชัดเจน ) ไตรกลีเซอไรด์ในซีรั่มสูงและระดับไลโปโปรตีนความหนาแน่นสูง (HDL) ต่ำ ALOX12 และเมตาโบไลต์ 12( S )-HETE มีระดับสูงขึ้นในเกาะลังเกอร์ฮันส์ของผู้ป่วยโรคเบาหวานประเภทที่ 1หรือประเภทที่ 2เช่นเดียวกับในเซลล์ไขมันของเนื้อเยื่อไขมันสีขาวของผู้ป่วยโรคเบาหวานประเภทที่ 2 ที่เป็นโรคอ้วนอย่างรุนแรง[ 8 ]เซลล์PP (เช่น เซลล์แกมมา) ของเกาะตับอ่อนดูเหมือนจะเป็นบริเวณหลัก หากไม่ใช่บริเวณเดียวที่ ALOX12 แสดงออกในผู้ป่วยเหล่านี้[ 8 ]การศึกษาเสนอว่าในเกาะลังเกอร์ฮันส์ ALOX12 และผลิตภัณฑ์ 12( S )-HETE ของมันทำให้เกิดการผลิตอนุมูลอิสระและการอักเสบมากเกินไป ซึ่งนำไปสู่การสูญเสียเซลล์เบต้าที่หลั่งอินซูลินและทำให้เกิดโรคเบาหวานประเภท 1 และ 2 และในเนื้อเยื่อไขมัน การมี ALOX12, 12( S )-HETE, อนุมูลอิสระ และการอักเสบมากเกินไปจะนำไปสู่ความผิดปกติของเซลล์ไขมัน (ดูเพิ่มเติมที่ 12-HETE#การอักเสบและโรคอักเสบและ12-HETE#เบาหวาน ) อันที่จริง ในการศึกษาหนึ่ง โพลีมอร์ฟิซึมแบบนิวคลีโอไทด์เดี่ยว rs2073438 [ 24 ]ซึ่งตั้งอยู่ใน บริเวณ อินตรอนของ ยีน ALOX12มีความสัมพันธ์อย่างมีนัยสำคัญกับมวลไขมันทั้งหมดและร้อยละของไขมันในผู้ชายชาวจีนหนุ่มที่เป็นโรคอ้วนเมื่อเทียบกับผู้ชายชาวจีนหนุ่มที่ไม่เป็นโรคอ้วน[ 8 ] [ 13 ] [ 18 ] ALOX12 และ 12( S )-HETE ก็มีส่วนเกี่ยวข้องกับความดันโลหิตสูงเช่นกัน (ดูหัวข้อถัดไป) ดังนั้น ALOX12 และเมตาโบไลต์ของมันอาจมีส่วนทำให้เกิดและ/หรือทำให้โรคอ้วน เบาหวาน ความดันโลหิตสูง และ/หรือกลุ่มอาการเมตาบอลิกรุนแรงขึ้น
หลอดเลือด
สารยับยั้ง ALOX12 ที่เลือกแต่ไม่เฉพาะเจาะจงทั้งหมดช่วยลดการตอบสนองการเจริญเติบโตของเซลล์เยื่อบุผนังหลอดเลือด มนุษย์ที่เพาะเลี้ยง ต่อปัจจัยการเจริญเติบโตของไฟโบรบลาสต์พื้นฐานและปัจจัยการเจริญเติบโตของเยื่อบุผนังหลอดเลือด (VEGF) การลดลงนี้ถูกย้อนกลับบางส่วนโดย 12( S )-HETE; 12( S )-HETE ยังกระตุ้นเซลล์ต่อมลูกหมากของมนุษย์ให้ผลิต VEGF [ 19 ]ผลลัพธ์เหล่านี้ชี้ให้เห็นว่าการตอบสนองการเจริญเติบโตต่อปัจจัยการเจริญเติบโตทั้งสองเกี่ยวข้องกับการกระตุ้นการผลิต 12( S )-HETE โดยเซลล์เยื่อบุผนังหลอดเลือด ดังนั้น ALOX12 อาจเป็นเป้าหมายในการลดการสร้างหลอดเลือดใหม่ที่ส่งเสริมโรคข้ออักเสบและมะเร็ง 12( S )-HETE ยังขยายหลอดเลือดฝอยในหัวใจของมนุษย์โดยการกระตุ้นช่องโพแทสเซียม BKca ของกล้ามเนื้อเรียบของหลอดเลือดเหล่านี้ และจึงถูกเสนอให้เป็น ปัจจัยไฮเปอร์โพลาไรซ์ ที่ได้จากเยื่อบุผนังหลอดเลือด[ 9 ] [ 19 ]สุดท้ายนี้การกลายพันธุ์ของนิวคลีโอไทด์เดี่ยวใน ยีน ALOX12 (R261Q [3957 G>A]) เกี่ยวข้องกับความดันโลหิตสูงชนิดไม่ทราบสาเหตุและการเพิ่มขึ้นของการขับถ่าย 12( S )-HETE ในปัสสาวะของมนุษย์ และอาจเป็นปัจจัยที่ทำให้เกิดความดันโลหิตสูงชนิดไม่ทราบสาเหตุ (ดูเพิ่มเติมที่12-HETE#ความดันโลหิต ) [ 9 ] [ 25 ]
โรคอัลไซเมอร์
ผู้ป่วยโรคอัลไซเมอร์หรือภาวะสมองเสื่อมรูปแบบอื่นๆ มีระดับ 12( S )-HETE (และ 15( S )-HETE) ในน้ำไขสันหลังสูงกว่าอย่างมีนัยสำคัญเมื่อเทียบกับบุคคลปกติที่มีอายุเท่ากัน การศึกษาเพิ่มเติมในแบบจำลองสัตว์ฟันแทะที่มียีนกลายพันธุ์ของมนุษย์สำหรับโปรตีนสารตั้งต้นอะไมลอยด์และ/หรือโปรตีนเทา (ดูโปรตีนเทา#ความสำคัญทางคลินิก ) ที่ทำให้เกิดกลุ่มอาการคล้ายภาวะสมองเสื่อมแบบอัลไซเมอร์บ่งชี้ว่า 12( S )-HETE, 15( S )-HETE และเอนไซม์ชนิด 12/15-ลิโปออกซิเจเนสมีส่วนเกี่ยวข้องกับการพัฒนาและความก้าวหน้าของอาการและผลการค้นพบที่คล้ายโรคอัลไซเมอร์ในสัตว์เหล่านี้[ 23 ]ในการศึกษาหนึ่ง พบว่า mRNA ของ ALOX12 สูงขึ้นในเนื้อเยื่อสมองของผู้ป่วยโรคอัลไซเมอร์เมื่อเทียบกับผู้ป่วยกลุ่มควบคุม[ 13 ]ผลลัพธ์เหล่านี้ชี้ให้เห็นว่า ALOX12 (หรือ ALOX15) อาจมีส่วนทำให้เกิดโรคอัลไซเมอร์ในมนุษย์
มะเร็ง
การศึกษาเกี่ยวกับมะเร็งต่อมลูกหมากพบว่าเซลล์มะเร็งต่อมลูกหมากของมนุษย์ที่เพาะเลี้ยงในห้องปฏิบัติการมีการแสดงออกของ ALOX12 มากเกินไป ผลิต 12( S )-HETE มากเกินไป และตอบสนองต่อ 12( S )-HETE โดยการเพิ่มอัตราการแพร่พันธุ์ เพิ่มการแสดงออกของอินทิกริน บนพื้นผิวเซลล์ เพิ่มการอยู่รอดและชะลอการตาย ของเซลล์ และเพิ่มการผลิตปัจจัยการเจริญเติบโตของหลอดเลือดและMMP9 (เช่น เมทริกซ์เมทัลโลเปปติเดส 9) สารยับยั้ง ALOX12 ที่เลือก (แต่ไม่ทั้งหมด) สามารถลดการแพร่กระจายและการอยู่รอดของเซลล์เหล่านี้ได้ (ดูเพิ่มเติมที่12-HETE#มะเร็งต่อมลูกหมาก ) การค้นพบเหล่านี้ชี้ให้เห็นว่า ALOX12 และผลิตภัณฑ์ 12( S )-HETE อาจมีส่วนช่วยในการเจริญเติบโตและการแพร่กระจายของมะเร็งต่อมลูกหมากในมนุษย์[ 19 ]เมื่อเร็วๆ นี้ การเกิดเมทิลเลชั่นมากเกินไปของ ยีน ALOX12ในเนื้อเยื่อมะเร็งต่อมลูกหมากมีความสัมพันธ์กับตัวทำนายทางคลินิกสำหรับอัตราการเกิดโรคซ้ำสูง[ 26 ]การศึกษาบางชิ้นพบว่า 12( S )-HETE ยังส่งเสริมการเจริญเติบโตและ/หรือพฤติกรรมที่เกี่ยวข้องกับมะเร็งร้ายของเซลล์มะเร็งชนิดอื่นๆ ที่เพาะเลี้ยง (ดู12-HETE#มะเร็งชนิดอื่นๆ ) [ 19 ] ALOX12 ได้รับการแสดงให้เห็นว่ามีปฏิสัมพันธ์กับเคราติน 5และLMNAตามที่คัดกรองในไลบรารีปฏิสัมพันธ์แบบยีสต์ทูไฮบริดจากเซลล์มะเร็งผิวหนังชนิด A431 ของมนุษย์ โปรตีนเหล่านี้เป็นตัวเลือกสำหรับการควบคุม 12-LOX โดยเฉพาะในเซลล์เนื้องอก[ 27 ]
หน้าที่ของเกล็ดเลือด
แม้ว่าจะถูกระบุครั้งแรกในเกล็ดเลือดของมนุษย์ บทบาทของ ALOX12 และเมตาโบไลต์หลัก 12( S )-HpETE และ 12( S )-HETE ในการทำงานของเกล็ดเลือดยังคงเป็นที่ถกเถียงและไม่ชัดเจน เป็นไปได้ว่าเส้นทางการเผาผลาญ ALOX12-12( S )-HETE มีหน้าที่สองอย่างในการส่งเสริมหรือยับยั้งการตอบสนองของเกล็ดเลือดขึ้นอยู่กับตัวกระตุ้นและการตอบสนองที่ศึกษา แต่การยับยั้ง ALOX12 อาจมีประโยชน์ในการยับยั้งการแข็งตัวของเลือดที่เกี่ยวข้องกับเกล็ดเลือดในที่สุด[ 19 ]
สมาคมอื่นๆ
ยีนALOX12มีอัลลีลที่ทำให้เกิดความเสี่ยง (rs6502997, [ 28 ] rs312462, [ 29 ] rs6502998, [ 30 ]และ rs434473 [ 31 ] ) สำหรับโรคปรสิต โรคท็อกโซพลาสโมซิสแต่กำเนิดในมนุษย์[ 13 ] [ 32 ] ทารก ในครรภ์ที่มียีนอัลลีลเหล่านี้จึงมีความเสี่ยงต่อโรคนี้เพิ่มขึ้น
ลิงก์ภายนอก
- ตำแหน่งจีโนมของยีน ALOX12ในมนุษย์และรายละเอียดของยีนALOX12 ใน UCSC Genome Browser
อ่านเพิ่มเติม
- Yoshimoto T, Arakawa T, Hada T, Yamamoto S, Takahashi E (ธันวาคม 1992). "โครงสร้างและตำแหน่งโครโมโซมของยีนอะราคิโดเนต 12-ลิโปออกซิเจเนสของมนุษย์"วารสารเคมีชีวภาพ 267 ( 34): 24805– 9. doi : 10.1016/S0021-9258(18)35835-6 . PMID 1447217 .
- Izumi T, Hoshiko S, Rådmark O, Samuelsson B (ตุลาคม 1990). "การโคลน cDNA สำหรับ 12-ลิโปออกซิเจเนสของมนุษย์" . Proceedings of the National Academy of Sciences of the United States of America . 87 (19): 7477– 81. Bibcode : 1990PNAS...87.7477I . doi : 10.1073/pnas.87.19.7477 . PMC 54770 . PMID 2217179 .
- Funk CD, Furci L, FitzGerald GA (สิงหาคม 1990). "การโคลนนิ่งระดับโมเลกุล โครงสร้างหลัก และการแสดงออกของ 12-ลิโปออกซิเจเนสของเกล็ดเลือดมนุษย์/เซลล์เม็ดเลือดขาวชนิดอีริโทรลูคีเมีย" Proceedings of the National Academy of Sciences of the United States of America . 87 (15): 5638– 42. Bibcode : 1990PNAS...87.5638F . doi : 10.1073/pnas.87.15.5638 . PMC 54382 . PMID 2377602 .
- Flatman S, Morgan A, McDonald-Gibson RG, Davey A, Jonas GE, Slater TF (พฤษภาคม 1988). "กิจกรรม 12-ลิโปออกซิเจเนสในปากมดลูกของมนุษย์" Prostaglandins, Leukotrienes, and Essential Fatty Acids . 32 (2): 87– 94. doi : 10.1016/0952-3278(88)90101-9 . PMID 3406043 .
- Wong PY, Westlund P, Hamberg M, Granström E, Chao PH, Samuelsson B (สิงหาคม 1985). " 15-ลิโปออกซิเจเนสในเกล็ดเลือดมนุษย์"วารสารเคมีชีวภาพ260 (16): 9162– 5. doi : 10.1016/S0021-9258(17)39346-8 . PMID 3926763 .
- Nakamura M, Ueda N, Kishimoto K, Yoshimoto T, Yamamoto S, Ishimura K (มีนาคม 1995). "การระบุตำแหน่งทางอิมมูโนไซโตเคมีของอะราคิโดเนต 12-ลิโปออกซิเจเนสชนิดเกล็ดเลือดในเซลล์เม็ดเลือดของหนู"วารสารฮิสโตเคมีและไซโตเคมี 43 ( 3): 237– 44. doi : 10.1177/43.3.7868854 . PMID 7868854 .
- Hussain H, Shornick LP, Shannon VR, Wilson JD, Funk CD, Pentland AP, Holtzman MJ (มกราคม 1994). "หนังกำพร้ามีลิโปออกซิเจเนสชนิดเกล็ดเลือด 12 ที่มีการแสดงออกมากเกินไปในเซลล์เคราติโนไซต์ของชั้นเจอร์มินัลในโรคสะเก็ดเงิน" วารสารสรีรวิทยาอเมริกัน 266 ( 1 Pt 1): C243-53. doi : 10.1152/ajpcell.1994.266.1.C243 . PMID 8304420 .
- Arora JK, Lysz TW, Zelenka PS (มิถุนายน 1996). "บทบาทของ 12(S)-HETE ในการตอบสนองของเซลล์เยื่อบุผิวเลนส์ตาของมนุษย์ต่อปัจจัยการเจริญเติบโตของผิวหนังและอินซูลิน" Investigative Ophthalmology & Visual Science . 37 (7): 1411– 8. PMID 8641843 .
- Hagmann W, Gao X, Timar J, Chen YQ, Strohmaier AR, Fahrenkopf C, Kagawa D, Lee M, Zacharek A, Honn KV (พฤศจิกายน 1996). "12-ลิโปออกซิเจเนสในเซลล์ A431: เอกลักษณ์ทางพันธุกรรม การปรับเปลี่ยนการแสดงออก และตำแหน่งภายในเซลล์" Experimental Cell Research . 228 (2): 197– 205. doi : 10.1006/excr.1996.0317 . PMID 8912711 .
- Nakamura M, Yamamoto S, Ishimura K (พฤษภาคม 1997). "การระบุตำแหน่งย่อยเซลล์ของอะราคิโดเนต 12-ลิโปออกซิเจเนสและผลกระทบทางสัณฐานวิทยาของการแสดงออกมากเกินไปในเคราติโนไซต์ของหนู" Cell and Tissue Research . 288 (2): 327– 34. doi : 10.1007/s004410050818 . PMID 9082968 . S2CID 23548308 .
- Nigam S, Kumar GS, Sutherland M, Schewe T, Ikawa H, Yamasaki Y, Ueda N, Yamamoto S (กันยายน 1999). "การยับยั้งการเผาผลาญของไลโปออกซิเจเนสชนิด 12 เกล็ดเลือดในปากมดลูกของมนุษย์ที่มีมะเร็งลุกลาม" International Journal of Cancer . 82 (6): 827– 31. doi : 10.1002/(SICI)1097-0215(19990909)82:6<827::AID-IJC10>3.0.CO;2-Q . PMID 10446449 . S2CID 22417409 .
- Tornhamre S, Elmqvist A, Lindgren JA (เมษายน 2543). "15-ลิโปออกซิเจเนชันของลิวโคไตรอีน A 4 : การศึกษาประสิทธิภาพของ 12- และ 15-ลิโปออกซิเจเนสในการเร่งปฏิกิริยาการก่อตัวของลิโปซิ น" Biochimica et Biophysica Acta (BBA) - ชีววิทยาโมเลกุลและเซลล์ของไขมัน1484 ( 2– 3): 298– 306. doi : 10.1016/S1388-1981(00)00017-2 . PMID 10760478 .
- Chen BK, Tsai TY, Huang HS, Chen LC, Chang WC, Tsai SB, Chang WC (2003). "บทบาทเชิงหน้าที่ของการกระตุ้น extracellular signal-regulated kinase และการเหนี่ยวนำ c-Jun ในการกระตุ้นโปรโมเตอร์ของยีน 12(S)-lipoxygenase ของมนุษย์ที่เหนี่ยวนำโดยฟอร์บอลเอสเทอร์" วารสารวิทยาศาสตร์ชีวการแพทย์9 (2): 156– 65. doi : 10.1159/000048212 . PMID 11914583 . S2CID 46753449 .
- Winer I, Normolle DP, Shureiqi I, Sondak VK, Johnson T, Su L, Brenner DE (ตุลาคม 2545). "การแสดงออกของ 12-ลิโปออกซิเจเนสเป็นตัวบ่งชี้ทางชีวภาพสำหรับการเกิดมะเร็งผิวหนัง" Melanoma Research . 12 (5): 429– 34. doi : 10.1097/00008390-200209000-00003 . PMID 12394183 . S2CID 27336312 .
- Gu J, Wen Y, Mison A, Nadler JL (กุมภาพันธ์ 2546). "วิถี 12-ลิโปออกซิเจเนสเพิ่มการผลิตอัลโดสเตอโรน การฟอสโฟรีเลชันของโปรตีนที่จับกับองค์ประกอบการตอบสนองของ 3',5'-ไซคลิกอะดีโนซีนโมโนฟอสเฟต และการกระตุ้น p38 มิทโทเจนแอคติเวเตดโปรตีนไคเนสในเซลล์ต่อมหมวกไตของมนุษย์ H295R" Endocrinology . 144 ( 2 ): 534– 43. doi : 10.1210/en.2002-220580 . PMID 12538614 .
- ฟริดแมน ซี, โอโจปิ อีพี, เกรโกริโอ เอสพี, อิเคนากะ เอเอช, โมเรโน ดีเอช, เดเมทริโอ เอฟเอ็น, กิมาไรส์ พีอี, วัลลาดา เอชพี, กัตตาซ WF, ดิอาส เนโต อี (กุมภาพันธ์ 2546) "การเชื่อมโยงของความหลากหลายใหม่ในยีน ALOX12 กับโรคไบโพลาร์" หอจดหมายเหตุแห่งจิตเวชและประสาทวิทยาศาสตร์แห่งยุโรป253 (1): 40– 3. ดอย : 10.1007/s00406-003-0404- y PMID12664313 . S2CID 21064663 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ALOX12
ALOX12 ( EC 1.13.11.31 ) หรือที่รู้จักกันในชื่อ อาราคิโดเนต 12-ลิโปออกซิ เจเนส , 12-ลิโปออกซิเจเน ส , 12 S -ลิ โปออกซิเจเนส , 12-LOX และ 12 S -LOX เป็น เอนไซม์ ประเภท ลิโปออก...
การตั้งชื่อ
ชื่อทางระบบ อื่นๆของ ALOX12 ได้แก่ 12S-ลิโปออกซิเจเนส, 12-ลิโปออกซิเจเนสชนิดเกล็ดเลือด, อะราคิโดเนต:ออกซิเจน 12-ออกซิโดรีดักเทส, เดลต้า 12-ลิโปออกซิเจเนส, 12เดลต้า-ลิโปออกซิเจเนส และ C-12 ลิโปออกซิเจเนส ALOX12 ซึ่งมักเรียกว่า...
การค้นพบ
ALOX12 ซึ่งเดิมเรียกว่า อะราคิโดเนต 12-ลิโปออกซิเจเนส ได้รับการระบุลักษณะครั้งแรกโดยผู้ได้รับรางวัลโน เบล เบงต์ ไอ.
การกระจายตัวของเนื้อเยื่อ
โดยอาศัยการมีอยู่ของ mRNA เป็นหลัก ALOX12 ของมนุษย์จะกระจายตัวอยู่เป็นส่วนใหญ่ในเกล็ดเลือดและ เม็ดเลือดขาว และในระดับที่ต่ำกว่าในชั้นฐานของหนังกำพร้า (โดยเฉพาะในรอยโรคของ โรคสะเก็ดเงิน ) เกาะลังเกอร์ฮันส์ ภายในตับอ่อน และมะเร็งบางชนิด [ 13 ]