อ่าน 18 นาที
ปมประสาทฐาน
ฐานสมอง ( BG ) หรือนิวเคลียสฐานเป็นกลุ่มของนิวเคลียสใต้เปลือกสมอง (กลุ่มของเซลล์ประสาท) ที่พบในสมองของสัตว์มีกระดูกสันหลัง
ปมประสาทฐาน
| ปมประสาทฐาน | |
|---|---|
 แผนภาพแสดงโครงสร้างของปุ่มสมองส่วนฐานซ้อนทับอยู่บนเปลือกสมอง | |
 ปมประสาทฐานสมอง มองจากด้านหน้า | |
| รายละเอียด | |
| ส่วนหนึ่งของ | มันสมอง |
| ตัวระบุ | |
| ละติน | นิวเคลียสฐาน |
| คำย่อ | บีจี |
| เมช | D001479 |
| นิวโรเนมส์ | 224 , 2677 |
| รหัสNeuroLex | เบิร์นเล็กซ์_826 |
| TA98 | A14.1.09.501 |
| ทีเอ2 | 5559 |
| เอฟเอ็มเอ | 84013 |
| คำศัพท์ทางกายวิภาคศาสตร์ของระบบประสาท | |
ฐานสมอง ( BG ) หรือนิวเคลียสฐานเป็นกลุ่มของนิวเคลียสใต้เปลือกสมอง (กลุ่มของเซลล์ประสาท) ที่พบในสมองของสัตว์มีกระดูกสันหลัง ตั้งอยู่บริเวณฐานของสมองส่วนหน้าและส่วนบนของสมองส่วนกลางมีการเชื่อมต่อที่แน่นหนากับเปลือกสมองทาลามัส ก้านสมองและบริเวณสมองอื่นๆ ฐานสมองเกี่ยวข้องกับหน้าที่ต่างๆ มากมาย รวมถึงการควบคุมการเคลื่อนไหวของกล้ามเนื้อ โดยสมัครใจ การ เรียน รู้เชิงกระบวนการการสร้างนิสัยการเรียนรู้แบบมีเงื่อนไข[ 1 ]การ เคลื่อนไหว ของดวงตาการรับรู้[ 2 ]และอารมณ์ [ 3 ]
ส่วนประกอบการทำงานหลักของฐานสมองประกอบด้วยสไตรอาตัมซึ่งประกอบด้วยสไตรอาตัมส่วนหลัง ( นิวเคลียสคอเดตและพิวทาเมน ) และสไตรอาตัมส่วนหน้า ( นิวเคลียสแอคคัมเบนส์และปุ่มรับกลิ่น ) โกลบัสพัลลิ ดัส เวนทรัลพัล ลิดัม ซับส แตนเชียไนกราและนิวเคลียสซับทาลามิก [ 4 ] แต่ละส่วนประกอบเหล่านี้มีโครงสร้างทางกายวิภาคและทางเคมีประสาทภายในที่ซับซ้อน ส่วนประกอบที่ใหญ่ที่สุดคือ สไตรอาตัม (ส่วนหลังและส่วนหน้า) รับข้อมูลจากบริเวณต่างๆ ของสมอง แต่ส่งเฉพาะข้อมูลออกไปยังส่วนประกอบอื่นๆ ของฐานสมองเท่านั้น โกลบัสพัลลิดัสรับข้อมูลจากสไตรอาตัมและส่งเอาต์พุตยับยั้งไปยังบริเวณที่เกี่ยวข้องกับการเคลื่อนไหวหลายแห่ง ซับสแตนเชียไนกราเป็นแหล่งที่มาของข้อมูลเข้าของสไตรอาตัมของสารสื่อประสาทโดปามีนซึ่งมีบทบาทสำคัญในการทำงานของฐานสมอง นิวเคลียสซับทาลามิกส่วนใหญ่รับข้อมูลจากสไตรอาตัมและเปลือกสมอง และส่งต่อไปยังโกลบัสพัลลิดัส
เชื่อกันว่าฐานสมองมีบทบาทสำคัญในการเลือกการกระทำช่วยในการเลือกพฤติกรรมที่จะดำเนินการ โดยเฉพาะอย่างยิ่ง ฐานสมองจะควบคุมบริเวณคอร์เทกซ์มอเตอร์และพรีมอเตอร์ ทำให้การเคลื่อนไหวโดยสมัครใจเป็นไปอย่างราบรื่น[ 2 ] [ 5 ] การศึกษาเชิงทดลองแสดงให้เห็นว่าฐานสมองมีอิทธิพลยับยั้ง ระบบมอเตอร์หลายระบบและการปลดปล่อยการยับยั้งนี้ทำให้ระบบมอเตอร์สามารถทำงานได้ การ "สลับพฤติกรรม" ที่เกิดขึ้นภายในฐานสมองได้รับอิทธิพลจากสัญญาณจากหลายส่วนของสมอง รวมถึงคอร์เทกซ์ส่วนหน้าซึ่งมีบทบาทสำคัญในหน้าที่การบริหารจัดการ [ 3 ] [ 6 ] นอกจากนี้ยังมีการตั้งสมมติฐานว่าฐานสมองไม่เพียงแต่รับผิดชอบในการเลือกการกระทำทางมอเตอร์เท่านั้น แต่ยังรับผิดชอบในการเลือกการกระทำทางปัญญาอีกด้วย[ 7 ] [ 8 ] [ 9 ]แบบจำลองทางคอมพิวเตอร์ของการเลือกการกระทำในฐานสมองได้รวมสิ่งนี้ไว้ด้วย[ 10 ]
ปมประสาทฐานมีความสำคัญอย่างยิ่งต่อการทำงานของสมองและพฤติกรรมปกติ การทำงานผิดปกติของปมประสาทฐาน ส่งผลให้เกิด ภาวะทางระบบประสาท ที่หลากหลาย รวมถึงความผิดปกติของการควบคุมพฤติกรรมและการเคลื่อนไหว ตลอดจนความบกพร่องทางสติปัญญาที่คล้ายกับที่เกิดจากความเสียหายต่อ เปลือกสมอง ส่วน หน้า [ 11 ] ความผิดปกติ ทางพฤติกรรม ได้แก่ กลุ่มอาการทูเร็ตต์โรค ย้ำ คิดย้ำทำและการเสพติด ความ ผิดปกติ ของการเคลื่อนไหวได้แก่โรคพาร์กินสันซึ่งเกี่ยวข้องกับการเสื่อมของเซลล์ที่สร้างโดปามีนในซับสแตนเซียไนกราโรคฮันติงตันซึ่งส่วนใหญ่เกี่ยวข้องกับความเสียหายต่อสไตรอาตัม[ 2 ] [ 4 ] โรคดี สโทเนีย และโรค เฮมิบอลลิสมัสซึ่งพบได้น้อยกว่า ปมประสาท ฐานมี ส่วน ลิมบิกซึ่งส่วนประกอบต่างๆ ได้รับชื่อที่แตกต่างกัน ได้แก่นิวเคลียสแอคคัมเบนส์ พัลลิดัมส่วนท้องและบริเวณเท็กเมนตัลส่วนท้อง (VTA) มีหลักฐานมากมายที่บ่งชี้ว่าส่วนลิมบิกนี้มีบทบาทสำคัญในการเรียนรู้รางวัลรวมถึงการรับรู้และ การทำงาน ของสมองส่วนหน้าผ่านทางเส้นทางเมโซลิมบิกจาก VTA ไปยังนิวเคลียสแอคคัมเบนส์ที่ใช้สารสื่อประสาทโดปามีน และเส้นทางเมโซคอร์ติคัลยาเสพติดที่มีฤทธิ์เสพติดสูงหลายชนิด รวมถึงโคเคนแอมเฟตามีนและนิโคตินเชื่อกันว่าออกฤทธิ์โดยการเพิ่มประสิทธิภาพของสัญญาณโดปามีนนี้ นอกจากนี้ยังมีหลักฐานที่บ่งชี้ว่าการทำงานมากเกินไปของการฉายภาพโดปามีนของ VTA มีส่วนเกี่ยวข้องกับโรคจิตเภท[ 12 ]
โครงสร้าง
ในแง่ของการพัฒนาระบบประสาทในมนุษย์ระบบ ประสาทส่วนกลางมักถูกจัดประเภทตามถุงสมอง ดั้งเดิมสามถุงแรก : ถุงหลักเหล่านี้ก่อตัวขึ้นในการพัฒนาตามปกติของท่อประสาทของตัวอ่อนและในขั้นต้นประกอบด้วยโปรเซนเซฟาลอนเมเซนเซฟาลอนและรอมเบนเซฟาลอนในแนวจากด้านหน้าไปด้านหลัง (จากหัวไปหาง) ต่อมาในการพัฒนาแต่ละส่วนจะกลายเป็นส่วนประกอบที่เล็กลง ในระหว่างการพัฒนา เซลล์ที่เคลื่อนที่ในแนวสัมผัสเพื่อสร้างฐานสมองจะถูกควบคุมโดยปุ่มประสาทด้านข้างและด้านใน[ 13 ]ตารางต่อไปนี้แสดงการจัดประเภทการพัฒนานี้และติดตามไปยังโครงสร้างทางกายวิภาคที่พบในฐานสมอง[ 2 ] [ 4 ] [ 14 ]โครงสร้างที่เกี่ยวข้องกับฐานสมองแสดงด้วยตัว หนา
| การแบ่งตัวขั้นต้นของท่อประสาท | การแบ่งย่อยขั้นที่สอง | ส่วนสุดท้ายในร่างกายมนุษย์ผู้ใหญ่ |
|---|---|---|
| สมองส่วนหน้า |
| |
| สมองส่วนกลาง |
|
|
| สมองส่วนหลัง |
|

ปมประสาทฐานเป็นส่วนประกอบพื้นฐานของสมองใหญ่ตรงกันข้ามกับ ชั้น คอร์เท็กซ์ที่เรียงตัวอยู่บนพื้นผิวของสมองส่วนหน้า ปมประสาทฐานเป็นกลุ่มของมวลเนื้อสีเทา ที่แตกต่างกัน ซึ่งอยู่ลึกเข้าไปในสมองไม่ไกลจากจุดเชื่อมต่อของทาลามัสพวกมันอยู่ด้านข้างและล้อมรอบทาลามัส[ 15 ]เช่นเดียวกับส่วนอื่นๆ ของสมอง ปมประสาทฐานประกอบด้วยด้านซ้ายและด้านขวาซึ่งเป็นภาพสะท้อนเสมือนจริงของกันและกัน
ในทางกายวิภาคศาสตร์ กลุ่มปมประสาทฐาน (basal ganglia) แบ่งออกเป็นสี่โครงสร้างที่แตกต่างกัน ขึ้นอยู่กับ ตำแหน่งที่อยู่ ด้านบนหรือด้านหน้า (กล่าวคือ ขึ้นอยู่กับว่าอยู่ใกล้กับส่วนบนของศีรษะมากแค่ไหน) สองส่วนคือ สไตรอาตัม (striatum)และพัลลิดัม (pallidum ) มีขนาดค่อนข้างใหญ่ ส่วนอีกสองส่วนคือ ซับสแตนเซีย นิกรา (substantia nigra) และนิวเคลียสซับทาลามิก (subthalamic nucleus ) มีขนาดเล็กกว่า ในภาพประกอบด้านขวา ภาพตัดขวาง สอง ส่วนของสมองมนุษย์แสดงตำแหน่งของส่วนประกอบของกลุ่มปมประสาทฐาน ที่น่าสังเกต (และไม่ปรากฏในภาพตัดขวางนี้) คือ นิวเคลียสซับทาลามิกและซับสแตนเซีย นิกราอยู่ลึกเข้าไปด้านหลัง ( posteriorly ) ในสมองมากกว่าสไตรอาตัมและพัลลิดัม
สไตรอาตัม

สไตรอาตัมเป็นโครงสร้างใต้เปลือกสมองที่โดยทั่วไปแบ่งออกเป็นสไตรอาตัมส่วนหลังและสไตรอาตัมส่วนหน้าสไตรอาตัมส่วนหลังยังแบ่งออกเป็นส ไตร อาตัมส่วนหลังด้านในและ สไตรอาตั มส่วนหลังด้าน นอกอีกด้วย [ 16 ] [ 17 ] [ 18 ]
สไตรอาตัมประกอบด้วยเซลล์ประสาทหนามขนาดกลาง เป็นส่วนใหญ่ เซลล์ประสาท GABAergicเหล่านี้ส่งสัญญาณไปยังโกลบัสพัลลิดัสภายนอก (ด้านข้าง) และโกลบัสพัลลิดัสภายใน (ตรงกลาง) รวมถึงซับสแตนเซียไนกราพาร์สเรติคูลาตาการส่งสัญญาณไปยังโกลบัสพัลลิดัสและซับสแตนเซียไนกราส่วนใหญ่เป็นโดปามีน แม้ว่า จะมีการแสดงออกของ เอนเคฟา ลิ นไดนอร์ฟินและสาร Pสไตรอาตัมยังมีเซลล์ประสาทระหว่างเซลล์ซึ่งจัดอยู่ในกลุ่มเซลล์ประสาทไนตริก (เนื่องจากการใช้ไนตริกออกไซด์เป็นสารสื่อประสาท ) เซลล์ประสาทระหว่างเซลล์โคลินเนอร์จิกที่ทำงานอย่างต่อเนื่อง (เช่น ปล่อยสารสื่อประสาทอย่างต่อเนื่องเว้นแต่จะถูกยับยั้ง) เซลล์ประสาท ที่แสดงออก พาร์วาลบูมินและเซลล์ประสาทที่แสดงออกแคลเรตินิ น [ 19 ]สไตรอาตัมส่วนหลังได้รับ อินพุต กลูตาเมอร์จิก จำนวนมาก จากคอร์เทกซ์ รวมถึง อินพุต โดปามีนจากซับสแตนเซียไนกราพาร์สคอมแพคตา โดยทั่วไปแล้ว striatum ส่วนหลังถือว่าเกี่ยวข้องกับกิจกรรมรับรู้และสั่งการ striatum ส่วนหน้าได้รับอินพุตกลูตาเมอร์จิกจากบริเวณลิมบิก เช่นเดียวกับอินพุตโดปามีนเนอร์จิกจาก VTA ผ่านทางเส้นทางเมโซลิมบิกเชื่อกันว่า striatum ส่วนหน้ามีบทบาทในการให้รางวัลและหน้าที่ลิมบิกอื่นๆ[ 20 ] striatum ส่วนหลังถูกแบ่งออกเป็นcaudateและputamenโดยแคปซูลภายในในขณะที่ striatum ส่วนหน้าประกอบด้วย นิวเคลียส แอคคัมเบนส์และปุ่มรับกลิ่น[ 21 ] [ 22 ] caudate มีบริเวณการเชื่อมต่อหลักสามบริเวณ โดยส่วนหัวของ caudate แสดงการเชื่อมต่อกับคอร์เทกซ์พรีฟรอนทัล คอร์เทกซ์ซิงกู เลต และอะมิกดาลา ส่วน ลำตัวและหางแสดงความแตกต่างระหว่างขอบด้านหลังและด้านข้าง และ caudate ส่วนหน้า ซึ่งฉายไปยังบริเวณรับรู้และสั่งการและลิมบิกของ striatum ตามลำดับ[ 23 ]เส้นใยสไตรอาโตพัลลิดัลเชื่อมต่อสไตรอาตัมกับพัลลิดัส
พัลลิดัม
พัลลิดัมประกอบด้วยโครงสร้างขนาดใหญ่ที่เรียกว่าโกลบัส พัลลิดัส ("ลูกโลกสีซีด") พร้อมกับส่วนขยายด้านล่างที่เล็กกว่าที่เรียกว่าเวนทรัล พัลลิดัม โกลบัส พัลลิดัสปรากฏเป็นมวลประสาทเดียว แต่สามารถแบ่งออกเป็นสองส่วนที่มีหน้าที่แตกต่างกัน คือโกลบัส พัลลิดัสภายใน (GPi) และโกลบัส พัลลิดัสภายนอก (GPe) [ 2 ]ทั้งสองส่วนประกอบด้วยเซลล์ประสาท GABAergic เป็นหลัก ซึ่งจึงมีผลยับยั้งต่อเป้าหมายของพวกมัน ทั้งสองส่วนมีส่วนร่วมในวงจรประสาท ที่แตกต่างกัน GPe รับอินพุตส่วนใหญ่จากสไตรอาตัม และส่งสัญญาณไปยังนิวเคลียสซับทาลามิก GPi รับสัญญาณจากสไตรอาตัมผ่านทางเส้นทาง "ตรง" และ "อ้อม" เซลล์ประสาทพัลลิดัมทำงานโดยใช้หลักการปลดปล่อยการยับยั้ง เซลล์ประสาทเหล่านี้จะยิงในอัตราสูงคงที่เมื่อไม่มีอินพุต และสัญญาณจากสไตรอาตัมจะทำให้พวกมันหยุดหรือลดอัตราการยิงลง เนื่องจากเซลล์ประสาทพัลลิดัมเองมีผลยับยั้งต่อเป้าหมายของมัน ผลสุทธิของการป้อนข้อมูลจากสไตรอาตัมไปยังพัลลิดัมจึงเป็นการลดการยับยั้งแบบต่อเนื่องที่เซลล์พัลลิดัมกระทำต่อเป้าหมาย (การลดการยับยั้ง) พร้อมกับอัตราการยิงที่เพิ่มขึ้นในเป้าหมาย
สารสีดำ

ซับสแตนเชีย นิกรา (Substantia nigra) เป็น ส่วนเนื้อสีเทา ในสมองส่วนกลางของฐานสมอง (basal ganglia) ซึ่งประกอบด้วยสองส่วน คือ พาร์ส คอมแพคตา ( Pars Compacta หรือ SNc) และ พาร์ส เรติคูลาตา (Pars Reticulataหรือ SNr) SNr มักทำงานร่วมกับ GPi และกลุ่ม SNr–GPi จะยับยั้งทาลามัส อย่างไรก็ตาม ซับสแตนเชีย นิกรา พาร์ส คอมแพคตา (SNc) ผลิตสารสื่อประสาทโดปามีนซึ่งมีความสำคัญมากในการรักษาสมดุลในเส้นทางสไตรอาตัม (striatal pathway) ส่วนวงจรด้านล่างจะอธิบายบทบาทและการเชื่อมต่อวงจรของแต่ละส่วนประกอบของฐานสมอง
นิวเคลียสซับทาลามิก
นิวเคลียสซับทาลามิกเป็น ส่วนของเนื้อเทา ในไดเอนเซฟาลอนของฐานสมอง และเป็นส่วนเดียวของฐานสมองที่สร้างสารสื่อประสาทกระตุ้นอย่างกลูตาเมตบทบาทของนิวเคลียสซับทาลามิกคือการกระตุ้นคอมเพล็กซ์ SNr–GPi และเป็นส่วนหนึ่งของเส้นทางทางอ้อม นิวเคลียสซับทาลามิกรับสัญญาณยับยั้งจากส่วนภายนอกของโกลบัสพัลลิดัสและส่งสัญญาณกระตุ้นไปยัง GPi
การเชื่อมต่อวงจร
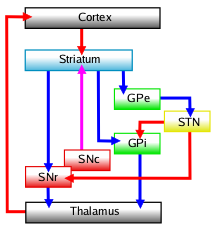


มีการเสนอแบบจำลองวงจรและการทำงานของฐานสมองหลายแบบ แม้ว่าจะมีการตั้งคำถามเกี่ยวกับการแบ่งแยกเส้นทางตรงและเส้นทางอ้อม อย่างเคร่งครัด การทับซ้อนที่เป็นไปได้ และการควบคุม[ 24 ]แบบจำลองวงจรได้พัฒนาขึ้นตั้งแต่แบบจำลองแรกที่เสนอโดยDeLong ในช่วงทศวรรษ 1990 ในแบบจำลองการประมวลผล แบบขนาน ซึ่งคอร์เทกซ์และซับสแตนเซียไนกราพาร์สคอมแพคตาฉายภาพไปยังดอร์ซัลสไตรอาตัมทำให้เกิดเส้นทางอ้อมที่ยับยั้งและเส้นทางตรงที่กระตุ้น
- กลไกการยับยั้งทางอ้อมเกี่ยวข้องกับการยับยั้ง globus pallidus externusซึ่งทำให้ globus pallidus internus ถูกปลดปล่อยการยับยั้ง (ผ่านทาง STN) ส่งผลให้globus pallidus internusสามารถยับยั้ง thalamus ได้
- เส้นทางตรงหรือเส้นทางกระตุ้นเกี่ยวข้องกับการปลดปล่อยการยับยั้งของทาลามัสผ่านการยับยั้งของ GPi/SNr อย่างไรก็ตาม ความเร็วของเส้นทางตรงจะไม่สอดคล้องกับเส้นทางอ้อมในแบบจำลองนี้ ซึ่งนำไปสู่ปัญหา เพื่อแก้ไขปัญหานี้ จึงมีการเสนอเส้นทางไฮเปอร์ไดเร็กต์ที่คอร์เทกซ์ส่งการฉายภาพกลูตาเมอร์จิกผ่านนิวเคลียสซับทาลามิกเพื่อกระตุ้น GPe ที่ยับยั้งภายใต้แบบจำลองศูนย์กลางล้อมรอบ ตลอด จนเส้นทางอ้อมที่สั้นกว่า
แม้ว่าจะถูกนำไปใช้เป็นแบบไล่ระดับโดยไม่มีขอบเขตที่แน่นอน (หรือผนังกั้นภายในนิวเคลียส) วงจรของฐานสมองมักจะถูกแบ่งออกเป็นห้าเส้นทาง ได้แก่ เส้นทางลิมบิกหนึ่งเส้นทาง เส้นทางเชื่อมโยง (พรีฟรอนทัล) สองเส้นทาง เส้นทางควบคุมการเคลื่อนไหวของดวงตาหนึ่งเส้นทาง และเส้นทางมอเตอร์หนึ่งเส้นทาง[ 25 ] บางครั้งเส้นทางมอเตอร์และเส้นทางควบคุมการเคลื่อนไหวของดวงตาจะถูกจัดกลุ่มเป็นเส้นทางมอเตอร์เดียว นอกจากนี้ แผนผังแบบง่ายๆ ที่แบ่งออกเป็นสามโดเมน (มอเตอร์ เชื่อมโยง และลิมบิก) ก็ได้รับความนิยม[ 26 ]เส้นทางทั่วไปทั้งห้าเส้นทางถูกจัดเรียงดังนี้: [ 27 ]
- วงจรการทำงานของมอเตอร์เกี่ยวข้องกับการส่งสัญญาณจากบริเวณเสริมการเคลื่อนไหว (supplementary motor area) , บริเวณพรีมอเตอร์โค้ง (arcuate premotor area), เปลือกสมองส่วนการเคลื่อนไหวหลัก (primary motor cortex) และเปลือกสมองส่วนรับความรู้สึก (somatosensory cortex) ไปยังพิวทาเมน (putamen) ซึ่งส่งสัญญาณต่อไปยัง GPi ด้านข้างส่วนล่าง (ventrolateral GPi) และ SNr ด้านข้างส่วนท้าย (caudolateral SNr) ซึ่งส่งสัญญาณต่อไปยังเปลือกสมองผ่านทาง ventralis lateralis pars medialis และ ventralis lateralis pars oralis
- วงจรควบคุมการเคลื่อนไหวของดวงตาเกี่ยวข้องกับการส่งสัญญาณจากบริเวณควบคุมการเคลื่อนไหวของดวงตาด้านหน้าคอร์เทกซ์พรีฟรอนทัลด้านข้างส่วนบน (DLPFC) และคอร์เทกซ์ข้างขมับส่วนหลังไปยังนิวเคลียสคอเดต ไปยัง GPi ด้านหลังส่วนกลางและ SNr ด้านข้างส่วนล่าง สุดท้ายวนกลับเข้าสู่คอร์เทกซ์ผ่านทาง ventralis anterior pars magnocellularis ด้านข้าง (VAmc)
- เส้นทางการรับรู้/การเชื่อมโยงแรกเสนอเส้นทางจาก DLPFC ไปยัง caudate dorsolateral ตามด้วยการฉายภาพไปยัง GPi lateral dorsomedial และ SNr rostral ก่อนที่จะฉายภาพไปยัง VAmc lateral และ pars magnocellularis medial
- เส้นทางการรับรู้/การเชื่อมโยงที่สองที่เสนอคือวงจรที่ฉายจากคอร์เทกซ์วงโคจร ด้านข้าง ไจรัสขมับ และคอร์เทกซ์ซิงกูเลตด้านหน้าไปยังคอเดตเวนโทรมีเดียล ตามด้วยการฉายไปยัง GPi ด้านข้างและ SNr ด้านหน้าและด้านข้าง ก่อนที่จะวนกลับเข้าสู่คอร์เทกซ์ผ่านทาง VAmc ด้านในและแมกโนเซลลูลาริสด้านใน
- วงจรลิมบิกที่เกี่ยวข้องกับการฉายภาพจาก ACC, ฮิปโปแคมปัส , เอนโทไรนัลคอร์ เทกซ์ และอินซูลา ไปยังเวนทรัลสไตรอาตั มจากนั้นไปยังรอสโทรดอร์ซัล GPi, เวนทรัลพัลลิดัมและรอสโทรดอร์ซัล SNr ตามด้วยวงจรวนกลับเข้าสู่คอร์เทกซ์ผ่านส่วนหลังด้านในของนิวเคลียสดอร์ซัลส่วนกลาง[ 28 ]อย่างไรก็ตาม มีการเสนอการแบ่งย่อยของวงจรเพิ่มเติมมากถึง 20,000 วงจร[ 29 ]
เป็นที่ทราบกันว่าวงจรเหล่านี้มีปฏิสัมพันธ์กัน (อย่างน้อย) ในระดับคอร์ติโค-คอร์ติคัล (เส้นใย U) ระดับคอร์ติโค-สไตรเอตัม (โดยการฉายภาพแบบกระจายจากคอร์เทกซ์ไปยังสไตรเอตัม) ระดับทาลามัส-คอร์ติคัล (โดยการเชื่อมต่อแบบกระจายระหว่างทาลามัสและคอร์เทกซ์) และระดับสไตรเอตัม-นิกราล[ 30 ]ปฏิสัมพันธ์หลังนี้ได้รับการอธิบายในรายละเอียดมากขึ้นโดยSuzanne Haberและเพื่อนร่วมงานใน 'แบบจำลองเกลียว' ของพวกเขา ซึ่งตั้งสมมติฐานว่าสไตรเอตัมส่วนล่าง (วงจรลิมบิก) สามารถมีอิทธิพลต่อสไตรเอตัมส่วนบน (วงจรการเคลื่อนไหว) ผ่านเซลล์โดปามีนในสมองส่วนกลาง (บริเวณเท็กเมนทัลส่วนล่าง ซับสแตนเซีย นิกรา พาร์ส คอมแพคตา และบริเวณอื่นๆ) ในแบบจำลองนี้ การเชื่อมต่อจากบริเวณเท็กเมนทัลส่วนล่างไปยังบริเวณเปลือกของนิวเคลียสแอคคัมเบนส์ก่อให้เกิดวงจรแบบ "ปิด" ที่เชื่อมต่อกัน อย่างไรก็ตาม การฉายภาพเหล่านี้ยังขยายออกไปด้านข้างเพื่อส่งผลต่อเซลล์ประสาทโดปามีนที่ส่งสัญญาณไปยังส่วนที่เหลือของสไตรอาตัมส่วนล่าง ทำให้เกิดส่วนเริ่มต้นของวงจรป้อนไปข้างหน้าหรือ 'เกลียว' เกลียวนี้จะดำเนินต่อไปผ่านเส้นทางสไตรอาโต-นิกโร-สไตรอาตัม โดยที่ VS ส่งผลต่อบริเวณสไตรอาตัมที่เกี่ยวข้องกับการรับรู้และการเคลื่อนไหวผ่านเซลล์ประสาทโดปามีนในสมองส่วนกลาง[ 31 ] [ 32 ]
เส้นทางตรงซึ่งมีต้นกำเนิดมาจากดอร์ซัลส ไตรอาตัมจะยับยั้ง GPi และ SNr ส่งผลให้เกิดการปลดปล่อยการยับยั้งหรือการกระตุ้นสุทธิของทาลามัส เส้นทางนี้ประกอบด้วยเซลล์ประสาทหนามขนาดกลาง (MSNs) ที่แสดงออกถึงตัวรับโดปามีน D1 ตัวรับอะเซทิลโคลีนมัสคารินิก M4และตัวรับอะดีโนซีน A1 [ 33 ] เส้นทางตรงนี้ได้รับการเสนอให้ช่วยอำนวยความสะดวกในการเคลื่อนไหว การกำหนดเวลาของการเคลื่อนไหว การควบคุมหน่วยความจำในการทำงานและการตอบสนองการเคลื่อนไหวต่อสิ่งเร้าเฉพาะ[ 29 ]
เส้นทางทางอ้อม (ยาว) เริ่มต้นจากดอร์ซัลสไตรอาตัมและยับยั้ง GPe ส่งผลให้ GPi คลายการยับยั้งและยับยั้งทาลามัสได้ เส้นทางนี้ประกอบด้วย MSNs ที่แสดงออกถึงตัวรับโดปามีน D2 , ตัวรับอะเซทิลโคลีนมัสคารินิก M1และตัวรับอะดีโนซีน A2a [ 33 ] เส้นทางนี้ได้รับการเสนอว่าส่งผลให้เกิดการยับยั้งการเคลื่อนไหวโดยรวม (การยับยั้งกิจกรรมการเคลื่อนไหวทั้งหมด) และการยุติการตอบสนอง นอกจากนี้ยังมีการเสนอเส้นทางทางอ้อมที่สั้นกว่า ซึ่งเกี่ยวข้องกับการกระตุ้นคอร์เทกซ์ของนิวเคลียสซับทาลามิกส่งผลให้เกิดการกระตุ้น GPe โดยตรงและยับยั้งทาลามัส เส้นทางนี้ได้รับการเสนอว่าส่งผลให้เกิดการยับยั้งโปรแกรมการเคลื่อนไหวเฉพาะตามการเรียนรู้แบบเชื่อมโยง[ 29 ]
การรวมกันของเส้นทางทางอ้อมเหล่านี้ส่งผลให้เกิดเส้นทางไฮเปอร์ไดเร็กต์ซึ่งส่งผลให้เกิดการยับยั้งอินพุตของฐานสมองนอกเหนือจากจุดโฟกัสเฉพาะจุดหนึ่ง ได้รับการเสนอให้เป็นส่วนหนึ่งของทฤษฎีศูนย์กลางล้อมรอบ [ 34 ] [ 35 ] เส้นทางไฮเปอร์ไดเร็กต์นี้ได้รับการเสนอให้ยับยั้งการตอบสนองก่อนกำหนด หรือยับยั้งฐานสมองโดยรวมเพื่อให้คอร์เทกซ์สามารถควบคุมจากบนลงล่างได้อย่างเฉพาะเจาะจงมากขึ้น[ 29 ]
ปฏิสัมพันธ์ของเส้นทางเหล่านี้กำลังเป็นที่ถกเถียงกันอยู่ บางคนกล่าวว่าเส้นทางทั้งหมดจะต่อต้านกันโดยตรงในลักษณะ "ผลักและดึง" ในขณะที่บางคนสนับสนุนทฤษฎีศูนย์กลางล้อมรอบซึ่งอินพุตที่มุ่งเน้นไปยังคอร์เทกซ์จะได้รับการปกป้องโดยการยับยั้งอินพุตที่แข่งขันกันโดยเส้นทางทางอ้อมที่เหลือ[ 29 ]
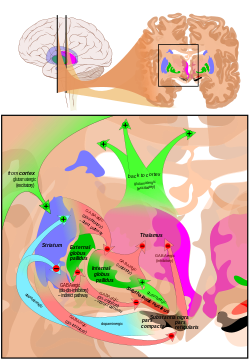
สารสื่อประสาท
ฐานสมองได้รับ อินพุต กลูตาเมต จำนวนมาก โดยมีเส้นใยนำออกGABAergic เป็นหลัก เส้นทาง โคลินเนอร์จิก ที่ปรับเปลี่ยน โดปามีนจำนวนมากในเส้นทางที่มาจากบริเวณเท็กเมนทัลด้านล่างและซับสแตน เซียไนกรา รวมถึง นิวโร เปปไทด์ ต่างๆ นิวโรเปป ไทด์ที่พบในฐานสมอง ได้แก่สาร P , นิวโรคินิน A , โคลีซิสโตคินิน , นิวโรเท นซิน , นิวโร คินิน B , นิวโรเปปไทด์ Y , โซมาโตสแต ติน , ไดนอร์ฟินและ เอนเค ฟาลิน นิวโรโมดูเลเตอร์อื่นๆ ที่พบในฐานสมอง ได้แก่ไนตริกออกไซด์ , คาร์บอนมอนอกไซด์และฟีนิลเอทิลามีน[ 36 ]
การเชื่อมต่อเชิงฟังก์ชัน
การเชื่อมต่อการทำงานที่วัดโดยการทำงานร่วมกันของภูมิภาคในระหว่างการศึกษาภาพประสาทการทำงานนั้น สอดคล้องกับแบบจำลองการประมวลผลแบบขนานของการทำงานของปมประสาทฐานโดยทั่วไป พูตาเมนโดยทั่วไปจะทำงานร่วมกับบริเวณมอเตอร์ เช่นบริเวณมอเตอร์เสริมคอร์เทกซ์ซิงกูเลตด้านหน้าส่วนท้ายและคอร์เทกซ์มอเตอร์หลักในขณะที่คอเดตและพูตาเมนส่วนหน้ามักจะทำงานร่วมกับ ACC ส่วนหน้าและ DLPFC บ่อยกว่า สไตรอาตัมส่วนล่างมีความสัมพันธ์อย่างมีนัยสำคัญกับอะมิกดาลาและฮิปโปแคมปัส ซึ่งแม้ว่าจะไม่ได้รวมอยู่ในแบบจำลองปมประสาทฐานในขั้นต้น แต่ก็เป็นส่วนเพิ่มเติมในแบบจำลองล่าสุด[ 37 ]
การทำงาน
การเคลื่อนไหวของดวงตา
หน้าที่หนึ่งที่ได้รับการศึกษาอย่างเข้มข้นของฐานสมองคือบทบาทในการควบคุม การเคลื่อนไหว ของดวงตา[ 38 ]การเคลื่อนไหวของดวงตาได้รับอิทธิพลจากเครือข่ายบริเวณสมองที่กว้างขวางซึ่งมาบรรจบกันที่บริเวณสมองส่วนกลาง ที่เรียกว่า superior colliculus (SC) SC เป็นโครงสร้างแบบหลายชั้นซึ่งชั้นต่างๆ สร้างแผนที่ เรตินาโทปิกสองมิติของพื้นที่การมองเห็น "การกระแทก" ของกิจกรรมประสาทในชั้นลึกของ SC จะขับเคลื่อนการเคลื่อนไหวของดวงตาที่มุ่งไปยังจุดที่สอดคล้องกันในอวกาศ
SC ได้รับการฉายภาพยับยั้งที่แข็งแกร่งจากฐานสมอง ซึ่งมีต้นกำเนิดมาจากซับสแตนเซียนิกรา พาร์ส เรติคูลาตา (SNr) [ 38 ]เซลล์ประสาทใน SNr มักจะยิงอย่างต่อเนื่องในอัตราที่สูง แต่เมื่อเริ่มมีการเคลื่อนไหวของดวงตา เซลล์ประสาทเหล่านั้นจะ "หยุดชั่วคราว" ทำให้ SC หลุดพ้นจากการยับยั้ง การเคลื่อนไหวของดวงตาทุกประเภทเกี่ยวข้องกับการ "หยุดชั่วคราว" ใน SNr อย่างไรก็ตาม เซลล์ประสาท SNr แต่ละเซลล์อาจเกี่ยวข้องกับการเคลื่อนไหวบางประเภทมากกว่าประเภทอื่น เซลล์ประสาทในบางส่วนของนิวเคลียสคอเดตยังแสดงกิจกรรมที่เกี่ยวข้องกับการเคลื่อนไหวของดวงตา เนื่องจากเซลล์ส่วนใหญ่ในนิวเคลียสคอเดตยิงในอัตราที่ต่ำมาก กิจกรรมนี้จึงมักปรากฏเป็นการเพิ่มขึ้นของอัตราการยิง ดังนั้น การเคลื่อนไหวของดวงตาจึงเริ่มต้นด้วยการกระตุ้นในนิวเคลียสคอเดต ซึ่งยับยั้ง SNr ผ่านการฉายภาพ GABAergic โดยตรง ซึ่งจะปลดปล่อยการยับยั้ง SC ในที่สุด
บทบาทในการสร้างแรงจูงใจ
โดปามีนนอกเซลล์ในฐานสมองส่วนกลาง (basal ganglia) มีความเชื่อมโยงกับสภาวะแรงจูงใจในสัตว์ฟันแทะ โดยระดับสูงเชื่อมโยงกับสภาวะอิ่มเอมใจ ระดับปานกลางเชื่อมโยงกับการแสวงหา และระดับต่ำเชื่อมโยงกับความรังเกียจ วงจรลิมบิก-ฐานสมองส่วนกลางได้รับอิทธิพลอย่างมากจากโดปามีน นอกเซลล์ การเพิ่มขึ้นของโดปามีนส่งผลให้เกิดการยับยั้งของเวน ท รัลพัลลิดัม (Ventral pallidum ) เอนโทเพดุนคูลาร์นิวเคลียส (entopeduncular nucleus) และ ซับสแตนเซียไนกราพาร์สเรติคูลาตา (substantia nigra pars reticulata ) ส่งผลให้เกิดการปลดปล่อยการยับยั้งของทาลามัส (thalamus) แบบจำลองของเส้นทาง D1 โดยตรงและเส้นทาง D2 โดยอ้อมนี้อธิบายว่าทำไมสารกระตุ้นเฉพาะของตัวรับแต่ละตัวจึงไม่ให้รางวัล เนื่องจากกิจกรรมในทั้งสองเส้นทางจำเป็นต่อการปลดปล่อยการยับยั้ง การปลดปล่อยการยับยั้งของทาลามัสจะนำไปสู่การกระตุ้นของเปลือกสมองส่วนหน้า (prefrontal cortex)และเวนทรัลสไตรอาตัม (ventral striatum)ซึ่งมีความเฉพาะเจาะจงต่อกิจกรรม D1 ที่เพิ่มขึ้นซึ่งนำไปสู่รางวัล[ 28 ]นอกจากนี้ยังมีหลักฐานจากการศึกษาทางสรีรวิทยาไฟฟ้าของไพรเมตที่ไม่ใช่มนุษย์และมนุษย์ที่แสดงให้เห็นว่าโครงสร้างฐานสมองอื่นๆ รวมถึง globus pallidus internus และ subthalamic nucleus มีส่วนเกี่ยวข้องในการประมวลผลรางวัล[ 39 ]
การตัดสินใจ
มีการเสนอแบบจำลองสองแบบสำหรับบทบาทของฐานสมองในการตัดสินใจ แบบจำลองแรก การกระทำถูกสร้างขึ้นโดย "นักวิจารณ์" ในสไตรอาตัมส่วนล่างที่ประเมินค่า และการกระทำถูกดำเนินการโดย "ผู้กระทำ" ในสไตรอาตัมส่วนบน แบบจำลองที่สอง ฐานสมองทำหน้าที่เป็นกลไกการเลือก โดยการกระทำถูกสร้างขึ้นในคอร์เทกซ์และถูกเลือกตามบริบทโดยฐานสมอง[ 40 ]วงจรCBGTCยังเกี่ยวข้องกับการลดทอนรางวัล โดยการยิงจะเพิ่มขึ้นเมื่อได้รับรางวัลที่ไม่คาดคิดหรือมากกว่าที่คาดไว้[ 41 ]บทวิจารณ์หนึ่งสนับสนุนแนวคิดที่ว่าคอร์เทกซ์มีส่วนเกี่ยวข้องกับการเรียนรู้การกระทำโดยไม่คำนึงถึงผลลัพธ์ ในขณะที่ฐานสมองมีส่วนเกี่ยวข้องกับการเลือกการกระทำที่เหมาะสมโดยอาศัยการเรียนรู้แบบลองผิดลองถูกตามรางวัลแบบเชื่อมโยง[ 42 ]
หน่วยความจำใช้งาน
มีการเสนอว่าฐานสมองส่วนกลางทำหน้าที่ควบคุมสิ่งที่เข้าและไม่เข้าในหน่วยความจำใช้งานสมมติฐานหนึ่งเสนอว่าเส้นทางตรง (Go หรือกระตุ้น) อนุญาตให้ข้อมูลเข้าสู่คอร์เทกซ์ส่วนหน้าซึ่งข้อมูลจะคงอยู่โดยอิสระจากเส้นทาง ในขณะที่อีกทฤษฎีหนึ่งเสนอว่าเพื่อให้ข้อมูลคงอยู่ในคอร์เทกซ์ส่วนหน้า เส้นทางตรงจำเป็นต้องสะท้อนกลับอย่างต่อเนื่อง มีการเสนอว่าเส้นทางอ้อมสั้นๆ ทำหน้าที่ปิดประตูสู่คอร์เทกซ์ส่วนหน้า ในลักษณะการผลักและดึงโดยตรงที่ขัดแย้งกับเส้นทางตรง กลไกเหล่านี้ร่วมกันควบคุมการโฟกัสของหน่วยความจำใช้งาน[ 29 ]
ความสำคัญทางคลินิก
โรคของปมประสาทฐานเป็นกลุ่มของความผิดปกติของการเคลื่อนไหวที่เกิดจากผลลัพธ์ที่มากเกินไปจากปมประสาทฐานไปยังทาลามัส (สำหรับความผิดปกติแบบไฮโปไค เนติก ) หรือจากผลลัพธ์ที่ไม่เพียงพอ (สำหรับความผิดปกติแบบไฮเปอร์ไคเนติก ) ความผิดปกติแบบไฮโปไคเนติกเกิดจากผลลัพธ์ที่มากเกินไปจากปมประสาทฐาน ซึ่งยับยั้งผลลัพธ์จากทาลามัสไปยังคอร์เทกซ์ และทำให้การเคลื่อนไหวโดยสมัครใจถูกจำกัด ความผิดปกติแบบไฮเปอร์ไคเนติกเกิดจากผลลัพธ์ที่น้อยเกินไปจากปมประสาทฐานไปยังทาลามัส ซึ่งไม่ยับยั้งการฉายภาพของทาลามัสไปยังคอร์เทกซ์อย่างเพียงพอ ทำให้เกิดการเคลื่อนไหวที่ควบคุมไม่ได้/ไม่เป็นไปตามความตั้งใจ การทำงานผิดปกติของวงจรปมประสาทฐานยังสามารถนำไปสู่ความผิดปกติอื่นๆ ได้อีกด้วย[ 43 ]
ต่อไปนี้เป็นรายการของความผิดปกติ สภาวะ และอาการที่เชื่อมโยงกับฐานสมอง: [ 44 ]
- การเสพติด[ 45 ] [ 46 ]
- อะเทโทซิส[ 47 ]
- กลุ่มอาการขาดฮอร์โมนไทมัส ( กลุ่มอาการ PAP )
- โรคสมาธิสั้น (ADHD)
- อาการกระตุกเปลือกตา
- การนอนกัดฟัน
- อัมพาตสมอง : ความเสียหายของปุ่มสมองส่วนฐานระหว่างไตรมาสที่สองและสามของการตั้งครรภ์
- โรคชัก[ 47 ]
- โรคดิสโทเนีย[ 47 ] [ 48 ]
- โรคลมชัก[ 49 ]
- โรคของ Fahr [ 45 ]
- กลุ่มอาการสำเนียงต่างชาติ (FAS)
- โรคฮันติงตัน[ 45 ] [ 48 ]
- ภาวะดีซ่านในเคอร์นิคเทอรัส
- คลาโซมาเนีย
- กลุ่มอาการเลช-นัยฮาน
- โรคซึมเศร้าขั้นรุนแรง[ 45 ] [ 50 ]
- โรคย้ำคิดย้ำทำ[ 45 ] [ 51 ] [ 52 ]
- ความผิดปกติทางวิตกกังวลอื่นๆ[ 46 ] [ 52 ]
- โรคแพนดาส (ความผิดปกติ)
- โรคพาร์กินสัน[ 45 ] [ 48 ]
- กลุ่มอาการขาอยู่ไม่สุข
- โรคจิตเภท[ 45 ] [ 46 ]
- ภาวะเสียงแหบเกร็ง
- การพูดติดอ่าง[ 53 ]
- โรคชักกระตุกของไซเดนแฮม
- อาการเคลื่อนไหวผิดปกติแบบเรื้อรัง (Tardive dyskinesia ) เกิดจากการรักษาด้วยยาต้านโรคจิต เป็นเวลานาน
- กลุ่มอาการทูเร็ตต์[ 45 ]
- โรควิลสัน[ 45 ] [ 48 ]
ประวัติศาสตร์
ต้องใช้เวลากว่าที่ฐานสมองจะได้รับการยอมรับว่าเป็นระบบของนิวเคลียสสมองที่มีความสัมพันธ์กันในเชิงหน้าที่การระบุโครงสร้างใต้เปลือกสมองที่แตกต่างกันทางกายวิภาค ครั้งแรกได้รับการตีพิมพ์โดย โทมัส วิลลิสในปี ค.ศ. 1664 [ 54 ]เป็นเวลาหลายปีที่คำว่าcorpus striatum [ 55 ]ถูกใช้เพื่ออธิบายกลุ่มองค์ประกอบใต้เปลือกสมองขนาดใหญ่ ซึ่งบางส่วนถูกค้นพบในภายหลังว่าไม่มีความสัมพันธ์กันในเชิงหน้าที่[ 56 ]ในทำนองเดียวกัน เป็นเวลาหลายปีที่พิวทาเมนและนิวเคลียสคอเดตไม่ได้เชื่อมโยงกัน แต่พิวทาเมนกลับเชื่อมโยงกับพัลลิดัมในสิ่งที่เรียกว่านิวเคลียสเลนติคูลาริสหรือนิวเคลียสเลนติฟอร์มิส
การทบทวนอย่างละเอียดโดยCécileและOskar Vogt (1941) ได้ทำให้คำอธิบายของปมประสาทฐานง่ายขึ้น โดยเสนอคำว่าstriatumเพื่ออธิบายกลุ่มโครงสร้างที่ประกอบด้วยนิวเคลียสคอเดต พูตาเมน และมวลที่เชื่อมต่อพวกมันทางด้านล่าง คือ นิวเคลียสแอคคัมเบนส์ชื่อ striatum มาจากลักษณะที่เป็นลาย (เป็นเส้น) ที่เกิดจากการแผ่กระจายของกลุ่มแอกซอน striato-pallido-nigral ที่หนาแน่น ซึ่งนักกายวิภาคศาสตร์Samuel Alexander Kinnier Wilson (1912) อธิบายว่า "เหมือนดินสอ"
การเชื่อมโยงทางกายวิภาคของสไตรเอตัมกับเป้าหมายหลักคือพัลลิดัมและซับสแตนเชีย นิกราถูกค้นพบในภายหลัง ชื่อโกลบัส พัลลิดัส (globus pallidus ) ถูกตั้ง โดยเด อเจอรีน (Déjerine) ในปี 1822 แต่ตระกูลโวกต์ (Vogts) เสนอคำที่ง่ายกว่า คือ พั ลลิดัม (pallidum) ส่วนคำว่า โลคัส ไนเจอร์ (locus niger)ถูกนำเสนอโดย เฟลิกซ์ วิค-ดาซีร์ (Félix Vicq-d'Azyr)ในชื่อ ทาเช นัว ร์ (tache noire ) ในปี 1786 อย่างไรก็ตาม โครงสร้างนั้นต่อมาเป็นที่รู้จักกันในชื่อซับสแตนเชีย นิกรา (substantia nigra) เนื่องจากการมีส่วนร่วมของ ฟอน ซอมเมอริง ( Von Sömmering ) ในปี 1788 ความคล้ายคลึงกันทางโครงสร้างระหว่างซับสแตนเชีย นิกราและโกลบัส พัลลิดัสถูกบันทึกไว้โดยมีร์โต (Mirto) ในปี 1896 ทั้งสองส่วนนี้รวมกันเรียกว่า พัลลิโดนิกราล แอนเซมเบิล (pallidonigral ensemble) ซึ่งเป็นแกนหลักของฐานสมอง (basal ganglia) โดยรวมแล้ว โครงสร้างหลักของปมประสาทฐานเชื่อมต่อกันด้วยมัดเส้นใยสไตรอาโต-พัลลิโด-นิกราล ซึ่งผ่านพัลลิดัมข้ามแคปซูลภายในในลักษณะ "มัดเส้นใยหวีของเอดิงเกอร์" และในที่สุดก็ไปถึงซับสแตนเซีย นิกรา
โครงสร้างเพิ่มเติมที่ต่อมามีความเกี่ยวข้องกับปมประสาทฐาน ได้แก่ "ร่างกายของลุยส์" (ค.ศ. 1865) (หรือนิวเคลียสของลุยส์) หรือนิวเคลียสซับทาลามิกซึ่งหากเกิดความเสียหายกับนิวเคลียสนี้จะทำให้เกิดความผิดปกติของการเคลื่อนไหว เมื่อไม่นานมานี้ บริเวณอื่นๆ เช่นนิวเคลียสเซนโทรมีเดียนและคอมเพล็กซ์เพดุนคูโลพอนไทน์ก็ถูกมองว่าเป็นตัวควบคุมปมประสาทฐานเช่นกัน
ในช่วงต้นศตวรรษที่ 20 ระบบฐานสมองถูกเชื่อมโยงกับหน้าที่การเคลื่อนไหวเป็นครั้งแรก เนื่องจากรอยโรคในบริเวณเหล่านี้มักส่งผลให้เกิดความผิดปกติในการเคลื่อนไหวในมนุษย์ (เช่นโรคชักกระตุก โรคอะ ที โทซิ สโรคพาร์กินสัน )
ศัพท์เฉพาะ
การตั้งชื่อระบบฐานสมองและส่วนประกอบต่างๆ นั้นเป็นปัญหามาโดยตลอด นักกายวิภาคศาสตร์ในยุคแรกๆ เห็นเพียงโครงสร้างทางกายวิภาคในระดับมหภาค แต่ไม่รู้เกี่ยวกับสถาปัตยกรรมระดับเซลล์หรือเคมีประสาท จึงจัดกลุ่มส่วนประกอบต่างๆ เข้าด้วยกัน ซึ่งปัจจุบันเชื่อกันว่ามีหน้าที่แตกต่างกัน (เช่น ส่วนภายในและภายนอกของโกลบัส พัลลิดัส) และตั้งชื่อที่แตกต่างกันให้กับส่วนประกอบต่างๆ ซึ่งปัจจุบันเชื่อกันว่าเป็นส่วนหนึ่งของโครงสร้างเดียวกันในเชิงหน้าที่ (เช่น นิวเคลียสคอเดตและพูตาเมน)
คำว่า "basal" มาจากข้อเท็จจริงที่ว่าองค์ประกอบส่วนใหญ่ตั้งอยู่ในส่วนฐานของสมองส่วนหน้า คำว่าgangliaเป็นคำที่ไม่ถูกต้อง: ในการใช้งานสมัยใหม่ กลุ่มเซลล์ประสาทเรียกว่า "ganglia" เฉพาะในระบบประสาทส่วนปลาย เท่านั้น ในระบบประสาทส่วนกลางเรียกว่า "nuclei" ด้วยเหตุนี้ ganglia ฐานจึงบางครั้งเรียกว่า "basal nuclei" [ 57 ] Terminologia Anatomica (1998) ซึ่งเป็นหน่วยงานระหว่างประเทศด้านการตั้งชื่อทางกายวิภาคศาสตร์ ยังคงใช้ "nuclei basales" แต่คำนี้ไม่ได้ใช้กันทั่วไป
สมาคมฐานสมองระหว่างประเทศ (IBAGS) [ 58 ]ถือว่าฐานสมองประกอบด้วยstriatum , pallidum (มีนิวเคลียสสองอัน), substantia nigra (มีส่วนประกอบสองส่วนที่แตกต่างกัน) และนิวเคลียส subthalamic อย่างไม่เป็นทางการ ในขณะที่ Terminologia anatomica ไม่รวมสองส่วนหลังนี้ นักประสาทวิทยาบางคนได้รวมนิวเคลียส centromedianของ thalamus เป็นส่วนหนึ่งของฐานสมอง[ 59 ] [ 60 ]และบางคนก็รวมนิวเคลียสpedunculopontine ด้วย [ 61 ]
สัตว์อื่นๆ
ปมประสาทฐานเป็นส่วนประกอบพื้นฐานอย่างหนึ่งของสมองส่วนหน้าและสามารถพบได้ในสัตว์มีกระดูกสันหลังทุกชนิด[ 62 ]แม้แต่ในปลาแลมเพรย์ (ซึ่งโดยทั่วไปถือว่าเป็นสัตว์มีกระดูกสันหลังที่ดั้งเดิมที่สุดชนิดหนึ่ง) ก็สามารถระบุองค์ประกอบของสไตรอาตัม พัลลิดัล และนิกราล ได้จากกายวิภาคศาสตร์และเคมี เนื้อเยื่อ [ 63 ]
ชื่อที่ใช้เรียกนิวเคลียสต่างๆ ของฐานสมองจะแตกต่างกันไปในแต่ละสายพันธุ์ ในแมวและหนู นิวเคลียส globus pallidus ภายในเรียกว่านิวเคลียสentopeduncular [ 64 ]ในนก striatum เรียกว่าpaleostriatum augmentatumและนิวเคลียส globus pallidus ภายนอกเรียกว่าpaleostriatum primitivum
ประเด็นสำคัญที่เกิดขึ้นอย่างชัดเจนในกายวิภาคเปรียบเทียบของปมประสาทฐานคือการพัฒนาระบบนี้ผ่านวิวัฒนาการในฐานะวงจรย้อนกลับเข้าสู่เปลือกสมองแบบบรรจบกัน ควบคู่ไปกับการพัฒนาและการขยายตัวของเปลือกสมอง อย่างไรก็ตาม ยังมีข้อถกเถียงเกี่ยวกับขอบเขตของการประมวลผลแบบคัดเลือกที่บรรจบกันเมื่อเทียบกับการประมวลผลแบบขนานที่แยกส่วนภายในวงจรปิดย้อนกลับของปมประสาทฐาน ไม่ว่าอย่างไรก็ตาม การเปลี่ยนแปลงของปมประสาทฐานไปเป็นระบบย้อนกลับเข้าสู่เปลือกสมองในวิวัฒนาการของสัตว์เลี้ยงลูกด้วยนมเกิดขึ้นผ่านการเปลี่ยนทิศทางของเอาต์พุตจากพัลลิดัล (หรือ "paleostriatum primitivum") จากเป้าหมายในสมองส่วนกลาง เช่น superior colliculus ดังที่เกิดขึ้นใน สมองของซอ รอปซิดไปยังบริเวณเฉพาะของทาลามัสส่วนล่าง และจากนั้นกลับไปยังบริเวณที่กำหนดของเปลือกสมองที่ประกอบเป็นกลุ่มย่อยของบริเวณเปลือกสมองเหล่านั้นที่ฉายภาพไปยังสไตรอาตัม การเปลี่ยนทิศทางอย่างฉับพลันของเส้นทางจากส่วนภายในของโกลบัส พัลลิดัสไปยังทาลามัสส่วนล่าง—ผ่านทางเส้นทางของแอนซา เลนติคูลาริส —อาจมองได้ว่าเป็นร่องรอยของการเปลี่ยนแปลงทางวิวัฒนาการของการไหลออกของปมประสาทฐานและอิทธิพลที่กำหนดเป้าหมาย
ดูเพิ่มเติม
รูปภาพเพิ่มเติม
 บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวระนาบ
บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวระนาบ บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวระนาบซาจิตัล (sagittal)
บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวระนาบซาจิตัล (sagittal) บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวขวาง
บริเวณฐานสมอง (Basal ganglia) ถูกเน้นด้วยสีเขียวในภาพ MRI T1 แนวขวาง แสดงปมประสาทฐาน (สีแดง) และโครงสร้างที่เกี่ยวข้อง (สีน้ำเงิน)
แสดงปมประสาทฐาน (สีแดง) และโครงสร้างที่เกี่ยวข้อง (สีน้ำเงิน)




ลิงก์ภายนอก
- การถ่ายภาพบริเวณฐานสมองที่USUHS
- Houk J (2007). "แบบจำลองของปมประสาทฐาน" . Scholarpedia . 2 (10): 1633. Bibcode : 2007SchpJ...2.1633H . doi : 10.4249/scholarpedia.1633 .
- คู่มือ Merck แบบโต้ตอบ แสดงภาพการผ่าตัดปมประสาทฐานแบบ 3 มิติ พร้อมส่วนที่เลือกได้
- สมาคมฐานสมองนานาชาติ
- Basal ganglia – วารสารอย่างเป็นทางการของ LIMPE (Lega Italiana per la Lotta Contro la Malattia di Parkinson, le Sindromi Extrapiramidali e le Demenze, อิตาลี), German Parkinson Society (DPG, Deutsche Parkinson Gesellschaft) และ Japanese Basal Ganglia Society (JBAGS Japan Basal Ganglia Society)
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ปมประสาทฐาน
ฐานสมอง ( BG ) หรือนิวเคลียสฐานเป็นกลุ่มของนิวเคลียสใต้เปลือกสมอง (กลุ่มของเซลล์ประสาท) ที่พบในสมองของสัตว์มีกระดูกสันหลัง
โครงสร้าง
ในแง่ของการ พัฒนาระบบประสาทในมนุษย์ ระบบ ประสาทส่วนกลาง มักถูกจัดประเภทตาม ถุงสมอง ดั้งเดิมสามถุงแรก : ถุงหลักเหล่านี้ก่อตัวขึ้นในการพัฒนาตามปกติของ ท่อประสาท ของ ตัวอ่อน และในขั้นต้นประกอบด้วย โปรเซนเซฟาลอน เม เซนเซฟาลอน และ รอมเบนเซฟาลอน...
สไตรอาตัม
สไตรอาตัมเป็นโครงสร้างใต้เปลือกสมองที่โดยทั่วไปแบ่งออกเป็น สไตรอาตัมส่วนหลัง และ สไตรอาตัมส่วนหน้า สไตรอาตัมส่วนหลังยังแบ่งออกเป็นส ไตร อาตัมส่วนหลังด้านใน และ สไตรอาตั ม ส่วนหลังด้าน นอกอีกด้วย [ 16 ] [ 17 ] [ 18 ]
พัลลิดัม
พัลลิดัมประกอบด้วยโครงสร้างขนาดใหญ่ที่เรียกว่า โกลบัส พัลลิดัส ("ลูกโลกสีซีด") พร้อมกับส่วนขยายด้านล่างที่เล็กกว่าที่เรียกว่าเวน ทรัล พัลลิดัม โก ลบัส พัลลิดัสปรากฏเป็นมวลประสาทเดียว แต่สามารถแบ่งออกเป็นสองส่วนที่มีหน้าที่แตกต่างกัน คือ โกลบัส พัลลิดัสภายใน...