อ่าน 25 นาที
รหัสพันธุกรรม
รหัสพันธุกรรม คือชุดของกฎที่ เซลล์สิ่ง มีชีวิตใช้ ใน การแปล ข้อมูลที่เข้ารหัสอยู่ในสารพันธุกรรม ( ลำดับ ดีเอ็นเอ หรือ อาร์เอ็นเอ ของกลุ่มนิวคลีโอไทด์สามตัวหรือ โคดอน ) ไปเป็น...
รหัสพันธุกรรม

รหัสพันธุกรรมคือชุดของกฎที่เซลล์สิ่ง มีชีวิตใช้ ในการแปลข้อมูลที่เข้ารหัสอยู่ในสารพันธุกรรม ( ลำดับ ดีเอ็นเอหรืออาร์เอ็นเอของกลุ่มนิวคลีโอไทด์สามตัวหรือโคดอน ) ไปเป็นโปรตีนกระบวนการแปลนี้เกิดขึ้นโดยไรโบโซมซึ่งเชื่อมต่อกรดอะมิโนที่สร้างโปรตีนตามลำดับที่กำหนดโดยอาร์เอ็นเอส่งสาร (mRNA) โดยใช้ โมเลกุล อาร์เอ็นเอถ่ายโอน (tRNA) ในการนำกรดอะมิโนและอ่าน mRNA ทีละสามนิวคลีโอไทด์รหัสพันธุกรรมมีความคล้ายคลึงกันอย่างมากในสิ่งมีชีวิตทุกชนิดและสามารถแสดงได้ในตารางอย่างง่ายที่มี 64 รายการ
โคดอนระบุว่ากรดอะมิโนใดจะถูกเพิ่มต่อไปในระหว่างการสังเคราะห์โปรตีนโดยมีข้อยกเว้นบางประการ[ 1 ] โคดอนสามนิวคลีโอไทด์ในลำดับกรดนิวคลีอิกจะระบุกรดอะมิโนเพียงตัวเดียว ยีนส่วนใหญ่ถูกเข้ารหัสด้วยรูปแบบเดียว (ดูตารางโคดอน RNA ) รูปแบบนั้นมักเรียกว่ารหัสพันธุกรรมแบบแคนอนิกหรือมาตรฐาน หรือเรียกง่ายๆ ว่ารหัสพันธุกรรม แม้ว่า จะมี รหัสที่แตกต่างกัน (เช่นใน ไมโทคอน เดรีย ) อยู่ ก็ตาม
ประวัติศาสตร์
ความพยายามที่จะเข้าใจว่าโปรตีนถูกเข้ารหัสอย่างไร เริ่มขึ้นหลังจาก มีการค้นพบ โครงสร้างของ DNAในปี 1953 ผู้ค้นพบที่สำคัญคือฟรานซิส คริก นักชีวฟิสิกส์ชาวอังกฤษ และเจมส์ วัตสัน นักชีววิทยาชาวอเมริกัน ซึ่งทำงานร่วมกันที่ห้องปฏิบัติการคาเวนดิชมหาวิทยาลัยเคมบริดจ์ ได้ตั้งสมมติฐานว่าข้อมูลไหลมาจาก DNA และมีความเชื่อมโยงระหว่าง DNA กับโปรตีน[ 2 ]จอร์จ กาโมว์นักฟิสิกส์ชาวโซเวียต-อเมริกันเป็นคนแรกที่เสนอแผนการสังเคราะห์โปรตีนจาก DNA ที่ใช้งานได้จริง[ 3 ]เขาตั้งสมมติฐานว่าต้องใช้ชุดเบสสามตัว (ทริปเล็ต) ในการเข้ารหัสกรดอะมิโนมาตรฐาน 20 ชนิดที่เซลล์สิ่งมีชีวิตใช้ในการสร้างโปรตีน ซึ่งจะทำให้มีกรดอะมิโนได้สูงสุด4³ = 64ชนิด (การเรียงสับเปลี่ยนทั้งหมดของเบสทั้งสี่ อ่านทีละสามตัว) [ 4 ]เขาตั้งชื่อปฏิสัมพันธ์ระหว่าง DNA กับโปรตีนนี้ (รหัสพันธุกรรมดั้งเดิม) ว่า "รหัสเพชร" [ 5 ]
ในปี พ.ศ. 2497 Gamow ได้ก่อตั้งองค์กรวิทยาศาสตร์ที่ไม่เป็นทางการชื่อRNA Tie Clubตามคำแนะนำของ Watson สำหรับนักวิทยาศาสตร์จากหลากหลายสาขาที่สนใจว่าโปรตีนถูกสังเคราะห์จากยีนได้อย่างไร อย่างไรก็ตาม สโมสรนี้สามารถมีสมาชิกถาวรได้เพียง 20 คนเพื่อเป็นตัวแทนของกรดอะมิโนทั้ง 20 ชนิด และมีสมาชิกกิตติมศักดิ์เพิ่มเติมอีก 4 คนเพื่อเป็นตัวแทนของนิวคลีโอไทด์ทั้ง 4 ชนิดของ DNA [ 6 ]
ผลงานทางวิทยาศาสตร์ชิ้นแรกของชมรม ซึ่งต่อมาได้รับการบันทึกว่าเป็น "หนึ่งในบทความที่ยังไม่ได้รับการตีพิมพ์ที่สำคัญที่สุดในประวัติศาสตร์วิทยาศาสตร์" [ 7 ]และ "บทความที่ยังไม่ได้รับการตีพิมพ์ที่มีชื่อเสียงที่สุดในบันทึกของชีววิทยาโมเลกุล" [ 8 ]นั้น มาจาก Crick Crick ได้นำเสนอเอกสารที่พิมพ์ดีดชื่อ "เกี่ยวกับแม่แบบที่เสื่อมสภาพและสมมติฐานอะแดปเตอร์: บันทึกสำหรับชมรม RNA Tie" [ 9 ]ให้กับสมาชิกของชมรมในเดือนมกราคม พ.ศ. 2498 ซึ่ง "เปลี่ยนวิธีคิดของเราเกี่ยวกับการสังเคราะห์โปรตีนไปโดยสิ้นเชิง" ดังที่ Watson เล่า[ 10 ]สมมติฐานนี้ระบุว่ารหัสสามตัวไม่ได้ถูกส่งต่อไปยังกรดอะมิโนอย่างที่ Gamow คิด แต่ถูกส่งผ่านโมเลกุลอื่น ซึ่งเป็นอะแดปเตอร์ ที่ทำปฏิกิริยากับกรดอะมิโน[ 8 ]ต่อมาอะแดปเตอร์นี้ถูกระบุว่าเป็น tRNA [ 11 ]
โคดอน
การทดลอง ของCrick, Brenner, Barnett และ Watts-Tobinเป็นครั้งแรกที่แสดงให้เห็นว่าโคดอนประกอบด้วยเบส DNA สามชนิด
Marshall NirenbergและJ. Heinrich Matthaeiเป็นคนแรกที่เปิดเผยธรรมชาติของโคดอนในปี 1961 [ 12 ]พวกเขาใช้ระบบที่ปราศจากเซลล์เพื่อแปล ลำดับ RNA โพลียูราซิล (เช่น UUUUU...) และค้นพบว่าพอลิเปปไทด์ที่พวกเขาสังเคราะห์ขึ้นนั้นประกอบด้วยกรดอะมิโนฟีนิลอะลานีนเพียง อย่างเดียว [ 13 ]พวกเขาจึงสรุปได้ว่าโคดอน UUU ระบุกรดอะมิโนฟีนิลอะลานีน
ต่อมามีการทดลองในห้องปฏิบัติการของSevero Ochoa ซึ่งแสดงให้เห็นว่าลำดับ RNA ของโพลีอะ ดีนีน (AAAAA...) เข้ารหัสสำหรับโพลีเปปไทด์โพลีไลซีน[ 14 ]และลำดับ RNA ของโพลีไซโตซีน (CCCCC...) เข้ารหัสสำหรับโพลีเปปไทด์โพลีโพรลีน [ 15 ] ดังนั้นโคดอน AAA จึงระบุกรดอะมิโนไลซีนและโคดอน CCC ระบุกรดอะมิโนโพรลีนจากนั้นจึงกำหนดโคดอนที่เหลือส่วนใหญ่ โดยใช้ โคพอลิเมอร์ต่างๆ
งานวิจัยต่อมาของHar Gobind Khoranaได้ระบุรหัสพันธุกรรมส่วนที่เหลือ หลังจากนั้นไม่นานRobert W. Holleyได้กำหนดโครงสร้างของtransfer RNA (tRNA) ซึ่งเป็นโมเลกุลอะแดปเตอร์ที่ช่วยอำนวยความสะดวกในกระบวนการแปล RNA เป็นโปรตีน งานวิจัยนี้อิงตามการศึกษาของ Ochoa ก่อนหน้านี้ ซึ่งส่งผลให้ Ochoa ได้รับรางวัลโนเบลสาขาสรีรวิทยาหรือการแพทย์ในปี 1959 จากงานวิจัยเกี่ยวกับเอนไซม์วิทยาของการสังเคราะห์ RNA [ 16 ]
จากการขยายงานนี้ Nirenberg และPhilip Lederได้เปิดเผยลักษณะสามตัวอักษรของรหัสและถอดรหัสโคดอน ในการทดลองเหล่านี้mRNA หลายชุด ถูกส่งผ่านตัวกรองที่มีไรโบโซมซึ่งเป็นส่วนประกอบของเซลล์ที่แปล RNA เป็นโปรตีน สามตัวอักษรที่ไม่ซ้ำกันส่งเสริมการจับกันของ tRNA เฉพาะกับไรโบโซม Leder และ Nirenberg สามารถกำหนดลำดับของโคดอน 54 จาก 64 โคดอนในการทดลองของพวกเขาได้[ 17 ] Khorana, Holley และ Nirenberg ได้รับรางวัลโนเบล (1968) จากผลงานของพวกเขา[ 18 ]
โคดอนหยุดทั้งสามตัวได้รับการตั้งชื่อโดยผู้ค้นพบ Richard Epstein และ Charles Steinberg โดย "Amber" ได้รับการตั้งชื่อตามเพื่อนของพวกเขา Harris Bernstein ซึ่งนามสกุลของเขามีความหมายว่า "อำพัน" ในภาษาเยอรมัน[ 19 ]โคดอนหยุดอีกสองตัวได้รับการตั้งชื่อว่า "ochre" และ "opal" เพื่อให้สอดคล้องกับธีม "ชื่อสี"
รหัสพันธุกรรมที่ขยายใหญ่ขึ้น (ชีววิทยาเชิงสังเคราะห์)
ในแวดวงวิชาการโดยทั่วไป แนวคิดเรื่องวิวัฒนาการของรหัสพันธุกรรมจากรหัสพันธุกรรมดั้งเดิมที่ไม่ชัดเจนไปสู่รหัสที่กำหนดไว้อย่างชัดเจน ("แช่แข็ง") พร้อมด้วยกรดอะมิโนมาตรฐาน 20 (+2) ชนิด ได้รับการยอมรับอย่างกว้างขวาง[ 20 ] อย่างไรก็ตาม มีความคิดเห็น แนวคิด วิธีการ และความคิดที่แตกต่างกันเกี่ยวกับวิธีที่ดีที่สุดในการเปลี่ยนแปลงรหัสพันธุกรรมในเชิงทดลอง แม้แต่แบบจำลองที่เสนอขึ้นมาก็ยังทำนาย "จุดเริ่มต้น" สำหรับการบุกรุกของกรดอะมิโนสังเคราะห์ในรหัสพันธุกรรม[ 21 ]
ตั้งแต่ปี พ.ศ. 2544 มีการเพิ่มกรดอะมิโนที่ไม่เป็นธรรมชาติ 40 ชนิดเข้าไปในโปรตีนโดยการสร้างโคดอนที่ไม่ซ้ำกัน (การเข้ารหัสใหม่) และคู่ transfer-RNA:aminoacyl – tRNA-synthetase ที่สอดคล้องกันเพื่อเข้ารหัสด้วยคุณสมบัติทางกายภาพเคมีและชีวภาพที่หลากหลาย เพื่อใช้เป็นเครื่องมือในการสำรวจโครงสร้างและหน้าที่ของโปรตีน หรือเพื่อสร้างโปรตีนใหม่หรือโปรตีนที่ได้รับการปรับปรุง[ 22 ] [ 23 ]
H. Murakami และ M. Sisido ได้ขยายโคดอนบางส่วนให้มีเบสสี่และห้าเบสSteven A. Bennerได้สร้างโคดอนที่ 65 ที่ใช้งานได้ ( ในร่างกาย ) [ 24 ]
ในปี 2015 N. Budisa , D. Söllและเพื่อนร่วมงานได้รายงานการแทนที่อย่างสมบูรณ์ของ สารตกค้าง ทริปโตแฟน ทั้งหมด 20,899 ตัว (รหัส UGG) ด้วยไทเอโนไพร์โรล-อะลานีนที่ไม่เป็นธรรมชาติในรหัสพันธุกรรมของแบคทีเรียE. coli [ 25 ]
ในปี 2016 สิ่งมีชีวิตกึ่งสังเคราะห์ที่เสถียรตัวแรกถูกสร้างขึ้น มันเป็นแบคทีเรีย (เซลล์เดียว) ที่มีเบสสังเคราะห์สองชนิด (เรียกว่า X และ Y) เบสเหล่านี้สามารถอยู่รอดได้แม้การแบ่งเซลล์[ 26 ] [ 27 ]
ในปี 2017 นักวิจัยในเกาหลีใต้รายงานว่าพวกเขาได้ดัดแปลงพันธุกรรมหนูด้วยรหัสพันธุกรรมที่ขยายออกไปซึ่งสามารถผลิตโปรตีนที่มีกรดอะมิโนที่ไม่เป็นธรรมชาติได้[ 28 ]
ในเดือนพฤษภาคม พ.ศ. 2562 นักวิจัยรายงานการสร้างสายพันธุ์ใหม่ของ แบคทีเรีย E. coliที่ชื่อว่า "Syn61" สายพันธุ์นี้มี จีโนม สังเคราะห์ ทั้งหมด ที่ได้รับการปรับโครงสร้างใหม่ (ขยายส่วนที่ทับซ้อนกันทั้งหมด) เข้ารหัสใหม่ (ลบการใช้โคดอน 3 ใน 64 โคดอนออกไปโดยสมบูรณ์) และได้รับการดัดแปลงเพิ่มเติมเพื่อลบ tRNA และปัจจัยการปลดปล่อยที่ไม่จำเป็นออกไป สายพันธุ์นี้ยังคงมีชีวิตอยู่ได้และเติบโตช้ากว่าสายพันธุ์ดั้งเดิม " MDS42 " ถึง 1.6 เท่า [ 29 ] [ 30 ]
ในปี 2025 นักวิจัยรายงานสายพันธุ์ "Syn57" ใหม่ ซึ่งกำจัดการใช้โคดอน 7 จาก 64 โคดอนออกไปโดยสมบูรณ์[ 31 ]
คุณสมบัติ

กรอบอ่านหนังสือ
กรอบการอ่านถูกกำหนดโดยกลุ่มนิวคลีโอไทด์สามตัวแรกซึ่งเป็นจุดเริ่มต้นของการแปลรหัส กรอบนี้จะกำหนดกรอบสำหรับลำดับของโคดอนที่ต่อเนื่องกันและไม่ทับซ้อนกัน ซึ่งเรียกว่า " กรอบการอ่านแบบเปิด " (ORF) ตัวอย่างเช่น ลำดับ 5'-AAATGAACG-3' (ดูรูป) หากอ่านจากตำแหน่งแรก จะมีโคดอน AAA, TGA และ ACG หากอ่านจากตำแหน่งที่สอง จะมีโคดอน AAT และ GAA และหากอ่านจากตำแหน่งที่สาม จะมีโคดอน ATG และ AAC ดังนั้น ทุกลำดับสามารถอ่านได้ในทิศทาง 5' → 3'ในสามกรอบการอ่านซึ่งแต่ละกรอบจะสร้างลำดับกรดอะมิโนที่อาจแตกต่างกันได้ ในตัวอย่างที่ให้มา คือ Lys (K)-Trp (W)-Thr (T), Asn (N)-Glu (E) หรือ Met (M)-Asn (N) ตามลำดับ (เมื่อแปลรหัสด้วยรหัสไมโทคอนเดรียของสัตว์มีกระดูกสันหลัง ) เมื่อ DNA เป็นสายคู่ จะมีการกำหนด เฟรมการอ่าน ที่เป็นไปได้หกเฟรม โดยสามเฟรมอยู่ในทิศทางไปข้างหน้าบนสายหนึ่ง และสามเฟรมอยู่ในทิศทางย้อนกลับบนสายตรงข้าม[ 33 ] : 330 เฟรมการเข้ารหัสโปรตีนถูกกำหนดโดยโคดอนเริ่มต้นซึ่งโดยปกติจะเป็นโคดอน AUG ตัวแรกในลำดับ RNA (ATG ใน DNA)
ในสิ่งมีชีวิตยูคาริโอต กรอบการอ่านแบบเปิดในเอ็กซอนมักถูกขัดจังหวะด้วยอินทรอน
รหัสเริ่มต้นและรหัสหยุด
การแปลเริ่มต้นด้วยโคดอนเริ่มต้นสายโซ่หรือโคดอนเริ่มต้นโคดอนเริ่มต้นเพียงอย่างเดียวไม่เพียงพอที่จะเริ่มต้นกระบวนการ ลำดับใกล้เคียง เช่น ลำดับ Shine-DalgarnoในE. coliและปัจจัยเริ่มต้นก็จำเป็นสำหรับการเริ่มต้นการแปล โคดอนเริ่มต้นที่พบบ่อยที่สุดคือ AUG ซึ่งอ่านเป็นเมไทโอนีนหรือฟอร์มิลเมไทโอนีน (ในแบคทีเรีย ไมโตคอนเดรีย และพลาสติด) โคดอนเริ่มต้นทางเลือกขึ้นอยู่กับสิ่งมีชีวิต ได้แก่ "GUG" หรือ "UUG" โคดอนเหล่านี้โดยปกติจะแทนวาลีนและลิวซีนตามลำดับ แต่ในฐานะโคดอนเริ่มต้น พวกมันจะถูกแปลเป็นเมไทโอนีนหรือฟอร์มิลเมไทโอนีน[ 34 ]
โคดอนหยุดทั้งสามมีชื่อเรียก: UAG คือamber , UGA คือopal (บางครั้งก็เรียกว่าumber ) และ UAA คือochreโคดอนหยุดยังเรียกว่า "โคดอนยุติ" หรือ "โคดอนไร้ความหมาย" พวกมันส่งสัญญาณให้โพลีเปปไทด์ที่เกิดขึ้นใหม่หลุดออกจากไรโบโซม เนื่องจากไม่มี tRNA ที่เกี่ยวข้องที่มีแอนติโคดอนที่เสริมกับสัญญาณหยุดเหล่านี้ ทำให้ปัจจัยปลดปล่อยสามารถจับกับไรโบโซมได้แทน[ 35 ]
ผลกระทบของการกลายพันธุ์
ในระหว่างกระบวนการจำลองดีเอ็นเอข้อผิดพลาดอาจเกิดขึ้นเป็นครั้งคราวในการสร้างพอลิเมอร์ของสายที่สอง ข้อผิดพลาดเหล่านี้ ซึ่งก็คือการกลายพันธุ์ สามารถส่งผลต่อ ฟีโนไทป์ของสิ่งมีชีวิตโดยเฉพาะอย่างยิ่งหากเกิดขึ้นภายในลำดับการเข้ารหัสโปรตีนของยีน อัตราข้อผิดพลาดโดยทั่วไปคือ 1 ข้อผิดพลาดในทุกๆ 10–100 ล้านเบส เนื่องมาจากความสามารถในการ " ตรวจสอบความถูกต้อง " ของดีเอ็นเอพอลิเมอเรส[ 37 ] [ 38 ]
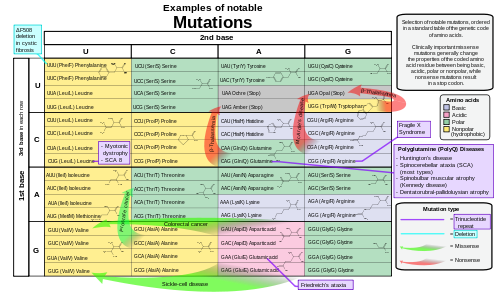
การกลายพันธุ์แบบมิสเซนส์และการกลายพันธุ์แบบนันเซนส์เป็นตัวอย่างของการกลายพันธุ์แบบจุดที่สามารถทำให้เกิดโรคทางพันธุกรรม เช่นโรคโลหิตจางเคียวและธาลัสซีเมียตามลำดับ[ 39 ] [ 40 ]การกลายพันธุ์แบบมิสเซนส์ที่มีความสำคัญทางคลินิกโดยทั่วไปจะเปลี่ยนคุณสมบัติของกรดอะมิโนที่เข้ารหัสระหว่างสถานะพื้นฐาน กรด ขั้ว หรือไม่มีขั้ว ในขณะที่การกลายพันธุ์แบบนันเซนส์ส่งผลให้เกิดรหัสหยุด[ 33 ]
การกลายพันธุ์ที่รบกวนลำดับเฟรมการอ่านโดยการแทรกหรือลบ เบสนิวคลี โอไทด์ที่ไม่ใช่จำนวนเท่าของ 3 เรียกว่าการกลายพันธุ์แบบเฟรมชิฟต์การกลายพันธุ์เหล่านี้มักส่งผลให้เกิดการแปลที่แตกต่างไปจากเดิมอย่างสิ้นเชิง และมีแนวโน้มที่จะทำให้เกิด การอ่าน รหัสหยุดซึ่งทำให้โปรตีนสั้นลง[ 41 ]การกลายพันธุ์เหล่านี้อาจทำให้การทำงานของโปรตีนบกพร่อง ดังนั้นจึงพบได้น้อยใน ลำดับการเข้ารหัสโปรตีน ในร่างกายเหตุผลหนึ่งที่การถ่ายทอดการกลายพันธุ์แบบเฟรมชิฟต์นั้นหายากก็คือ หากโปรตีนที่ถูกแปลนั้นมีความสำคัญต่อการเจริญเติบโตภายใต้แรงกดดันในการคัดเลือกที่สิ่งมีชีวิตเผชิญ การขาดโปรตีนที่ทำงานได้อาจทำให้สิ่งมีชีวิตตายก่อนที่มันจะสามารถอยู่รอดได้[ 42 ] การกลายพันธุ์แบบเฟรมชิฟต์ อาจส่งผลให้เกิดโรคทางพันธุกรรมที่รุนแรง เช่นโรคเทย์-แซคส์[ 43 ]
แม้ว่าการกลายพันธุ์ส่วนใหญ่ที่เปลี่ยนแปลงลำดับโปรตีนจะเป็นอันตรายหรือไม่มีผลใดๆ แต่การกลายพันธุ์บางอย่างก็มีประโยชน์[ 44 ]การกลายพันธุ์เหล่านี้อาจทำให้สิ่งมีชีวิตกลายพันธุ์สามารถทนต่อความเครียดจากสิ่งแวดล้อมบางอย่างได้ดีกว่า สิ่งมีชีวิต สายพันธุ์ปกติหรือสามารถสืบพันธุ์ได้เร็วขึ้น ในกรณีเหล่านี้ การกลายพันธุ์มักจะแพร่หลายมากขึ้นในประชากรผ่านการคัดเลือกโดยธรรมชาติ [ 45 ] ไวรัสที่ใช้RNAเป็นสารพันธุกรรมมีอัตราการกลายพันธุ์ที่รวดเร็ว[ 46 ]ซึ่งอาจเป็นข้อได้เปรียบ เนื่องจากไวรัสเหล่านี้จึงวิวัฒนาการได้อย่างรวดเร็ว และหลีกเลี่ยงการตอบสนองการป้องกัน ของ ระบบภูมิคุ้มกัน[ 47 ]ในประชากรขนาดใหญ่ของสิ่งมีชีวิตที่สืบพันธุ์แบบไม่อาศัยเพศ เช่นE. coliการกลายพันธุ์ที่เป็นประโยชน์หลายอย่างอาจเกิดขึ้นพร้อมกัน ปรากฏการณ์นี้เรียกว่าการรบกวนของโคลนและทำให้เกิดการแข่งขันระหว่างการกลายพันธุ์[ 48 ]
ความเสื่อมโทรม

{kind=link}

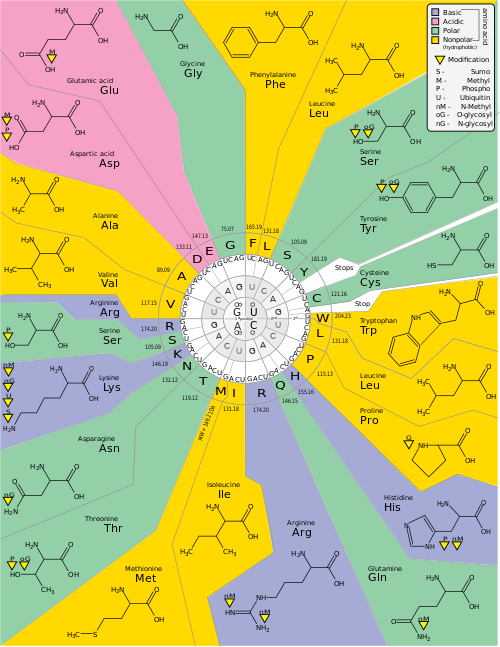
ความซ้ำซ้อน (Degeneracy) คือความซ้ำซ้อนของรหัสพันธุกรรม คำนี้ตั้งโดย Bernfield และ Nirenberg รหัสพันธุกรรมมีความซ้ำซ้อนแต่ไม่มีความกำกวม (ดูตารางโคดอนด้านล่างสำหรับความสัมพันธ์ทั้งหมด) ตัวอย่างเช่น แม้ว่าโคดอน GAA และ GAG ต่างก็ระบุถึงกรดกลูตามิก (ความซ้ำซ้อน) แต่ก็ไม่มีโคดอนใดระบุถึงกรดอะมิโนอื่น (ไม่มีความกำกวม) โคดอนที่เข้ารหัสกรดอะมิโนหนึ่งตัวอาจแตกต่างกันในตำแหน่งใดตำแหน่งหนึ่งในสามตำแหน่ง ตัวอย่างเช่น กรดอะมิโนลิวซีนระบุโดย โคดอน Y U Rหรือ CU N (UUA, UUG, CUU, CUC, CUA หรือ CUG) (ความแตกต่างในตำแหน่งที่หนึ่งหรือที่สามระบุโดยใช้สัญกรณ์ IUPAC ) ในขณะที่กรดอะมิโนซีรีนระบุโดยโคดอน UC Nหรือ AG Y (UCA, UCG, UCC, UCU, AGU หรือ AGC) (ความแตกต่างในตำแหน่งที่หนึ่ง สอง หรือสาม) [ 49 ]ผลที่ตามมาในทางปฏิบัติของความซ้ำซ้อนคือ ข้อผิดพลาดในตำแหน่งที่สามของรหัสสามตัวจะทำให้เกิดการกลายพันธุ์แบบเงียบหรือข้อผิดพลาดที่ไม่ส่งผลกระทบต่อโปรตีน เนื่องจากคุณสมบัติชอบน้ำหรือไม่ชอบน้ำยังคงอยู่โดยการแทนที่กรดอะมิโนที่เทียบเท่ากัน ตัวอย่างเช่น รหัส NUN (โดยที่ N = นิวคลีโอไทด์ใดๆ) มักจะเข้ารหัสกรดอะมิโนที่ไม่ชอบน้ำ NCN ให้กรดอะมิโนที่มีขนาดเล็กและมีคุณสมบัติชอบน้ำปานกลาง NAN เข้ารหัสกรดอะมิโนที่ชอบน้ำขนาดเฉลี่ย รหัสพันธุกรรมมีโครงสร้างที่ดีมากสำหรับคุณสมบัติชอบน้ำ จนกระทั่งการวิเคราะห์ทางคณิตศาสตร์ ( การแยกส่วนค่าเอกพจน์ ) ของตัวแปร 12 ตัว (นิวคลีโอไทด์ 4 ตัว x 3 ตำแหน่ง) ให้ความสัมพันธ์ที่น่าทึ่ง (C = 0.95) สำหรับการทำนายคุณสมบัติชอบน้ำของกรดอะมิโนที่เข้ารหัสโดยตรงจากลำดับนิวคลีโอไทด์สามตัวโดยไม่ต้องแปล[ 50 ] [ 51 ] โปรดสังเกตในตารางด้านล่าง กรดอะมิโนแปดชนิดไม่ได้รับผลกระทบจากการกลายพันธุ์ที่ตำแหน่งที่สามของโคดอนเลย ในขณะที่ในรูปด้านบน การกลายพันธุ์ที่ตำแหน่งที่สองมีแนวโน้มที่จะทำให้เกิดการเปลี่ยนแปลงอย่างมากในคุณสมบัติทางกายภาพและเคมีของกรดอะมิโนที่เข้ารหัส อย่างไรก็ตาม การเปลี่ยนแปลงที่ตำแหน่งแรกของโคดอนมีความสำคัญมากกว่าการเปลี่ยนแปลงที่ตำแหน่งที่สองในภาพรวม[ 52 ] เหตุผลอาจเป็นเพราะการกลับประจุ (จากประจุบวกเป็นประจุลบหรือในทางกลับกัน) สามารถเกิดขึ้นได้เฉพาะกับการกลายพันธุ์ที่ตำแหน่งแรกของโคดอนบางชนิดเท่านั้น แต่ไม่เกิดขึ้นกับการเปลี่ยนแปลงที่ตำแหน่งที่สองของโคดอนใดๆ การกลับประจุดังกล่าวอาจมีผลกระทบอย่างมากต่อโครงสร้างหรือการทำงานของโปรตีน แง่มุมนี้อาจถูกประเมินต่ำเกินไปโดยการศึกษาก่อนหน้านี้[ 52 ]
ความลำเอียงในการใช้โคดอน
ความถี่ของโคดอน หรือที่รู้จักกันในชื่อความลำเอียงในการใช้โคดอนสามารถแตกต่างกันไปในแต่ละสปีชีส์ ซึ่งมีผลต่อการควบคุมการแปลโคดอนจะแตกต่างกันไปตามสิ่งมีชีวิต ตัวอย่างเช่น โคดอนโพรลีนที่พบมากที่สุดใน E. coli คือ CCG ในขณะที่ในมนุษย์ โคดอนโพรลีนนี้เป็นโคดอนที่ใช้น้อยที่สุด[ 53 ]
ตารางความถี่โคดอนจีโนมมนุษย์[ 54 ] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
รหัสพันธุกรรมทางเลือก
กรดอะมิโนที่ไม่เป็นมาตรฐาน
ในโปรตีนบางชนิด กรดอะมิโนที่ไม่เป็นมาตรฐานจะถูกแทนที่ด้วยรหัสหยุดมาตรฐาน ขึ้นอยู่กับลำดับสัญญาณที่เกี่ยวข้องในอาร์เอ็นเอส่งสาร ตัวอย่างเช่น UGA สามารถเข้ารหัสซีลีโนซิสเทอีนและ UAG สามารถเข้ารหัสไพ โร ไลซีน ซีลีโนซิสเทอีนจึงถูกมองว่าเป็นกรดอะมิโนตัวที่ 21 และไพโรไลซีนเป็นตัวที่ 22 [ 55 ]ทั้งซีลีโนซิสเทอีนและไพโรไลซีนอาจมีอยู่ในสิ่งมีชีวิตเดียวกัน[ 55 ]แม้ว่ารหัสพันธุกรรมโดยปกติจะคงที่ในสิ่งมีชีวิต แต่โปรคาริโอตอะคีอัลAcetohalobium arabaticumสามารถขยายรหัสพันธุกรรมจาก 20 เป็น 21 กรดอะมิโน (โดยรวมไพโรไลซีน) ภายใต้สภาวะการเจริญเติบโตที่แตกต่างกัน[ 56 ]
การเปลี่ยนแปลง
เดิมทีมีข้อโต้แย้งที่เรียบง่ายและเป็นที่ยอมรับกันอย่างกว้างขวางว่ารหัสพันธุกรรมควรเป็นสากล กล่าวคือ การเปลี่ยนแปลงใดๆ ในรหัสพันธุกรรมจะเป็นอันตรายถึงชีวิตต่อสิ่งมีชีวิต (แม้ว่า Crick จะระบุว่าไวรัสเป็นข้อยกเว้น) นี่เป็นที่รู้จักกันในชื่อข้อโต้แย้ง "อุบัติเหตุแช่แข็ง" สำหรับความเป็นสากลของรหัสพันธุกรรม อย่างไรก็ตาม ในบทความสำคัญของเขาเกี่ยวกับต้นกำเนิดของรหัสพันธุกรรมในปี 1968 ฟรานซิส คริก ยังคงระบุว่าความเป็นสากลของรหัสพันธุกรรมในสิ่งมีชีวิตทั้งหมดเป็นสมมติฐานที่ยังไม่ได้รับการพิสูจน์ และอาจไม่เป็นความจริงในบางกรณี เขาทำนายว่า "รหัสเป็นสากล (เหมือนกันในสิ่งมีชีวิตทั้งหมด) หรือเกือบจะเป็นเช่นนั้น" [ 58 ]การเปลี่ยนแปลงครั้งแรกถูกค้นพบในปี 1979 โดยนักวิจัยที่ศึกษาเกี่ยวกับ ยีนไมโทคอนเดรีย ของมนุษย์[ 59 ]หลังจากนั้นก็มีการค้นพบรูปแบบที่แตกต่างกันเล็กน้อยมากมาย[ 60 ]รวมถึงรหัสไมโทคอนเดรียทางเลือกต่างๆ[ 61 ]ตัวอย่างเช่น ตัวแปรย่อยเหล่านี้เกี่ยวข้องกับการแปลรหัสพันธุกรรม UGA เป็นทริปโตแฟนใน สายพันธุ์ ไมโคพลาสมาและการแปล CUG เป็นซีรีนแทนที่จะเป็นลิวซีนในยีสต์ของ "กลุ่ม CTG" (เช่นCandida albicans ) [ 62 ] [ 63 ] [ 64 ]เนื่องจากไวรัสต้องใช้รหัสพันธุกรรมเดียวกันกับโฮสต์ การดัดแปลงรหัสพันธุกรรมมาตรฐานอาจรบกวนการสังเคราะห์โปรตีนหรือการทำงานของไวรัส อย่างไรก็ตาม ไวรัสเช่นโทติไวรัสได้ปรับตัวให้เข้ากับการดัดแปลงรหัสพันธุกรรมของโฮสต์ แล้ว [ 65 ]ในแบคทีเรียและอาร์เคีย GUG และ UUG เป็นรหัสเริ่มต้นทั่วไป ในบางกรณีที่หายาก โปรตีนบางชนิดอาจใช้รหัสเริ่มต้นทางเลือกอื่น[ 60 ] น่าประหลาดใจที่ความแปรผันในการตีความรหัสพันธุกรรมยังมีอยู่ในยีนที่เข้ารหัสในนิวเคลียสของมนุษย์ด้วย: ในปี 2016 นักวิจัยที่ศึกษาการแปลของมาเลตดีไฮโดรจีเนสพบว่าใน mRNA ประมาณ 4% ที่เข้ารหัสเอนไซม์นี้ โคดอนหยุดถูกใช้ตามธรรมชาติเพื่อเข้ารหัสกรดอะมิโนทริปโตเฟนและอาร์จินีน[ 66 ]การเข้ารหัสใหม่ประเภทนี้ถูกกระตุ้นโดยบริบทโคดอนหยุดที่มีการอ่านผ่านสูง[ 67 ]และเรียกว่า การอ่านผ่านการ แปลเชิงฟังก์ชัน[ 68 ]
แม้จะมีความแตกต่างกัน แต่รหัสที่เกิดขึ้นตามธรรมชาติที่รู้จักทั้งหมดก็มีความคล้ายคลึงกันมาก กลไกการเข้ารหัสเหมือนกันสำหรับสิ่งมีชีวิตทั้งหมด: โคดอนสามเบส, tRNA , ไรโบโซม, การอ่านทิศทางเดียว และการแปลโคดอนเดี่ยวเป็นกรดอะมิโนเดี่ยว[ 69 ]ความแปรผันที่รุนแรงที่สุดเกิดขึ้นในซีลิเอตบางชนิด ซึ่งความหมายของโคดอนหยุดขึ้นอยู่กับตำแหน่งภายใน mRNA เมื่ออยู่ใกล้ปลาย 3' โคดอนหยุดจะทำหน้าที่เป็นตัวยุติ ในขณะที่อยู่ในตำแหน่งภายใน โคดอนหยุดจะเข้ารหัสกรดอะมิโน เช่นในCondylostoma magnum [ 70 ] หรือกระตุ้นการเลื่อนเฟรมของไรโบโซมเช่นในEuplotes [ 71 ]
ที่มาและความแปรผันของรหัสพันธุกรรม รวมถึงกลไกเบื้องหลังความสามารถในการวิวัฒนาการของรหัสพันธุกรรม ได้รับการศึกษาอย่างกว้างขวาง[ 72 ] [ 73 ]และมีการศึกษาบางส่วนที่ดำเนินการทดลองวิวัฒนาการรหัสพันธุกรรมของสิ่งมีชีวิตบางชนิด[ 74 ] [ 75 ] [ 76 ]
การอนุมาน
รหัสพันธุกรรมที่แตกต่างกันซึ่งสิ่งมีชีวิตใช้สามารถอนุมานได้โดยการระบุยีนที่มีการอนุรักษ์สูงซึ่งเข้ารหัสอยู่ในจีโนมนั้น และเปรียบเทียบการใช้โคดอนกับกรดอะมิโนในโปรตีนที่คล้ายคลึงกันของสิ่งมีชีวิตอื่น ตัวอย่างเช่น โปรแกรม FACIL อนุมานรหัสพันธุกรรมโดยการค้นหากรดอะมิโนในโดเมนโปรตีนที่คล้ายคลึงกันที่มักจะเรียงตัวกับโคดอนแต่ละตัวมากที่สุด ความน่าจะเป็นของกรดอะมิโน (หรือโคดอนหยุด) ที่ได้สำหรับแต่ละโคดอนจะแสดงในโลโก้รหัสพันธุกรรม[ 57 ]
ณ เดือนมกราคม 2022 การสำรวจรหัสพันธุกรรมที่สมบูรณ์ที่สุดดำเนินการโดย Shulgina และ Eddy ซึ่งคัดกรองจีโนมโปรคาริโอต 250,000 จีโนมโดยใช้เครื่องมือ Codetta ของพวกเขา เครื่องมือนี้ใช้วิธีการที่คล้ายกับ FACIL โดยใช้ ฐานข้อมูล Pfam ที่ใหญ่กว่า แม้ว่า NCBI จะมีตารางการแปล 27 ตารางแล้วก็ตาม ผู้เขียนก็สามารถค้นพบความแปรผันของรหัสพันธุกรรมใหม่ 5 รายการ (ได้รับการยืนยันโดยการกลายพันธุ์ของ tRNA) และแก้ไขการระบุที่ไม่ถูกต้องหลายรายการ[ 77 ]ต่อมา Codetta ถูกนำมาใช้ในการวิเคราะห์การเปลี่ยนแปลงรหัสพันธุกรรมในซีลิเอต[ 78 ]
ต้นทาง
รหัสพันธุกรรมเป็นส่วนสำคัญของประวัติศาสตร์ของสิ่งมีชีวิตภายใต้สมมติฐานโลกของ RNAโมเลกุล RNA ที่จำลองตัวเองได้นั้นมาก่อนการใช้โปรตีนอย่างมีนัยสำคัญ ภายใต้สมมติฐานโลกของนิวคลีโอเปปไทด์ การใช้เปปไทด์อย่างมีนัยสำคัญนั้นมาก่อนรหัสพันธุกรรมและเกิดขึ้นพร้อมกับการใช้ RNA ที่ซับซ้อนของสิ่งมีชีวิตยุคแรก[ 79 ]โมเลกุล transfer RNA ดูเหมือนจะวิวัฒนาการมาก่อนaminoacyl-tRNA synthetasesใน ปัจจุบัน [ 80 ]เป็นไปได้ว่า synthetases เข้ามาแทนที่ระบบribozymes (เอนไซม์ RNA) ในยุคก่อนหน้า หรือว่ากรดอะมิโนได้รับการจดจำโดยช่องเฉพาะในโครงสร้างตติยภูมิของ proto-tRNAs [ 81 ]ยังไม่ทราบว่าเหตุใดรหัสพันธุกรรมจึงใช้เฉพาะกรดอะมิโน L และไม่ใช้กรดอะมิโน D [ 82 ]
แบบจำลองวิวัฒนาการใดๆ สำหรับต้นกำเนิดของรหัสจะต้องคำนึงถึงความทนทานของโปรตีนที่เข้ารหัสต่อข้อผิดพลาดระหว่างการจำลองดีเอ็นเอและระหว่างการแปล ข้อผิดพลาดของนิวคลีโอไทด์เดี่ยวจำนวนมากเป็นข้อผิดพลาดแบบเดียวกันและข้อผิดพลาดที่ไม่ใช่แบบนั้นมักจะทำให้เกิดการแทนที่กรดอะมิโนที่คล้ายคลึงกันทางชีวเคมีแม้จะรักษาสภาพโครงสร้างของรหัสให้เหมือนเดิมโดยที่กลุ่มของโคดอนเข้ารหัสกรดอะมิโนตัวเดียวกัน แต่กรดอะมิโนใดถูกเข้ารหัสโดยชุดของโคดอนใดนั้นมีโอกาส "หนึ่งในล้าน" ในแง่ของความทนทาน[ 83 ]กรดอะมิโนที่คล้ายคลึงกันทางชีวเคมีมักจะมีนิวคลีโอไทด์ตรงกลางเหมือนกัน ในขณะที่การเปลี่ยนแปลงแบบเดียวกันโดยทั่วไปจะเกิดขึ้นที่นิวคลีโอไทด์ตัวที่สาม
กรดอะมิโนที่มีวิถีการสังเคราะห์ทางชีวภาพเดียวกันมักจะมีเบสแรกในโคดอนเหมือนกัน นี่อาจเป็นร่องรอยทางวิวัฒนาการของรหัสพันธุกรรมยุคแรกที่เรียบง่ายกว่าซึ่งมีกรดอะมิโนน้อยกว่า ซึ่งต่อมาได้วิวัฒนาการเพื่อสร้างรหัสกรดอะมิโนที่ใหญ่ขึ้น[ 84 ] นอกจากนี้ ยังอาจสะท้อนถึงคุณสมบัติทางกายภาพและทางเคมีที่มีผลต่อโคดอนในระหว่างวิวัฒนาการ กรดอะมิโนที่มีคุณสมบัติทางกายภาพคล้ายกันมักจะมีโคดอนที่คล้ายกันด้วย[ 85 ] [ 86 ]ซึ่งช่วยลดปัญหาที่เกิดจากการกลายพันธุ์แบบจุดและการแปลผิด[ 83 ]
สมมติฐานหลักสามประการกล่าวถึงที่มาของรหัสพันธุกรรม แบบจำลองจำนวนมากเป็นของสมมติฐานใดสมมติฐานหนึ่งหรือเป็นลูกผสม: [ 87 ]
- การแช่แข็งแบบสุ่ม: รหัสพันธุกรรมถูกสร้างขึ้นแบบสุ่ม ตัวอย่างเช่น ไรโบไซม์ที่คล้ายกับ tRNA ในยุคแรก อาจมีความสัมพันธ์กับกรดอะมิโนที่แตกต่างกัน โดยมีโคดอนเกิดขึ้นจากส่วนอื่นของไรโบไซม์ที่แสดงความแปรปรวนแบบสุ่ม เมื่อ มีการเข้ารหัส เปปไทด์ มากพอ การเปลี่ยนแปลงแบบสุ่มครั้งใหญ่ใดๆ ในรหัสพันธุกรรมจะเป็นอันตรายถึงชีวิต ดังนั้นรหัสพันธุกรรมจึง "หยุดนิ่ง" [ 58 ]
- ความสัมพันธ์เชิงสเตอริโอเคมี: รหัสพันธุกรรมเป็นผลมาจากความสัมพันธ์ที่สูงระหว่างกรดอะมิโนแต่ละตัวกับโคดอนหรือแอนติโคดอน ตัวเลือกหลังนี้หมายความว่าโมเลกุล pre-tRNA จับคู่กับกรดอะมิโนที่สอดคล้องกันโดยความสัมพันธ์นี้ ต่อมาในระหว่างวิวัฒนาการ การจับคู่นี้ค่อยๆ ถูกแทนที่ด้วยการจับคู่โดยอะมิโนเอซิล-tRNA ซินเทส[ 82 ] [ 88 ] [ 89 ]
- ความเหมาะสมที่สุด: รหัสพันธุกรรมยังคงวิวัฒนาการต่อไปหลังจากการสร้างครั้งแรก เพื่อให้รหัสปัจจุบันเพิ่ม ฟังก์ชัน ความเหมาะสม สูงสุด ซึ่งโดยปกติแล้วจะเป็นการลดข้อผิดพลาดบางประเภท[ 82 ] [ 87 ] [ 90 ]
สมมติฐานได้กล่าวถึงสถานการณ์ต่างๆ ไว้มากมาย: [ 91 ]
- หลักการทางเคมีควบคุมปฏิสัมพันธ์เฉพาะของ RNA กับกรดอะมิโน การทดลองกับแอพทาเมอร์แสดงให้เห็นว่ากรดอะมิโนบางชนิดมีความสัมพันธ์ทางเคมีแบบเลือกเฉพาะกับโคดอนของพวกมัน[ 92 ]การทดลองแสดงให้เห็นว่าจากกรดอะมิโน 8 ชนิดที่ทดสอบ มี 6 ชนิดที่แสดงการเชื่อมโยงระหว่าง RNA สามตัวกับกรดอะมิโน[ 93 ] [ 89 ]
- การขยายตัวทางชีวสังเคราะห์ รหัสพันธุกรรมเติบโตจากรหัสที่เรียบง่ายกว่าในอดีตผ่านกระบวนการ "การขยายตัวทางชีวสังเคราะห์" สิ่งมีชีวิตดั้งเดิม "ค้นพบ" กรดอะมิโนใหม่ (ตัวอย่างเช่น เป็นผลพลอยได้จากกระบวนการเผาผลาญ ) และต่อมาได้รวมกรดอะมิโนเหล่านี้บางส่วนเข้ากับกลไกการเข้ารหัสทางพันธุกรรม[ 73 ]แม้ว่าจะมีหลักฐานแวดล้อมมากมายที่บ่งชี้ว่ามีการใช้กรดอะมิโนประเภทน้อยกว่าในอดีต[ 94 ] แต่ สมมติฐานที่แม่นยำและละเอียดเกี่ยวกับกรดอะมิโนใดที่เข้าสู่รหัสในลำดับใดนั้นยังคงเป็นที่ถกเถียงกันอยู่[ 95 ] [ 96 ]อย่างไรก็ตาม การศึกษาหลายชิ้นได้แนะนำว่า Gly, Ala, Asp, Val, Ser, Pro, Glu, Leu, Thr อาจอยู่ในกลุ่มของกรดอะมิโนที่เพิ่มเข้ามาในช่วงต้น ในขณะที่ Cys, Met, Tyr, Trp, His, Phe อาจอยู่ในกลุ่มของกรดอะมิโนที่เพิ่มเข้ามาในภายหลัง[ 97 ] [ 98 ] [ 99 ] [ 100 ]การวิเคราะห์ทางเลือกเกี่ยวกับการใช้กรดอะมิโนในบรรพบุรุษร่วมสากลสุดท้ายสรุปได้ว่ากรดอะมิโนมาในลำดับต่อไปนี้: Val, Gly, Ile, Met, Ala, Thr, His, Glu, Cys, Pro, Lys, Ser, Asp, Leu, Asn, Arg, Phe, Tyr, Gln, Trp [ 101 ]มีการชี้ให้เห็นว่าการปรากฏตัวของซิสเทอีนและเมไทโอนีนที่มีกำมะถันในภายหลังนั้นสรุปได้บางส่วนจากการที่ไม่มีการทดลอง Miller–Ureyที่ปราศจากกำมะถัน เชื่อกันว่าสิ่งมีชีวิตในยุคแรกใช้S-adenosyl methionineและในขณะที่ฮิสติดีนสร้างได้ยากในทางอชีวภาพ แต่สามารถสังเคราะห์ได้ง่ายในสิ่งมีชีวิตที่มี RNA ที่ซับซ้อนอยู่แล้วและมีการสังเคราะห์พิวรีน[ 101 ]
- การคัดเลือกโดยธรรมชาตินำไปสู่การกำหนดโคดอนของรหัสพันธุกรรมที่ลดผลกระทบของการกลายพันธุ์ให้ น้อยที่สุด [ 102 ]สมมติฐานล่าสุด[ 103 ]ชี้ให้เห็นว่ารหัสสามตัวถูกพัฒนามาจากรหัสที่ใช้โคดอนที่ยาวกว่าสามตัว (เช่น โคดอนสี่ตัว) การถอดรหัสที่ยาวกว่าสามตัวจะเพิ่มความซ้ำซ้อนของโคดอนและจะทนต่อข้อผิดพลาดได้มากขึ้น คุณลักษณะนี้อาจช่วยให้การถอดรหัสแม่นยำโดยไม่ต้องใช้กลไกการแปลที่ซับซ้อน เช่นไรโบโซมเช่น ก่อนที่เซลล์จะเริ่มสร้างไรโบโซม
- ช่องทางข้อมูล: แนวทาง ทฤษฎีข้อมูลจำลองกระบวนการแปลรหัสพันธุกรรมเป็นกรดอะมิโนที่สอดคล้องกันเป็นช่องทางข้อมูลที่มีข้อผิดพลาด[ 104 ]สัญญาณรบกวนโดยธรรมชาติ (นั่นคือ ข้อผิดพลาด) ในช่องทางทำให้สิ่งมีชีวิตต้องเผชิญกับคำถามพื้นฐาน: จะสร้างรหัสพันธุกรรมให้ทนต่อสัญญาณรบกวนได้อย่างไร[ 105 ]ในขณะที่แปลข้อมูลได้อย่างแม่นยำและมีประสิทธิภาพ? แบบจำลอง"อัตรา-การบิดเบือน" เหล่านี้ [ 106 ]ชี้ให้เห็นว่ารหัสพันธุกรรมมีต้นกำเนิดมาจากการทำงานร่วมกันของแรงผลักดันวิวัฒนาการที่ขัดแย้งกันสามประการ ได้แก่ ความต้องการกรดอะมิโนที่หลากหลาย[ 107 ]ความทนทานต่อข้อผิดพลาด[ 102 ]และต้นทุนทรัพยากรที่น้อยที่สุด รหัสเกิดขึ้นในช่วงเปลี่ยนผ่านเมื่อการจับคู่โคดอนกับกรดอะมิโนไม่เป็นแบบสุ่ม การเกิดขึ้นของรหัสถูกควบคุมโดยโทโพโลยีที่กำหนดโดยข้อผิดพลาดที่อาจเกิดขึ้นและเกี่ยวข้องกับปัญหาการระบายสีแผนที่[ 108 ]
- ทฤษฎีเกม: แบบจำลองที่อิงตามเกมการส่งสัญญาณรวมองค์ประกอบของทฤษฎีเกม การคัดเลือกโดยธรรมชาติ และช่องทางข้อมูล แบบจำลองดังกล่าวถูกนำมาใช้เพื่อเสนอแนะว่าโพลีเปปไทด์แรกน่าจะมีขนาดสั้นและมีฟังก์ชันที่ไม่ใช่เอนไซม์ แบบจำลองทฤษฎีเกมชี้ให้เห็นว่าการจัดระเบียบสาย RNA ให้เป็นเซลล์อาจมีความจำเป็นเพื่อป้องกันการใช้รหัสพันธุกรรมในทางที่ "หลอกลวง" กล่าวคือ การป้องกันสิ่งที่เทียบเท่ากับไวรัสในยุคโบราณจากการครอบงำโลก RNA [ 109 ]
- โคดอนหยุด: โคดอนสำหรับการหยุดการแปลเป็นอีกแง่มุมที่น่าสนใจของปัญหาต้นกำเนิดของรหัสพันธุกรรม ตัวอย่างเช่น ในการกล่าวถึงวิวัฒนาการของโคดอนหยุด มีการเสนอแนะว่าโคดอนหยุดมีแนวโน้มที่จะยุติการแปลก่อนกำหนดในกรณีที่เกิดข้อผิดพลาดการเลื่อนเฟรม[ 110 ]ในทางตรงกันข้าม แบบจำลองโมเลกุลเชิงสเตอริโอเคมีบางแบบอธิบายต้นกำเนิดของโคดอนหยุดว่า "ไม่สามารถระบุได้" [ 82 ]
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gilbert WM (1999). บทนำสู่การวิเคราะห์ทางพันธุกรรม (ฉบับที่ 7). ซานฟรานซิสโก: WH Freeman. ISBN 978-0-7167-3771-1เก็บถาวรจากต้นฉบับเมื่อวันที่ 24 ธันวาคม 2544
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). ชีววิทยาโมเลกุลของเซลล์ (ฉบับที่ 4). นิวยอร์ก: Garland Science. ISBN 978-0-8153-3218-3.
- โลดิช HF, เบิร์ก เอ, ซิเพอร์สกี้ เอสแอล, มัตสึไดระ พี, บัลติมอร์ ดี, ดาร์เนล เจอี (2000) อณูเซลล์ชีววิทยา (ฉบับที่ 4) ซานฟรานซิสโก: WH ฟรีแมนไอเอสบีเอ็น 978-0-7167-3706-3.
- Caskey CT, Leder P (เมษายน 2014). "รหัส RNA: ศิลาโรเซตตาแห่งธรรมชาติ" . Proceedings of the National Academy of Sciences of the United States of America . 111 (16): 5758– 5759. Bibcode : 2014PNAS..111.5758C . doi : 10.1073/pnas.1404819111 . PMC 4000803 . PMID 24756939 .
ลิงก์ภายนอก
- รหัสพันธุกรรม: ตารางรหัสพันธุกรรม
- ฐานข้อมูลการใช้โคดอน — ตารางความถี่ของโคดอนสำหรับสิ่งมีชีวิตหลายชนิด
- ประวัติการถอดรหัสพันธุกรรม เก็บถาวรเมื่อวันที่ 21 ตุลาคม 2550 ที่Wayback Machine
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ รหัสพันธุกรรม
รหัสพันธุกรรม คือชุดของกฎที่ เซลล์สิ่ง มีชีวิตใช้ ใน การแปล ข้อมูลที่เข้ารหัสอยู่ในสารพันธุกรรม ( ลำดับ ดีเอ็นเอ หรือ อาร์เอ็นเอ ของกลุ่มนิวคลีโอไทด์สามตัวหรือ โคดอน ) ไปเป็น...
ประวัติศาสตร์
ความพยายามที่จะเข้าใจว่าโปรตีนถูกเข้ารหัสอย่างไร เริ่มขึ้นหลังจาก มีการค้นพบ โครงสร้างของ DNA ในปี 1953 ผู้ค้นพบที่สำคัญคือ ฟรานซิส คริก นักชีวฟิสิกส์ชาวอังกฤษ และ เจมส์ วัตสัน นักชีววิทยาชาวอเมริกัน ซึ่งทำงานร่วมกันที่ ห้องปฏิบัติการคาเวนดิช...
โคดอน
การทดลอง ของ Crick, Brenner, Barnett และ Watts-Tobin เป็นครั้งแรกที่แสดงให้เห็นว่า โคดอน ประกอบด้วยเบส DNA สามชนิด
รหัสพันธุกรรมที่ขยายใหญ่ขึ้น (ชีววิทยาเชิงสังเคราะห์)
ในแวดวงวิชาการโดยทั่วไป แนวคิดเรื่องวิวัฒนาการของรหัสพันธุกรรมจากรหัสพันธุกรรมดั้งเดิมที่ไม่ชัดเจนไปสู่รหัสที่กำหนดไว้อย่างชัดเจน ("แช่แข็ง") พร้อมด้วยกรดอะมิโนมาตรฐาน 20 (+2) ชนิด ได้รับการยอมรับอย่างกว้างขวาง [ 20 ] อย่างไรก็ตาม มีความคิดเห็น แนวคิด วิธีการ...