อ่าน 9 นาที
การเชื่อมโยงทางพันธุกรรม
การเชื่อมโยงทางพันธุกรรมคือแนวโน้มของลำดับดีเอ็นเอที่อยู่ใกล้กันบนโครโมโซมที่จะถูกถ่ายทอดไปด้วยกันในระหว่าง ระยะ
การเชื่อมโยงทางพันธุกรรม
การเชื่อมโยงทางพันธุกรรมคือแนวโน้มของลำดับดีเอ็นเอที่อยู่ใกล้กันบนโครโมโซมที่จะถูกถ่ายทอดไปด้วยกันในระหว่าง ระยะ ไมโอซิสของการสืบพันธุ์แบบอาศัยเพศเครื่องหมายทางพันธุกรรมสองตัวที่อยู่ใกล้กันทางกายภาพมีโอกาสน้อยที่จะแยกออกจากกันไปยังโครมาทิด ที่แตกต่างกัน ในระหว่างการไขว้กันของโครโมโซมดังนั้นจึงกล่าวได้ว่ามีการเชื่อมโยงกันมากกว่าเครื่องหมายที่อยู่ห่างกัน กล่าวอีกนัยหนึ่ง ยิ่งยีน สองตัว อยู่ใกล้กันบนโครโมโซมมากเท่าใด โอกาสในการเกิดการรวมตัวใหม่ระหว่างยีนเหล่านั้นก็จะยิ่งน้อยลง และยิ่งมีแนวโน้มที่จะถูกถ่ายทอดไปด้วยกันมากขึ้นเท่านั้น เครื่องหมายบนโครโมโซมที่แตกต่างกันจะไม่เชื่อมโยงกัน อย่างสมบูรณ์ แม้ว่าการแสดงออกของอัลลีลที่อาจเป็นอันตรายอาจได้รับอิทธิพลจากการมีอยู่ของอัลลีลอื่นๆ และอัลลีลอื่นๆ เหล่านี้อาจอยู่บนโครโมโซมอื่นที่ไม่ใช่โครโมโซมที่อัลลีลที่อาจเป็นอันตรายนั้นตั้งอยู่[ 1 ]
การเชื่อมโยงทางพันธุกรรมเป็นข้อยกเว้นที่โดดเด่นที่สุดของกฎการแยกตัวอย่างอิสระของเกรเกอร์ เมนเดลการทดลองครั้งแรกที่แสดงให้เห็นถึงการเชื่อมโยงเกิดขึ้นในปี ค.ศ. 1905 ในเวลานั้น สาเหตุที่ลักษณะบางอย่างมักถูกถ่ายทอดทางพันธุกรรมร่วมกันยังไม่เป็นที่ทราบแน่ชัด งานวิจัยในภายหลังได้เปิดเผยว่ายีนเป็นโครงสร้างทางกายภาพที่สัมพันธ์กันด้วยระยะทางทางกายภาพ
หน่วยวัดการเชื่อมโยงทางพันธุกรรมโดยทั่วไปคือเซนติมอร์แกน (cM) ระยะห่าง 1 cM ระหว่างเครื่องหมายสองตัวหมายความว่าเครื่องหมายเหล่านั้นแยกไปอยู่บนโครโมโซมที่แตกต่างกันโดยเฉลี่ยหนึ่งครั้งต่อผลผลิตไมโอซิส 100 ตัว หรือหนึ่งครั้งต่อไมโอซิส 50 ครั้ง
การค้นพบ
กฎการแยกตัวอย่างอิสระของเกรเกอร์ เมนเดลระบุว่าลักษณะทุกอย่างได้รับการถ่ายทอดทางพันธุกรรมอย่างอิสระจากลักษณะอื่นๆ แต่ไม่นานหลังจากที่งานของเมนเดลได้รับการค้นพบใหม่ก็พบข้อยกเว้นของกฎนี้ ในปี ค.ศ. 1905 นักพันธุศาสตร์ชาวอังกฤษวิลเลียม เบตสัน , เอดิธ รีเบคกา ซอนเดอร์สและเรจินัลด์ พุนเน็ตต์ ได้ผสมพันธุ์ต้นถั่วลันเตาในการทดลองที่คล้ายกับของเมนเดล[ 2 ] [ 3 ]พวกเขาสนใจการถ่ายทอดลักษณะทางพันธุกรรมในถั่วลันเตาหวาน และกำลังศึกษายีนสองยีน ได้แก่ ยีนสำหรับสีดอก ( Pสีม่วง และpสีแดง) และยีนที่มีผลต่อรูปร่างของละอองเรณู ( Lยาว และlกลม) พวกเขาผสมพันธุ์สายพันธุ์บริสุทธิ์PPLLและppllแล้วผสมตัวเองกับสายพันธุ์ PpLl ที่ได้
ตามหลักพันธุศาสตร์ของเมนเดลฟีโนไทป์ที่คาดหวังจะเกิดขึ้นในอัตราส่วน 9:3:3:1 ของ PL:Pl:pL:pl แต่ที่น่าประหลาดใจคือ พวกเขาพบว่าความถี่ของ PL และ pl เพิ่มขึ้น และความถี่ของ Pl และ pL ลดลง
| ฟีโนไทป์และจีโนไทป์ | สังเกต | คาดหวังจากอัตราส่วน 9:3:3:1 |
|---|---|---|
| สีม่วง ยาว ( P_L_ ) | 284 | 216 |
| สีม่วง ทรงกลม ( P_ll ) | 21 | 72 |
| สีแดง ยาว ( ppL_ ) | 21 | 72 |
| สีแดง กลม ( ppll ) | 55 | 24 |
การทดลองของพวกเขาเผยให้เห็นความเชื่อมโยงระหว่าง อัลลีล PและLและ อัลลีล pและlความถี่ของ การเกิด Pร่วมกับLและ การเกิด pร่วมกับlนั้นมากกว่าความถี่ของการเกิดPlและpL ที่เกิดจากการรวมตัวใหม่ ความถี่ของการรวมตัวใหม่นั้นคำนวณได้ยากกว่าในการผสมพันธุ์ F2 มากกว่าการผสมพันธุ์แบบย้อนกลับ[ 4 ]แต่ความไม่สอดคล้องกันระหว่างจำนวนลูกหลานที่สังเกตได้และที่คาดหวังในตารางข้างต้นบ่งชี้ว่าน้อยกว่า 50% ซึ่งแสดงให้เห็นว่าปัจจัยสองอย่างมีปฏิสัมพันธ์กันในบางลักษณะเพื่อสร้างความแตกต่างนี้โดยการบดบังการปรากฏของฟีโนไทป์อีกสองแบบ สิ่งนี้นำไปสู่ข้อสรุปว่าลักษณะบางอย่างมีความสัมพันธ์กันเนื่องจากอยู่ใกล้กันบนโครโมโซม
ความเข้าใจเกี่ยวกับการเชื่อมโยงของยีนได้รับการขยายความโดยผลงานของโทมัส ฮันต์ มอร์แกนการสังเกตของมอร์แกนที่ว่าปริมาณการไขว้กันระหว่างยีนที่เชื่อมโยงกันนั้นแตกต่างกัน นำไปสู่แนวคิดที่ว่าความถี่ของการไขว้กันอาจบ่งชี้ถึงระยะห่างระหว่างยีนบนโครโมโซมหน่วยเซนติมอร์แกนซึ่งแสดงถึงความถี่ของการไขว้กัน ได้รับการตั้งชื่อเพื่อเป็นเกียรติแก่เขา
แผนที่เชื่อมโยง
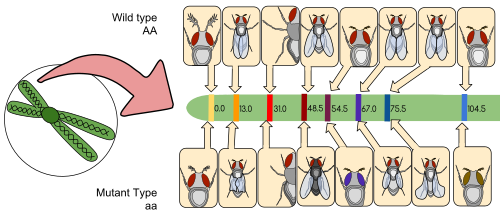
แผนที่เชื่อมโยง (หรือที่เรียกว่าแผนที่พันธุกรรม ) คือตารางสำหรับสิ่งมีชีวิตหรือประชากรทดลองที่แสดงตำแหน่งของยีนหรือเครื่องหมายทางพันธุกรรม ที่ทราบแล้ว สัมพันธ์กันในแง่ของความถี่ของการเกิดการรวมตัวใหม่ แทนที่จะเป็นระยะทางทางกายภาพที่เฉพาะเจาะจงตามแต่ละโครโมโซม แผนที่เชื่อมโยงได้รับการพัฒนาขึ้นครั้งแรกโดยอัลเฟรด สเตอร์เทแวนท์ศิษย์ของโทมัส ฮันท์ มอร์แกน
แผนที่เชื่อมโยง (Linkage map) คือแผนที่ที่สร้างขึ้นจากความถี่ของการเกิดการแลกเปลี่ยนชิ้นส่วน (recombination) ระหว่างเครื่องหมายทางพันธุกรรมในระหว่างการไขว้กันของโครโมโซมคู่เหมือน ยิ่งความถี่ของการเกิดการแลกเปลี่ยนชิ้นส่วน (การแยกตัว) ระหว่างเครื่องหมายทางพันธุกรรมสองตัวสูงเท่าไร ก็ยิ่งสันนิษฐานได้ว่าเครื่องหมายทั้งสองอยู่ห่างกันมากขึ้นเท่านั้น ในทางกลับกัน ยิ่งความถี่ของการเกิดการแลกเปลี่ยนชิ้นส่วนระหว่างเครื่องหมายต่ำเท่าไร ระยะห่างทางกายภาพระหว่างเครื่องหมายทั้งสองก็จะยิ่งน้อยลงเท่านั้น ในอดีต เครื่องหมายที่ใช้ในตอนแรกคือลักษณะ ที่ตรวจจับได้ (การผลิตเอนไซม์ สีตา) ซึ่งได้มาจาก ลำดับ ดีเอ็นเอที่เข้ารหัส ต่อ มาได้มีการใช้ลำดับ ดีเอ็นเอที่ไม่เข้ารหัสที่ได้รับการยืนยันหรือสันนิษฐานแล้วเช่นไมโครแซทเทล ไลต์ หรือลำดับที่สร้างความหลากหลายของความยาวชิ้นส่วนที่ได้จากการตัดด้วยเอนไซม์จำกัด ( RFLPs )
แผนที่การเชื่อมโยงช่วยให้นักวิจัยสามารถระบุตำแหน่งเครื่องหมายอื่นๆ เช่น ยีนอื่นๆ โดยการทดสอบการเชื่อมโยงทางพันธุกรรมของเครื่องหมายที่ทราบอยู่แล้ว ในช่วงเริ่มต้นของการพัฒนาแผนที่การเชื่อมโยง ข้อมูลจะถูกนำมาใช้เพื่อประกอบกลุ่มการเชื่อมโยงซึ่งเป็นชุดของยีนที่ทราบว่ามีการเชื่อมโยงกัน เมื่อความรู้ก้าวหน้าขึ้น สามารถเพิ่มเครื่องหมายเพิ่มเติมลงในกลุ่มได้ จนกระทั่งกลุ่มนั้นครอบคลุมโครโมโซมทั้งหมด[ 6 ]สำหรับสิ่งมีชีวิตที่ได้รับการศึกษามาเป็นอย่างดี กลุ่มการเชื่อมโยงจะสอดคล้องกับโครโมโซมแบบหนึ่งต่อหนึ่ง
แผนที่การเชื่อมโยงไม่ใช่แผนที่ทางกายภาพ (เช่น แผนที่ ลูกผสมที่ลดทอนด้วยรังสี ) หรือแผนที่ยีน
การวิเคราะห์ความเชื่อมโยง
การวิเคราะห์การเชื่อมโยงเป็นวิธีการทางพันธุกรรมที่ค้นหาส่วนของโครโมโซมที่ถ่ายทอดร่วมกับลักษณะอาการของโรคผ่านทางครอบครัว[ 7 ]สามารถใช้ในการทำแผนที่ยีนสำหรับลักษณะทั้งแบบไบนารีและเชิงปริมาณ[ 7 ]การวิเคราะห์การเชื่อมโยงอาจเป็นแบบพาราเมตริก (หากเรารู้ความสัมพันธ์ระหว่างความคล้ายคลึงกันของลักษณะทางฟีโนไทป์และพันธุกรรม) หรือแบบไม่พาราเมตริก การวิเคราะห์การเชื่อมโยงแบบพาราเมตริกเป็นวิธีการแบบดั้งเดิม โดยศึกษาความน่าจะเป็นที่ยีนสำคัญต่อโรคจะเชื่อมโยงกับเครื่องหมายทางพันธุกรรมผ่านคะแนน LOD ซึ่งประเมินความน่าจะเป็นที่แผนผังครอบครัวที่กำหนด ซึ่งโรคและเครื่องหมายมีการถ่ายทอดร่วมกัน เกิดจากการมีอยู่ของการเชื่อมโยง (ด้วยค่าการเชื่อมโยงที่กำหนด) หรือเกิดจากความบังเอิญ ในทางกลับกัน การวิเคราะห์การเชื่อมโยงแบบไม่พาราเมตริกจะศึกษาความน่าจะเป็นที่อัลลีลจะเหมือนกันโดยการสืบทอดกับตัวมันเอง

การวิเคราะห์การเชื่อมโยงแบบพาราเมตริก
คะแนนLOD (ลอการิทึมฐาน 10 ของอัตราต่อรอง) ซึ่งพัฒนาโดยNewton Morton [ 8 ] เป็นการทดสอบทางสถิติที่มักใช้สำหรับการวิเคราะห์การเชื่อมโยงในประชากรมนุษย์ สัตว์ และพืช คะแนน LOD เปรียบเทียบความน่าจะเป็นของการได้รับข้อมูลทดสอบหากตำแหน่งทั้งสองเชื่อมโยงกันจริง กับความน่าจะเป็นของการสังเกตข้อมูลเดียวกันโดยบังเอิญ (นั่นคือ เป็น (ลอการิทึมของ) อัตราส่วนความน่าจะ เป็น ) ดังนั้น เราสามารถเพิ่มคะแนน LOD เข้ากับลอการิทึมของอัตราต่อรองของความน่าจะเป็นก่อนหน้าสำหรับสมมติฐานเพื่อปรับปรุงให้เป็นลอการิทึมของอัตราต่อรองภายหลังตามที่กำหนดโดยกฎของเบย์สนั่นคือ คะแนน LOD คือความแข็งแกร่งของหลักฐาน (ในฐาน 10 แทนที่จะเป็นบิต) สำหรับการเชื่อมโยงในปริมาณที่เท่ากับปริมาณที่สังเกตได้ (โดยพื้นฐานแล้วคือค่าความน่าจะเป็นสูงสุดหรือ ค่า ภายหลังสูงสุด ) คะแนน LOD ที่เป็นบวกสนับสนุนการมีอยู่ของการเชื่อมโยง ในขณะที่คะแนน LOD ที่เป็นลบแสดงว่าการเชื่อมโยงมีโอกาสน้อยลง การวิเคราะห์ค่า LOD ด้วยคอมพิวเตอร์เป็นวิธีที่ง่ายในการวิเคราะห์แผนผังลำดับวงศ์ตระกูลที่ซับซ้อน เพื่อกำหนดความเชื่อมโยงระหว่าง ลักษณะทาง พันธุกรรมแบบเมนเดล (หรือระหว่างลักษณะหนึ่งกับเครื่องหมายทางพันธุกรรม หรือเครื่องหมายทางพันธุกรรมสองตัว)
วิธีการนี้ได้รับการอธิบายโดยละเอียดเพิ่มเติมโดย Strachan และ Read [1]โดยสรุป วิธีการนี้ทำงานดังนี้:
- สร้างประวัติสายพันธุ์
- ทำการประมาณค่าความถี่ของการเกิดการรวมตัวใหม่หลายๆ ค่า
- คำนวณค่า LOD สำหรับแต่ละการประมาณ
- ค่าประมาณที่มีคะแนน LOD สูงที่สุดจะถือเป็นค่าประมาณที่ดีที่สุด
คะแนน LOD คำนวณได้ดังนี้:

NR หมายถึงจำนวนลูกหลานที่ไม่เกิดการรวมตัวใหม่ และ R หมายถึงจำนวนลูกหลานที่เกิดการรวมตัวใหม่ เหตุผลที่ใช้ 0.5 ในตัวส่วนก็เพราะว่าอัลลีลใดๆ ที่ไม่เชื่อมโยงกันโดยสมบูรณ์ (เช่น อัลลีลบนโครโมโซมที่แยกจากกัน) มีโอกาสเกิดการรวมตัวใหม่ 50% เนื่องจากการแยกตัวอย่างอิสระθ คือเศษส่วนของการรวมตัวใหม่ กล่าว คือ เศษส่วนของการเกิดที่เกิดการรวมตัวใหม่ระหว่างเครื่องหมายทางพันธุกรรมที่ศึกษาและยีนที่คาดว่าเกี่ยวข้องกับโรค ดังนั้นจึงเท่ากับR / ( NR + R )
ตามธรรมเนียมปฏิบัติ ค่า LOD ที่มากกว่า 3.0 ถือเป็นหลักฐานของการเชื่อมโยง เนื่องจากบ่งชี้ว่ามีโอกาส 1,000 ต่อ 1 ที่การเชื่อมโยงที่สังเกตได้นั้นไม่ได้เกิดขึ้นโดยบังเอิญ ในทางกลับกัน ค่า LOD ที่น้อยกว่า -2.0 ถือเป็นหลักฐานในการยกเว้นการเชื่อมโยง แม้ว่าจะเป็นไปได้ยากมากที่จะได้ค่า LOD เท่ากับ 3 จากแผนผังครอบครัวเดียว แต่คุณสมบัติทางคณิตศาสตร์ของการทดสอบอนุญาตให้รวมข้อมูลจากแผนผังครอบครัวจำนวนมากเข้าด้วยกันโดยการบวกค่า LOD เข้าด้วยกัน ค่า LOD เท่ากับ 3 แปลงเป็นค่าpประมาณ 0.05 [ 9 ]และไม่ จำเป็นต้อง มีการแก้ไขการทดสอบหลายครั้ง (เช่นการแก้ไข Bonferroni ) [ 10 ]
ข้อจำกัด
การวิเคราะห์การเชื่อมโยงมีข้อจำกัดทางระเบียบวิธีและทฤษฎีหลายประการที่สามารถเพิ่ม อัตรา ความผิดพลาดประเภทที่ 1 อย่างมีนัยสำคัญ และลดพลังในการระบุตำแหน่งยีนควบคุมลักษณะเชิงปริมาณของมนุษย์ (QTL) [ 11 ]แม้ว่าการวิเคราะห์การเชื่อมโยงจะถูกนำมาใช้เพื่อระบุตัวแปรทางพันธุกรรมที่ก่อให้เกิดโรคหายาก เช่นโรคฮันติงตัน ได้สำเร็จ แต่ก็ไม่ได้ผลดีนักเมื่อนำไปใช้กับโรคที่พบได้บ่อยกว่า เช่นโรคหัวใจ หรือ มะเร็งชนิดต่างๆ[ 12 ] คำอธิบายสำหรับเรื่องนี้คือกลไกทางพันธุกรรมที่ส่งผลต่อโรคที่พบได้บ่อย นั้นแตกต่างจากกลไกที่ก่อให้เกิดโรคหายากบางชนิด[ 13 ]
ความถี่ของการรวมตัวใหม่
ความถี่ของการเกิดการรวมตัวใหม่ (Recombination frequency) เป็นตัววัดความเชื่อมโยงทางพันธุกรรม และใช้ในการสร้างแผนที่ความเชื่อมโยงทางพันธุกรรม ความถี่ของการเกิดการรวมตัวใหม่ ( θ ) คือความถี่ที่การไขว้กันของโครโมโซม เพียงครั้งเดียว จะเกิดขึ้นระหว่างยีนสองยีนในระหว่างการแบ่งเซลล์แบบไมโอซิสเซนติมอร์แกน ( cM) เป็นหน่วยที่อธิบายความถี่ของการเกิดการรวมตัวใหม่ 1% ด้วยวิธีนี้เราสามารถวัดระยะทางทางพันธุกรรมระหว่างตำแหน่งสองตำแหน่งโดยอาศัยความถี่ของการเกิดการรวมตัวใหม่ ซึ่งเป็นการประมาณระยะทางที่แท้จริงได้ดี การไขว้กันสองครั้งจะกลายเป็นไม่มีการรวมตัวใหม่ ในกรณีนี้เราไม่สามารถบอกได้ว่ามีการไขว้กันเกิดขึ้นหรือไม่ หากตำแหน่งที่เรากำลังวิเคราะห์อยู่ใกล้กันมาก (น้อยกว่า 7 cM) การไขว้กันสองครั้งนั้นไม่น่าจะเกิดขึ้น เมื่อระยะทางสูงขึ้น ความน่าจะเป็นของการไขว้กันสองครั้งก็จะเพิ่มขึ้น เมื่อความน่าจะเป็นของการไขว้กันสองครั้งเพิ่มขึ้น เราอาจประเมินระยะทางทางพันธุกรรมระหว่างตำแหน่งสองตำแหน่งต่ำกว่าความเป็นจริงอย่างเป็นระบบ เว้นแต่เราจะใช้แบบจำลองทางคณิตศาสตร์ที่เหมาะสม
การเชื่อมโยงแบบคู่เป็นข้อกังวลทางประวัติศาสตร์สำหรับพืชมากกว่า ในสัตว์ การไขว้กันแบบคู่เกิดขึ้นได้ยาก ตัวอย่างเช่น ในมนุษย์ โครโมโซมหนึ่งคู่มีการไขว้กันสองครั้งโดยเฉลี่ยในระหว่างการแบ่งเซลล์แบบไมโอซิส ยิ่งไปกว่านั้น นักพันธุศาสตร์สมัยใหม่มีจำนวนยีนมากพอที่จะต้องวิเคราะห์การเชื่อมโยงเฉพาะยีนที่อยู่ใกล้เคียงเท่านั้น ซึ่งแตกต่างจากในยุคแรกๆ ที่รู้จักยีนเพียงไม่กี่ตัว[ 14 ]
ในระหว่างกระบวนการแบ่งเซลล์แบบไมโอซิส โครโมโซมจะแยกตัวแบบสุ่มเข้าไปในเซลล์สืบพันธุ์โดยที่การแยกตัวของอัลลีลของยีนหนึ่งจะเป็นอิสระจากการแยกตัวของอัลลีลของยีนอีกยีนหนึ่ง นี่คือสิ่งที่ระบุไว้ในกฎข้อที่สองของเมนเดลและรู้จักกันในชื่อกฎการแยกตัวอย่างอิสระกฎการแยกตัวอย่างอิสระนี้เป็นจริงเสมอสำหรับยีนที่อยู่บนโครโมโซมต่างกัน แต่สำหรับยีนที่อยู่บนโครโมโซมเดียวกันนั้น กฎนี้อาจไม่เป็นจริงเสมอไป
ตัวอย่างของการแยกตัวอย่างอิสระ คือ การผสมพันธุ์ระหว่างสายพันธุ์พ่อแม่ที่เป็นโฮโมไซกัส บริสุทธิ์ที่มี จีโนไทป์AABBกับสายพันธุ์บริสุทธิ์อีกสายพันธุ์หนึ่งที่มีจีโนไทป์aabbโดยที่ A และ a และ B และ b แทนอัลลีลของยีน A และ B การผสมพันธุ์ระหว่างสายพันธุ์พ่อแม่ที่เป็นโฮโมไซกัสเหล่านี้จะทำให้ได้ลูกหลานรุ่น F1 ที่เป็นเฮเทโรไซกัส คู่ ที่มีจีโนไทป์ AaBb ลูกหลานรุ่น F1 ที่มีจีโนไทป์ AaBb จะสร้างแกมีตที่เป็นAB , Ab , aBและabด้วยความถี่เท่ากัน (25%) เนื่องจากอัลลีลของยีน A แยกตัวอย่างอิสระจากอัลลีลของยีน B ในระหว่างการแบ่งเซลล์แบบไมโอซิส โปรดสังเกตว่า 2 ใน 4 แกมีต (50%)— AbและaB —ไม่มีอยู่ในรุ่นพ่อแม่ แกมีตเหล่านี้เป็นแกมีตลูกผสม แกมีตลูกผสมคือแกมีตที่แตกต่างจากแกมีต แฮพลอยด์ทั้งสองที่ประกอบขึ้นเป็นเซลล์ดิพลอยด์ ดั้งเดิม ในตัวอย่างนี้ ความถี่ของการเกิดการรวมตัวใหม่คือ 50% เนื่องจากแกมีต 2 ใน 4 ตัวเป็นแกมีตที่เกิดจากการรวมตัวใหม่
ความถี่ของการเกิดการรวมตัวใหม่จะมีค่า 50% เมื่อยีนสองยีนอยู่บนโครโมโซม ที่แตกต่างกัน หรือเมื่อยีนทั้งสองอยู่ห่างกันมากบนโครโมโซมเดียวกัน นี่เป็นผลมาจากการแยกตัวอย่างอิสระ
เมื่อยีนสองยีนอยู่ใกล้กันบนโครโมโซมเดียวกัน ยีนเหล่านั้นจะไม่แยกตัวออกจากกันอย่างอิสระและเรียกว่ายีนที่เชื่อมโยงกัน ในขณะที่ยีนที่อยู่บนโครโมโซมต่างกันจะแยกตัวออกจากกันอย่างอิสระและมีอัตราการเกิดการรวมตัวใหม่ 50% แต่ยีนที่เชื่อมโยงกันจะมีอัตราการเกิดการรวมตัวใหม่น้อยกว่า 50%
ตัวอย่างของการเชื่อมโยง ลองพิจารณาการทดลองคลาสสิกของWilliam BatesonและReginald Punnett [ 15 ] พวก เขาสนใจการถ่ายทอดลักษณะทางพันธุกรรมในถั่วลันเตาหวาน และกำลังศึกษายีนสองตัว คือ ยีนสำหรับสีดอก ( Pสีม่วง และpสีแดง) และยีนที่มีผลต่อรูปร่างของละอองเรณู ( Lยาว และlกลม) พวกเขาผสมพันธุ์สายพันธุ์บริสุทธิ์PPLLและppllแล้วผสมตัวเองกับ สายพันธุ์ PpLl ที่ได้ ตามพันธุศาสตร์ของเมนเดล ฟีโนไทป์ที่คาดหวังจะเกิดขึ้นในอัตราส่วน 9:3:3:1 ของ PL:Pl:pL:pl แต่ที่น่าประหลาดใจคือ พวกเขาพบความถี่ของ PL และ pl เพิ่มขึ้น และความถี่ของ Pl และ pL ลดลง (ดูตารางด้านล่าง)
| ฟีโนไทป์และจีโนไทป์ | สังเกต | คาดหวังจากอัตราส่วน 9:3:3:1 |
|---|---|---|
| สีม่วง ยาว ( P_L_ ) | 284 | 216 |
| สีม่วง ทรงกลม ( P_ll ) | 21 | 72 |
| สีแดง ยาว ( ppL_ ) | 21 | 72 |
| สีแดง กลม ( ppll ) | 55 | 24 |

การทดลองของพวกเขาเผยให้เห็นการเชื่อมโยงระหว่าง อัลลีล PและLและ อัลลีล pและlความถี่ของ การเกิด Pร่วมกับLและ การเกิด pร่วมกับlนั้นมากกว่าความถี่ของการเกิดPlและpL ที่เป็นรีคอม บิแนนท์ ความถี่ของการเกิดรีคอมบิแนนท์นั้นคำนวณได้ยากกว่าในการผสมพันธุ์ F2 มากกว่าการผสมพันธุ์แบบย้อนกลับ[ 4 ]แต่การที่จำนวนลูกหลานที่สังเกตได้และที่คาดหวังในตารางข้างต้นไม่ตรงกัน แสดงให้เห็นว่าความถี่นั้นน้อยกว่า 50%
ในกรณีนี้ ลูกหลานได้รับอัลลีลเด่นสองตัวที่เชื่อมโยงกันบนโครโมโซมเดียวกัน (เรียกว่าการจับคู่หรือการจัดเรียงแบบซิส ) อย่างไรก็ตาม หลังจากครอสโอเวอร์ ลูกหลานบางตัวอาจได้รับโครโมโซมจากพ่อแม่ตัวหนึ่งที่มีอัลลีลเด่นสำหรับลักษณะหนึ่ง (เช่น สีม่วง) เชื่อมโยงกับอัลลีลด้อยสำหรับลักษณะที่สอง (เช่น กลม) ในขณะที่โครโมโซมอีกตัวหนึ่งเป็นไปในทางตรงกันข้าม (เช่น สีแดงและยาว) สิ่งนี้เรียกว่าการผลักกันหรือการจัดเรียงแบบทรานส์ ฟีโนไทป์ในกรณีนี้จะยังคงเป็นสีม่วงและยาว แต่การผสมข้ามพันธุ์ระหว่างตัวนี้กับพ่อแม่ที่มีลักษณะด้อยจะทำให้ได้ลูกหลานที่มีสัดส่วนของฟีโนไทป์ครอสโอเวอร์ทั้งสองแบบมากขึ้น แม้ว่าปัญหาดังกล่าวอาจดูไม่น่าเกิดขึ้นจากตัวอย่างนี้ แต่การเชื่อมโยงแบบผลักกันที่ไม่พึงประสงค์ก็ปรากฏขึ้นเมื่อทำการผสมพันธุ์เพื่อต้านทานโรคในพืชบางชนิด
การจัดเรียงอัลลีลสองแบบที่เป็นไปได้ คือ ซิส (cis) และ ทรานส์ (trans) ในเฮเทโรไซโกตคู่ เรียกว่าเฟสของเซลล์สืบพันธุ์และการกำหนดเฟสคือ กระบวนการในการพิจารณาว่าเซลล์สืบพันธุ์แต่ละเซลล์มีเฟสแบบใดอยู่
เมื่อยีนสองยีนอยู่บนโครโมโซมเดียวกัน โอกาสที่จะ เกิดการไขว้กันจนเกิดการรวม ตัวใหม่ระหว่างยีนนั้นจะสัมพันธ์กับระยะห่างระหว่างยีนทั้งสอง ดังนั้น การใช้ความถี่ของการรวมตัวใหม่จึงถูกนำมาใช้ในการพัฒนาแผนที่เชื่อมโยงหรือแผนที่ทางพันธุกรรม
อย่างไรก็ตาม ความถี่ของการรวมตัวใหม่มักจะประเมินระยะห่างระหว่างยีนที่เชื่อมโยงกันสองยีนต่ำกว่าความเป็นจริง เนื่องจากเมื่อยีนทั้งสองอยู่ห่างกันมากขึ้น โอกาสที่จะเกิดการไขว้กันสองครั้งหรือจำนวนคู่ระหว่างยีนทั้งสองก็จะเพิ่มขึ้นด้วย การไขว้กันสองครั้งหรือจำนวนคู่ระหว่างยีนทั้งสองส่งผลให้ยีนทั้งสองถูกแยกไปอยู่ในเซลล์สืบพันธุ์เดียวกัน ทำให้ได้ลูกหลานที่มีลักษณะเหมือนพ่อแม่ แทนที่จะเป็นลูกหลานที่เกิดจากการรวมตัวใหม่ตามที่คาดไว้ ดังที่กล่าวไว้ข้างต้นการแปลง Kosambi และ Haldaneพยายามแก้ไขการไขว้กันหลายครั้ง[ 16 ]
การเชื่อมโยงของตำแหน่งทางพันธุกรรมภายในยีน
ในช่วงต้นทศวรรษ 1950 มุมมองที่แพร่หลายคือยีนในโครโมโซมเป็นหน่วยที่แยกจากกัน ไม่สามารถแบ่งแยกได้ด้วยการรวมตัวทางพันธุกรรมและเรียงตัวเหมือนลูกปัดบนเส้นด้าย ในช่วงปี 1955 ถึง 1959 เบนเซอร์ได้ทำการ ทดลอง การรวมตัวทางพันธุกรรมโดยใช้ แบค ทีริโอเฟจ T4 กลายพันธุ์rIIเขาพบว่าจากการทดสอบการรวมตัวทางพันธุกรรม ตำแหน่งของการกลายพันธุ์สามารถระบุได้ในลำดับเชิงเส้น[ 17 ] [ 18 ]ผลลัพธ์นี้เป็นหลักฐานสนับสนุนแนวคิดหลักที่ว่ายีนมีโครงสร้างเชิงเส้นเทียบเท่ากับความยาวของDNAที่มีหลายตำแหน่งที่สามารถกลายพันธุ์ได้อย่างอิสระ
Edgar และคณะ[ 19 ]ได้ทำการทดลองการทำแผนที่ด้วย r mutants ของแบคทีริโอเฟจ T4 โดยแสดงให้เห็นว่าความถี่ของการรวมตัวใหม่ระหว่าง rII mutants นั้นไม่ได้เป็นการบวกกันอย่างเคร่งครัด ความถี่ของการรวมตัวใหม่จากการผสมข้ามของ rII mutants สองตัว (axd) มักจะน้อยกว่าผลรวมของความถี่ของการรวมตัวใหม่สำหรับช่วงย่อยภายในที่อยู่ติดกัน (axb) + (bxc) + (cxd) แม้ว่าจะไม่เป็นการบวกกันอย่างเคร่งครัด แต่ก็สังเกตเห็นความสัมพันธ์ที่เป็นระบบ[ 20 ]ซึ่งน่าจะสะท้อนถึงกลไกโมเลกุลพื้นฐานของ การรวมตัว ทาง พันธุกรรม
ความแปรผันของความถี่การรวมตัวใหม่
แม้ว่าการรวมตัวใหม่ของโครโมโซมจะเป็นกระบวนการที่จำเป็นในระหว่างการแบ่งเซลล์แบบไมโอซิส แต่ความถี่ของการไขว้กันนั้นมีความหลากหลายมากในสิ่งมีชีวิตต่างๆ และภายในสายพันธุ์เดียวกัน อัตราการรวมตัวใหม่ที่แตกต่างกันระหว่างเพศเรียกว่า heterochiasmy และพบได้บ่อยกว่าอัตราปกติระหว่างเพศผู้และเพศเมีย ในสัตว์เลี้ยงลูกด้วยนม เพศเมียมักมีอัตราการรวมตัวใหม่สูงกว่าเพศผู้ มีทฤษฎีว่ามีการคัดเลือกเฉพาะหรือตัวขับเคลื่อนไมโอซิสที่ส่งผลต่อความแตกต่างของอัตรา ความแตกต่างของอัตราอาจสะท้อนถึงสภาพแวดล้อมและเงื่อนไขของการแบ่งเซลล์แบบไมโอซิสที่แตกต่างกันอย่างมากในกระบวนการสร้างไข่และการสร้างอสุจิ[ 21 ]
ยีนที่มีผลต่อความถี่ของการเกิดการรวมตัวใหม่
การกลายพันธุ์ในยีนที่เข้ารหัสโปรตีนที่เกี่ยวข้องกับการประมวลผล DNA มักส่งผลต่อ ความถี่ ของการรวมตัวใหม่ในแบคทีริโอเฟจ T4การกลายพันธุ์ที่ลดการแสดงออกของDNA polymerase ที่ใช้ในการ จำลองแบบ [ผลิตภัณฑ์ยีน 43 (gp43)] จะเพิ่มการรวมตัวใหม่ (ลดการเชื่อมโยง) หลายเท่า[ 22 ] [ 23 ] การเพิ่มขึ้นของการรวมตัวใหม่อาจเกิดจาก ข้อผิดพลาด ในการจำลองแบบโดย DNA polymerase ที่บกพร่อง ซึ่งเป็นเหตุการณ์การรวมตัวใหม่เช่นกัน เช่น การสลับแม่แบบ กล่าวคือ เหตุการณ์การรวมตัวใหม่แบบเลือกสำเนา[ 24 ]การรวมตัวใหม่ยังเพิ่มขึ้นจากการกลายพันธุ์ที่ลดการแสดงออกของDNA ligase (gp30) [ 25 ] [ 23 ]และ dCMP hydroxymethylase (gp42) [ 22 ] [ 23 ]ซึ่งเป็นเอนไซม์สองชนิดที่ใช้ใน การ สังเคราะห์ DNA
การรวมตัวใหม่ลดลง (การเชื่อมโยงเพิ่มขึ้น) โดยการกลายพันธุ์ในยีนที่เข้ารหัสโปรตีนที่มี ฟังก์ชัน นิวคลีเอส (gp46 และ gp47) [ 25 ] [ 23 ]และโปรตีนที่จับกับ DNA (gp32) [ 23 ] การกลายพันธุ์ในยีน uvsX ของแบคทีริโอเฟจยังลดการรวมตัวใหม่ลงอย่างมาก[ 26 ] ยีน uvsX คล้ายคลึงกับยีนrecA ที่ได้รับการศึกษาอย่างดีของ Escherichia coliซึ่งมีบทบาทสำคัญในการรวมตัวใหม่[ 27 ]
ตัวบ่งชี้การแบ่งเซลล์แบบไมโอซิส
ด้วยแผนผังลำดับวงศ์ตระกูลขนาดใหญ่ หรือข้อมูลเครื่องหมายทางพันธุกรรมที่หนาแน่นมาก เช่น จากการลำดับจีโนมทั้งหมด ทำให้สามารถระบุตำแหน่งของการเกิดการรวมตัวใหม่ได้อย่างแม่นยำ การวิเคราะห์ทางพันธุกรรมแบบนี้ จะกำหนดตัวบ่งชี้การแบ่งเซลล์แบบไมโอซิสให้กับแต่ละตำแหน่งของจีโนมในแต่ละไมโอซิสในแผนผังลำดับวงศ์ตระกูล ตัวบ่งชี้จะระบุว่าโครโมโซมจากพ่อแม่ชุดใดมีส่วนในการสร้างเซลล์สืบพันธุ์ในตำแหน่งนั้น ตัวอย่างเช่น หากอัลลีลจากโครโมโซมชุดที่ 'แรก' ของพ่อแม่ถูกส่งต่อ อาจกำหนดค่า '0' ให้กับไมโอซิสนั้น หากอัลลีลจากโครโมโซมชุดที่ 'ที่สอง' ของพ่อแม่ถูกส่งต่อ จะกำหนดค่า '1' ให้กับไมโอซิสนั้น อัลลีลทั้งสองในพ่อแม่มาจากปู่ย่าตายายคนละชุด ตัวบ่งชี้เหล่านี้จะถูกนำมาใช้เพื่อกำหนดสถานะการสืบทอดทางพันธุกรรมที่เหมือนกัน (IBD) หรือสถานะการถ่ายทอดทางพันธุกรรม ซึ่งจะนำไปใช้ในการระบุยีนที่ก่อให้เกิดโรคต่อไป
ดูเพิ่มเติม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การเชื่อมโยงทางพันธุกรรม
การเชื่อมโยงทางพันธุกรรมคือแนวโน้มของลำดับดีเอ็นเอที่อยู่ใกล้กันบนโครโมโซมที่จะถูกถ่ายทอดไปด้วยกันในระหว่าง ระยะ
การค้นพบ
กฎการแยกตัวอย่างอิสระ ของ เกรเกอร์ เมนเดล ระบุว่าลักษณะทุกอย่างได้รับการถ่ายทอดทางพันธุกรรมอย่างอิสระจากลักษณะอื่นๆ แต่ไม่นานหลังจากที่ งานของเมนเดลได้รับการค้นพบใหม่ ก็พบข้อยกเว้นของกฎนี้ ในปี ค.ศ.
แผนที่เชื่อมโยง
แผนที่ เชื่อมโยง (หรือที่เรียกว่า แผนที่พันธุกรรม ) คือตารางสำหรับสิ่งมีชีวิตหรือประชากรทดลองที่แสดงตำแหน่งของ ยีน หรือ เครื่องหมายทางพันธุกรรม ที่ทราบแล้ว สัมพันธ์กันในแง่ของความถี่ของการเกิดการรวมตัวใหม่...
การวิเคราะห์ความเชื่อมโยง
การวิเคราะห์การเชื่อมโยงเป็นวิธีการทางพันธุกรรมที่ค้นหาส่วนของโครโมโซมที่ ถ่ายทอดร่วม กับลักษณะอาการของโรคผ่านทางครอบครัว [ 7 ] สามารถใช้ในการทำแผนที่ยีนสำหรับลักษณะทั้งแบบไบนารีและเชิงปริมาณ [ 7 ] การวิเคราะห์การเชื่อมโยงอาจเป็นแบบพาราเมตริก...