อ่าน 12 นาที
ตัวรับอินซูลิน
ตัว รับอินซูลิน ( IR ) เป็น ตัวรับแบบทรานส์เมมเบรน ที่ถูกกระตุ้นโดย อินซูลิน , IGF-I , IGF-II และอยู่ในกลุ่มใหญ่ของ ตัวรับไทโรซีนไคเนส [ 5 ] ใน ทางเมตาบอลิซึม...
ตัวรับอินซูลิน
| อินเอสอาร์ | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | INSR , CD220, HHF5, ตัวรับอินซูลิน | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 147670 ; เอ็มจีไอ : 96575 ; โฮโมโลยีน : 20090 ; GeneCards : INSR ; OMA : INSR - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




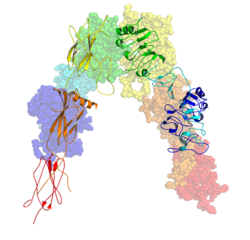
ตัวรับอินซูลิน ( IR ) เป็นตัวรับแบบทรานส์เมมเบรนที่ถูกกระตุ้นโดยอินซูลิน , IGF-I , IGF-IIและอยู่ในกลุ่มใหญ่ของตัวรับไทโรซีนไคเนส [ 5 ] ในทางเมตาบอลิซึม ตัวรับอินซูลินมีบทบาทสำคัญในการควบคุมภาวะสมดุลของกลูโคสซึ่งเป็นกระบวนการทำงานที่หากอยู่ในสภาวะที่เสื่อมลง อาจส่งผลให้เกิดอาการทางคลินิกต่างๆ รวมถึงโรคเบาหวานและมะเร็ง[ 6 ] [ 7 ]การส่งสัญญาณของอินซูลินควบคุมการเข้าถึงกลูโคสในเลือดในเซลล์ร่างกาย เมื่ออินซูลินลดลง โดยเฉพาะในผู้ที่มีความไวต่ออินซูลินสูง เซลล์ร่างกายจะเริ่มเข้าถึงเฉพาะไขมันที่ไม่ต้องขนส่งผ่านเยื่อหุ้มเซลล์ ดังนั้น ในแง่นี้ อินซูลินจึงเป็นตัวควบคุมหลักของการเผาผลาญไขมันเช่นกัน ในทางชีวเคมี ตัวรับอินซูลินถูกเข้ารหัสโดยยีน เดี่ยว INSRซึ่ง การ ตัดต่อแบบสลับกัน ระหว่างการถอดรหัสส่งผลให้เกิด ไอโซฟอร์ม IR-A หรือ IR- B [ 8 ]เหตุการณ์หลังการแปลรหัสปลายทางของไอโซฟอร์มใดไอโซฟอร์มหนึ่งส่งผลให้เกิดการสร้างซับยูนิต α และ β ที่ถูกตัดด้วยโปรตีโอไลติก ซึ่งเมื่อรวมกันแล้วจะสามารถเกิดการรวมตัวกันเป็นโฮโมไดเมอร์หรือเฮเทโรไดเมอร์เพื่อสร้างตัวรับอินซูลินทรานส์เมมเบรนที่เชื่อมด้วยไดซัลไฟด์ขนาด ≈320 kDa ได้ในที่สุด[ 8 ]
โครงสร้าง
ในขั้นต้นการถอดรหัสของตัวแปรการต่อเชื่อมทางเลือกที่ได้มาจาก ยีน INSRจะถูกแปลเป็นโปรตีนโมโนเมอร์ 2 ชนิด ได้แก่ IR-A ซึ่ง ไม่มี เอ็กซอน 11 และ IR-B ซึ่งมีเอ็กซอน 11 การรวมเอ็กซอน 11 ส่งผลให้มีการเพิ่มกรดอะมิโน 12 ตัวก่อนหน้าตำแหน่งการตัดของเอนไซม์ฟูริน

เมื่อเกิดการไดเมอไรเซชันของตัวรับ หลังจากการตัดแยกโปรตีนออกเป็นโซ่ α และ β กรดอะมิโนเพิ่มเติม 12 ตัวจะยังคงอยู่บริเวณปลาย Cของโซ่ α (เรียกว่า αCT) ซึ่งคาดว่าจะส่งผลต่อปฏิสัมพันธ์ ระหว่างตัวรับและ ลิแกนด์[ 9 ]
โมโนเมอร์ไอโซเมตริกแต่ละตัวมีโครงสร้างที่จัดเรียงเป็น 8 โดเมนที่แตกต่างกัน ประกอบด้วย โดเมนที่มีลิวซีนเป็นองค์ประกอบหลัก (L1, กรดอะมิโนตำแหน่งที่ 1–157), บริเวณที่มีซิสเทอีนเป็นองค์ประกอบหลัก (CR, กรดอะมิโนตำแหน่งที่ 158–310), โดเมนที่มีลิวซีนเป็นองค์ประกอบหลักเพิ่มเติม (L2, กรดอะมิโนตำแหน่งที่ 311–470), และโดเมนไฟโบรเนคตินชนิด III สามโดเมน ได้แก่ FnIII-1 (กรดอะมิโนตำแหน่งที่ 471–595), FnIII-2 (กรดอะมิโนตำแหน่งที่ 596–808) และ FnIII-3 (กรดอะมิโนตำแหน่งที่ 809–906) นอกจากนี้ ยังมีโดเมนแทรก (ID, กรดอะมิโนตำแหน่งที่ 638–756) อยู่ภายใน FnIII-2 ซึ่งมีตำแหน่งการตัดของฟูริน α/β โดยการย่อยสลายโปรตีนจะทำให้เกิดโดเมน IDα และ IDβ ขึ้น ภายในโซ่ β ปลายน้ำของโดเมน FnIII-3 มีเกลียวทรานส์เมมเบรน (TH) และบริเวณจุกซ์ตาเมมเบรนภายในเซลล์ (JM) อยู่เหนือโดเมนเร่งปฏิกิริยาไทโรซีนไคเนส (TK) ภายในเซลล์ ซึ่งรับผิดชอบเส้นทางการส่งสัญญาณภายในเซลล์ต่อไป[ 10 ]
เมื่อโมโนเมอร์แยกออกเป็นโซ่ α และ β ตามลำดับ การสร้างไดเมอร์แบบเฮเทอโรหรือโฮโมของตัวรับจะคงอยู่ด้วยพันธะโควาเลนต์ระหว่างโซ่ด้วยพันธะไดซัลไฟด์เดี่ยว และระหว่างโมโนเมอร์ในไดเมอร์ด้วยพันธะไดซัลไฟด์สองพันธะที่ยื่นออกมาจากโซ่ α แต่ละโซ่ โครงสร้าง เอกโต โดเมน 3 มิติโดยรวม ซึ่งมีไซต์การจับลิแกนด์สี่ไซต์ มีลักษณะคล้ายตัว 'V' คว่ำ โดยแต่ละโมโนเมอร์หมุนประมาณ 2 เท่ารอบแกนที่ขนานกับตัว 'V' คว่ำ และโดเมน L2 และ FnIII-1 จากแต่ละโมโนเมอร์จะก่อตัวเป็นยอดของตัว 'V' คว่ำ[ 10 ] [ 11 ]
การจับตัวของลิแกนด์


ลิแกนด์ภายในของตัวรับอินซูลิน ได้แก่อินซูลิน , IGF-IและIGF-IIการใช้cryo-EMทำให้ได้ข้อมูลเชิงลึกเชิงโครงสร้างเกี่ยวกับการเปลี่ยนแปลงรูปร่างเมื่อมีการจับกับอินซูลิน การจับของลิแกนด์กับ α-chain ของ ectodomain แบบไดเมอร์ของ IR จะเปลี่ยนรูปร่างจากรูปตัววีคว่ำเป็นรูปตัว T และการเปลี่ยนแปลงนี้จะแพร่กระจายไปยังโดเมนทรานส์เมมเบรน ซึ่งจะเข้าใกล้กันมากขึ้น จนในที่สุดนำไปสู่การฟอสโฟรีเลชันตัวเองของสารตกค้างไทโรซีนต่างๆ ภายในโดเมน TK ภายในเซลล์ของ β-chain [ 12 ]การเปลี่ยนแปลงเหล่านี้ช่วยอำนวยความสะดวกในการดึงดูดโปรตีนอะแดปเตอร์ เฉพาะ เช่น โปรตีนซับสเตรตของตัวรับอินซูลิน (IRS) นอกเหนือจากSH2-B ( Src Homology 2 - B), APSและโปรตีนฟอสฟาเทส เช่นPTP1Bซึ่งในที่สุดจะส่งเสริมกระบวนการปลายน้ำที่เกี่ยวข้องกับการรักษาสมดุลของระดับน้ำตาลในเลือด[ 14 ]
กล่าวอย่างเคร่งครัด ความสัมพันธ์ระหว่าง IR และลิแกนด์แสดงคุณสมบัติอัลโลสเตอริกที่ซับซ้อน สิ่งนี้ได้รับการระบุโดยการใช้แผนภาพ Scatchardซึ่งระบุว่าการวัดอัตราส่วนของลิแกนด์ที่จับกับ IR ต่อลิแกนด์ที่ไม่จับนั้นไม่เป็นไปตามความสัมพันธ์เชิงเส้นกับการเปลี่ยนแปลงความเข้มข้นของลิแกนด์ที่จับกับ IR ซึ่งชี้ให้เห็นว่า IR และลิแกนด์ที่เกี่ยวข้องมีความสัมพันธ์ของการจับแบบร่วมมือกัน [ 15 ] นอกจากนี้ การสังเกตว่าอัตราการแยกตัวของ IR-ลิแกนด์เร่งขึ้นเมื่อเติมลิแกนด์ที่ไม่จับนั้นหมายความว่าลักษณะของการร่วมมือนี้เป็นลบ กล่าวอีกนัยหนึ่ง การจับเริ่มต้นของลิแกนด์กับ IR จะยับยั้งการจับเพิ่มเติมกับไซต์ที่ใช้งานที่สองของมัน ซึ่งเป็นการแสดงออกของการยับยั้งแบบอัลโลสเตอริก[ 15 ]
แบบจำลองเหล่านี้ระบุว่าโมโนเมอร์ IR แต่ละตัวมีไซต์การจับอินซูลิน 2 ไซต์ ไซต์ 1 ซึ่งจับกับพื้นผิวการจับแบบ 'คลาสสิก' ของอินซูลินประกอบด้วยโดเมน L1 บวก αCT และไซต์ 2 ซึ่งประกอบด้วยลูปที่จุดเชื่อมต่อของ FnIII-1 และ FnIII-2 ที่คาดว่าจะจับกับไซต์การจับแบบ 'ใหม่' ของหน้าเฮกซาเมอร์ของอินซูลิน[ 5 ]เนื่องจากโมโนเมอร์แต่ละตัวที่ประกอบเป็นเอกโตโดเมนของ IR แสดงความสมบูรณ์แบบ 'สะท้อน' 3 มิติ ไซต์ N-เทอร์มินัล 1 ของโมโนเมอร์หนึ่งจะหันหน้าเข้าหาไซต์ C-เทอร์มินัล 2 ของโมโนเมอร์ที่สอง ซึ่งเป็นความจริงสำหรับส่วนเติมเต็มแบบสะท้อนของแต่ละโมโนเมอร์ (ด้านตรงข้ามของโครงสร้างเอกโตโดเมน) วรรณกรรมปัจจุบันแยกแยะไซต์การจับส่วนเติมเต็มโดยกำหนดชื่อไซต์ 1 และไซต์ 2 ของโมโนเมอร์ที่สองเป็นไซต์ 3 และไซต์ 4 หรือเป็นไซต์ 1' และไซต์ 2' ตามลำดับ[ 5 ] [ 14 ] ด้วยเหตุนี้ แบบจำลองเหล่านี้จึงระบุว่า IR แต่ละตัวอาจจับกับโมเลกุลอินซูลิน (ซึ่งมีพื้นผิวการจับสองพื้นผิว) ผ่าน 4 ตำแหน่ง ได้แก่ ตำแหน่งที่ 1, 2, (3/1') หรือ (4/2') เนื่องจากตำแหน่งที่ 1 แต่ละตำแหน่งหันหน้าเข้าหาตำแหน่งที่ 2 อย่างใกล้ชิด เมื่ออินซูลินจับกับตำแหน่งเฉพาะ จะคาดการณ์ว่าจะเกิด 'การเชื่อมโยงข้าม'ผ่านลิแกนด์ระหว่างโมโนเมอร์ (เช่น [โมโนเมอร์ 1 ตำแหน่งที่ 1 - อินซูลิน - โมโนเมอร์ 2 ตำแหน่ง (4/2')] หรือ [โมโนเมอร์ 1 ตำแหน่งที่ 2 - อินซูลิน - โมโนเมอร์ 2 ตำแหน่ง (3/1')]) ตามแบบจำลองทางคณิตศาสตร์ปัจจุบันของจลนศาสตร์ IR-อินซูลิน มีผลลัพธ์ที่สำคัญสองประการต่อเหตุการณ์การเชื่อมโยงข้ามของอินซูลิน 1. จากการสังเกตความร่วมมือเชิงลบระหว่าง IR และลิแกนด์ที่กล่าวมาข้างต้น การจับลิแกนด์กับ IR ในภายหลังจะลดลง และ 2. การกระทำทางกายภาพของการเชื่อมโยงข้ามทำให้เอ็กโทโดเมนอยู่ในโครงสร้างที่จำเป็นสำหรับการเกิดฟอสโฟรีเลชันของไทโรซีนภายในเซลล์ (กล่าวคือ เหตุการณ์เหล่านี้ทำหน้าที่เป็นข้อกำหนดสำหรับการกระตุ้นตัวรับและการรักษาสมดุลของระดับน้ำตาลในเลือดในที่สุด) [ 14 ]
การแสดงภาพของคอมเพล็กซ์ IR แบบเต็มความยาวยังไม่สามารถทำได้เนื่องจากข้อจำกัดหลายประการ การแสดงภาพของคอมเพล็กซ์ IR–อินซูลินแบบเต็มความยาวยังไม่สามารถทำได้เนื่องจากการเชื่อมโยงที่ยืดหยุ่นของโดเมนทรานส์เมมเบรน (TM) กับโดเมนภายนอกเซลล์และโดเมนภายในเซลล์ โดเมนทรานส์เมมเบรน (TM) มีความสำคัญต่อการกระตุ้นและการส่งสัญญาณปลายทาง การทำให้โดเมน TM มีเสถียรภาพอาจเป็นผลมาจากฟอสฟาติดิลอิโนซิทอล ในขณะเดียวกัน การแสดงภาพของโปรตีน IR–ปลายทางแบบเต็มความยาวเป็นเรื่องท้าทายเนื่องจากลักษณะชั่วคราวของการเชื่อมโยง ความต้องการตัวรับฟอสโฟรีเลชัน และการวางแนวสัมพัทธ์ที่ไม่คงที่[ 16 ]
การใช้ cryo-EM และ การจำลอง พลศาสตร์โมเลกุลของตัวรับที่สร้างขึ้นใหม่ในนาโนดิสก์ทำให้สามารถมองเห็นโครงสร้างของเอกโตโดเมนของตัวรับอินซูลินแบบไดเมอร์ทั้งหมดที่มีโมเลกุลอินซูลินสี่โมเลกุลจับอยู่ ซึ่งเป็นการยืนยันและแสดงให้เห็นโดยตรงถึงตำแหน่งการจับ 4 ตำแหน่งที่ทำนายไว้ทางชีวเคมี[ 13 ]
อะโกนิสต์
มีการระบุสารกระตุ้นตัวรับอินซูลินโมเลกุลขนาดเล็กจำนวนหนึ่ง[ 17 ]
เส้นทางการส่งสัญญาณ
ตัวรับอินซูลินเป็น ตัวรับไทโรซีนไคเนสชนิดหนึ่งซึ่งการจับกับลิแกนด์ที่เป็นตัวกระตุ้นจะทำให้เกิดการฟอสโฟรีเลชันตัวเองของหมู่ไทโรซีน โดยแต่ละหน่วยย่อยจะฟอสโฟรีเลตคู่ของมัน การเพิ่มหมู่ฟอสเฟตจะสร้างตำแหน่งการจับสำหรับสารตั้งต้นของตัวรับอินซูลิน (IRS-1) ซึ่งจะถูกกระตุ้นในภายหลังผ่านการฟอสโฟรีเลชัน IRS-1 ที่ถูกกระตุ้นจะเริ่มต้นเส้นทางการส่งสัญญาณและจับกับฟอสโฟอิโนซิไทด์ 3-ไคเนส (PI3K) ซึ่งจะทำให้ PI3K ถูกกระตุ้น จากนั้น PI3K จะเร่งปฏิกิริยาการเปลี่ยนฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟตเป็นฟอสฟาติดิลอิโนซิทอล 3,4,5-ไตรฟอสเฟต ( PIP3 ) PIP 3ทำหน้าที่เป็นตัวส่งสัญญาณรองและกระตุ้นการทำงานของโปรตีนไคเนสที่ขึ้นอยู่กับฟอสฟาติดิลอิโนซิทอล ซึ่งจะกระตุ้นไคเนสอื่นๆ อีกหลายตัว โดยเฉพาะอย่างยิ่งโปรตีนไคเนส B (PKB หรือที่รู้จักกันในชื่อ Akt) PKB กระตุ้นการเคลื่อนย้ายของเวสิเคิลที่มีตัวขนส่งกลูโคส ( GLUT4 ) ไปยังเยื่อหุ้มเซลล์ ผ่านการกระตุ้น โปรตีน SNAREเพื่ออำนวยความสะดวกในการแพร่ของกลูโคสเข้าสู่เซลล์ นอกจากนี้ PKB ยังฟอสโฟรีเลตและยับยั้ง ไกลโคเจน ซินเท สไค เนส ซึ่งเป็นเอนไซม์ที่ยับยั้งไกลโคเจนซินเทสดังนั้น PKB จึงทำหน้าที่เริ่มต้นกระบวนการสร้างไกลโคเจน ซึ่งในที่สุดจะช่วยลดความเข้มข้นของกลูโคสในเลือด[ 18 ]
 ผลของอินซูลินต่อการดูดซึมและการเผาผลาญกลูโคสอินซูลินจับกับตัวรับของมัน (1) ซึ่งในทางกลับกันจะเริ่มต้นกระบวนการกระตุ้นโปรตีนหลายอย่าง (2) ซึ่งรวมถึง: การเคลื่อนย้ายตัวขนส่ง Glut-4 ไปยังเยื่อหุ้มพลาสมาและการไหลเข้าของกลูโคส (3) การสังเคราะห์ไกลโคเจน (4) ไกลโคไลซิส (5) และการสังเคราะห์กรดไขมัน (6)
ผลของอินซูลินต่อการดูดซึมและการเผาผลาญกลูโคสอินซูลินจับกับตัวรับของมัน (1) ซึ่งในทางกลับกันจะเริ่มต้นกระบวนการกระตุ้นโปรตีนหลายอย่าง (2) ซึ่งรวมถึง: การเคลื่อนย้ายตัวขนส่ง Glut-4 ไปยังเยื่อหุ้มพลาสมาและการไหลเข้าของกลูโคส (3) การสังเคราะห์ไกลโคเจน (4) ไกลโคไลซิส (5) และการสังเคราะห์กรดไขมัน (6)


การทำงาน
การควบคุมการแสดงออกของยีน
โปรตีน IRS-1 ที่ถูกกระตุ้นจะทำหน้าที่เป็นตัวส่งสัญญาณรองภายในเซลล์เพื่อกระตุ้นการถอดรหัสของยีนที่ควบคุมโดยอินซูลิน ขั้นแรก โปรตีน Grb2 จะจับกับหมู่ P-Tyr ของ IRS-1 ในโดเมน SH2ของ มัน จากนั้น Grb2จะสามารถจับกับ SOS ซึ่งจะเร่งปฏิกิริยาการแทนที่ GDP ที่จับอยู่กับ GTP บน Ras ซึ่งเป็นโปรตีน Gโปรตีนนี้จะเริ่มกระบวนการฟอสโฟรีเลชันแบบต่อเนื่อง ซึ่งจะสิ้นสุดลงด้วยการกระตุ้นของไมโทเจนแอคติเวเตดโปรตีนไคเนส ( MAPK ) ซึ่งจะเข้าสู่นิวเคลียสและฟอสโฟรีเลตปัจจัยการถอดรหัสในนิวเคลียสต่างๆ (เช่นElk1 )
การกระตุ้นการสังเคราะห์ไกลโคเจน
การสังเคราะห์ไกลโคเจนยังถูกกระตุ้นโดยตัวรับอินซูลินผ่านทาง IRS-1 ในกรณีนี้ โดเมน SH2 ของPI-3 kinase (PI-3K) จะจับกับ P-Tyr ของ IRS-1 เมื่อ PI-3K ถูกกระตุ้นแล้ว มันสามารถเปลี่ยนฟอสฟาติดิลอิโนซิทอล 4,5-บิสฟอสเฟต (PIP2 ) ซึ่งเป็น ลิปิด ในเยื่อหุ้มเซลล์ ให้เป็นฟอสฟาติดิลอิโนซิทอล 3,4,5-ไตรฟอสเฟต (PIP3 )ซึ่งจะกระตุ้นโปรตีนไคเนส PKB ( Akt ) โดยอ้อมผ่านการฟอสฟอริเลชัน จากนั้น PKB จะฟอสฟอริเลตโปรตีนเป้าหมายหลายชนิด รวมถึงไกลโคเจนซินเทสไคเนส 3 (GSK-3) GSK-3 มีหน้าที่ในการฟอสฟอริเลต (และทำให้ไม่ทำงาน) ไกลโคเจนซินเทส เมื่อ GSK-3 ถูกฟอสฟอริเลต มันจะถูกทำให้ไม่ทำงาน และป้องกันไม่ให้ไปทำให้ไกลโคเจนซินเทสไม่ทำงาน ด้วยวิธีการทางอ้อมเช่นนี้ อินซูลินจึงเพิ่มการสังเคราะห์ไกลโคเจน
การสลายตัวของอินซูลิน
เมื่อโมเลกุลอินซูลินจับกับตัวรับและออกฤทธิ์แล้ว อาจถูกปล่อยกลับเข้าสู่สภาพแวดล้อมภายนอกเซลล์หรืออาจถูกย่อยสลายโดยเซลล์ การย่อยสลายโดยปกติเกี่ยวข้องกับการนำสารเชิงซ้อนอินซูลิน-ตัวรับเข้าสู่เซลล์ ตามด้วยการทำงานของเอนไซม์ย่อยสลายอินซูลินโมเลกุลอินซูลินส่วนใหญ่จะถูกย่อยสลายโดย เซลล์ ตับมีการประมาณการว่าโมเลกุลอินซูลินทั่วไปจะถูกย่อยสลายในที่สุดประมาณ 71 นาทีหลังจากถูกปล่อยเข้าสู่ระบบไหลเวียนโลหิตครั้งแรก[ 19 ]
ระบบภูมิคุ้มกัน
นอกจากหน้าที่การเผาผลาญแล้ว ตัวรับอินซูลินยังแสดงออกบนเซลล์ภูมิคุ้มกัน เช่น แมโครฟาจ เซลล์บี และเซลล์ที บนเซลล์ที การแสดงออกของตัวรับอินซูลินจะตรวจไม่พบในสภาวะพัก แต่จะเพิ่มขึ้นเมื่อมี การกระตุ้น ตัวรับเซลล์ที (TCR) อันที่จริงอินซูลินได้รับการพิสูจน์แล้วว่าเมื่อให้จากภายนอกจะส่งเสริม การเพิ่มจำนวนเซลล์ทีในหลอด ทดลองในแบบจำลองสัตว์ การส่งสัญญาณของตัวรับอินซูลินมีความสำคัญต่อการเพิ่มผลกระทบสูงสุดของเซลล์ทีในระหว่างการติดเชื้อเฉียบพลันและการอักเสบ[ 20 ] [ 21 ]
พยาธิวิทยา
กิจกรรมหลักของการกระตุ้นตัวรับอินซูลินคือการกระตุ้นการดูดซึมกลูโคส ด้วยเหตุนี้ "ภาวะดื้อต่ออินซูลิน" หรือการลดลงของการส่งสัญญาณของตัวรับอินซูลิน จึงนำไปสู่โรคเบาหวานชนิดที่ 2 – เซลล์ไม่สามารถดูดซึมกลูโคสได้ ส่งผลให้เกิดภาวะน้ำตาลในเลือดสูง (ระดับกลูโคสในกระแสเลือดสูงขึ้น) และภาวะแทรกซ้อนต่างๆ ที่เกิดจากโรคเบาหวาน
ผู้ป่วยที่มีภาวะดื้อต่ออินซูลินอาจแสดงอาการผิวหนังดำคล้ำ (acanthosis nigricans )
มี ผู้ป่วยจำนวนหนึ่งที่มีการกลายพันธุ์แบบโฮโมไซกัสใน ยีน INSRซึ่งทำให้เกิดกลุ่มอาการโดโนฮิวหรือโรคเลเปรคาวนิ สม์ ความผิด ปกติทางพันธุกรรมแบบออโตโซมัลรีเซสซี ฟนี้ ส่งผลให้ตัวรับอินซูลินไม่ทำงานโดยสิ้นเชิง ผู้ป่วยเหล่านี้มีหูที่อยู่ต่ำกว่าปกติ มักจะโป่งออกมา รูจมูกบาน ริมฝีปากหนา และการเจริญเติบโตช้าอย่างรุนแรง ในกรณีส่วนใหญ่ ผู้ป่วยเหล่านี้มีโอกาสรอดชีวิตต่ำมาก โดยเสียชีวิตภายในปีแรกของชีวิต การกลายพันธุ์อื่นๆ ของยีนเดียวกันนี้ทำให้เกิดกลุ่มอาการแร็บสัน-เมนเดนฮอลล์ซึ่งมีความรุนแรงน้อยกว่า โดยผู้ป่วยจะมีฟันที่ผิดปกติเหงือก บวม และต่อมไพเนียล ขยายใหญ่ขึ้น โรคทั้งสองนี้มีอาการ ระดับ น้ำตาล ในเลือดผันผวน : หลังรับประทานอาหาร ระดับน้ำตาลในเลือดจะสูงมากในตอนแรก แล้วลดลงอย่างรวดเร็วจนต่ำกว่าปกติ[ 22 ]การกลายพันธุ์ทางพันธุกรรมอื่นๆ ของยีนตัวรับอินซูลินอาจทำให้เกิดภาวะดื้อต่ออินซูลินอย่างรุนแรง[ 23 ]
ปฏิสัมพันธ์
มีการแสดงให้เห็นว่าตัวรับอินซูลินมีปฏิสัมพันธ์กับ
อ่านเพิ่มเติม
- Pearson RB, Kemp BE (1991). "[3] ลำดับไซต์ฟอสโฟรีเลชันของโปรตีนไคเนสและโมทีฟความจำเพาะที่เป็นฉันทามติ: ตาราง" ลำดับไซต์ฟอสโฟรีเล ชัน ของ โปรตีนไคเนสและโมทีฟความจำเพาะที่เป็นฉันทามติ: ตารางวิธีการทางเอนไซม์วิทยา เล่มที่ 200 หน้า 62–81 doi : 10.1016/0076-6879(91)00127- I ISBN 9780121821012. PMID 1956339 .
- Joost HG (กุมภาพันธ์ 1995). "ความแตกต่างเชิงโครงสร้างและหน้าที่ของตัวรับอินซูลิน". Cellular Signalling . 7 (2): 85– 91. doi : 10.1016/0898-6568(94)00071-I . PMID 7794689 .
- O'Dell SD, Day IN (กรกฎาคม 1998). "ปัจจัยการเจริญเติบโตคล้ายอินซูลิน II (IGF-II)". วารสารนานาชาติชีวเคมีและชีววิทยาของเซลล์30 (7): 767– 71. doi : 10.1016/S1357-2725(98)00048-X . PMID 9722981 .
- Lopaczynski W (1999). "การควบคุมที่แตกต่างกันของเส้นทางการส่งสัญญาณสำหรับอินซูลินและอินซูลินไลค์โกรทแฟคเตอร์ I" Acta Biochimica Polonica . 46 (1): 51– 60. doi : 10.18388/abp.1999_4183 . PMID 10453981 .
- Sasaoka T, Kobayashi M (สิงหาคม 2000) "ความสำคัญเชิงหน้าที่ของ Shc ในการส่งสัญญาณอินซูลินในฐานะสารตั้งต้นของตัวรับอินซูลิน " วารสารต่อมไร้ท่อ . 47 (4): 373– 81. ดอย : 10.1507 / endocrj.47.373 PMID11075717 .
- Perz M, Torlińska T (2001). "ตัวรับอินซูลิน - ลักษณะโครงสร้างและหน้าที่การทำงาน" Medical Science Monitor . 7 (1): 169– 77. PMID 11208515 .
- Benaim G, Villalobo A (สิงหาคม 2545). "การฟอสฟอริเลชันของแคลโมดูลิน ความหมายเชิงหน้าที่" European Journal of Biochemistry . 269 (15): 3619– 31. doi : 10.1046/j.1432-1033.2002.03038.x . hdl : 10261/79981 . PMID 12153558 .
ลิงก์ภายนอก
- Insulin+receptorที่ US National Library of Medicine Medical Subject Headings (MeSH)
- ภาพรวมของข้อมูลโครงสร้างทั้งหมดที่มีอยู่ในPDBสำหรับUniProt : P06213 (ตัวรับอินซูลิน) ที่PDBe- KB
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ตัวรับอินซูลิน
ตัว รับอินซูลิน ( IR ) เป็น ตัวรับแบบทรานส์เมมเบรน ที่ถูกกระตุ้นโดย อินซูลิน , IGF-I , IGF-II และอยู่ในกลุ่มใหญ่ของ ตัวรับไทโรซีนไคเนส [ 5 ] ใน ทางเมตาบอลิซึม...
โครงสร้าง
ในขั้นต้น การถอดรหัส ของตัวแปรการต่อเชื่อมทางเลือกที่ได้มาจาก ยีน INSR จะ ถูกแปล เป็นโปรตีนโมโนเมอร์ 2 ชนิด ได้แก่ IR-A ซึ่ง ไม่มี เอ็กซอน 11 และ IR-B ซึ่งมีเอ็กซอน 11 การรวมเอ็กซอน 11 ส่งผลให้มีการเพิ่มกรดอะมิโน 12 ตัวก่อนหน้าตำแหน่งการตัดของเอนไซม์ ฟูริน
การจับตัวของลิแกนด์
ลิแกนด์ภายในของตัวรับอินซูลิน ได้แก่ อินซูลิน , IGF-I และ IGF-II การใช้ cryo-EM ทำให้ได้ข้อมูลเชิงลึกเชิงโครงสร้างเกี่ยวกับการเปลี่ยนแปลงรูปร่างเมื่อมีการจับกับอินซูลิน การจับของลิแกนด์กับ α-chain ของ ectodomain แบบไดเมอร์ของ IR...
อะโกนิสต์
มีการระบุสารกระตุ้นตัวรับอินซูลินโมเลกุล ขนาดเล็ก จำนวนหนึ่ง [ 17 ]