อ่าน 16 นาที
ไมโทเจนแอคติเวตโปรตีนไคเนส
โปรตีน ไคเนสที่กระตุ้นด้วยไมโทเจน ( MAPK หรือ MAP kinase ) เป็น โปรตีนไคเนส ชนิดหนึ่ง ที่จำเพาะต่อเซริน/ทรีโอนีน...
ไมโทเจนแอคติเวตโปรตีนไคเนส
| ไมโทเจนแอคติเวตโปรตีนไคเนส | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| ตัวระบุ | |||||||||
| หมายเลข EC | 2.7.11.24 | ||||||||
| หมายเลข CAS | 142243-02-5 | ||||||||
| ฐานข้อมูล | |||||||||
| อินท์เอ็นซ์ | มุมมองของ IntEnz | ||||||||
| เบรนด้า | เบรนด้าเข้าร่วม | ||||||||
| เอ็กซ์แพซี่ | มุมมองของ NiceZyme | ||||||||
| เคกก์ | รายการ KEGG | ||||||||
| เมตาไซค์ | วิถีการเผาผลาญ | ||||||||
| ไพรแอม | ประวัติโดยย่อ | ||||||||
| โครงสร้างPDB | RCSB PDB PDBe PDBsum | ||||||||
| ออนโทโลยีของยีน | อามิโก้ / ควิกโก้ | ||||||||
| |||||||||
โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน ( MAPKหรือMAP kinase ) เป็น โปรตีนไคเนสชนิดหนึ่ง ที่จำเพาะต่อเซริน/ทรีโอนีน ซึ่งมีส่วนเกี่ยวข้องในการควบคุมการตอบสนองของเซลล์ต่อสิ่งเร้าที่หลากหลาย เช่นไมโทเจนความเครียดจากออสโมซิสความร้อนช็อกและไซโตไคน์ที่ก่อให้เกิดการอักเสบ พวกมันควบคุมการทำงานของเซลล์ รวมถึงการเพิ่มจำนวนการแสดงออกของยีนการแบ่งเซลล์ไมโทซิสการอยู่รอดของเซลล์ และอะพอพโทซิส[ 1 ]
MAP kinase พบได้เฉพาะในยูคาริโอตเท่านั้น แต่มีความหลากหลายค่อนข้างมากและพบได้ในสัตว์ทุกชนิด เชื้อรา และพืช รวมถึงแม้แต่ในยูคาริโอตเซลล์เดียวหลายชนิด
MAPK จัดอยู่ในกลุ่มไคเนส CMGC (CDK/MAPK/GSK3/CLK) ญาติสนิทที่สุดของ MAPK คือไคเนสที่ขึ้นอยู่กับไซคลิน (CDK) [ 2 ]
การค้นพบ
โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจนตัวแรกที่ถูกค้นพบคือERK1 (MAPK3) ในสัตว์เลี้ยงลูกด้วยนม เนื่องจาก ERK1 และERK2 (MAPK1) ซึ่งเป็นญาติใกล้ชิดกัน ต่างก็มีส่วนเกี่ยวข้องกับการส่งสัญญาณของปัจจัยการเจริญเติบโต จึงเรียกตระกูลนี้ว่า "โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน" แต่เมื่อมีการค้นพบสมาชิกอื่นๆ แม้กระทั่งจากสิ่งมีชีวิตที่อยู่ห่างไกลกัน (เช่น พืช) ก็ยิ่งชัดเจนขึ้นว่าชื่อนี้ไม่ถูกต้อง เพราะ MAPK ส่วนใหญ่มีส่วนเกี่ยวข้องกับการตอบสนองต่อสิ่งกระตุ้นความเครียดจากสิ่งแวดล้อมที่ไม่ใช่สิ่งมีชีวิต (เช่น ภาวะออสโมซิสสูง ความเครียดจากออกซิเดชัน ความเสียหายของดีเอ็นเอ ภาวะออสโมลาริตีต่ำ การติดเชื้อ ฯลฯ) เนื่องจากพืชไม่สามารถ "หนี" จากความเครียดได้ พืชบกจึงมีจำนวนยีน MAPK ต่อสิ่งมีชีวิตมากที่สุดเท่าที่เคยพบมา ดังนั้นบทบาทของไคเนส ERK1/2 ในสัตว์เลี้ยงลูกด้วยนมในฐานะตัวควบคุมการเพิ่มจำนวนเซลล์จึงไม่ใช่หน้าที่ทั่วไป แต่เป็นหน้าที่เฉพาะทางสูง
ประเภท
MAPK ส่วนใหญ่มีลักษณะร่วมกันหลายประการ เช่น การกระตุ้นขึ้นอยู่กับ การ ฟอสโฟรีเลชัน สองครั้ง โครงสร้างเส้นทางแบบสามระดับ และไซต์การจดจำซับสเตรตที่คล้ายคลึงกัน เหล่านี้คือ MAP kinase แบบ "คลาสสิก" แต่ก็ยังมี MAPK รุ่นเก่าบางส่วนที่อยู่นอกกลุ่มดังที่กล่าวมาข้างต้น ซึ่งไม่มีไซต์การฟอสโฟรีเลชันคู่ มีเพียงเส้นทางแบบสองระดับ และขาดคุณสมบัติที่จำเป็นสำหรับ MAPK อื่นๆ ในการจับกับซับสเตรต โดยทั่วไปแล้ว MAPK เหล่านี้จะถูกเรียกว่า MAPK แบบ "ผิดปกติ" [ 3 ]ยังไม่ชัดเจนว่า MAPK แบบผิดปกติเหล่านี้จัดอยู่ในกลุ่มเดียวกันหรือไม่ เมื่อเทียบกับ MAPK แบบคลาสสิก
ตระกูล MAPK ของเอนไซม์ไคเนสในสัตว์เลี้ยงลูกด้วยนมประกอบด้วยสามตระกูลย่อย:
- เอนไซม์ไคเนสที่ควบคุมสัญญาณภายนอกเซลล์ (ERKs)
- ซี-จุน เอ็น-เทอร์มินัล ไคเนส (JNKs)
- โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน p38 (p38s) [ 4 ] [ 5 ]
โดยทั่วไป ERK จะถูกกระตุ้นโดยปัจจัยการเจริญเติบโตและไมโทเจนในขณะที่ความเครียดของเซลล์และไซโตไคน์ที่ก่อให้เกิดการอักเสบจะกระตุ้น JNK และ p38 [ 4 ]
การเปิดใช้งาน

โปรตีนไคเนสที่ถูกกระตุ้นด้วยไมโทเจนนั้นไม่มีฤทธิ์เร่งปฏิกิริยาในรูปทรงพื้นฐาน เพื่อให้มีฤทธิ์ โปรตีนไคเนสเหล่านี้ต้องการการเติมหมู่ฟอสเฟต (อาจเกิดขึ้นหลายครั้ง) ในวงจรการกระตุ้น กระบวนการนี้ดำเนินการโดยเอนไซม์เฉพาะทางในกลุ่มโปรตีนไคเนส STE ด้วยวิธีนี้พลวัตของโปรตีนสามารถเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างของโปรตีนผ่านกลไกอัลโลสเตอรี ระยะไกล ได้
ในกรณีของ MAP kinase แบบคลาสสิกวงจรการกระตุ้นจะมีโมทีฟ TxY (ทรีโอนีน-x-ไทโรซีน) ที่เป็นลักษณะเฉพาะ (TEY ในERK1และERK2 ของสัตว์เลี้ยงลูกด้วยนม , TDY ในERK5 , TPY ในJNKs , TGY ในp38 kinases ) ซึ่งจำเป็นต้องได้รับการฟอสฟอริเลชันทั้งบนหมู่ทรีโอนีนและไทโรซีนเพื่อล็อกโดเมนของเอนไซม์ให้อยู่ในโครงสร้างที่พร้อมสำหรับการเร่งปฏิกิริยา ทั้งในร่างกาย และในหลอดทดลองการฟอสฟอริเลชันของไทโรซีนมักเกิดขึ้นก่อนการฟอสฟอริเลชันของทรีโอนีน แม้ว่าการฟอสฟอริเลชันของหมู่ใดหมู่หนึ่งสามารถเกิดขึ้นได้โดยปราศจากอีกหมู่หนึ่งก็ตาม
การฟอสฟอริเลชัน ของวงจรการกระตุ้นแบบคู่ขนานนี้(ซึ่งมีการเสนอว่าอาจเป็นแบบกระจายหรือแบบต่อเนื่อง ขึ้นอยู่กับสภาพแวดล้อมของเซลล์) ดำเนินการโดยสมาชิกของตระกูลโปรตีนไคเนส Ste7 หรือที่รู้จักกันในชื่อMAP2 ไคเนส MAP2 ไคเนสเองก็ถูกกระตุ้นด้วยการฟอสฟอริเลชันโดยไคเนสเซริน-ทรีโอนีนต้นน้ำหลายตัว ( MAP3 ไคเนส ) เนื่องจาก MAP2 ไคเนสแสดงกิจกรรมน้อยมากกับสารตั้งต้นอื่นนอกเหนือจาก MAPK ที่เกี่ยวข้อง ดังนั้นวิถี MAPK แบบคลาสสิกจึงก่อตัวเป็นวิถีหลายระดับ แต่ค่อนข้างเป็นเส้นตรง วิถีเหล่านี้สามารถส่งผ่านสิ่งเร้าจากเยื่อหุ้มเซลล์ (ซึ่งมี MAP3K จำนวนมากถูกกระตุ้น) ไปยังนิวเคลียส (ซึ่งมีเพียง MAPK เท่านั้นที่สามารถเข้าไปได้) หรือไปยังเป้าหมายย่อยภายในเซลล์อื่นๆ ได้อย่างมีประสิทธิภาพ
เมื่อเปรียบเทียบกับวิถี MAPK แบบคลาสสิกสามระดับ พบว่า MAP kinase ที่ผิดปกติบางชนิดมีระบบสองระดับที่เก่าแก่กว่าERK3 (MAPK6) และERK4 (MAPK4) เพิ่งได้รับการแสดงให้เห็นว่าถูกฟอสฟอริเลตโดยตรงและถูกกระตุ้นโดยPAK kinase (ที่เกี่ยวข้องกับ MAP3 kinase อื่นๆ) [ 6 ]ในทางตรงกันข้ามกับ MAP kinase แบบคลาสสิก MAPK ที่ผิดปกติเหล่านี้ต้องการเพียงสารตกค้างเดียวในลูปการกระตุ้นเพื่อถูกฟอสฟอริเลต รายละเอียดของ การกระตุ้น NLKและERK7 (MAPK15) ยังคงไม่เป็นที่ทราบแน่ชัด
การยับยั้งการทำงานของ MAPK เกิดขึ้นจาก ฟอสฟาเทสหลายชนิดฟอสฟาเทสเฉพาะกลุ่มที่มีการอนุรักษ์ไว้อย่างดีคือฟอสฟาเทส MAP kinase (MKPs) ซึ่งเป็นกลุ่มย่อยของฟอสฟาเทสแบบจำเพาะคู่ (DUSPs) [ 7 ]ดังที่ชื่อบ่งบอก เอนไซม์เหล่านี้สามารถไฮโดรไลซ์ฟอสเฟตจากทั้งฟอสโฟไทโรซีนและฟอสโฟทรีโอนีนได้ เนื่องจากการกำจัดหมู่ฟอสเฟตใดหมู่หนึ่งจะลดกิจกรรมของ MAPK ลงอย่างมาก ทำให้การส่งสัญญาณหยุดชะงักโดยสิ้นเชิง ดังนั้นฟ อ สฟาเทสไทโรซีน บางชนิด จึงมีส่วนเกี่ยวข้องในการยับยั้งการทำงานของ MAP kinase ด้วย (เช่น ฟอสฟาเทสHePTP , STEPและPTPRRในสัตว์เลี้ยงลูกด้วยนม)
การส่งสัญญาณแบบต่อเนื่อง

ดังที่กล่าวมาข้างต้น MAPK มักจะสร้างเส้นทางหลายระดับ โดยรับอินพุตหลายระดับเหนือ MAP kinase จริง ในทางตรงกันข้ามกับกลไกการกระตุ้นที่ค่อนข้างง่ายซึ่งขึ้นอยู่กับการฟอสโฟรีเลชั่นของ MAPK และMAP2Kนั้น MAP3K มีการควบคุมที่ซับซ้อนอย่างน่าทึ่งMAP3K ที่รู้จักกันดีหลายตัว เช่นc-Raf , MEKK4หรือMLK3ต้องใช้หลายขั้นตอนในการกระตุ้น โดยทั่วไปแล้วเอนไซม์เหล่านี้จะถูกควบคุมแบบอัลโลสเตอริก และถูกล็อกไว้ในสถานะที่ไม่ทำงานโดยกลไกหลายอย่าง ขั้นตอนแรกในการกระตุ้นประกอบด้วยการปลดปล่อยการยับยั้งตัวเองโดยลิแกนด์ขนาดเล็กกว่า (เช่นRasสำหรับc-Raf , GADD45สำหรับMEKK4 [ 10 ]หรือCdc42สำหรับ MLK3 [ 11 ] ) โดยทั่วไป (แต่ไม่เสมอไป) กระบวนการนี้เกิดขึ้นที่เยื่อหุ้มเซลล์ ซึ่งเป็นบริเวณที่ตัวกระตุ้นส่วนใหญ่จับอยู่ (โปรดทราบว่าโปรตีน G ขนาดเล็กจะจับกับเยื่อหุ้มเซลล์อย่างต่อเนื่องเนื่องจากการพรีนิเลชัน ) ขั้นตอนนี้ตามมาด้วยการสร้างโฮโมไดเมอร์และเฮเทอโรไดเมอร์แบบข้างต่อข้างของโดเมนไคเนสที่เข้าถึงได้แล้ว โครงสร้างเชิงซ้อนที่เพิ่งได้รับการกำหนดเมื่อเร็ว ๆ นี้เผยให้เห็นว่าไดเมอร์ก่อตัวขึ้นในทิศทางที่ทำให้บริเวณการจับกับสารตั้งต้นของทั้งสองส่วนเป็นอิสระ[ 12 ]ที่สำคัญ เหตุการณ์การสร้างไดเมอร์นี้ยังบังคับให้โดเมนไคเนส MAP3 ปรับตัวให้มีโครงสร้างที่ทำงานได้บางส่วน กิจกรรมเต็มที่จะเกิดขึ้นได้ก็ต่อเมื่อไดเมอร์เหล่านี้ถ่ายโอนฟอสเฟตให้กันและกันบนลูปการกระตุ้น ขั้นตอนหลังนี้สามารถทำได้หรือได้รับความช่วยเหลือจากโปรตีนไคเนสเสริม (ไคเนส MAP4 สมาชิกของตระกูล Ste20) เมื่อไคเนส MAP3 ทำงานได้อย่างเต็มที่แล้ว มันอาจจะฟอสฟอริเลตไคเนส MAP2 ซึ่งเป็นสารตั้งต้นของมัน และในทางกลับกัน ไคเนส MAP2 ก็จะฟอสฟอริเลตไคเนส MAP ที่เป็นสารตั้งต้นของมันต่อไป
ในสัตว์

วิถีการส่งสัญญาณ ERK1 /2ในสัตว์เลี้ยงลูกด้วยนมเป็นระบบ MAPK ที่ได้รับการศึกษาอย่างละเอียดที่สุด ตัวกระตุ้นต้นน้ำที่สำคัญที่สุดของวิถีการส่งสัญญาณนี้คือโปรตีน Raf ( A-Raf , B-Rafหรือc-Raf ) ซึ่งเป็นตัวกลางสำคัญในการตอบสนองต่อปัจจัยการเจริญเติบโต ( EGF , FGF , PDGFเป็นต้น) แต่ MAP3K อื่นๆ เช่น c-Mos และTpl2/Cotก็สามารถมีบทบาทเดียวกันได้เช่นกัน เอนไซม์เหล่านี้ทั้งหมดจะทำการฟอสฟอริเลตและกระตุ้น ไคเนส MKK1และ/หรือMKK2ซึ่งเป็นตัวกระตุ้นที่มีความจำเพาะสูงสำหรับERK1และERK2 ไคเนสเหล่านี้จะทำการ ฟอสฟอริเลตสารตั้งต้นจำนวนมากที่มีความสำคัญต่อการ เพิ่มจำนวน เซลล์ การดำเนินไปของวงจรเซลล์การแบ่งเซลล์ และการเปลี่ยนแปลงรูปร่างของเซลล์ ( ไคเนส RSK , ปัจจัยการถอดรหัส Elk-1 เป็นต้น)
ตรงกันข้ามกับเส้นทาง ERK1/2 ที่ค่อนข้างแยกตัวออกจากกัน ไคเน ส p38และJNK ของสัตว์เลี้ยงลูกด้วยนมมีตัวกระตุ้นส่วนใหญ่ร่วมกันที่ระดับ MAP3K ( MEKK1 , MEKK4 , ASK1 , TAK1 , MLK3 , TAOK1เป็นต้น) นอกจากนี้ เอนไซม์ MAP2K บางตัวอาจกระตุ้นทั้ง p38 และ JNK ( MKK4 ) ในขณะที่บางตัวมีความจำเพาะต่อ JNK ( MKK7 ) หรือ p38 ( MKK3และMKK6 ) มากกว่าเนื่องจากกลไกการเชื่อมโยงเหล่านี้ จึงมีสิ่งกระตุ้นน้อยมากหรือแทบไม่มีเลยที่สามารถกระตุ้นการทำงานของ JNK โดยไม่กระตุ้น p38 หรือในทางกลับกันพร้อมกัน[ 13 ]ทั้งเส้นทางการส่งสัญญาณ JNK และ p38 ตอบสนองต่อสิ่งกระตุ้นความเครียด เช่นไซโตไคน์การฉายรังสีอัลตราไวโอเลตความร้อนช็อกและออสโมติกช็อกและเกี่ยวข้องกับการปรับตัวต่อความเครียดอะพอพโทซิสหรือการแบ่งเซลล์เอนไซม์ JNK มีสารตั้งต้นเฉพาะจำนวนหนึ่งที่พวกมันเท่านั้นที่สามารถฟอสฟอริเลตได้ (เช่นc-Jun , NFAT4เป็นต้น) ในขณะที่เอนไซม์ p38 ก็มีเป้าหมายเฉพาะบางอย่างเช่นกัน (เช่น ไคเนส MAPKAP MK2และMK3 ) ทำให้มั่นใจได้ว่าจำเป็นต้องมีทั้งสองเอนไซม์เพื่อตอบสนองต่อสิ่งเร้าที่ก่อให้เกิดความเครียด
ERK5เป็นส่วนหนึ่งของวิถีทางที่แยกออกจากกันอย่างชัดเจนในสัตว์เลี้ยงลูกด้วยนม ตัวกระตุ้นต้นน้ำเฉพาะตัวเดียวของมัน คือ MKK5จะถูกเปิดใช้งานเพื่อตอบสนองต่อไคเนส MAP3 MEKK2และMEKK3ความจำเพาะของการโต้ตอบเหล่านี้เกิดจากโครงสร้างที่เป็นเอกลักษณ์ของ MKK5 และ MEKK2/3 ซึ่งทั้งคู่มีโดเมน PB1 ที่ปลาย N ทำให้เกิดการสร้างเฮเทอโรไดเมอร์โดยตรงระหว่างกัน[ 14 ]โดเมน PB1 ของ MKK5 ยังมีส่วนร่วมในการโต้ตอบระหว่าง ERK5-MKK5 ด้วย โดยให้ส่วนต่อประสานพิเศษ (นอกเหนือจากโมทีฟ Dที่พบใน MKK5) ซึ่ง MKK5 สามารถจดจำสารตั้งต้น ERK5 ได้อย่างจำเพาะเจาะจง[ 15 ]แม้ว่ารายละเอียดในระดับโมเลกุลจะยังไม่เป็นที่รู้จักดีนัก แต่ MEKK2 และ MEKK3 ตอบสนองต่อสัญญาณการพัฒนาบางอย่างเพื่อชี้นำ การสร้าง เอนโดเทลและการ สร้าง รูปร่างของหัวใจแม้ว่าจะเกี่ยวข้องกับการพัฒนาสมองเช่นกัน แต่การเสียชีวิตของตัวอ่อนจากการปิดใช้งาน ERK5 อันเนื่องมาจากความผิดปกติของหัวใจเน้นย้ำถึงบทบาทสำคัญของมันในการสร้างหลอดเลือด ในสัตว์เลี้ยงลูกด้วย นม[ 16 ]เป็นที่น่าสังเกตว่าการกำจัด ERK5 แบบมีเงื่อนไขในสัตว์โตเต็มวัยก็เป็นอันตรายถึงชีวิตเช่นกัน เนื่องจากการหยุดชะงักอย่างกว้างขวางของอุปสรรค ของ เยื่อบุผนังหลอดเลือด[ 17 ]เชื่อกันว่าการกลายพันธุ์ในส่วนประกอบต้นน้ำของเส้นทาง ERK5 (คอมเพล็กซ์ CCM) เป็นสาเหตุของความผิดปกติของหลอดเลือดในสมองในมนุษย์
ในเชื้อรา
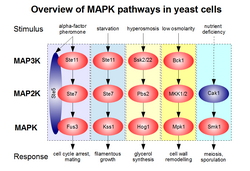
วิถีการส่งสัญญาณ MAPK ของเชื้อราก็ได้รับการศึกษามาอย่างดีเช่นกัน ในยีสต์ MAPK Fus3 มีหน้าที่ควบคุมการหยุดวงจรเซลล์และการผสมพันธุ์เพื่อตอบสนองต่อการกระตุ้นด้วยฟีโรโมน ฟีโรโมนอัลฟาแฟคเตอร์จะถูกตรวจจับโดยตัวรับแบบเจ็ดทรานส์เมมเบ รน การดึงดูดและการกระตุ้นส่วนประกอบของวิถีการส่งสัญญาณ Fus3 ขึ้นอยู่กับ การกระตุ้น โปรตีน G แบบเฮเทอโรไตรเม อริกอย่างเคร่งครัด วิถีการส่งสัญญาณ MAPK สำหรับการผสมพันธุ์ประกอบด้วยสามระดับ (Ste11-Ste7-Fus3) แต่ไคเนส MAP2 และ MAP3 นั้นใช้ร่วมกับวิถีการส่งสัญญาณอื่น คือ Kss1 หรือวิถีการเจริญเติบโตแบบเส้นใย แม้ว่า Fus3 และ Kss1 จะเป็นไคเนสชนิด ERK ที่มีความสัมพันธ์ใกล้ชิดกัน แต่เซลล์ยีสต์ก็ยังสามารถกระตุ้นพวกมันแยกกันได้ โดยอาศัยโปรตีนโครงสร้าง Ste5 ที่ถูกดึงดูดโดยโปรตีน G ของวิถีการส่งสัญญาณสำหรับการผสมพันธุ์อย่างเลือกสรร เคล็ดลับคือ Ste5 สามารถเชื่อมโยงและ "ปลดล็อก" Fus3 สำหรับ Ste7 ในฐานะสารตั้งต้นในคอมเพล็กซ์ตติยภูมิ ในขณะที่มันไม่ทำเช่นเดียวกันกับ Kss1 ทำให้เส้นทางการเติบโตของเส้นใยถูกเปิดใช้งานเฉพาะในกรณีที่ไม่มีการดึงดูด Ste5 [ 18 ]
เชื้อรายังมีวิถีการส่งสัญญาณที่คล้ายคลึงกับการส่งสัญญาณ JNK/p38 ของสัตว์เลี้ยงลูกด้วยนม วิถีนี้คือวิถี Hog1 ซึ่งถูกกระตุ้นโดยความเข้มข้นของสารละลายสูง (ในSaccharomyces cerevisiae ) หรือความเครียดจากปัจจัยทางชีวภาพอื่นๆ (ในSchizosaccharomyces pombe ) ไคเนส MAP2 ของวิถีนี้เรียกว่า Pbs2 (เกี่ยวข้องกับ MKK3/4/6/7 ของสัตว์เลี้ยงลูกด้วยนม) และไคเนส MAP3 ที่เกี่ยวข้องกับการกระตุ้นโดยเฉพาะคือ Ssk2 และ SSk22 ระบบในS. cerevisiaeถูกกระตุ้นโดยโมดูลตรวจจับความเข้มข้นของสารละลายที่ซับซ้อนซึ่งประกอบด้วยโปรตีน Sho1 และ Sln1 แต่ยังไม่ชัดเจนว่าสิ่งกระตุ้นอื่นๆ สามารถกระตุ้นการทำงานของ Hog1 ได้อย่างไร ยีสต์ยังแสดงวิถี MAPK อื่นๆ อีกหลายวิถีที่ไม่มีโฮโมล็อกใกล้เคียงในสัตว์ เช่น วิถีความสมบูรณ์ของผนังเซลล์ (Mpk1/Slt2) หรือ วิถี การสร้างสปอร์ (Smk1) [ 19 ]
ในพืช
แม้จะมีจำนวนยีน MAPK สูง แต่เส้นทาง MAPK ของพืชชั้นสูงกลับได้รับการศึกษาน้อยกว่าในสัตว์หรือเชื้อรา แม้ว่าการส่งสัญญาณจะดูซับซ้อนมาก แต่ไคเนส MPK3, MPK4 และ MPK6 ของArabidopsis thalianaเป็นตัวกลางสำคัญในการตอบสนองต่อภาวะช็อกจากออสโม ซิ สความเครียดจากออกซิเดชันการตอบสนองต่อความเย็น และเกี่ยวข้องกับการตอบสนองต่อเชื้อโรค[ 20 ] [ 21 ] [ 22 ]แบบจำลองภูมิคุ้มกันที่ควบคุมโดย MAPK ของAsai et al. 2002 ส่งสัญญาณการรับรู้ เอฟเฟกต์จากFLS2 ⇨ MEKK1 ⇨ MKK4หรือMKK5 ⇨ MPK3 และ MPK6 ⇨ WRKY22 หรือ WRKY29 [ 22 ]อย่างไรก็ตาม งานของ Mészáros et al. 2006 และ Suarez-Rodriguez et al.ในปี 2007 มีคำสั่งอื่นสำหรับเส้นทางนี้ และเป็นไปได้ว่าเส้นทางเหล่านี้เป็นเส้นทางคู่ขนานที่ทำงานพร้อมกัน[ 22 ]พวกมันยังมีส่วนเกี่ยวข้องกับการสร้างรูปร่างเนื่องจากตัวกลายพันธุ์ MPK4 แสดง อาการ แคระแกร็น อย่างรุนแรง [ 23 ]
ความสัมพันธ์เชิงวิวัฒนาการ

สมาชิกของตระกูล MAPK สามารถพบได้ในสิ่งมีชีวิตยูคาริโอตทุกชนิดที่ได้รับการตรวจสอบมาจนถึงปัจจุบัน โดยเฉพาะอย่างยิ่ง MAP kinase ทั้งแบบคลาสสิกและแบบผิดปกติสามารถสืบย้อนกลับไปถึงรากเหง้าของการวิวัฒนาการของกลุ่มยูคาริโอตหลัก พืชบกมี MAPK แบบคลาสสิกสี่กลุ่ม (MAPK-A, MAPK-B, MAPK-C และ MAPK-D) ซึ่งเกี่ยวข้องกับการตอบสนองต่อความเครียดจากสิ่งแวดล้อมมากมาย[ 25 ]อย่างไรก็ตาม ไม่มีกลุ่มใดที่สามารถเทียบเคียงได้โดยตรงกับกลุ่มของ MAPK แบบคลาสสิกที่พบในโอพิสโทคอนต์ (เชื้อราและสัตว์) ในกลุ่มหลังนี้ กลุ่มย่อยหลักของ MAPK แบบคลาสสิกก่อตัวเป็นสาขาคล้าย ERK/Fus3 (ซึ่งแบ่งย่อยเพิ่มเติมในเมตาโซแอนออกเป็นกลุ่มย่อย ERK1/2 และ ERK5) และไคเนสคล้าย p38/Hog1 (ซึ่งแยกออกเป็นกลุ่มย่อย p38 และ JNK ในสัตว์หลายเซลล์) [ 26 ]นอกจากนี้ ยังมี MAPK หลายชนิดในทั้งเชื้อราและสัตว์ ซึ่งต้นกำเนิดไม่ชัดเจนนัก อาจเนื่องมาจากความแตกต่างสูง (เช่น NLK) หรืออาจเป็นสาขาแรกเริ่มของตระกูล MAPK ทั้งหมด (ERK3, ERK4, ERK7) ในสัตว์มีกระดูกสันหลัง เนื่องจากการจำลองจีโนมทั้งหมดสองครั้งหลังจากการแยกตัวของเซฟาโลคอร์เดต/สัตว์มีกระดูกสันหลัง[ 27 ]จึงมีพาราโลกหลายตัวในแต่ละกลุ่ม ดังนั้น ERK1 และ ERK2 จึงสอดคล้องกับไคเนส rolled ของ Drosophila ส่วน JNK1, JNK2 และ JNK3 ล้วนเป็นออร์โธโลกั สกับยีนbasketในDrosophilaแม้ว่าในกลุ่ม p38 นั้น p38 อัลฟาและเบตาจะเป็นคู่พาราโลกัสอย่างชัดเจน และ p38 แกมมาและเดลต้าก็เช่นกันในสัตว์มีกระดูกสันหลัง แต่จังหวะเวลาของการแยกฐานนั้นไม่ชัดเจนนัก เนื่องจากสัตว์หลายเซลล์หลายชนิดมีโฮโมล็อก p38 หลายตัวอยู่แล้ว (มีไคเนสชนิด p38 สามตัวในDrosophilaได้แก่Mpk2 ( p38a ), p38bและp38c ) โปรตีน ERK5 ตัวเดียวดูเหมือนจะทำหน้าที่เฉพาะทางมาก (จำเป็นต่อการพัฒนาหลอดเลือดในสัตว์มีกระดูกสันหลัง) ไม่ว่าจะอยู่ที่ใดก็ตาม สายพันธุ์นี้ถูกลบออกไปในโปรโตสโตมพร้อมกับส่วนประกอบของเส้นทางต้นน้ำ (MEKK2/3, MKK5) แม้ว่าจะพบได้อย่างชัดเจนในไนดาเรียนฟองน้ำและแม้แต่ในสิ่งมีชีวิตเซลล์เดียวบางชนิด (เช่นโคอาโนแฟลเจลเลตMonosiga brevicollis ) ที่เกี่ยวข้องอย่างใกล้ชิดกับต้นกำเนิดของสัตว์หลายเซลล์[ 28 ]
การแยกตัวระหว่าง MAP kinase แบบคลาสสิกและแบบผิดปกติบางชนิดเกิดขึ้นค่อนข้างเร็ว สิ่งนี้ได้รับการเสนอแนะไม่เพียงแต่จากความแตกต่างสูงระหว่างยีนที่มีอยู่ แต่ยังรวมถึงการค้นพบ MAPK แบบผิดปกติในยูคาริโอตดั้งเดิมและพื้นฐานเมื่อไม่นานมานี้ การจัดลำดับจีโนมของGiardia lambliaเผยให้เห็นการมีอยู่ของยีน MAPK สองยีน โดยยีนหนึ่งคล้ายกับ MAPK ของสัตว์เลี้ยงลูกด้วยนมที่รู้จักกันดีอยู่แล้ว (ERK, p38 เป็นต้น) ส่วนอีกยีนหนึ่งแสดงความคล้ายคลึงกับโปรตีน ERK7 ของสัตว์เลี้ยงลูกด้วยนม[ 29 ]สถานการณ์คล้ายกันในอะมีบาหลายเซลล์Dictyostelium discoideumซึ่งโปรตีน ddERK1 ดูเหมือนจะเป็น MAPK แบบคลาสสิก ในขณะที่ ddERK2 คล้ายกับโปรตีน ERK7 และ ERK3/4 ของเรามากกว่า[ 30 ] MAPK แบบผิดปกติยังสามารถพบได้ในพืชชั้นสูง แม้ว่าจะยังไม่เป็นที่รู้จักดีนัก เช่นเดียวกับสถานการณ์ในสัตว์เลี้ยงลูกด้วยนม ลักษณะส่วนใหญ่ของ MAPK ที่ผิดปกติยังไม่ได้รับการศึกษาอย่างละเอียด เนื่องจากขาดการวิจัยที่มุ่งเน้นในด้านนี้
การรับรู้พื้นผิวและพันธมิตร

โดยทั่วไปแล้ว ไคเนสกลุ่ม CMGC จะมีลำดับคอนเซนซัสที่หลวมมากสำหรับสารตั้งต้นที่ ตำแหน่งเร่งปฏิกิริยาของ MAP ไคเนส เช่นเดียวกับญาติทั้งหมด พวกมันต้องการเพียง กรดอะมิโน เซริน / ทรีโอนีน เป้าหมาย ตามด้วยกรดอะมิโนขนาดเล็ก โดยเฉพาะอย่างยิ่งโพรลีน ("ไคเนสที่กำกับโดยโพรลีน") แต่เนื่องจากตำแหน่ง SP/TP พบได้ทั่วไปในโปรตีนทั้งหมด กลไกการจดจำสารตั้งต้นเพิ่มเติมจึงได้รับการพัฒนาขึ้นเพื่อให้มั่นใจถึงความแม่นยำในการส่งสัญญาณ[ 31 ]แตกต่างจากญาติที่ใกล้เคียงที่สุดอย่างไคเนสที่ขึ้นอยู่กับไซคลิน (CDK) ซึ่งสารตั้งต้นจะถูกจดจำโดย หน่วยย่อย ไซคลิน MAPK จะเชื่อมโยงกับสารตั้งต้นผ่านบริเวณการจับเสริมบนโดเมนไคเนส บริเวณที่สำคัญที่สุดดังกล่าวประกอบด้วยร่องเชื่อมต่อแบบไฮโดรโฟบิกและบริเวณ CD ที่มีประจุลบ ร่วมกันพวกมันจะจดจำสิ่งที่เรียกว่า MAPK docking หรือ D-motifs (เรียกอีกอย่างว่า kinase interaction motif / KIM) โดยพื้นฐานแล้ว D-motif ประกอบด้วยกรดอะมิโนที่มีประจุบวกหนึ่งหรือสองตัว ตามด้วยสารตกค้างที่ไม่ชอบน้ำสลับกัน (ส่วนใหญ่เป็นลิวซีน) โดยทั่วไปจะอยู่เหนือไซต์ฟอสโฟรีเลชันประมาณ 10–50 กรดอะมิโน[ 32 ]สารตั้งต้นของ MAPK ที่รู้จักหลายชนิดมี D-motif ดังกล่าว ซึ่งไม่เพียงแต่สามารถจับกับ MAPK บางชนิดได้เท่านั้น แต่ยังให้การจดจำที่เฉพาะเจาะจงอีกด้วย D-motif ไม่ได้จำกัดอยู่เฉพาะสารตั้งต้นเท่านั้น ไคเนส MAP2 ก็มี motif ดังกล่าวที่ปลายN-terminusซึ่งจำเป็นอย่างยิ่งสำหรับการโต้ตอบระหว่าง MAP2K-MAPK และการกระตุ้น MAPK [ 33 ]ในทำนองเดียวกัน ทั้งฟอสฟาเทส MAP ไคเนสแบบจำเพาะคู่และฟอสฟาเทสไทโรซีนที่จำเพาะต่อ MAP จะจับกับ MAP ไคเนสผ่านทางไซต์เชื่อมต่อเดียวกัน[ 34 ] [ 35 ] D-motif ยังสามารถพบได้ในตัวควบคุมเส้นทาง MAPK และโครงสร้างบางอย่าง (เช่น ในโปรตีน JIP ของสัตว์เลี้ยงลูกด้วยนม)
นอกจากนี้ยังมีไซต์จับซับสเตรตอื่นๆ ที่ยังไม่ได้รับการระบุลักษณะอย่างชัดเจน ไซต์หนึ่งดังกล่าว (ไซต์ DEF) เกิดจากลูปการกระตุ้น (เมื่ออยู่ในโครงสร้างที่ทำงานอยู่) และส่วนแทรกเฉพาะของ MAP kinase ที่อยู่ด้านล่าง ไซต์นี้สามารถรองรับเปปไทด์ที่มีลำดับคอนเซนซัส FxFP ซึ่งโดยทั่วไปจะอยู่ถัดจากไซต์ฟอสโฟรีเลชัน[ 36 ]โปรดทราบว่าไซต์หลังนี้สามารถพบได้เฉพาะในโปรตีนที่จำเป็นต้องจดจำ MAP kinase ที่ทำงานอยู่เท่านั้น ดังนั้นจึงพบได้เกือบทั้งหมดในซับสเตรต โมทีฟที่แตกต่างกันอาจทำงานร่วมกันได้ เช่นในตระกูลปัจจัยการถอดรหัส Elk ซึ่งมีทั้งโมทีฟ D และโมทีฟ FxFP การมีอยู่ของโมทีฟ FxFP ในโปรตีนโครงร่าง KSR1 ยังทำหน้าที่ทำให้มันเป็นซับสเตรตของ ERK1/2 ซึ่งเป็นกลไกป้อนกลับเชิงลบเพื่อกำหนดความแรงที่ถูกต้องของการกระตุ้น ERK1/2
โปรตีนโครงสร้าง
นับตั้งแต่การค้นพบ Ste5 ในยีสต์ นักวิทยาศาสตร์ได้ออกค้นหาโปรตีนที่ทำหน้าที่เป็นโครงสร้างรองรับแบบไม่ใช้เอนไซม์ที่คล้ายคลึงกันในสัตว์เลี้ยงลูกด้วยนม ที่จริงแล้วมีโปรตีนจำนวนหนึ่งที่เกี่ยวข้องกับการส่งสัญญาณ ERK ซึ่งสามารถจับกับองค์ประกอบหลายอย่างในเส้นทางการส่งสัญญาณได้ เช่น MP1จับกับทั้ง MKK1/2 และ ERK1/2, KSR1และKSR2สามารถจับกับ B-Raf หรือ c-Raf, MKK1/2 และ ERK1/2 นอกจากนี้ยังมีการค้นพบโปรตีนที่คล้ายคลึงกันในเส้นทางการส่งสัญญาณ JNK ด้วย เช่น โปรตีนในตระกูล JIP1 / JIP2 และJIP3 /JIP4 ซึ่งแสดงให้เห็นว่าสามารถจับกับ MLK, MKK7 และไคเนส JNK ใดๆ ก็ได้ แต่โชคไม่ดีที่กลไกการควบคุมการกระตุ้น MAPK ของโปรตีนเหล่านี้ยังไม่เป็นที่เข้าใจอย่างถ่องแท้ ต่างจาก Ste5 ในยีสต์ ในขณะที่ Ste5 สร้างสารประกอบเชิงซ้อนสามส่วนกับ Ste7 และ Fus3 เพื่อส่งเสริมการฟอสฟอริเลชันของ Fus3 แต่โปรตีนโครงสร้างรองรับที่รู้จักกันในสัตว์เลี้ยงลูกด้วยนมดูเหมือนจะทำงานด้วยกลไกที่แตกต่างกันมาก ตัวอย่างเช่น KSR1 และ KSR2 เป็นไคเนส MAP3 และเกี่ยวข้องกับโปรตีน Raf [ 37 ]แม้ว่า KSR เพียงอย่างเดียวจะแสดงกิจกรรมไคเนส MAP3 เพียงเล็กน้อย แต่โปรตีน KSR ก็ยังสามารถมีส่วนร่วมในการกระตุ้นไคเนส Raf ได้โดยการสร้างเฮเทอโรไดเมอร์แบบข้างต่อข้างกับไคเนสเหล่านั้น ทำให้เกิดคู่แบบอัลโลสเตอริกเพื่อเปิดใช้งานเอนไซม์แต่ละตัว[ 38 ] ในทางกลับกัน JIPs เห็นได้ชัดว่าเป็นโปรตีนขนส่ง ซึ่งรับผิดชอบในการเพิ่มความเข้มข้นของส่วนประกอบการส่งสัญญาณ MAPK ในบางส่วนของเซลล์ที่มีขั้ว[ 39 ]ในบริบทนี้ การฟอสโฟรีเลชันของ JIP1 (และอาจรวมถึง JIP2) ที่ขึ้นอยู่กับ JNK จะให้สัญญาณแก่ JIPs เพื่อปล่อยส่วนประกอบเส้นทางต้นน้ำที่จับกับ JIP และไม่ทำงาน ทำให้เกิดวงจรป้อนกลับเชิงบวกเฉพาะที่ที่แข็งแกร่ง[ 40 ]กลไกที่ซับซ้อนนี้เชื่อมโยงการขนส่งที่ขึ้นอยู่กับไคเนซินกับการกระตุ้น JNK เฉพาะที่ ไม่เพียงแต่ในสัตว์เลี้ยงลูกด้วยนมเท่านั้น แต่ยังรวมถึงในแมลงวันผลไม้Drosophila melanogasterด้วย[ 41 ]
ในฐานะเป้าหมายในการรักษา
เนื่องจากวิถีการส่งสัญญาณ ERKมีส่วนเกี่ยวข้องกับการเพิ่มจำนวนเซลล์ทั้งในสภาวะปกติและสภาวะผิดปกติ จึงเป็นเรื่องปกติที่สารยับยั้ง ERK1/2 จะเป็นกลุ่ม ยา ต้านมะเร็ง ที่น่าสนใจ อันที่จริง การกลายพันธุ์ของโปรโตออนโคจีนิก "ตัวขับเคลื่อน" หลายชนิดเชื่อมโยงกับการส่งสัญญาณ ERK1/2 เช่น ตัวรับไทโรซีนไคเนส ที่ทำงานอย่างต่อเนื่อง (กลายพันธุ์) โปรตีน RasหรือRafแม้ว่าจะไม่มีการพัฒนาสารยับยั้ง MKK1/2 หรือ ERK1/2 สำหรับใช้ในทางคลินิก แต่สารยับยั้งไคเนสที่ยับยั้งไคเนส Raf ด้วย( เช่น Sorafenib )ประสบความสำเร็จในการเป็นยาต้านมะเร็งหลายชนิด[ 42 ] [ 43 ] มีการศึกษาโคบิเมทิ นิบซึ่งเป็นสารยับยั้ง MEK ในแบบจำลองมะเร็งปอดก่อนการทดลองทางคลินิกโดยใช้ร่วมกับการยับยั้งวิถี PI3Kซึ่งยาทั้งสองชนิดนำไปสู่การตอบสนองแบบเสริมฤทธิ์กัน[ 44 ] [ 45 ]
ไคเนส JNKมีส่วนเกี่ยวข้องกับการพัฒนาภาวะดื้อต่ออินซูลินในบุคคลที่เป็นโรคอ้วน[ 46 ]รวมถึงความเป็นพิษต่อ สารสื่อประสาท หลังภาวะขาดเลือด การยับยั้ง JNK1 ช่วยบรรเทาภาวะดื้อต่ออินซูลินในแบบจำลองสัตว์บางชนิด หนูที่ได้รับการดัดแปลงพันธุกรรมให้ขาดจีน JNK3 ที่ทำงานได้ ซึ่งเป็นไอโซฟอร์มหลักในสมอง แสดงให้เห็นถึงความทนทานต่อภาวะขาดเลือดและการฟื้นตัวจากโรคหลอดเลือดสมองที่ดีขึ้น[ 47 ]แม้ว่าสารยับยั้ง JNK โมเลกุลขนาดเล็กจะอยู่ระหว่างการพัฒนา แต่ยังไม่มีตัวใดที่พิสูจน์ได้ว่ามีประสิทธิภาพในการทดสอบในมนุษย์ สารยับยั้ง JNK ที่ใช้เปปไทด์ (AM-111 ซึ่งเป็นเปปไทด์แบบย้อนกลับ D-motif จาก JIP1 ซึ่งเดิมรู้จักกันในชื่อ XG-102) ก็อยู่ระหว่างการพัฒนาทางคลินิกสำหรับการสูญเสียการได้ยินแบบประสาทรับรู้เช่น กัน [ 48 ]
เดิมทีเชื่อกันว่า p38เป็นเป้าหมายที่สมบูรณ์แบบสำหรับยาต้านการอักเสบ แต่ความล้มเหลวของสารประกอบที่แตกต่างกันทางเคมีมากกว่าสิบชนิดในระยะคลินิกแสดงให้เห็นว่าไคเนส p38 อาจไม่ใช่เป้าหมายการรักษาที่ดีในโรคภูมิต้านตนเองสารประกอบเหล่านี้หลายชนิดพบว่าเป็นพิษต่อตับในระดับต่างๆ และเกิดภาวะดื้อต่อฤทธิ์ต้านการอักเสบภายในไม่กี่สัปดาห์[ 49 ]แนวทางอื่นคือการประเมินศักยภาพในการกำหนดเป้าหมาย MAPK ต้นน้ำ เช่นASK1 [ 50 ]การศึกษาในแบบจำลองสัตว์ของโรคข้ออักเสบอักเสบให้ผลลัพธ์ที่น่าหวัง และเมื่อเร็วๆ นี้พบว่า ASK1 มีความพิเศษในบรรดา MAPK อื่นๆ ตรงที่สามารถถูกกระตุ้นได้ด้วยไซโตไคน์ที่ก่อให้เกิดการอักเสบ เช่นTNF - α [ 50 ]
ดูเพิ่มเติม
- การส่งสัญญาณ
- MAP ไคเนส ไคเนส
- MAP ไคเนส ไคเนส ไคเนส
- MAP ไคเนส ไคเนส ไคเนส ไคเนส
- MAPK1 (ERK2)
- MAPK3 (ERK1)
- MAPK7 (ERK5)
- MAPK8 (JNK1)
- MAPK9 (JNK2)
- MAPK10 (JNK3)
- MAPK11 (p38-เบต้า)
- MAPK12 (p38-gamma)
- MAPK13 (p38-delta)
- MAPK14 (p38-alpha)
- MAPK4 (ERK4: MAPK ที่ผิดปกติ)
- MAPK6 (ERK3: MAPK ที่ผิดปกติ)
- MAPK15 (ERK7/ERK8: MAPK ที่ผิดปกติ)
- NLK (Nemo-like kinase: atypical MAPK)
- ไคเนส ERK1/2
- เส้นทาง ERK1/2
- ไคเนส JNK
- พี38 เมพีไคเนส
- MEKK2 (MAP3K2)
- ASK1 (MAP3K5)
- เครือข่าย MAPK
ลิงก์ภายนอก
- แหล่ง ข้อมูลMAP Kinase
- ตารางรายชื่อเอนไซม์ไคเนสที่ถูกกระตุ้นด้วยไมโทเจน
- ภาพลำดับชั้นของ MAPK
- Mitogen-Activated+Protein+Kinases ที่ หัวข้อทางการ แพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- แบบจำลองความไวสูงพิเศษของ MAPK ในฐานข้อมูล BioModels
- แมลงหวี่กลิ้งตัว – แมลงหวี่แบบโต้ตอบได้
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ไมโทเจนแอคติเวตโปรตีนไคเนส
โปรตีน ไคเนสที่กระตุ้นด้วยไมโทเจน ( MAPK หรือ MAP kinase ) เป็น โปรตีนไคเนส ชนิดหนึ่ง ที่จำเพาะต่อเซริน/ทรีโอนีน...
การค้นพบ
โปรตีนไคเนสที่กระตุ้นด้วยไมโทเจนตัวแรกที่ถูกค้นพบคือ ERK1 (MAPK3) ในสัตว์เลี้ยงลูกด้วยนม เนื่องจาก ERK1 และ ERK2 (MAPK1) ซึ่งเป็นญาติใกล้ชิดกัน ต่างก็มีส่วนเกี่ยวข้องกับการส่งสัญญาณของปัจจัยการเจริญเติบโต จึงเรียกตระกูลนี้ว่า...
ประเภท
MAPK ส่วนใหญ่มีลักษณะร่วมกันหลายประการ เช่น การกระตุ้นขึ้นอยู่กับ การ ฟอสโฟรีเลชัน สองครั้ง โครงสร้างเส้นทางแบบสามระดับ และไซต์การจดจำซับสเตรตที่คล้ายคลึงกัน เหล่านี้คือ MAP kinase แบบ "คลาสสิก" แต่ก็ยังมี MAPK...
การเปิดใช้งาน
โปรตีนไคเนสที่ถูกกระตุ้นด้วยไมโทเจนนั้นไม่มีฤทธิ์เร่งปฏิกิริยาในรูปทรงพื้นฐาน เพื่อให้มีฤทธิ์ โปรตีนไคเนสเหล่านี้ต้องการการเติมหมู่ฟอสเฟต (อาจเกิดขึ้นหลายครั้ง) ในวงจรการกระตุ้น กระบวนการนี้ดำเนินการโดยเอนไซม์เฉพาะทางในกลุ่มโปรตีนไคเนส STE ด้วยวิธีนี้...