อ่าน 12 นาที
ปาก
ในทางพฤกษศาสตร์ปากใบ ( พหูพจน์: stomata มาจาก ภาษา กรีกστόμαแปลว่า "ปาก") หรือเรียกว่าปากใบ หลายรู ( พหูพจน์ : stomates ) คือรูที่พบในชั้นผิวของใบ ลำต้น และอวัยวะอื่นๆ...
ปาก
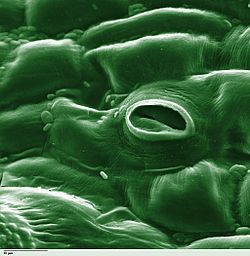


ในทางพฤกษศาสตร์ปากใบ ( พหูพจน์: stomata มาจาก ภาษา กรีกστόμαแปลว่า "ปาก") หรือเรียกว่าปากใบ หลายรู ( พหูพจน์ : stomates ) คือรูที่พบในชั้นผิวของใบ ลำต้น และอวัยวะอื่นๆ ซึ่งควบคุมอัตราการแลกเปลี่ยนก๊าซ ระหว่างช่องว่างอากาศภายในใบกับบรรยากาศ รูนี้ถูกล้อมรอบด้วยเซลล์ พาเรนไคมาชนิดพิเศษคู่หนึ่งที่เรียกว่าเซลล์คุมซึ่งทำหน้าที่ควบคุมขนาดของช่องเปิดปากใบ
โดยทั่วไปแล้ว คำนี้มักใช้ในความหมายรวมๆ กัน หมายถึงกลุ่มปากใบทั้งหมด ซึ่งประกอบด้วยเซลล์ยามคู่และรูพรุน ซึ่งเรียกว่าช่องเปิดปากใบ[ 1 ]อากาศที่มีออกซิเจนซึ่งใช้ในการหายใจและคาร์บอนไดออกไซด์ซึ่งใช้ในการสังเคราะห์แสงจะผ่านปากใบโดยการแพร่ ของก๊าซ ไอ น้ำจะแพร่ผ่านปากใบเข้าสู่บรรยากาศในกระบวนการที่เรียกว่าการ คายน้ำ
ปากใบมีอยู่ใน ระยะ สปอโรไฟต์ ของ พืชบกส่วนใหญ่ยกเว้นลิเวอร์เวิร์ตรวมถึงมอ ส และฮอร์นเวิร์ต บาง ชนิด ในพืชมีท่อลำเลียง จำนวน ขนาด และการกระจายตัวของปากใบมีความแตกต่างกันอย่างมาก พืชใบเลี้ยงคู่มักมีปากใบอยู่บนพื้นผิวด้านล่างของใบมากกว่าพื้นผิวด้านบน พืช ใบเลี้ยงเดี่ยวเช่นหัวหอมข้าวโอ๊ตและข้าวโพดอาจมีจำนวนปากใบเท่าๆ กันบนทั้งสองด้านของใบ[ 2 ] : 5 ในพืชที่มีใบลอย ปากใบอาจพบได้เฉพาะบนผิวใบด้านบนเท่านั้น และใบที่จมอยู่ใต้น้ำอาจไม่มีปากใบเลย พืชยืนต้นส่วนใหญ่มีปากใบอยู่เฉพาะบนพื้นผิวด้านล่างของใบเท่านั้น[ 3 ]ใบที่มีปากใบทั้งบนพื้นผิวด้านบนและด้านล่างเรียกว่าใบแอมฟิสโตมาตั ส ใบที่มีปากใบเฉพาะบนพื้นผิวด้านล่างเรียกว่าไฮโปสโตมา ตัส และใบที่มีปากใบเฉพาะบนพื้นผิวด้านบนเรียกว่าเอปิสโตมาตัสหรือไฮเปอร์สโตมาตัส [ 3 ] ขนาดแตกต่างกันไปในแต่ละชนิด โดยมีความยาวจากปลายถึงปลายตั้งแต่ 10 ถึง 80 ไมโครเมตรและความกว้างตั้งแต่ไม่กี่ไมโครเมตรถึง 50 ไมโครเมตร[ 4 ]
การทำงาน

การได้ รับ CO2 และการสูญเสียน้ำ
คาร์บอนไดออกไซด์ซึ่งเป็นสารตั้งต้นสำคัญในการสังเคราะห์แสงมีอยู่ในชั้นบรรยากาศที่ความเข้มข้นประมาณ 425 ppm พืชส่วนใหญ่ต้องการให้ปากใบเปิดในเวลากลางวัน ช่องว่างอากาศในใบจะอิ่มตัวด้วยไอน้ำซึ่งจะออกจากใบผ่านปากใบในกระบวนการที่เรียกว่าการคายน้ำดังนั้น พืชจึงไม่สามารถได้รับคาร์บอนไดออกไซด์โดยไม่สูญเสียไอน้ำไปพร้อมกัน[ 5 ]
แนวทางทางเลือก
โดยปกติแล้ว คาร์บอนไดออกไซด์จะถูกตรึงเข้ากับไรบูโลส 1,5-บิสฟอสเฟต (RuBP) โดยเอนไซม์RuBisCOใน เซลล์มีโซ ฟิลล์ที่สัมผัสโดยตรงกับช่องว่างอากาศภายในใบ ซึ่งทำให้ปัญหาการคายน้ำรุนแรงขึ้นด้วยเหตุผลสองประการ คือ ประการแรก RuBisCO มีความสัมพันธ์กับคาร์บอนไดออกไซด์ค่อนข้างต่ำ และประการที่สอง มันตรึงออกซิเจนเข้ากับ RuBP ทำให้สิ้นเปลืองพลังงานและคาร์บอนในกระบวนการที่เรียกว่าการหายใจระดับแสง (photorespiration ) ด้วยเหตุผลทั้งสองประการนี้ RuBisCO จึงต้องการความเข้มข้นของคาร์บอนไดออกไซด์สูง ซึ่งหมายถึงการเปิดของปากใบกว้าง และส่งผลให้สูญเสียน้ำมาก
สามารถใช้รูเปิดปากใบที่แคบลงร่วมกับโมเลกุลตัวกลางที่มีความสามารถในการจับคาร์บอนไดออกไซด์สูง เช่นฟอสโฟอีโนลไพรูเวตคาร์บอกซิเลส (PEPcase) ได้ อย่างไรก็ตาม การดึงผลิตภัณฑ์จากการตรึงคาร์บอนจาก PEPCase เป็นกระบวนการที่ต้องใช้พลังงานสูง ดังนั้น การใช้ PEPCase จึงเหมาะสมเฉพาะในกรณีที่น้ำมีจำกัดแต่แสงสว่างเพียงพอ หรือในกรณีที่อุณหภูมิสูงทำให้ความสามารถในการละลายของออกซิเจนเพิ่มขึ้นเมื่อเทียบกับคาร์บอนไดออกไซด์ ซึ่งจะทำให้ปัญหาการเติมออกซิเจนของ RuBisCo รุนแรงขึ้น
พืช CAM

พืชกลุ่มหนึ่งซึ่งส่วนใหญ่เป็นพืชทะเลทราย เรียกว่าพืช "CAM" ( กระบวนการเผาผลาญกรดแครสซูลาเซียนตามชื่อวงศ์ Crassulaceae ซึ่งเป็นวงศ์ที่ค้นพบกระบวนการ CAM เป็นครั้งแรก) จะเปิดปากใบในเวลากลางคืน (เมื่อน้ำระเหยจากใบช้าลงสำหรับระดับการเปิดปากใบที่กำหนด) ใช้เอนไซม์ PEPcase ในการตรึงคาร์บอนไดออกไซด์และเก็บผลิตภัณฑ์ไว้ในแวคิวโอลขนาดใหญ่ วันรุ่งขึ้น พวกมันจะปิดปากใบและปล่อยคาร์บอนไดออกไซด์ที่ตรึงไว้ในคืนก่อนออกมาสู่สภาวะที่มี RuBisCO ซึ่งจะทำให้ RuBisCO อิ่มตัวด้วยคาร์บอนไดออกไซด์ ทำให้เกิดการหายใจระดับแสงน้อยที่สุด อย่างไรก็ตาม วิธีการนี้มีข้อจำกัดอย่างมากในด้านความสามารถในการเก็บคาร์บอนที่ตรึงไว้ในแวคิวโอล ดังนั้นจึงเหมาะสมเฉพาะในกรณีที่ขาดแคลนน้ำอย่างรุนแรงเท่านั้น
การเปิดและปิด

อย่างไรก็ตาม พืชส่วนใหญ่ไม่มี CAM ดังนั้นจึงต้องเปิดและปิดปากใบในเวลากลางวันเพื่อตอบสนองต่อสภาวะที่เปลี่ยนแปลง เช่น ความเข้มของแสง ความชื้น และความเข้มข้นของคาร์บอนไดออกไซด์ เมื่อสภาวะเอื้อต่อการเปิดปากใบ (เช่น ความเข้มของแสงสูงและความชื้นสูง) ปั๊มโปรตอนจะขับโปรตอน (H + ) จากเซลล์ยาม ซึ่งหมายความว่า ศักย์ไฟฟ้าของเซลล์จะลดลงเรื่อยๆ ศักย์ไฟฟ้าลบนี้จะเปิดช่องโพแทสเซียมที่ควบคุมด้วยแรงดันไฟฟ้า ทำให้เกิดการดูดซึม ไอออน โพแทสเซียม (K + ) เพื่อรักษาระดับศักย์ไฟฟ้าลบภายในนี้เพื่อไม่ให้การเข้าของไอออนโพแทสเซียมหยุดลง ไอออนลบจะปรับสมดุลกับการไหลเข้าของโพแทสเซียม ในบางกรณี ไอออนคลอไรด์จะเข้า ในขณะที่ในพืชชนิดอื่น ไอออนอินทรีย์มาเลตจะถูกผลิตขึ้นในเซลล์ยาม การเพิ่มขึ้นของความเข้มข้นของสารละลายนี้จะลดศักย์น้ำภายในเซลล์ ซึ่งส่งผลให้เกิดการแพร่ของน้ำเข้าสู่เซลล์ผ่านกระบวนการออสโมซิสทำให้ ปริมาตรและ ความดันเต่งของเซลล์เพิ่มขึ้นจากนั้น เนื่องจากวงแหวนของไมโครไฟบริล เซลลูโลส ที่ป้องกันไม่ให้ความกว้างของเซลล์ยามบวม และทำให้แรงดันเต่งตัวที่เพิ่มขึ้นเท่านั้นที่ทำให้เซลล์ยามยืดออก โดยที่ปลายของเซลล์ยามถูกยึดไว้อย่างแน่นหนาโดย เซลล์ ผิวหนัง โดยรอบ เซลล์ยามทั้งสองจึงยืดออกโดยการโค้งออกจากกัน ทำให้เกิดรูพรุนเปิดที่ก๊าซสามารถแพร่ผ่านได้[ 6 ]
เมื่อรากเริ่มรับรู้ถึงการขาดแคลนน้ำในดินกรดแอบซิสิก (ABA) จะถูกปล่อยออกมา[ 7 ] ABA จะจับกับโปรตีนตัวรับในเยื่อหุ้มพลาสมาและไซโตซอลของเซลล์ยาม ซึ่งจะทำให้ค่า pH ของไซโตซอลของเซลล์สูงขึ้นก่อน และทำให้ความเข้มข้นของ Ca 2+อิสระเพิ่มขึ้นในไซโตซอลเนื่องจากการไหลเข้าจากภายนอกเซลล์และการปล่อย Ca 2+จากแหล่งเก็บภายใน เช่น เอนโดพลาสมิกเรติคูลัมและแวคิวโอล[ 8 ]สิ่งนี้ทำให้คลอไรด์ (Cl − ) และไอออนอินทรีย์ออกจากเซลล์ ประการที่สอง สิ่งนี้จะหยุดการดูดซึม K + เพิ่มเติม เข้าสู่เซลล์ และส่งผลให้ K + สูญเสียไป การสูญเสียสารละลายเหล่านี้ทำให้ ศักยภาพของน้ำเพิ่มขึ้นซึ่งส่งผลให้เกิดการแพร่ของน้ำกลับออกจากเซลล์โดยออสโมซิสทำให้เซลล์เกิดการหดตัวซึ่งส่งผลให้รูเปิดปากใบปิดลง
เซลล์ยามมีคลอโรพลาสต์มากกว่าเซลล์ผิวหนังอื่นๆ ที่เซลล์ยามได้รับมาจาก หน้าที่ของพวกมันยังเป็นที่ถกเถียงกันอยู่[ 9 ] [ 10 ]
การอนุมานพฤติกรรมของปากใบจากการแลกเปลี่ยนก๊าซ
ระดับความต้านทานของปากใบสามารถกำหนดได้โดยการวัดการแลกเปลี่ยนก๊าซของใบ อัตรา การคายน้ำขึ้นอยู่กับ ความต้านทาน การแพร่กระจายที่เกิดจากรูปากใบและขึ้นอยู่กับ ความแตกต่าง ของความชื้นระหว่างช่องว่างอากาศภายในใบกับอากาศภายนอก ดังนั้นความต้านทานของปากใบ (หรือค่าผกผันของความต้านทาน คือการนำไฟฟ้าของปากใบ ) จึงสามารถคำนวณได้จากอัตราการคายน้ำและความแตกต่างของความชื้น ซึ่งช่วยให้นักวิทยาศาสตร์สามารถตรวจสอบการตอบสนองของปากใบต่อการเปลี่ยนแปลงของสภาพแวดล้อม เช่น ความเข้มของแสงและความเข้มข้นของก๊าซ เช่น ไอน้ำ คาร์บอนไดออกไซด์ และโอโซนได้อย่างไร[ 11 ]การระเหย ( E ) สามารถคำนวณได้ดังนี้[ 12 ]

โดยที่e iและe aคือความดันย่อยของน้ำในใบและในอากาศโดยรอบตามลำดับPคือความดันบรรยากาศ และrคือความต้านทานของปากใบ ค่าผกผันของrคือค่าการนำไฟฟ้าของไอน้ำ ( g ) ดังนั้นสมการจึงสามารถจัดเรียงใหม่ได้เป็น[ 12 ]

และแก้หาg : [ 12 ]

การดูด ซับ CO2 ในกระบวนการสังเคราะห์แสง( A ) สามารถคำนวณได้จาก

โดยที่C aและC i คือความดันย่อยของ CO 2ในบรรยากาศและใต้ปากใบตามลำดับ อัตราการระเหยจากใบสามารถกำหนดได้โดยใช้ระบบการสังเคราะห์แสงเครื่องมือทางวิทยาศาสตร์เหล่านี้จะวัดปริมาณไอน้ำที่ออกจากใบและความดันไอของอากาศโดยรอบ ระบบการสังเคราะห์แสงอาจคำนวณประสิทธิภาพการใช้น้ำ ( A / E ), g , ประสิทธิภาพการใช้น้ำที่แท้จริง ( A / g ) และC iเครื่องมือทางวิทยาศาสตร์เหล่านี้มักใช้โดยนักสรีรวิทยาพืชเพื่อวัดการดูดซับ CO 2และวัดอัตราการสังเคราะห์แสง[ 13 ] [ 14 ]
วิวัฒนาการ
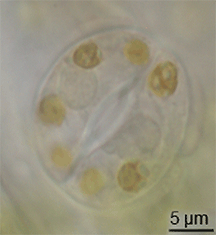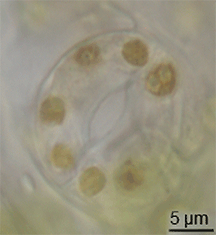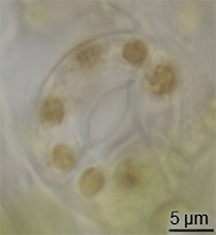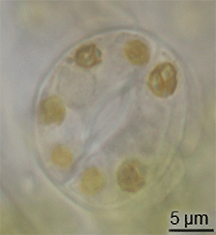
มีหลักฐานเพียงเล็กน้อยเกี่ยวกับการวิวัฒนาการของปากใบในบันทึกฟอสซิล แต่ปากใบได้ปรากฏขึ้นในพืชบกในช่วงกลางของยุคไซลูเรียน[ 15 ]พวกมันอาจวิวัฒนาการมาจากการปรับเปลี่ยนคอนเซปแทเคิลจากบรรพบุรุษของพืชที่มีลักษณะคล้ายสาหร่าย[ 16 ] อย่างไรก็ตาม การวิวัฒนาการของปากใบต้องเกิดขึ้นพร้อมกับ การวิวัฒนาการของ คิวติเคิล ที่เป็นขี้ผึ้ง ซึ่งลักษณะทั้งสองนี้รวมกันถือเป็นข้อได้เปรียบที่สำคัญสำหรับพืชบกในยุคแรก
การพัฒนา
เซลล์ชั้นนอกสุด (epidermal cells) มีอยู่ 3 ชนิดหลัก ซึ่งทั้งหมดล้วนมีต้นกำเนิดมาจากเนื้อเยื่อชั้นนอกสุด (L1) ของเนื้อเยื่อเจริญปลายยอด (shoot apical meristem ) ที่เรียกว่าเซลล์โปรโตเดอร์มัล (protodermal cells) ได้แก่ไตรโคม (trichomes) , เซลล์พื้นผิว (pavement cells)และเซลล์ยาม (guard cells) ซึ่งเรียงตัวกันอย่างไม่สุ่ม
การแบ่งเซลล์แบบไม่สมมาตรเกิดขึ้นในเซลล์โปรโตเดอร์มัล ส่งผลให้เกิดเซลล์ขนาดใหญ่หนึ่งเซลล์ซึ่งจะกลายเป็นเซลล์พื้นผิว และเซลล์ขนาดเล็กกว่าที่เรียกว่าเมริสเตมอยด์ ซึ่งในที่สุดจะแตกต่างไปเป็นเซลล์ยามที่ล้อมรอบปากใบ เมริสเตมอยด์นี้จะแบ่งตัวแบบไม่สมมาตรหนึ่งถึงสามครั้งก่อนที่จะแตกต่างไปเป็นเซลล์แม่ยาม เซลล์แม่ยามจะแบ่งตัวแบบสมมาตรหนึ่งครั้ง ซึ่งก่อให้เกิดเซลล์ยามคู่หนึ่ง[ 17 ]การแบ่งเซลล์ถูกยับยั้งในบางเซลล์ ดังนั้นจึงมีเซลล์อย่างน้อยหนึ่งเซลล์อยู่ระหว่างปากใบเสมอ[ 18 ]
รูปแบบปากใบถูกควบคุมโดยปฏิสัมพันธ์ของ ส่วนประกอบ การส่งสัญญาณ หลายอย่าง เช่นEPF (Epidermal Patterning Factor), ERL (ERecta Like) และYODA ( MAP kinase kinase kinase ที่คาดการณ์ไว้ ) [ 18 ]การกลายพันธุ์ในยีนใดๆ ที่เข้ารหัสปัจจัยเหล่านี้อาจเปลี่ยนแปลงการพัฒนาของปากใบในชั้นหนังกำพร้า[ 18 ]ตัวอย่างเช่น การกลายพันธุ์ในยีนหนึ่งทำให้เกิดปากใบมากขึ้นและรวมกลุ่มกัน จึงเรียกว่า Too Many Mouths ( TMM ) [ 17 ]ในขณะที่การหยุดชะงักของ ยีน SPCH (SPeecCHless) ป้องกันการพัฒนาของปากใบโดยสิ้นเชิง[ 18 ] การยับยั้งการสร้างปากใบสามารถเกิดขึ้นได้จากการกระตุ้น EPF1 ซึ่งกระตุ้น TMM/ERL ซึ่งร่วมกันกระตุ้น YODA YODA ยับยั้ง SPCH ทำให้กิจกรรมของ SPCH ลดลง ป้องกันการแบ่งเซลล์แบบไม่สมมาตรที่เริ่มต้นการสร้างปากใบ[ 18 ] [ 19 ]การพัฒนาของปากใบยังได้รับการประสานงานโดยสัญญาณเปปไทด์ของเซลล์ที่เรียกว่า stomagen ซึ่งส่งสัญญาณกระตุ้น SPCH ส่งผลให้จำนวนปากใบเพิ่มขึ้น[ 20 ]
ปัจจัยด้านสิ่งแวดล้อมและฮอร์โมนสามารถส่งผลต่อการพัฒนาของปากใบได้ แสงจะเพิ่มการพัฒนาของปากใบในพืช ในขณะที่พืชที่ปลูกในที่มืดจะมีปากใบน้อยลงออกซินจะยับยั้งการพัฒนาของปากใบโดยส่งผลต่อการพัฒนาในระดับตัวรับ เช่น ตัวรับ ERL และ TMM อย่างไรก็ตาม ความเข้มข้นต่ำของออกซินจะช่วยให้เซลล์แม่ของเซลล์ยามแบ่งตัวอย่างเท่าเทียมกันและเพิ่มโอกาสในการสร้างเซลล์ยาม[ 21 ]
ต้นไม้ดอกส่วนใหญ่มีปากใบเฉพาะบนผิวใบด้านล่างเท่านั้นต้นป็อปลาร์และต้นวิลโลว์มีปากใบทั้งสองด้าน เมื่อใบพัฒนาปากใบทั้งสองด้าน ปากใบที่ผิวใบด้านล่างมักจะมีขนาดใหญ่กว่าและมีจำนวนมากกว่า แต่ขนาดและความถี่ของปากใบอาจแตกต่างกันไปในแต่ละชนิดและสายพันธุ์ ใบของ ต้นเถ้าขาวและ ต้น เบิร์ชขาวมีปากใบน้อยกว่าแต่มีขนาดใหญ่กว่า ในทางกลับกันต้นเมเปิลน้ำตาลและต้นเมเปิลเงินมีปากใบขนาดเล็กแต่มีจำนวนมากกว่า[ 22 ]
ประเภท
มีการจำแนกประเภทของปากใบที่แตกต่างกันหลายประเภท ประเภทหนึ่งที่ใช้กันอย่างแพร่หลายนั้นอิงตามประเภทที่Julien Joseph Vesqueนำเสนอในปี พ.ศ. 2432 ซึ่งได้รับการพัฒนาเพิ่มเติมโดย Metcalfe และ Chalk [ 23 ]และต่อมาได้รับการเสริมเพิ่มเติมโดยผู้เขียนคนอื่นๆ โดยอิงตามขนาด รูปร่าง และการจัดเรียงของเซลล์ย่อยที่ล้อมรอบเซลล์ยามสองเซลล์[ 24 ] พวกเขาแยกแยะสำหรับพืชใบเลี้ยงคู่ดังนี้ :
- ปากใบ แบบแอคติโนไซติก (หมายถึงแบบเซลล์รูปดาว) มีเซลล์คุมที่ล้อมรอบด้วยเซลล์แผ่รัศมีอย่างน้อยห้าเซลล์ forming เป็นวงกลมคล้ายดาว นี่เป็นชนิดที่หายาก ซึ่งพบได้ในวงศ์Ebenaceaeเป็นต้น
- ปากใบ แบบอะนิโซไซติก (หมายถึงเซลล์ไม่เท่ากัน) มีเซลล์คุมอยู่ระหว่างเซลล์ข้างเคียงขนาดใหญ่สองเซลล์และเซลล์ข้างเคียงขนาดเล็กกว่าอีกหนึ่งเซลล์ ปากใบชนิดนี้พบได้ในพืชใบเลี้ยงคู่มากกว่าสามสิบวงศ์ รวมถึงวงศ์Brassicaceae,SolanaceaeและCrassulaceaeบางครั้งเรียกว่าชนิดกะหล่ำ
- ปากใบ แบบอะโนโมไซติก (หมายถึงเซลล์ที่มีรูปร่างไม่สม่ำเสมอ) มีเซลล์คุมที่ล้อมรอบด้วยเซลล์ที่มีขนาด รูปร่าง และการจัดเรียงเหมือนกับเซลล์ชั้นนอกสุดอื่นๆ ปากใบชนิดนี้พบได้ในพืชใบเลี้ยงคู่มากกว่าร้อยวงศ์ เช่น วงศ์Apocynaceae,Boraginaceae,ChenopodiaceaeและCucurbitaceaeบางครั้งเรียกว่าแบบรานันคูเลเซียส
- ปากใบแบบ ไดอะไซติก (หมายถึงมีเซลล์ไขว้กัน) มีเซลล์คุมล้อมรอบด้วยเซลล์ย่อยสองเซลล์ ซึ่งแต่ละเซลล์จะล้อมรอบปลายด้านหนึ่งของช่องเปิดและสัมผัสกันตรงข้ามกับกึ่งกลางของช่องเปิด ปากใบชนิดนี้พบได้ในพืชใบเลี้ยงคู่มากกว่าสิบวงศ์ เช่น วงศ์แคริโอฟิลลาซี (Caryophyllaceaeและวงศ์อะแคนทาซี (Acanthaceae) บางครั้งเรียกว่าชนิดแคริโอฟิลลาเซียส (Caryophyllaceous type)
- ปากใบ แบบเฮมิพาราไซติกมีเซลล์ข้างเคียงเพียงเซลล์เดียวที่แตกต่างจากเซลล์ชั้นนอกที่อยู่รอบข้าง โดยมีความยาวขนานกับช่องเปิดของปากใบ ปากใบชนิดนี้พบได้ในพืชวงศ์ MolluginaceaeและAizoaceaeเป็นต้น
- ปากใบ แบบพาราไซติก (หมายถึงเซลล์ขนาน) มีเซลล์ย่อยหนึ่งเซลล์หรือมากกว่านั้นขนานกับช่องเปิดระหว่างเซลล์คุม เซลล์ย่อยเหล่านี้อาจยื่นเลยเซลล์คุมหรือไม่ก็ได้ ปากใบชนิดนี้พบได้ในพืชใบเลี้ยงคู่มากกว่าร้อยวงศ์ เช่นวงศ์ Rubiaceae,ConvolvulaceaeและFabaceaeบางครั้งเรียกว่าแบบรูเบียซ (rubiaceous type)
ในพืชใบเลี้ยงเดี่ยวพบปากใบหลายประเภท เช่น:
- ปากใบ แบบแกรมมินัสหรือแกรมมิโนอิด (หมายถึงคล้ายหญ้า ) มีเซลล์คุมสองเซลล์ล้อมรอบด้วยเซลล์ย่อยรูปเลนส์สองเซลล์ เซลล์คุมจะแคบลงตรงกลางและโป่งออกที่ปลายทั้งสองข้าง ส่วนกลางนี้หนาขึ้นอย่างเห็นได้ชัด แกนของเซลล์ย่อยขนานกับช่องเปิดปากใบ[ 25 ]ประเภทนี้สามารถพบได้ในวงศ์พืชใบเลี้ยงเดี่ยวได้แก่PoaceaeและCyperaceae [ 26 ]
- ปากใบแบบ เฮกซาไซติก (หมายถึงมีหกเซลล์) มีเซลล์ย่อยหกเซลล์ล้อมรอบเซลล์คุมทั้งสองเซลล์ โดยมีเซลล์ย่อยหนึ่งเซลล์อยู่ที่ปลายแต่ละด้านของช่องเปิดปากใบ เซลล์ย่อยหนึ่งเซลล์อยู่ติดกับเซลล์คุมแต่ละเซลล์ และเซลล์ย่อยอีกหนึ่งเซลล์อยู่ระหว่างเซลล์ย่อยเซลล์สุดท้ายกับเซลล์ชั้นนอกสุดของใบ ปากใบแบบนี้พบได้ในพืชใบเลี้ยงเดี่ยวบางวงศ์
- ปากใบ แบบเทตราไซติก (หมายถึงมีสี่เซลล์) มีเซลล์ย่อยสี่เซลล์ เซลล์หนึ่งอยู่ที่ปลายแต่ละด้านของช่องเปิด และอีกเซลล์หนึ่งอยู่ข้างๆ เซลล์คุมแต่ละเซลล์ ปากใบแบบนี้พบได้ในพืชใบเลี้ยงเดี่ยวหลายวงศ์ แต่ก็พบได้ในพืชใบเลี้ยงคู่บางชนิด เช่นสกุล Tiliaและวงศ์Asclepiadaceae
ในเฟิร์นสามารถแบ่งออกได้เป็น 4 ประเภทหลัก:
- ปากใบ แบบไฮโปไซติกมีเซลล์คุมสองเซลล์เรียงตัวเป็นชั้นเดียวกับเซลล์เอพิเดอร์มิสธรรมดา แต่มีเซลล์เสริมสองเซลล์อยู่บนพื้นผิวด้านนอกของเอพิเดอร์มิส เรียงตัวขนานกับเซลล์คุม โดยมีรูพรุนอยู่ระหว่างเซลล์เสริมทั้งสอง และอยู่เหนือช่องเปิดของปากใบ
- ปากใบ แบบเพอริไซติกมีเซลล์คุมสองเซลล์ซึ่งถูกล้อมรอบด้วยเซลล์ย่อยต่อเนื่องหนึ่งเซลล์ (คล้ายโดนัท)
- ปากใบ แบบเดสโมไซติกมีเซลล์คุมสองเซลล์ที่ถูกล้อมรอบโดยสมบูรณ์ด้วยเซลล์ย่อยหนึ่งเซลล์ที่ปลายทั้งสองข้างยังไม่เชื่อมติดกัน (เหมือนไส้กรอก)
- ปากใบ แบบโพโลไซติกมีเซลล์คุมสองเซลล์ซึ่งถูกล้อมรอบด้วยเซลล์ข้างเคียงหนึ่งเซลล์เป็นส่วนใหญ่ แต่ยังสัมผัสกับเซลล์ผิวหนังชั้นนอกทั่วไปด้วย (คล้ายรูปตัว U หรือเกือกม้า)
แคตตาล็อกภาพพิมพ์ผิวใบที่แสดงปากใบจากพืชหลากหลายชนิด สามารถดูได้ในวิกิมีเดียคอมมอนส์https://commons.wikimedia.org/wiki/Category:Leaf_epidermis_and_stomata_prints
โพรงปากใบ
โพรงปากใบเป็นบริเวณที่ยุบตัวลงของผิวใบซึ่งก่อตัวเป็นโครงสร้างคล้ายห้องที่บรรจุปากใบหนึ่งหรือมากกว่า และบางครั้งอาจมีขนหรือการสะสมของขี้ผึ้งโพรงปากใบอาจเป็นการปรับตัวให้เข้ากับสภาพแห้งแล้งและสภาพอากาศแห้งเมื่อโพรงปากใบมีความเด่นชัดมาก อย่างไรก็ตาม สภาพอากาศแห้งไม่ใช่สถานที่เดียวที่สามารถพบได้ พืชต่อไปนี้เป็นตัวอย่างของสายพันธุ์ที่มีโพรงปากใบหรือห้องด้านหน้า: Nerium oleander , สน, Hakea [ 27 ]และDrimys winteriซึ่งเป็นสายพันธุ์ของพืชที่พบในป่าเมฆ[ 28 ]
ปากใบเป็นเส้นทางก่อโรค
ปากใบเป็นรูบนใบที่เชื้อโรคสามารถเข้าไปได้โดยไม่มีอุปสรรค อย่างไรก็ตาม ปากใบสามารถรับรู้ถึงการมีอยู่ของเชื้อโรคบางชนิดหรือทั้งหมดได้[ 29 ]อย่างไรก็ตาม แบคทีเรียก่อโรคที่นำมาใช้กับใบพืชArabidopsis สามารถปล่อยสารเคมี โคโรนาทีนซึ่งกระตุ้นให้ปากใบเปิดออกอีกครั้ง [ 30 ]
ปากใบและการเปลี่ยนแปลงสภาพภูมิอากาศ
การตอบสนองของปากใบต่อปัจจัยทางสิ่งแวดล้อม
การสังเคราะห์แสงการลำเลียงน้ำของพืช ( ไซเล็ม ) และการแลกเปลี่ยนก๊าซถูกควบคุมโดยการทำงานของปากใบ ซึ่งมีความสำคัญต่อการทำงานของพืช[ 31 ]
ปากใบตอบสนองต่อแสง โดยแสงสีฟ้ามีประสิทธิภาพในการกระตุ้นการตอบสนองของปากใบ มากกว่า แสงสีแดง เกือบ 10 เท่า งานวิจัยชี้ให้เห็นว่านี่เป็นเพราะการตอบสนองของปากใบต่อแสงสีฟ้าเป็นอิสระจากส่วนประกอบอื่นๆ ของใบ เช่น คลอโรฟิลล์โปรโตพลาสต์ของเซลล์ยาม จะบวมภายใต้แสงสี ฟ้าหากมีโพแทสเซียม เพียงพอ [ 32 ]การศึกษาหลายชิ้นพบว่าการเพิ่มความเข้มข้นของโพแทสเซียมอาจทำให้ปากใบเปิดมากขึ้นในตอนเช้า ก่อนที่กระบวนการสังเคราะห์แสงจะเริ่มต้น แต่ในช่วงบ่ายซูโครสจะมีบทบาทสำคัญมากขึ้นในการควบคุมการเปิดของปากใบ[ 33 ]ซีแซนทินในเซลล์ยามทำหน้าที่เป็นตัวรับแสงสีฟ้าซึ่งเป็นตัวกลางในการเปิดของปากใบ[ 34 ]ผลของแสงสีฟ้าต่อเซลล์ยามจะกลับกันเมื่อได้รับแสงสีเขียว ซึ่งจะทำให้ซีแซนทินเกิดไอโซเมอไรเซชัน[ 34 ]
ความหนาแน่นของปากใบและช่องเปิด (ความยาวของปากใบ) แตกต่างกันไปตามปัจจัยด้านสิ่งแวดล้อมหลายประการ เช่น ความเข้มข้นของ CO2 ในบรรยากาศความเข้มของแสง อุณหภูมิอากาศ และช่วงเวลากลางวัน (ระยะเวลากลางวัน) [ 35 ] [ 36 ]
การลดความหนาแน่นของปากใบเป็นวิธีหนึ่งที่พืชตอบสนองต่อการเพิ่มขึ้นของความเข้มข้นของ CO2 ในบรรยากาศ( [CO2 ] atm ) [ 37 ] แม้ว่าการเปลี่ยนแปลงในการตอบสนองต่อ [CO2 ] atm จะ เป็นกลไกที่เข้าใจได้น้อยที่สุด แต่การตอบสนองของปากใบนี้เริ่มถึงจุดสูงสุดแล้ว ซึ่งคาดว่าจะส่งผลกระทบต่อกระบวนการ คายน้ำและการสังเคราะห์แสงในพืชในไม่ช้า[ 31 ] [ 38 ]
ความแห้งแล้งยับยั้งการเปิดปากใบ แต่การวิจัยเกี่ยวกับถั่วเหลืองชี้ให้เห็นว่าความแห้งแล้งระดับปานกลางไม่มีผลกระทบอย่างมีนัยสำคัญต่อการปิดปากใบของใบ การปิดปากใบมีกลไกที่แตกต่างกัน ความชื้นต่ำทำให้เซลล์ยามเกิด ความเครียด ส่งผลให้สูญเสีย ความเต่งซึ่งเรียกว่าการปิดแบบไฮโดรพาสซีฟ การปิดแบบไฮโดรแอคทีฟนั้นตรงกันข้ามกับการที่ใบทั้งใบได้รับผลกระทบจากความเครียดจากความแห้งแล้ง ซึ่งเชื่อว่าน่าจะเกิดจากกรดแอบซิสิก[ 39 ]
การปรับตัวในอนาคตในช่วงการเปลี่ยนแปลงสภาพภูมิอากาศ
คาดว่า [CO 2 ] atmจะสูงถึง 500–1000 ppm ภายในปี 2100 [ 31 ] 96% ของ 400,000 ปีที่ผ่านมามี CO 2 ต่ำกว่า 280 ppm จากตัวเลขนี้ มีความเป็นไปได้สูงที่จีโนไทป์ของพืชในปัจจุบันจะแยกตัวออกจากญาติในยุคก่อนอุตสาหกรรม[ 31 ]
ยีนHIC (high carbon dioxide) เข้ารหัสตัวควบคุมเชิงลบสำหรับการพัฒนาปากใบในพืช[ 40 ]การวิจัยเกี่ยวกับยีนHIC โดยใช้ Arabidopsis thalianaพบว่าไม่มีการเพิ่มขึ้นของการพัฒนาปากใบในอัลลีล เด่น แต่ในอัลลีลด้อยแบบ 'wild type' แสดงให้เห็นการเพิ่มขึ้นอย่างมาก ทั้งสองกรณีตอบสนองต่อระดับ CO2ที่เพิ่มขึ้นในบรรยากาศ[ 40 ]การศึกษาเหล่านี้บ่งชี้ว่าการตอบสนองของพืชต่อการเปลี่ยนแปลงระดับ CO2 นั้นถูกควบคุมโดยพันธุกรรมเป็นส่วนใหญ่
ผลกระทบต่อภาคเกษตรกรรม
ผลของปุ๋ย CO2 ถูกประเมินค่าสูงเกินไปอย่างมากในระหว่างการ ทดลอง Free-Air Carbon dioxide Enrichment (FACE) ซึ่งผลลัพธ์แสดงให้เห็นว่าระดับ CO2 ที่เพิ่มขึ้นในบรรยากาศช่วยเพิ่มการสังเคราะห์แสง ลดการคายน้ำ และเพิ่มประสิทธิภาพการใช้น้ำ (WUE) [ 37 ]มวลชีวภาพที่เพิ่มขึ้นเป็นหนึ่งในผลกระทบ โดยการจำลองจากการทดลองคาดการณ์ว่าผลผลิตพืชจะเพิ่มขึ้น 5–20% ที่ระดับ CO2 550 ppm [ 41 ]อัตราการสังเคราะห์แสงของใบแสดงให้เห็นว่าเพิ่มขึ้น 30–50% ใน พืช C3และ 10–25% ในพืชC4ภายใต้ระดับCO2 ที่เพิ่มขึ้นเป็นสองเท่า [ 41 ]การมีอยู่ของกลไกป้อนกลับส่งผลให้เกิดความยืดหยุ่นทางฟีโนไทป์ในการตอบสนองต่อ [CO2 ] atm ซึ่งอาจเป็นลักษณะที่ปรับตัวได้ในวิวัฒนาการของการหายใจและการทำงานของพืช[ 31 ] [ 36 ]
การทำนายว่าปากใบจะทำงานอย่างไรในระหว่างการปรับตัวนั้นมีประโยชน์ต่อการทำความเข้าใจผลผลิตของระบบพืชทั้งในระบบธรรมชาติและระบบเกษตรกรรม[ 35 ]นักปรับปรุงพันธุ์พืชและเกษตรกรเริ่มทำงานร่วมกันโดยใช้การปรับปรุงพันธุ์พืชเชิงวิวัฒนาการและแบบมีส่วนร่วมเพื่อค้นหาสายพันธุ์ที่เหมาะสมที่สุด เช่น พันธุ์พืชที่ทนต่อความร้อนและภัยแล้งซึ่งสามารถวิวัฒนาการตามธรรมชาติเพื่อรับมือกับการเปลี่ยนแปลงในภาวะที่ต้องเผชิญกับความท้าทายด้านความมั่นคงทางอาหาร[ 37 ]
ลิงก์ภายนอก
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ปาก
ในทางพฤกษศาสตร์ปากใบ ( พหูพจน์: stomata มาจาก ภาษา กรีกστόμαแปลว่า "ปาก") หรือเรียกว่าปากใบ หลายรู ( พหูพจน์ : stomates ) คือรูที่พบในชั้นผิวของใบ ลำต้น และอวัยวะอื่นๆ...
การทำงาน
ภาพถ่ายอิเล็กตรอนไมโครสโคปของปากใบจาก ใบ ผักกาดขาว ( Brassica chinensis )
การได้ รับ CO2 และการสูญเสียน้ำ
คาร์บอนไดออกไซด์ ซึ่งเป็นสารตั้งต้นสำคัญใน การสังเคราะห์แสง มีอยู่ในชั้นบรรยากาศที่ความเข้มข้นประมาณ 425 ppm พืชส่วนใหญ่ต้องการให้ปากใบเปิดในเวลากลางวัน ช่องว่างอากาศใน ใบ จะอิ่มตัวด้วย ไอน้ำ ซึ่งจะออกจากใบผ่านปากใบในกระบวนการที่เรียกว่า การคายน้ำ ดังนั้น...
แนวทางทางเลือก
โดยปกติแล้ว คาร์บอนไดออกไซด์จะถูกตรึงเข้ากับ ไรบูโลส 1,5-บิสฟอสเฟต (RuBP) โดยเอนไซม์ RuBisCO ใน เซลล์มีโซ ฟิลล์ ที่สัมผัสโดยตรงกับช่องว่างอากาศภายในใบ ซึ่งทำให้ปัญหาการคายน้ำรุนแรงขึ้นด้วยเหตุผลสองประการ คือ ประการแรก RuBisCO...