อะโพลิโปโปรตีน บี
| เอพีโอบี | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | APOB , FLDB, LDLCQ4, apoB-100, apoB-48, อะโพลิโปโปรตีน บี, FCHL2 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 107730 ; เอ็มจีไอ : 88052 ; โฮโมโลยีน : 328 ; GeneCards : APOB ; OMA : APOB - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





อะโพลิโปโปรตีน บี ( ApoB ) เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนAPOB การวัดระดับของโปรตีนนี้มักใช้เพื่อตรวจหาความเสี่ยงของโรคหลอดเลือด หัวใจ ตีบ[ 5 ] [ 6 ]
ไอโซฟอร์ม
โปรตีนนี้พบในพลาสมาในสองไอโซฟอร์ม หลัก คือ ApoB48 และ ApoB100 ไอโซฟอร์มแรกถูกสังเคราะห์โดยลำไส้เล็ก เท่านั้น ไอโซฟอร์ม ที่สองถูกสังเคราะห์โดยตับ[ 7 ] ApoB -100 เป็นโปรตีนที่ใหญ่ที่สุดในกลุ่มโปรตีน apoB ประกอบด้วยกรดอะมิโน 4563 ตัว รวมถึงเปปไทด์สัญญาณ 27 ตัว และโปรตีนที่สมบูรณ์ 4536 ตัว[ 7 ]ทั้งสองไอโซฟอร์มถูกเข้ารหัสโดยAPOBและโดย ทรานสคริปต์ mRNA เดียว ที่มีขนาดใหญ่กว่า 16 kb ApoB48 ถูกสร้างขึ้นเมื่อ มีการสร้าง รหัสหยุด (UAA) ที่ตำแหน่ง 2153 โดยการแก้ไข RNAดูเหมือนว่าจะมียีนสไปลซิงเฉพาะเนื้อเยื่อที่ออกฤทธิ์แบบทรานส์ซึ่งกำหนดว่าไอโซฟอร์มใดจะถูกผลิตขึ้นในที่สุดหรืออีกทางหนึ่ง มีหลักฐานบางอย่างที่แสดงว่า องค์ประกอบ ที่ออกฤทธิ์แบบซิสหลายพันคู่เบสขึ้นไปเป็นตัวกำหนดว่าไอโซฟอร์มใดจะถูกผลิต ขึ้น
จากการแก้ไข RNA ทำให้ ApoB48 และ ApoB100 มี ลำดับ ปลายด้าน N ที่เหมือนกัน แต่ ApoB48 ขาด บริเวณจับกับ ตัวรับ LDL ที่ปลายด้าน C ของ ApoB100 ApoB48 ได้รับชื่อนี้เพราะมันประกอบขึ้นเป็น 48% ของลำดับของ ApoB100 ApoB48 เป็นโปรตีนที่มีลักษณะเฉพาะในไคโลไมครอนจากลำไส้เล็ก หลังจากที่ไขมันส่วนใหญ่ในไคโลไมครอนถูกดูดซึมไปแล้ว ApoB48 จะกลับไปยังตับในรูปของส่วนที่เหลือของไคโลไมครอน ซึ่งจะถูกดูดซึมเข้าสู่เซลล์และย่อยสลายไป
การทำงาน
อะโพลิโปโปรตีน บี (ApoB) เป็นอะ โพลิโปโปรตีนหลักของไคโลไมครอน , VLDL , Lp(a) , IDLและLDL (LDL—โดยทั่วไปรู้จักกันในชื่อ " คอเลสเตอรอล ไม่ดี " เมื่อกล่าวถึงโรคหัวใจและโรคหลอดเลือดโดยทั่วไป) ซึ่งมีหน้าที่ในการขนส่ง โมเลกุล ไขมัน ( ลิปิด ) รวมถึงคอเลสเตอรอลไปทั่วร่างกายไปยังเซลล์ ต่างๆ ในเนื้อเยื่อ ทั้งหมด แม้ว่าบทบาทหน้าที่ทั้งหมดของ ApoB ภายในอนุภาค LDL (และอนุภาคขนาดใหญ่กว่าทั้งหมด) ยังคงไม่ชัดเจนนัก แต่ก็เป็นโปรตีนหลักที่ทำหน้าที่จัดระเบียบ (ของเปลือกที่ซับซ้อนทั้งหมดที่ห่อหุ้ม/ขนส่งโมเลกุลไขมันอยู่ภายใน) ของอนุภาค และจำเป็นอย่างยิ่งสำหรับการก่อตัวของอนุภาคเหล่านี้ สิ่งที่ชัดเจนอีกอย่างคือ ApoB บนอนุภาค LDL ทำหน้าที่เป็นลิแกนด์สำหรับตัวรับ LDL ในเซลล์ต่างๆ ทั่วร่างกาย (กล่าวคือ ในทางที่ไม่เป็นทางการ ApoB บ่งชี้ว่าอนุภาคที่ขนส่งไขมันพร้อมที่จะเข้าสู่เซลล์ใดๆ ที่มีตัวรับ ApoB และส่งไขมัน ที่ ขนส่งอยู่ภายในเข้าไปในเซลล์เหล่านั้น)
บทบาทในระบบภูมิคุ้มกันโดยกำเนิด
ไลโปโปรตีนความหนาแน่นต่ำมากและไลโปโปรตีนความหนาแน่นต่ำขัดขวาง ระบบ การรับรู้จำนวนประชากรที่ควบคุมยีนที่จำเป็นสำหรับ การติดเชื้อ Staphylococcus aureus ที่รุกราน กลไกการต่อต้านเกี่ยวข้องกับการจับ ApoB กับ ฟีโรโมน ตัวกระตุ้นอัตโนมัติ ของ S. aureus ป้องกันการส่งสัญญาณผ่านตัวรับ หนูที่ขาด ApoB จะไวต่อการติดเชื้อแบคทีเรียที่รุกรานมากขึ้น[ 8 ]
ความสำคัญทางคลินิก
มีหลักฐานมากมายที่แสดงว่าความเข้มข้นของ ApoB [ 9 ] [ 10 ]และโดยเฉพาะอย่างยิ่งการตรวจวิเคราะห์ NMR [ 11 ] (เฉพาะสำหรับความเข้มข้นของอนุภาค LDL) เป็นตัวบ่งชี้ที่ดีกว่าของสรีรวิทยาที่ขับเคลื่อนโรคหลอดเลือด/หัวใจ มากกว่าคอเลสเตอรอลรวมหรือ LDL-คอเลสเตอรอล (ซึ่งได้รับการส่งเสริมโดยNIH มานาน แล้วตั้งแต่ต้นทศวรรษ 1970) อย่างไรก็ตาม ด้วยเหตุผลหลักด้านต้นทุน/ความซับซ้อนในอดีต คอเลสเตอรอล และLDL-คอเลสเตอรอล ที่ประมาณค่า โดยการคำนวณยังคงเป็นการทดสอบไขมันที่ได้รับการส่งเสริมมากที่สุดสำหรับปัจจัยเสี่ยงของหลอดเลือดแดงแข็ง ApoB จะถูกวัดเป็นประจำโดยใช้การตรวจวิเคราะห์ทางภูมิคุ้มกัน เช่นELISAหรือเนเฟโลเมตรี วิธีการ NMRที่ได้รับการปรับปรุงและอัตโนมัติช่วยให้สามารถวัดความแตกต่างระหว่างอนุภาค ApoB ที่แตกต่างกันมากมายได้
โรคทางพันธุกรรม
ระดับ ApoB ที่สูงมีความเกี่ยวข้องกับโรคหัวใจ ภาวะไฮโปเบตาไลโปโปรตีนีเมียเป็นความผิดปกติทางพันธุกรรมที่อาจเกิดจากการกลายพันธุ์ในยีน ApoB, APOB [ 12 ] ภาวะอะเบตาไลโปโปรตีนีเมียเกิดจากการกลายพันธุ์ใน ยีน โปรตีนถ่ายโอนไตรกลีเซอไรด์ไมโครโซม , MTTP [ 13 ]
การกลายพันธุ์ในยีนAPOB100อาจทำให้เกิดภาวะไขมันในเลือดสูงในครอบครัว ได้เช่นกัน [ 14 ]ซึ่งเป็นรูปแบบทางพันธุกรรม (ลักษณะเด่นของออโตโซม) ของความผิดปกติทางเมตาบอลิซึม ของไขมัน ในเลือดสูง
บทบาทในภาวะดื้อต่ออินซูลิน
การผลิตอะโพลิโปโปรตีนบีมากเกินไปอาจส่งผลให้เกิดความเครียดของเอนโดพลาสมิกเรติคูลัม ที่เกิดจากไขมัน และภาวะดื้อต่ออินซูลินในตับ[ 15 ]
บทบาทในไลโปโปรตีนและภาวะหลอดเลือดแดงแข็ง
ApoB100 พบในไลโปโปรตีนที่มาจากตับ ( VLDL , IDL , LDL [ 16 ] ) ที่สำคัญคือ มีโมเลกุล ApoB100 หนึ่งโมเลกุลต่อไลโปโปรตีนที่มาจากตับหนึ่งโมเลกุล ดังนั้น การใช้ข้อเท็จจริงนี้ เราสามารถหาปริมาณอนุภาคไลโปโปรตีนได้โดยการสังเกตความเข้มข้นรวมของ ApoB100 ในการไหลเวียน เนื่องจากมี ApoB100 เพียงหนึ่งโมเลกุลต่ออนุภาค จำนวนอนุภาคจึงสะท้อนโดยความเข้มข้นของ ApoB100 เทคนิคเดียวกันนี้สามารถนำไปใช้กับไลโปโปรตีนแต่ละประเภท (เช่น LDL) และทำให้สามารถนับจำนวนได้เช่นกัน
เป็นที่ทราบกันดีว่าระดับ ApoB100 มีความสัมพันธ์กับโรคหลอดเลือดหัวใจและเป็นตัวบ่งชี้ที่ดีกว่าความเข้มข้นของ LDL-C มาก[ 17 ] [ 18 ] [ 19 ]เหตุผล: LDL-C ไม่สะท้อนความเข้มข้นของอนุภาคที่แท้จริง และคอเลสเตอรอลไม่สามารถละลายหรือเคลื่อนที่ (ในน้ำ) ได้หากไม่มีอนุภาคพาหะ วิธีง่ายๆ ในการทำความเข้าใจข้อสังเกตนี้คือข้อเท็จจริงที่ว่า ApoB100 หนึ่งตัวต่ออนุภาค สะท้อนความเข้มข้นของอนุภาคไลโปโปรตีนที่แท้จริง (โดยไม่ขึ้นอยู่กับปริมาณคอเลสเตอรอลหรือไขมันอื่นๆ) ด้วยวิธีนี้ เราสามารถเข้าใจได้ว่าจำนวนอนุภาคไลโปโปรตีนที่มี ApoB100 ซึ่งสามารถนำไขมันเข้าสู่ผนังหลอดเลือดแดงได้นั้น เป็นตัวกำหนดที่สำคัญและเป็นตัวขับเคลื่อนของภาวะหลอดเลือดแดงแข็งและโรคหัวใจ
วิธีหนึ่งที่จะอธิบายเรื่องข้างต้นได้คือ การพิจารณาว่าอนุภาคไลโปโปรตีนจำนวนมาก โดยเฉพาะอย่างยิ่งอนุภาค LDL จำนวนมาก จะนำไปสู่การแข่งขันที่ตัวรับ ApoB100 (เช่น ตัวรับ LDL) ของเซลล์ส่วนปลาย เนื่องจากการแข่งขันดังกล่าวจะทำให้ระยะเวลาที่อนุภาค LDL อยู่ในระบบไหลเวียนโลหิตนานขึ้น จึงอาจนำไปสู่โอกาสที่อนุภาคเหล่านั้นจะเกิดปฏิกิริยาออกซิเดชันและ/หรือการดัดแปลงทางเคมีอื่นๆ มากขึ้น การดัดแปลงดังกล่าวอาจลดความสามารถของอนุภาคในการถูกกำจัดโดยตัวรับ LDL แบบดั้งเดิม และ/หรือเพิ่มความสามารถในการโต้ตอบกับตัวรับที่เรียกว่า "ตัวเก็บกวาด" ผลลัพธ์สุทธิคือการเบี่ยงเบนอนุภาค LDL ไปยังตัวรับเก็บกวาดเหล่านี้ ตัวรับเก็บกวาดมักพบในแมโครฟาจโดยแมโครฟาจที่เต็มไปด้วยคอเลสเตอรอลเป็นที่รู้จักกันดีในชื่อ " เซลล์โฟม " เซลล์โฟมเป็นลักษณะเฉพาะของรอยโรคหลอดเลือดแดงแข็ง นอกจากกลไกที่เป็นไปได้ของการสร้างเซลล์โฟมแล้ว การเพิ่มขึ้นของระดับอนุภาค LDL ที่ได้รับการดัดแปลงทางเคมีอาจนำไปสู่การเพิ่มขึ้นของ ความเสียหายต่อเยื่อบุหลอดเลือดได้เช่นกันปรากฏการณ์นี้เกิดขึ้นเนื่องจาก LDL ที่ถูกดัดแปลงมีฤทธิ์เป็นพิษต่อเยื่อบุหลอดเลือด รวมถึงความสามารถในการดึงดูดเซลล์ภูมิคุ้มกันและกระตุ้นการทำงานของเกล็ดเลือด
การศึกษา INTERHEART พบว่าอัตราส่วน ApoB100 / ApoA1 มีประสิทธิภาพมากกว่าในการทำนายความเสี่ยงต่อการเกิดโรคหัวใจวายในผู้ป่วยที่มีภาวะกล้ามเนื้อหัวใจขาดเลือดเฉียบพลัน เมื่อเทียบกับการวัดค่า ApoB100 หรือ ApoA1 เพียงอย่างเดียว[ 20 ] ( ApoA1เป็นโปรตีน HDL หลัก[ 21 ] ) ในประชากรทั่วไปยังไม่ชัดเจน แม้ว่าในการศึกษาล่าสุด ApoB จะเป็นตัวบ่งชี้ความเสี่ยงที่แข็งแกร่งที่สุดสำหรับเหตุการณ์เกี่ยวกับหลอดเลือดหัวใจ[ 22 ]
แนะนำให้รับประทาน อาหารเมดิเตอร์เรเนียนเพื่อลดระดับอะโพลิโปโปรตีนบี[ 23 ]
การศึกษาในหนู
หนูถูกใช้เป็นแบบจำลองสิ่งมีชีวิตในการศึกษา ApoB เนื่องจากพวกมันแสดงโปรตีนที่เทียบเท่ากันที่เรียกว่า ApoB ของหนู (mApoB) หนูที่แสดง mApoB มากเกินไปจะมีระดับ LDL เพิ่มขึ้นและระดับHDLลด ลง [ 24 ]หนูที่มีสำเนาของยีน mApoB ที่ใช้งานได้เพียงหนึ่งสำเนาจะแสดงผลตรงกันข้าม คือต้านทานต่อภาวะคอเลสเตอรอลสูงหนูที่ไม่มีสำเนาของยีนที่ใช้งานได้จะไม่สามารถมีชีวิตอยู่ได้[ 25 ]
ปฏิสัมพันธ์
มีการแสดงให้เห็นว่า ApoB มีปฏิสัมพันธ์กับ apo ( a) [ 26 ] PPIB [ 27 ]ตัวรับแคลซิโทนิน [ 27 ] [ 28 ] และ HSP90B1 [ 27 ] [ 28 ] เชื่อ กันว่า การมีปฏิสัมพันธ์ของ ApoB กับโปรตีโอไกลแคนคอลลาเจนและไฟโบรเนกตินทำให้เกิดภาวะหลอดเลือดแดงแข็ง[ 29 ] [ 30 ]
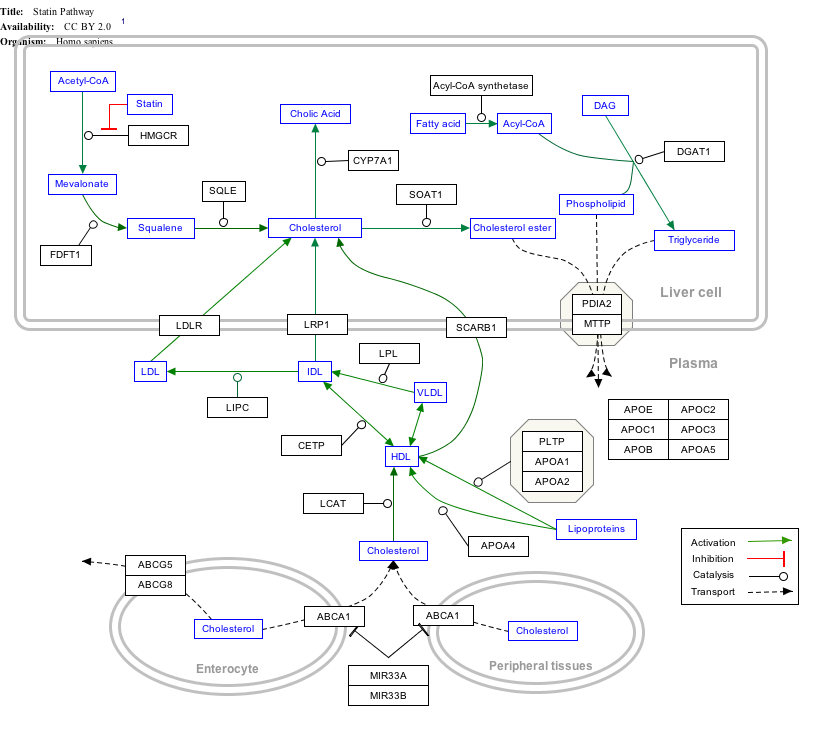
แผนที่เส้นทางแบบโต้ตอบ
คลิกที่ยีน โปรตีน และเมตาบอไลต์ด้านล่างเพื่อเชื่อมโยงไปยังบทความที่เกี่ยวข้อง[ § 1 ]
{kind=link}
- ↑แผนผังเส้นทางแบบโต้ตอบสามารถแก้ไขได้ที่ WikiPathways: " Statin_Pathway_WP430 "
ระเบียบข้อบังคับ
การแสดงออกของAPOBถูกควบคุมโดยองค์ประกอบควบคุมซิสในAPOB 5′ UTRและ 3′ UTR [ 31 ]
การแก้ไขอาร์เอ็นเอ
mRNA ที่เข้ารหัสโปรตีนนี้อยู่ภายใต้การแก้ไข RNA เฉพาะตำแหน่ง จากไซทิดีนเป็นยูริดีน (C เป็น U) ApoB100 และ ApoB48 ถูกเข้ารหัสโดยยีนเดียวกัน อย่างไรก็ตาม ความแตกต่างในโปรตีนที่แปลแล้วไม่ได้เกิดจากการสไปลซิงแบบทางเลือก แต่เกิดจากเหตุการณ์การแก้ไข RNA เฉพาะเนื้อเยื่อ การแก้ไข mRNA ของ ApoB เป็นตัวอย่างแรกของการแก้ไขที่พบในสัตว์มีกระดูกสันหลัง[ 32 ] การแก้ไข mRNA ของ ApoB เกิดขึ้นในสัตว์ เลี้ยงลูกด้วยนมที่มีรกทั้งหมด[ 33 ] การ แก้ไขเกิดขึ้นหลังการถอดรหัส เนื่องจากพอลินิวคลี โอไทด์ที่เกิดขึ้นใหม่ไม่มีนิวคลีโอไซด์ที่ได้รับการแก้ไข[ 34 ]
พิมพ์
การแก้ไข mRNA ของ ApoB จาก C เป็น U ต้องใช้คอมเพล็กซ์การแก้ไขหรือโฮโลเอนไซม์ (เอดิโทโซม) ซึ่งประกอบด้วยเอนไซม์แก้ไข mRNA ของ Apolipoprotein B, โพลีเปปไทด์เร่งปฏิกิริยา 1 (ApoBEC-1) รวมถึงปัจจัยเสริมอื่นๆ ApoBEC-1 เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนAPOBEC1 [ 35 ]เป็นสมาชิกของ ตระกูล ไซทิดีนดีอะมีเนส ApoBEC-1 เพียงอย่างเดียวไม่เพียงพอสำหรับการแก้ไข mRNA ของ ApoB [ 36 ]และต้องใช้ปัจจัยเสริมอย่างน้อยหนึ่งอย่าง คือปัจจัยเสริม APOBEC1 (A1CF) [ 37 ]เพื่อให้การแก้ไขเกิดขึ้น A1CF ประกอบด้วยส่วนที่ซ้ำกัน 3 ส่วน ทำหน้าที่เป็นหน่วยย่อยที่จับกับ RNA และนำทาง ApoBEC-1 ไปยัง mRNA ของ ApoB ที่อยู่ด้านล่างของไซทิดีนที่ได้รับการแก้ไข[ 38 ]ปัจจัยเสริมอื่นๆ เป็นที่ทราบกันว่าเป็นส่วนหนึ่งของโฮโลเอนไซม์ โปรตีนเหล่านี้บางส่วนได้รับการระบุแล้ว ได้แก่ โปรตีนที่จับกับ CUG 2 ( CUGBP2 ) [ 39 ] SYNCRIP (โปรตีนที่จับกับ RNA ที่อุดมไปด้วยไกลซีน-อาร์จินีน-ไทโรซีน, GRY-RBP) [ 40 ] ไรโบ นิวคลีโอโปรตีนนิวเคลียร์เฮเทอโรจีนัส (hnRNP)-C1 ( HNRNPC ) [ 41 ]โปรตีนที่จับกับ ApoBEC-1 ABBP1 ( HNRNPAB ), ABBP2 [ 42 ]โปรตีนที่จับกับตัวควบคุมการตัดต่อแบบ KH ( KHSRP ), Bcl-2-associated athanogene 4 ( BAG4 ) [ 43 ]และปัจจัยเสริม (AUX)240 [ 44 ]โปรตีนเหล่านี้ทั้งหมดได้รับการระบุโดยใช้การทดสอบการตรวจจับและได้รับการพิสูจน์แล้วว่ามีปฏิสัมพันธ์กับ ApoBEC-1, A1CF หรือ ApoB RNA หน้าที่ของโปรตีนเสริมเหล่านี้ในคอมเพล็กซ์การแก้ไขยังไม่เป็นที่ทราบ นอกจากการแก้ไข mRNA ของ ApoB แล้ว เอดิตโซม ApoBEC-1 ยังแก้ไข mRNA ของNF1ด้วย การแก้ไข mRNA ของ ApoB mRNA เป็นตัวอย่างที่ชัดเจนที่สุดของการแก้ไข RNA ประเภท C เป็น U ในมนุษย์
ที่ตั้ง
แม้ว่าจะเป็นทรานสคริปต์ที่มีความยาว 14,000 หน่วยย่อย แต่ไซทิดีนเพียงตัวเดียวก็ถูกกำหนดเป้าหมายสำหรับการแก้ไข ภายใน mRNA ของ ApoB พบลำดับที่ประกอบด้วยนิวคลีโอไทด์ 26 ตัวที่จำเป็นสำหรับการแก้ไข ซึ่งเรียกว่าโมทีฟการแก้ไข นิวคลีโอไทด์เหล่านี้ (6662–6687) ถูกกำหนดให้มีความสำคัญโดยการทดลองการกลายพันธุ์เฉพาะจุด[ 45 ]ส่วนของลำดับนี้ที่มีความยาว 11 นิวคลีโอไทด์ ซึ่งอยู่ห่างจากตำแหน่งการแก้ไขไปทางปลายน้ำ 4–5 นิวคลีโอไทด์ เป็นบริเวณสำคัญที่เรียกว่าลำดับการยึด[ 46 ]พบบริเวณที่เรียกว่าองค์ประกอบตัวเว้นวรรคที่อยู่ระหว่างนิวคลีโอไซด์ที่แก้ไขกับลำดับการยึดนี้ 2–8 นิวคลีโอไทด์[ 47 ]นอกจากนี้ยังมีลำดับควบคุมอยู่ทางด้าน 3′ ของตำแหน่งการแก้ไขด้วย เชื่อกันว่าบริเวณออกฤทธิ์ของ ApoBEC-1 ซึ่งเป็นส่วนประกอบเร่งปฏิกิริยาของเอนไซม์โฮโลเอนไซม์สำหรับการแก้ไข จะจับกับบริเวณที่อุดมไปด้วย AU ของลำดับการยึดเกาะ โดยอาศัย ACF ในการจับคอมเพล็กซ์กับ mRNA [ 48 ] หมู่ไซทิดีนที่ถูกแก้ไขจะอยู่ที่นิวคลีโอไทด์ 6666 ซึ่งอยู่ในเอ็กซอน 26 ของยีน การแก้ไขที่ตำแหน่งนี้ส่งผลให้เกิดการเปลี่ยนแปลงโคดอนจากโคดอนกลูตามีน (CAA) ไปเป็นโคดอนหยุด (UAA) ที่อยู่ในเฟรมเดียวกัน[ 32 ] การสร้างแบบจำลองด้วยคอมพิวเตอร์ตรวจพบว่าการแก้ไขจะเกิดขึ้นได้ก็ต่อเมื่อไซทิดีนที่ถูกแก้ไขอยู่ในลูป[ 46 ]การเลือกไซทิดีนที่ถูกแก้ไขยังขึ้นอยู่กับโครงสร้างทุติยภูมิของ RNA โดยรอบเป็นอย่างมาก นอกจากนี้ยังมีข้อบ่งชี้บางประการว่าบริเวณลูปนี้เกิดขึ้นระหว่างลำดับการยึดเกาะและบริเวณควบคุม 3′ ของ mRNA ของ ApoB [ 49 ]โครงสร้างทุติยภูมิที่คาดการณ์ไว้ซึ่งเกิดจาก mRNA ของ ApoB เชื่อกันว่าจะช่วยให้เกิดการสัมผัสระหว่างสารตกค้างที่จะได้รับการแก้ไขกับตำแหน่งที่ใช้งานของ APOBEC1 รวมถึงการจับกับ ACF และปัจจัยเสริมอื่นๆ ที่เกี่ยวข้องกับเอดิโทโซม
ระเบียบข้อบังคับ
การแก้ไข mRNA ของ ApoB ในมนุษย์นั้นถูกควบคุมโดยเนื้อเยื่อ โดย ApoB48 เป็นโปรตีน ApoB หลักของลำไส้เล็กในมนุษย์ พบในปริมาณที่น้อยกว่าในลำไส้ใหญ่ ไต และกระเพาะอาหาร พร้อมกับเวอร์ชันที่ไม่ได้รับการแก้ไข[ 50 ] การแก้ไขยังถูกควบคุมโดยการพัฒนา โดยเวอร์ชันที่ไม่ได้รับการแก้ไขจะถูกแปลเฉพาะในช่วงต้นของการพัฒนา แต่รูปแบบที่ได้รับการแก้ไขจะเพิ่มขึ้นในระหว่างการพัฒนาในเนื้อเยื่อที่สามารถเกิดการแก้ไขได้[ 51 ] [ 52 ] ระดับการแก้ไขของ mRNA ของ ApoB แสดงให้เห็นว่าแตกต่างกันไปตามการเปลี่ยนแปลงของอาหาร การสัมผัสกับแอลกอฮอล์ และระดับฮอร์โมน[ 53 ] [ 54 ] [ 55 ]
การอนุรักษ์
การแก้ไข mRNA ของ ApoB เกิดขึ้นในหนูและหนูแรตเช่นกัน ในทางตรงกันข้ามกับมนุษย์ การแก้ไขเกิดขึ้นในตับของหนูและหนูแรตด้วยความถี่สูงถึง 65% [ 56 ]ไม่พบในนกหรือสัตว์ชนิดที่เล็กกว่า[ 57 ]
ผลที่ตามมา
โครงสร้าง
การแก้ไขส่งผลให้เกิดการเปลี่ยนแปลงของโคดอน ทำให้เกิดโคดอนหยุดในเฟรม ส่งผลให้เกิดการแปลโปรตีนที่สั้นลง คือ ApoB48 โคดอนหยุดนี้ส่งผลให้เกิดการแปลโปรตีนที่ขาดปลายคาร์บอกซิล ซึ่งมีโดเมนการจับ LDLR ของโปรตีน โปรตีน ApoB100 ที่สมบูรณ์ ซึ่งมีกรดอะมิโนเกือบ 4500 ตัว พบได้ใน VLDL และ LDL เนื่องจากหลายส่วนของ ApoB100 อยู่ใน สภาวะ แอมฟิพาติกโครงสร้างของบางโดเมนจึงขึ้นอยู่กับสภาวะของไขมัน อย่างไรก็ตาม เป็นที่ทราบกันดีว่ามันมีการพับโดยรวมเหมือนกับ LDL โดยมีโดเมนหลักห้าโดเมน เมื่อเร็วๆ นี้ โครงสร้างแรกของ LDL ที่อุณหภูมิร่างกายมนุษย์ในสภาวะปกติถูกค้นพบโดยใช้กล้องจุลทรรศน์อิเล็กตรอนแบบไครโอที่ความละเอียด 16 อังสตรอม[ 58 ]การพับโดยรวมของ ApoB-100 ได้รับการยืนยันแล้ว และความไม่สม่เสมอในโครงสร้างเฉพาะที่ของโดเมนบางส่วนได้รับการทำแผนที่แล้ว
การทำงาน
การแก้ไขถูกจำกัดเฉพาะทรานสคริปต์ที่แสดงออกในลำไส้เล็กโปรตีนเวอร์ชันที่สั้นกว่านี้มีหน้าที่เฉพาะในลำไส้เล็ก หน้าที่หลักของ ApoB100 ที่แสดงออกใน ตับ แบบเต็มความยาว คือเป็นลิแกนด์สำหรับการกระตุ้น LDL-R อย่างไรก็ตาม การแก้ไขส่งผลให้โปรตีนขาดบริเวณการจับ LDL-R นี้ ซึ่งเปลี่ยนแปลงหน้าที่ของโปรตีนและโปรตีน ApoB48 ที่สั้นกว่า มีหน้าที่เฉพาะที่เกี่ยวข้องกับลำไส้เล็ก ApoB48 เหมือนกับ 48% ของปลายอะมิโนของ ApoB100 [ 59 ]หน้าที่ของไอโซฟอร์มนี้คือการดูดซึมไขมันของลำไส้เล็กและเกี่ยวข้องกับการสังเคราะห์ การประกอบ และการหลั่งไคโลไมครอน ไคโลไมครอนเหล่านี้ทำหน้าที่ขนส่งไขมันจากอาหารไปยังเนื้อเยื่อ ในขณะที่ไคโลไมครอนที่เหลืออยู่ พร้อมกับไขมันตกค้างที่เกี่ยวข้อง จะถูกดูดซึมโดยตับภายใน 2-3 ชั่วโมง ผ่านการทำงานร่วมกันของอะโพลิโปโปรตีนอี (ApoE) กับตัวรับไลโปโปรตีน ApoE เป็นโปรตีน ApoB ที่เด่นในลำไส้เล็กของสัตว์เลี้ยงลูกด้วยนมส่วนใหญ่ มันเป็นโปรตีนสำคัญในวิถีการเผาผลาญไลโปโปรตีนจากภายนอก โปรตีนในลำไส้ที่มี ApoB48 จะถูกเผาผลาญเป็นอนุภาคไคโลไมครอนที่เหลืออยู่ ซึ่งจะถูกดูดซึมโดยตัวรับที่เหลืออยู่
ดูเพิ่มเติม
อ่านเพิ่มเติม
- Mahley RW, Innerarity TL, Rall SC, Weisgraber KH (1985). "ไลโปโปรตีนในพลาสมา: โครงสร้างและหน้าที่ของอะโปไลโปโปรตีน" . J. Lipid Res . 25 (12): 1277– 1294. doi : 10.1016/S0022-2275(20)34443-6 . PMID 6099394 .
- อิทาคุระ เอช, มัตสึโมโตะ เอ (1995) "[อะโพลิโปโปรตีน บี]". นิปปอน รินโช . 52 (12): 3113– 3118. PMID 7853698 .
- Chumakova OS, Zateĭshchikov DA, Sidorenko BA (2006). "[Apolipoprotein B: โครงสร้าง หน้าที่ ความหลากหลายทางพันธุกรรม และความสัมพันธ์กับหลอดเลือดแดงแข็ง]". Kardiologiia . 45 (6): 43– 55. PMID 16007035 .
- Ye J (2007). "การพึ่งพาเส้นทางการเผาผลาญคอเลสเตอรอลของโฮสต์สำหรับวงจรชีวิตของไวรัสตับอักเสบซี" PLOS Pathog 3 ( 8) e108. doi : 10.1371/journal.ppat.0030108 . PMC 1959368 . PMID 17784784 .
ลิงก์ภายนอก
- ฐานข้อมูลการแก้ไข RNA (DARNED )
- การวิจัยประยุกต์เกี่ยวกับอะโพลิโปโปรตีน-บี
- หน้าแสดงตำแหน่งจีโนม APOBของมนุษย์และ รายละเอียดเกี่ยวกับยีน APOBในUCSC Genome Browser