อ่าน 13 นาที
การพัฒนาพืช
โครงสร้างที่สำคัญในการ พัฒนาของพืช ได้แก่ ตา หน่อ ราก ใบ และ ดอก พืชสร้างเนื้อเยื่อและโครงสร้างเหล่านี้ตลอดช่วงชีวิตจาก เนื้อเยื่อเจริญ [ 1 ] ซึ่งตั้งอยู่ที่ปลาย อวัยวะ...
การพัฒนาพืช
โครงสร้างที่สำคัญในการพัฒนาของพืชได้แก่ตาหน่อรากใบและดอกพืชสร้างเนื้อเยื่อและโครงสร้างเหล่านี้ตลอดช่วงชีวิตจากเนื้อเยื่อเจริญ[ 1 ]ซึ่งตั้งอยู่ที่ปลายอวัยวะหรือระหว่างเนื้อเยื่อที่เจริญเต็มที่ ดังนั้น พืชที่มีชีวิตจึงมีเนื้อเยื่อตัวอ่อนอยู่เสมอ ในทางตรงกันข้าม ตัวอ่อนของสัตว์จะสร้างส่วนต่างๆของร่างกายทั้งหมดที่จะมีในชีวิตของมันตั้งแต่เนิ่นๆ เมื่อสัตว์เกิดมา (หรือฟักออกจากไข่) มันจะมีส่วนต่างๆ ของร่างกายทั้งหมด และจากนั้นก็จะเติบโตใหญ่ขึ้นและเจริญเต็มที่มากขึ้น อย่างไรก็ตาม ทั้งพืชและสัตว์ต่างก็ผ่านระยะฟีโลไทป์ที่วิวัฒนาการอย่างอิสระ[ 2 ]ซึ่งก่อให้เกิดข้อจำกัดในการพัฒนาที่จำกัดความหลากหลายทางสัณฐานวิทยา[ 3 ] [ 4 ] [ 5 ] [ 6 ]
ตามที่นักสรีรวิทยาพืชA. Carl Leopoldกล่าวไว้ คุณสมบัติของการจัดระเบียบที่พบในพืชเป็นคุณสมบัติที่เกิดขึ้นใหม่ซึ่งมีมากกว่าผลรวมของส่วนประกอบแต่ละส่วน "การประกอบเนื้อเยื่อและหน้าที่เหล่านี้เข้าด้วยกันเป็นสิ่งมีชีวิตหลายเซลล์แบบ บูรณาการ ไม่เพียงแต่ให้ลักษณะเฉพาะของส่วนประกอบและกระบวนการที่แยกจากกันเท่านั้น แต่ยังให้ลักษณะเฉพาะชุดใหม่ที่ไม่สามารถคาดการณ์ได้จากการตรวจสอบส่วนประกอบที่แยกจากกัน" [ 7 ]
การเจริญเติบโต
พืชมีท่อลำเลียงเริ่ม ต้นจาก ไซโกตเซลล์เดียวที่เกิดจากการปฏิสนธิของเซลล์ไข่กับเซลล์อสุจิ จากนั้นเซลล์ไซโกตจะเริ่มแบ่งตัวเพื่อสร้างเอ็มบริโอ ของพืช ผ่านกระบวนการเกิดเอ็มบริโอเมื่อกระบวนการนี้เกิดขึ้น เซลล์ที่เกิดขึ้นจะจัดเรียงตัวเพื่อให้ปลายด้านหนึ่งกลายเป็นรากแรก ในขณะที่ปลายอีกด้านหนึ่งกลายเป็นปลายยอด ใน พืช มีเมล็ด เอ็มบริโอจะพัฒนาเป็น "ใบเลี้ยง" หนึ่งใบหรือมากกว่า ( cotyledons ) เมื่อสิ้นสุดกระบวนการเกิดเอ็มบริโอ พืชอ่อนจะมีส่วนประกอบทั้งหมดที่จำเป็นต่อการเริ่มต้นชีวิต[ 8 ]
การสร้างอวัยวะของพืช
เมื่อเอ็มบริโองอกออกมาจากเมล็ดหรือต้นแม่ มันจะเริ่มสร้างอวัยวะเพิ่มเติม (ใบ ลำต้น และราก) ผ่านกระบวนการสร้างอวัยวะรากใหม่จะงอกออกมาจากเนื้อเยื่อเจริญ ราก ที่อยู่ปลายราก และลำต้นและใบใหม่จะงอกออกมาจากเนื้อเยื่อ เจริญยอด ที่อยู่ปลายยอด[ 9 ]การแตกแขนงเกิดขึ้นเมื่อกลุ่มเซลล์เล็กๆ ที่เหลืออยู่จากเนื้อเยื่อเจริญ ซึ่งยังไม่ผ่านกระบวนการแบ่งเซลล์เพื่อสร้างเนื้อเยื่อเฉพาะ เริ่มเจริญเติบโตเป็นปลายรากหรือยอดใหม่ การเจริญเติบโตจากเนื้อเยื่อเจริญดังกล่าวที่ปลายรากหรือยอดเรียกว่าการเจริญเติบโตขั้นต้นและส่งผลให้รากหรือยอดนั้นยาวขึ้นการเจริญเติบโตขั้นทุติยภูมิส่งผลให้รากหรือยอดกว้างขึ้นจากการแบ่งเซลล์ในแคมเบียม[ 10 ]
การสร้างอวัยวะโดยตรง
การสร้างอวัยวะโดยตรง (Direct organogenesis) เป็นวิธี การเพาะเลี้ยง เนื้อเยื่อพืชชนิดหนึ่งที่อวัยวะต่างๆ เช่น รากและยอด พัฒนาขึ้นโดยตรงจากเซลล์เมริสเต็มหรือเซลล์ที่ไม่ใช่เมริสเต็ม โดยไม่ต้องผ่าน ขั้นตอนการสร้าง แคลลัสกระบวนการนี้เกิดขึ้นจากการกระตุ้นเมริสเต็มปลายยอดและปลายราก หรือตาข้างโดยได้รับอิทธิพลจากสารควบคุมการเจริญเติบโตของพืชทั้งภายในและภายนอกพืช ผลที่ได้คือ เซลล์ชนิดต่างๆ จะเกิดการเปลี่ยนแปลงไปเป็นโครงสร้างพืชที่สามารถเจริญเติบโตเป็นพืชทั้งต้นได้ เทคนิคนี้มักใช้ในการขยายพันธุ์พืชหลายชนิด รวมถึงผักผลไม้ไม้และพืชสมุนไพรโดยทั่วไปจะใช้ปลายยอดและส่วนข้อเป็นชิ้นส่วนเริ่มต้นในกระบวนการนี้ ในบางกรณี โครงสร้างที่เกิดขึ้นเองตามธรรมชาติอาจเกิดขึ้นจากเนื้อเยื่อร่างกายภายใต้สภาวะที่เฉพาะเจาะจง ทำให้เกิดการงอกใหม่ของยอดหรือรากในบริเวณที่ไม่สามารถเจริญเติบโตได้ตามธรรมชาติ แนวทางนี้มีประสิทธิภาพเป็นพิเศษในพืชล้มลุก และในขณะที่การสร้างใหม่แบบไม่ปกติอาจนำไปสู่การสร้างยอดในอัตราที่สูงขึ้น การขยายพันธุ์ยอดข้างยังคงเป็นวิธีการที่ใช้กันอย่างแพร่หลายที่สุดในการขยายพันธุ์แบบจุลภาคเนื่องจากมีประสิทธิภาพและใช้งานได้จริง ลำดับทั่วไปของการพัฒนาอวัยวะในกระบวนการนี้เป็นไปตามรูปแบบ: ชิ้นส่วนเริ่มต้น → เนื้อเยื่อเจริญ → ต้นอ่อนของอวัยวะ[ 11 ]
การสร้างอวัยวะทางอ้อม
การสร้างอวัยวะทางอ้อมเป็นกระบวนการพัฒนาที่เซลล์พืช undergo dedifferentiationทำให้เซลล์สามารถกลับคืนสู่สภาวะเฉพาะและเปลี่ยนไปสู่เส้นทางการพัฒนาใหม่ กระบวนการนี้มีลักษณะเฉพาะคือระยะแคลลัสขั้นกลาง ซึ่งเซลล์จะสูญเสียเอกลักษณ์ดั้งเดิมและปรับตัวทางสัณฐานวิทยาได้ ทำหน้าที่เป็นพื้นฐานสำหรับการสร้างอวัยวะ ความก้าวหน้าของการสร้างอวัยวะทางอ้อมเกี่ยวข้องกับหลายขั้นตอนสำคัญ เริ่มต้นด้วย dedifferentiation ซึ่งทำให้เซลล์มีความสามารถ ตามด้วยระยะการเหนี่ยวนำที่นำไปสู่สภาวะที่กำหนดอย่างสมบูรณ์ เมื่อบรรลุการกำหนดแล้ว เซลล์จะ undergo การเปลี่ยนแปลงทางสัณฐานวิทยา ในที่สุดจะก่อให้เกิดยอดหรือรากที่ใช้งานได้ กระบวนการนี้เป็นไปตามลำดับการพัฒนาที่มีโครงสร้าง: เอ็กซ์แพลนต์หลัก → แคลลัส → เมริสเตมอยด์ → พริมอร์เดียมอวัยวะ ทำให้มั่นใจได้ว่าการสร้างอวัยวะของพืชเป็นไปอย่างเป็นระเบียบ[ 12 ]
ปัจจัยที่มีผลต่อการสร้างอวัยวะ
การปลูกถ่าย
ความสามารถในการสร้างพืชใหม่ได้อย่างประสบความสำเร็จขึ้นอยู่กับการเลือกชิ้นส่วนพืชที่เหมาะสม ซึ่งแตกต่างกันไปตามชนิดและสายพันธุ์ของพืช ในการสร้างอวัยวะโดยตรง มักนิยมใช้ชิ้นส่วนพืชจากเนื้อเยื่อเจริญ เช่น ยอดอ่อน ตาข้าง ใบ ก้าน ใบราก และโครงสร้างดอก เนื่องจากมีความสามารถในการพัฒนาเป็นอวัยวะใหม่ได้อย่างรวดเร็ว เนื้อเยื่อเหล่านี้มีอัตราการรอดชีวิตสูง เจริญเติบโตเร็ว และมีศักยภาพในการสร้างใหม่ในหลอดทดลองสูง เนื้อเยื่อเจริญ ยอดอ่อน ตาข้าง ใบอ่อน และเอ็มบริโอ มีประสิทธิภาพเป็นพิเศษในการส่งเสริมการสร้างใหม่ในพืชหลากหลายชนิด
นอกจากนี้ ส่วนต่างๆ ของพืชที่เจริญเต็มที่แล้ว เช่น ใบ ลำต้น ราก ก้านใบ และกลีบดอก สามารถใช้เป็นชิ้นส่วนพืชเริ่มต้น (explant) ที่มีชีวิตสำหรับการสร้างอวัยวะใหม่ได้ภายใต้สภาวะที่เหมาะสม การงอกใหม่ของพืชเกิดขึ้นผ่านการสร้างแคลลัส ซึ่งเป็นกลุ่มเซลล์ที่ยังไม่แบ่งตัว และต่อมาจะพัฒนาไปเป็นอวัยวะใหม่ การสร้างแคลลัสสามารถกระตุ้นได้จากชิ้นส่วนพืชเริ่มต้นต่างๆ เช่นใบเลี้ยงลำต้นใบยอดอ่อนรากช่อดอกและโครงสร้างดอก เมื่อนำมาเพาะเลี้ยงภายใต้สภาวะที่ควบคุม ได้
โดยทั่วไปแล้ว เนื้อเยื่อที่ประกอบด้วยเซลล์ที่กำลังแบ่งตัวอย่างแข็งขันจะมีประสิทธิภาพมากกว่าในการเริ่มต้นการสร้างแคลลัส เนื่องจากมีศักยภาพในการปรับเปลี่ยนโปรแกรมของเซลล์ได้สูงกว่า เนื้อเยื่อที่ยังไม่เจริญเต็มที่มักจะปรับตัวได้ดีกว่าสำหรับการสร้างใหม่เมื่อเทียบกับเนื้อเยื่อที่เจริญเต็มที่แล้ว เนื่องจากมีความยืดหยุ่นในการพัฒนาที่มากขึ้น ขนาดและรูปร่างของเนื้อเยื่อก็มีอิทธิพลต่อความสำเร็จในการสร้างวัฒนธรรมเช่นกัน เนื่องจากเนื้อเยื่อที่มีขนาดใหญ่กว่าหรือมีโครงสร้างที่เหมาะสมกว่าอาจเพิ่มโอกาสในการอยู่รอดและการเจริญเติบโต การพัฒนาของแคลลัสส่วนใหญ่ถูกกระตุ้นโดยการบาดเจ็บและการมีอยู่ของฮอร์โมนพืช ซึ่งอาจมีอยู่ตามธรรมชาติในเนื้อเยื่อหรือเสริมในอาหารเลี้ยงเชื้อเพื่อกระตุ้นกิจกรรมของเซลล์และการสร้างอวัยวะ[ 13 ]
อาหารเลี้ยงเชื้อ สารควบคุมการเจริญเติบโตของพืช และสารก่อเจล
องค์ประกอบของอาหารเลี้ยงเชื้อมีความแตกต่างกันอย่างมากในด้านธาตุแร่และปริมาณวิตามิน เพื่อรองรับความต้องการของพืชหลากหลายชนิด อาหาร เลี้ยงเชื้อ Murashige and Skoog (MS) มีลักษณะเด่นคือมีปริมาณไนโตรเจนในรูปแอมโมเนียมสูง ซึ่งเป็นลักษณะที่ไม่พบในสูตรอื่นๆ โดยทั่วไปแล้วซูโครสจะทำหน้าที่เป็นแหล่งคาร์โบไฮเดรตหลักในอาหารเลี้ยงเชื้อประเภทต่างๆ[ 14 ]
ปฏิสัมพันธ์ระหว่างออกซินและไซโตไคนินในการควบคุมการสร้างอวัยวะได้รับการพิสูจน์แล้ว แม้ว่าการตอบสนองจะแตกต่างกันไปตามชนิดของพืช พืชบางชนิด เช่น ยาสูบ สามารถสร้างตาหน่อได้เองโดยไม่ต้องใช้สารควบคุมการเจริญเติบโตจากภายนอก ในขณะที่พืชชนิดอื่น เช่นScurrula pulverulenta , Lactuca sativaและBrassica junceaจำเป็นต้องเสริมฮอร์โมนอย่างเคร่งครัด ในการเพาะเลี้ยงใบเลี้ยงของ B. juncea เบนซิล อะมิโนพิวรีน (BAP) เพียงอย่างเดียวสามารถกระตุ้นการสร้างหน่อจากเนื้อเยื่อก้านใบได้ คล้ายกับต้นสนเรเดียตาที่ไซโตไคนินเพียงอย่างเดียวก็เพียงพอสำหรับการกระตุ้นการสร้างหน่อ[ 15 ]
งานวิจัยชี้ให้เห็นว่าความเข้มข้นของฮอร์โมนภายในร่างกายมากกว่าระดับการใช้ฮอร์โมนจากภายนอก เป็นตัวกำหนดการสร้างอวัยวะในท้ายที่สุด ในบรรดาไซโตไคนิน ต่างๆ (2iP, BAP, thidiazuron, kinetin และ zeatin) ที่ใช้ในการชักนำให้เกิดยอด BAP แสดงให้เห็นถึงประสิทธิภาพที่เหนือกว่าและมีการใช้งานอย่างแพร่หลาย ออกซินก็มีอิทธิพลต่อกระบวนการสร้างอวัยวะเช่นกัน โดย 2,4-D มักใช้ในการชักนำให้เกิดแคลลัสในธัญพืช แม้ว่าการสร้างอวัยวะโดยทั่วไปจะต้องย้ายไปยังอาหารเลี้ยงเชื้อที่มีIAAหรือNAAหรือไม่มี 2,4-D เลยก็ตาม อัตราส่วนของออกซินต่อไซโตไคนินเป็นตัวกำหนดส่วนใหญ่ว่าอวัยวะใดจะพัฒนาขึ้น
กรดจิบเบอเรลลิก (GA3) มีส่วนช่วยในการยืดตัวของเซลล์และการสร้างเนื้อเยื่อเจริญ ในขณะที่สารประกอบที่ไม่ธรรมดา เช่น กรดไตรไอโอโดเบนโซอิก (TIBA) กรดแอบซิสิก (ABA) คานามัยซิน และสารยับยั้งออกซิน ได้รับการพิสูจน์แล้วว่ามีประสิทธิภาพสำหรับพืชที่งอกยาก สารเติมแต่งจากธรรมชาติ เช่น ผง โสมสามารถเพิ่มความถี่ในการงอกใหม่ในบางการเพาะเลี้ยง เนื่องจากเอทิลีนมักจะยับยั้งการสร้างยอด สารยับยั้งการสังเคราะห์เอทิลีน เช่นอะมิโนเอทอกซีไวนิลไกลซีน (AVG) และซิลเวอร์ไนเตรต (AgNO3 )จึงมักถูกนำมาใช้เพื่อส่งเสริมการสร้างอวัยวะ โดยมีเอกสารยืนยันความสำเร็จในการเพาะเลี้ยง ข้าวสาลียาสูบและทานตะวัน
อะการ์ไม่ใช่ส่วนประกอบที่จำเป็นของอาหารเลี้ยงเชื้อ แต่คุณภาพและปริมาณของอะการ์เป็นปัจจัยสำคัญที่อาจกำหนดบทบาทในการสร้างอวัยวะ อะการ์ที่วางจำหน่ายในเชิงพาณิชย์อาจมีสิ่งเจือปน หากมีความเข้มข้นของอะการ์สูง อาหารเลี้ยงเชื้อจะแข็งตัวและขัดขวางการแพร่กระจายของสารอาหารไปยังเนื้อเยื่อที่กำลังเจริญเติบโต ส่งผลต่อกระบวนการสร้างอวัยวะโดยการสร้างรากพิเศษแคลลัสที่ไม่พึงประสงค์ที่โคนต้น หรือใบเหี่ยวเฉา ค่าpHเป็นอีกปัจจัยสำคัญที่อาจส่งผลต่อเส้นทางการสร้างอวัยวะ ค่า pH ของอาหารเลี้ยงเชื้อจะถูกปรับให้อยู่ระหว่าง 5.6 ถึง 5.8 ก่อนการฆ่าเชื้อ ค่า pH ของอาหารเลี้ยงเชื้อช่วยส่งเสริมหรือยับยั้งการดูดซึมสารอาหารในอาหารเลี้ยงเชื้อ ตัวอย่างเช่น การดูดซึมแอมโมเนียมในหลอดทดลองเกิดขึ้นที่ค่า pH คงที่ที่ 5.5 (Thorpe et al., 2008)
ปัจจัยอื่นๆ
ฤดูกาลของปี
ช่วงเวลาในการเก็บชิ้นส่วนพืชมีผลอย่างมากต่อความสามารถในการสร้างใหม่ในระบบเพาะเลี้ยงเนื้อเยื่อ โดยการเปลี่ยนแปลงตามฤดูกาลมีบทบาทสำคัญต่อความสำเร็จในการสร้างอวัยวะ ปรากฏการณ์นี้แสดงให้เห็นอย่างชัดเจนในลิลลี่ (Lilium speciosum ) ซึ่งเกล็ดหัวแสดงการตอบสนองในการสร้างใหม่ที่แตกต่างกันไปตามฤดูกาลในการเก็บ ชิ้นส่วนพืชที่เก็บในช่วงฤดูใบไม้ผลิและฤดูใบไม้ร่วงสามารถสร้างหัวเล็กๆ ได้ง่ายในหลอดทดลอง ในขณะที่ชิ้นส่วนพืชที่เก็บในช่วงฤดูร้อนหรือฤดูหนาวไม่สามารถสร้างหัวเล็กๆ ได้แม้จะมีสภาวะการเพาะเลี้ยงที่เหมือนกันก็ตาม
พบความสัมพันธ์ตามฤดูกาลที่คล้ายคลึงกันในChlorophytum borivillianumซึ่งเป็นพืชที่มีคุณค่าทางยา โดยแสดงให้เห็นว่าการสร้างหัวในหลอดทดลองเพิ่มขึ้นอย่างเห็นได้ชัดใน ช่วงฤดู มรสุมเมื่อเทียบกับช่วงเวลาอื่น ๆ ของปี ความแปรผันตามฤดูกาลในศักยภาพในการสร้างรูปร่างนี้อาจสะท้อนถึงความแตกต่างในสภาวะทางสรีรวิทยาของพืชต้นกำเนิด รวมถึงระดับฮอร์โมนภายใน ปริมาณคาร์โบไฮเดรตสำรอง และกิจกรรมการเผาผลาญที่ผันผวนตลอดวงจรการเจริญเติบโตประจำปี
ความแตกต่างของระดับออกซิเจน
ออกซิเจนมีบทบาทสำคัญในการเพาะเลี้ยงเนื้อเยื่อ ซึ่งส่งผลต่อการสร้างอวัยวะ ในการเพาะเลี้ยงบางชนิด การสร้างตาหน่อจะเกิดขึ้นเมื่อความเข้มข้นของออกซิเจนที่มีอยู่ภายในภาชนะเพาะเลี้ยงลดลง ในขณะที่การเหนี่ยวนำให้เกิดรากต้องอาศัยความเข้มข้นของออกซิเจนสูง[ 16 ]
แสงสว่าง
สภาพ แสงทั้งความเข้มและคุณภาพสเปกตรัม มีบทบาทสำคัญในการกระตุ้นการสร้างรูปร่างในระบบเพาะเลี้ยงเนื้อเยื่อพืช การวิจัยเกี่ยวกับองค์ประกอบสเปกตรัมเผยให้เห็นการตอบสนองที่แตกต่างกันตามความยาวคลื่น โดยทั่วไปแสงสีฟ้าจะส่งเสริมการสร้างยอด ในขณะที่แสงสีแดงมักส่งเสริมการเหนี่ยวนำให้เกิดราก การให้แสงตามลำดับ—แสงสีฟ้าตามด้วยแสงสีแดง—ได้รับการบันทึกไว้ว่าสามารถกระตุ้นกระบวนการสร้างอวัยวะเฉพาะในพืชบางชนิดได้อย่างมีประสิทธิภาพ
ผลกระทบด้านการควบคุมของความยาวคลื่นที่แตกต่างกันแสดงให้เห็นว่าคุณภาพของแสงสามารถควบคุมผลลัพธ์ทางด้านสัณฐานวิทยาได้อย่างเลือกสรรแสงไฟฟลูออ เรสเซนต์เทียม ก่อให้เกิดการตอบสนองที่แตกต่างกันไปขึ้นอยู่กับชนิดของพืช โดยส่งเสริมการสร้างรากในบางชนิดในขณะที่ยับยั้งการสร้างรากในชนิดอื่น พืชบางชนิดมีความต้องการแสงที่เฉพาะเจาะจง ดังที่พบใน ถั่วลันเตา ( Pisum sativum ) ซึ่งการเริ่มต้นของตาหน่อเกิดขึ้นได้ดีที่สุดในสภาพแวดล้อมที่มืด ก่อนที่การได้รับแสงจะกระตุ้นการพัฒนาต่อไป
สำหรับการเพาะเลี้ยงเนื้อเยื่อส่วนใหญ่ โปรโตคอลการให้แสงมาตรฐานมักแนะนำให้ใช้แสงที่มีความเข้มประมาณ 2,000-3,000 ลักซ์ โดยมีช่วงเวลาการให้แสง 16 ชั่วโมง อย่างไรก็ตาม บางชนิดมีความต้องการความเข้มของแสงสูงเป็นพิเศษ ตัวอย่างเช่น การ เพาะเลี้ยงเนื้อเยื่อแคลลัสของยาสูบ ( Nicotiana tabacum ) ซึ่งต้องการความเข้มของแสงสูงกว่ามากถึง 10,000-15,000 ลักซ์ เพื่อกระตุ้นการสร้างหน่อหรือการเกิดเอ็มบริโอจากเซลล์ร่างกาย
อุณหภูมิ
อุณหภูมิเป็นปัจจัยด้านสิ่งแวดล้อมที่สำคัญอย่างยิ่งในระบบการเพาะเลี้ยงเนื้อเยื่อพืช โดยอุณหภูมิการบ่มที่เหมาะสมจะแตกต่างกันอย่างมากในแต่ละชนิด ขึ้นอยู่กับความต้องการในถิ่นที่อยู่ตามธรรมชาติของพืชนั้นๆ แม้ว่า 25 องศาเซลเซียสจะเป็นอุณหภูมิการบ่มมาตรฐานที่เหมาะสมสำหรับพืชหลายชนิดในการเพาะเลี้ยงในหลอดทดลอง แต่ควรพิจารณาการปรับอุณหภูมิให้เหมาะสมกับแต่ละชนิดเพื่อเพิ่มศักยภาพในการสร้างอวัยวะให้สูงสุด
โดยทั่วไปแล้ว พืชหัวที่ขึ้นในเขตอบอุ่นต้องการอุณหภูมิที่ต่ำกว่ามาตรฐานที่กำหนดไว้ ตัวอย่างที่เห็นได้ชัด ได้แก่ พืชหัว เช่นดอกกาแลนทัส (ดอกสโนว์ดรอป) ซึ่งเจริญเติบโตได้ดีที่สุดที่อุณหภูมิประมาณ 15 องศาเซลเซียส ในขณะที่บางสายพันธุ์ของดอกนาร์ซิสซัส (ดอกแดฟโฟดิล) และ หัวหอมประดับ ( อัลเลียม ) แสดงประสิทธิภาพการงอกใหม่ที่ดีขึ้นที่อุณหภูมิประมาณ 18 องศาเซลเซียส
ในทางตรงกันข้าม พืชที่มีถิ่นกำเนิดในเขตร้อนโดยทั่วไปต้องการอุณหภูมิที่สูงขึ้นเพื่อการเจริญเติบโตและการสร้างอวัยวะที่ดีที่สุดในสภาพการเพาะ เลี้ยง ต้นปาล์มอินทผลัมเจริญเติบโตได้ดีที่อุณหภูมิ 27 องศาเซลเซียส ในขณะที่ ต้น มอนสเตรา เดลิซิ โอซา (ต้นชีสสวิส) แสดงประสิทธิภาพการงอกใหม่สูงสุดที่อุณหภูมิ 30 องศาเซลเซียส ความต้องการอุณหภูมิเหล่านี้สะท้อนถึงการปรับตัวตามวิวัฒนาการของพืชให้เข้ากับสภาพแวดล้อมดั้งเดิมของพวกมัน
ระดับพลอยดี
ความแปรปรวนของจำนวนโครโมโซมเช่น แอนยูพลอยดี โพลีพลอยดี เป็นต้น ในการเพาะเลี้ยงเซลล์พืชนั้นได้รับการบันทึกไว้เป็นอย่างดีในอดีต ความไม่เสถียรของโครโมโซมในเซลล์ส่งผลให้ศักยภาพในการสร้างรูปร่างของเนื้อเยื่อแคลลัสลดลงเรื่อยๆ ดังนั้น เพื่อรักษาศักยภาพในการสร้างอวัยวะของเนื้อเยื่อแคลลัสและความเสถียรของโครโมโซม จึงแนะนำให้ปฏิบัติตามระยะเวลาและความถี่ของการเพาะเลี้ยงย่อยอย่างสม่ำเสมอ
ยุคแห่งวัฒนธรรม
อายุของวัฒนธรรมมักเป็นกุญแจสำคัญสู่ความสำเร็จในการสร้างอวัยวะ วัฒนธรรมที่อายุน้อย/วัสดุที่เพาะเลี้ยงใหม่อาจสร้างอวัยวะได้บ่อยกว่าวัฒนธรรมที่อายุมาก เหตุผลที่เป็นไปได้คือการลดลงหรือการสูญเสียศักยภาพในการสร้างอวัยวะในวัฒนธรรมที่เก่า อย่างไรก็ตาม ในพืชบางชนิด ความสามารถในการสร้างใหม่ของพืชอาจคงอยู่ได้ไม่จำกัดเป็นเวลาหลายปี[ 17 ]
กระบวนการพัฒนา
การลดความแตกต่าง
ความสามารถของเซลล์ในการสร้างอวัยวะขึ้นอยู่กับการใช้สารควบคุมการเจริญเติบโตของพืช (PGRs) เป็นอย่างมาก ซึ่งมีอิทธิพลต่อทิศทางการพัฒนาของเนื้อเยื่อ ความสมดุลระหว่างออกซินและไซโตไคนินมีบทบาทสำคัญในการกำหนดว่าหน่อหรือรากจะเกิดขึ้น อัตราส่วนออกซินต่อไซโตไคนินที่ต่ำกว่าจะส่งเสริมการสร้างหน่อ ในขณะที่ความเข้มข้นของออกซินที่สูงกว่าจะส่งเสริมการสร้างราก ตัวอย่างเช่น ใน การเพาะเลี้ยง Medicago sativa (อัลฟัลฟา) ระดับคิเนตินที่สูงขึ้นร่วมกับความเข้มข้นต่ำของ 2,4-D (ออกซินสังเคราะห์) จะนำไปสู่การพัฒนาของหน่อ ในขณะที่การเพิ่ม 2,4-D ในขณะที่ลดความเข้มข้นของคิเนตินจะส่งเสริมการสร้างราก อย่างไรก็ตาม การสร้างอวัยวะที่ประสบความสำเร็จไม่ได้ขึ้นอยู่กับการรักษาด้วย PGR เพียงอย่างเดียวแคลลัสหรือเนื้อเยื่อที่กำลังพัฒนาจะต้องมีขนาดที่สำคัญก่อนที่การสร้างอวัยวะจะดำเนินต่อไปได้ ซึ่งสะท้อนให้เห็นถึงบทบาทของการส่งสัญญาณระหว่างเซลล์ในการประสานการพัฒนา[ 18 ]
การเหนี่ยวนำ
ระยะการเหนี่ยวนำในการสร้างอวัยวะแสดงถึงช่วงเปลี่ยนผ่านระหว่างเนื้อเยื่อที่บรรลุความสามารถและถูกกำหนดอย่างสมบูรณ์เพื่อเริ่มต้นการสร้างไพรมอร์เดีย ในระหว่างขั้นตอนนี้ เส้นทางทางพันธุกรรมแบบบูรณาการจะชี้นำกระบวนการพัฒนา ก่อนที่จะเกิดการแยกความแตกต่างทางสัณฐานวิทยา ปัจจัยทางเคมีและทางกายภาพสามารถรบกวนเส้นทางการพัฒนาที่ถูกกำหนดโดยพันธุกรรมและเปลี่ยนแปลงผลลัพธ์ทางสัณฐานวิทยา ในConvolvulus arvensisอิทธิพลดังกล่าวจะยับยั้งการสร้างยอดและนำไปสู่การพัฒนาแคลลัสแทน[ 19 ]
การสิ้นสุดของระยะการเหนี่ยวนำจะถูกทำเครื่องหมายโดยเซลล์หรือกลุ่มเซลล์ที่กำหนดให้เกิดการสร้างยอดหรือราก การกำหนดนี้ได้รับการทดสอบโดยการย้ายเนื้อเยื่อจากอาหารเลี้ยงเชื้อที่เสริมด้วยสารควบคุมการเจริญเติบโตไปยังอาหารเลี้ยงเชื้อพื้นฐานที่มีแร่ธาตุ วิตามิน และแหล่งคาร์บอนที่จำเป็น แต่ไม่มีสารควบคุมการเจริญเติบโตของพืช ในขั้นตอนนี้ เนื้อเยื่อจะเสร็จสิ้นกระบวนการเหนี่ยวนำและกำหนดชะตากรรมการพัฒนาของมันอย่างสมบูรณ์[ 20 ]
แนวคิดสำคัญในกระบวนการนี้คือการสร้างช่องทางซึ่งหมายถึงความสามารถของเส้นทางการพัฒนาในการสร้างฟีโนไทป์ ที่สม่ำเสมอ แม้จะมีความแปรปรวนทางพันธุกรรมหรือสิ่งแวดล้อม หากนำชิ้นส่วนพืชออกจากอาหารเลี้ยงเชื้อที่กระตุ้นการสร้างยอดก่อนที่การสร้างช่องทางจะเสร็จสมบูรณ์ การสร้างยอดจะลดลงอย่างรวดเร็วและการพัฒนาของรากจะเด่นกว่า ซึ่งสะท้อนถึงความยืดหยุ่นทางสัณฐานวิทยาของเนื้อเยื่อพืชในหลอดทดลอง[ 21 ]
ความแตกต่าง
การแบ่งแยกทางสัณฐานวิทยาเริ่มต้นในระยะนี้ นำไปสู่การก่อตัวของอวัยวะที่เพิ่งเกิดขึ้น การเริ่มต้นของการสร้างอวัยวะถูกทำเครื่องหมายด้วยการเปลี่ยนแปลงขั้วของเซลล์ตามด้วยการสร้างสมมาตรแบบรัศมีและการเติบโตตามแกนที่กำหนดใหม่ ทำให้เกิดส่วนนูนของโครงสร้างที่เป็นสัญญาณของการเริ่มต้นของอวัยวะ[ 22 ]
การพัฒนาตามลำดับของการสร้างอวัยวะสามารถสังเกตได้ในสายพันธุ์เช่นPinus oocarpa Schiedeซึ่งตาหน่อจะงอกใหม่โดยตรงจากใบเลี้ยงผ่านการสร้างอวัยวะโดยตรง อย่างไรก็ตาม รูปแบบการพัฒนาที่เฉพาะเจาะจงอาจแตกต่างกันไปในสายพันธุ์พืชต่าง ๆ ที่ปลูกในหลอดทดลอง ความก้าวหน้าของการสร้างอวัยวะรวมถึงการเปลี่ยนแปลงทางสัณฐานวิทยาที่ชัดเจน เริ่มต้นด้วยการเปลี่ยนแปลงในพื้นผิว การเกิดขึ้นของเมริสเตมอยด์ และการขยายตัวของ บริเวณ เมริสเตมทั้งในแนวตั้งหรือแนวนอน ตามมาด้วยการยื่นของบริเวณเมริสเตมออกไปนอกชั้นผิวหนัง การสร้างเมริสเตมที่มีโครงสร้างพร้อมใบอ่อนที่มองเห็นได้ และในที่สุด การพัฒนาอย่างสมบูรณ์ของตาที่เกิดขึ้นเอง[ 23 ]
ลักษณะเด่นอย่างหนึ่งของการเพาะเลี้ยงอวัยวะในหลอดทดลองคือการก่อตัวของเมริสเตมอยด์หลายอันพร้อมกันบนชิ้นส่วนเดียว โดยมีระดับการแยกแยะที่แตกต่างกัน ภายในชิ้นส่วนเดียวกัน อาจมีตาอยู่ในระยะการพัฒนาที่แตกต่างกัน ตั้งแต่เริ่มแรกจนถึงโครงสร้างที่พัฒนาเต็มที่ เมื่อหน่อมีความยาวเกินประมาณ 1 ซม. จะถูกย้ายไปยังวัสดุปลูกรากในหลอดทดลองหรือนอกหลอดทดลอง เพื่อ สร้างต้นกล้า ให้สมบูรณ์ [ 24 ]
ข้อดีและข้อจำกัด
ในการสร้างอวัยวะโดยตรงหน่อข้างเกิดขึ้นโดยตรงจากเนื้อเยื่อเจริญที่มีอยู่แล้วที่ปลายหน่อและข้อ ทำให้มีอัตราการขยายพันธุ์สูง เนื่องจากเนื้อเยื่อเจริญของหน่อที่จัดระเบียบแล้วแทบจะไม่สะสมการกลายพันธุ์ พืชที่ได้จึงมีความสม่ำเสมอทางพันธุกรรมและตรงตามสายพันธุ์ กล่าวคือ เป็นโคลน ทางพันธุกรรม ของพ่อแม่ ทำให้เทคนิคนี้มีประโยชน์อย่างยิ่งสำหรับการขยายพันธุ์และการอนุรักษ์พันธุ์พืชที่มีความสำคัญทางเศรษฐกิจและสิ่งแวดล้อม[ 25 ]
อย่างไรก็ตาม การสร้างอวัยวะมีข้อจำกัดบางประการความแปรปรวนของโซมาโคลนซึ่งอาจส่งผลให้เกิดความหลากหลายทางพันธุกรรมที่ไม่พึงประสงค์ เป็นปัญหาที่อาจเกิดขึ้นได้ โดยเฉพาะอย่างยิ่งในกระบวนการสร้างอวัยวะทางอ้อม นอกจากนี้ เทคนิคนี้อาจไม่เหมาะสมสำหรับ พืช ที่ดื้อต่อการรักษาซึ่งก็คือพืชที่ไม่ตอบสนองต่อการเพาะเลี้ยงในหลอดทดลองหรือโปรโตคอลการสร้างใหม่ได้ดี[ 26 ]ข้อจำกัดเหล่านี้เน้นย้ำถึงความจำเป็นในการวิจัยอย่างต่อเนื่องและการปรับปรุงวิธีการสำหรับพืชชนิดต่างๆ เพื่อเอาชนะความท้าทายเหล่านี้ใน การขยายพันธุ์ และการอนุรักษ์พืช[ 27 ]
การยืดตัวของเซลล์
นอกจากการเจริญเติบโตโดย การแบ่ง เซลล์แล้ว พืชยังสามารถเจริญเติบโตได้ด้วยการยืดตัวของเซลล์ซึ่งเกิดขึ้นเมื่อเซลล์แต่ละเซลล์หรือกลุ่มเซลล์เจริญเติบโตยาวขึ้น ไม่ใช่ว่าเซลล์พืชทุกเซลล์จะเจริญเติบโตจนมีความยาวเท่ากัน เมื่อเซลล์ด้านหนึ่งของลำต้นเจริญเติบโตยาวและเร็วกว่าเซลล์อีกด้านหนึ่ง ลำต้นจะโค้งงอไปทางด้านของเซลล์ที่เจริญเติบโตช้ากว่า การเจริญเติบโตแบบมีทิศทางนี้สามารถเกิดขึ้นได้จากการตอบสนองของพืชต่อสิ่งเร้าเฉพาะ เช่น แสง ( โฟโตโทรปิซึม ) แรงโน้มถ่วง ( กราวิโทรปิซึม ) น้ำ ( ไฮโดรโทรปิซึม ) และการสัมผัสทางกายภาพ ( ทิกโมโทรปิซึม ) [ 28 ]


การเจริญเติบโตและการพัฒนาของพืชถูกควบคุมโดยฮอร์โมนพืชและสารควบคุมการเจริญเติบโตของพืช (PGRs) ที่เฉพาะเจาะจง (Ross et al. 1983) [ 29 ]ระดับฮอร์โมนภายในได้รับอิทธิพลจากอายุของพืช ความทนทานต่อความหนาวเย็น การพักตัว และสภาวะการเผาผลาญอื่นๆช่วงเวลาแสงความชื้น อุณหภูมิ และสภาพแวดล้อมภายนอกอื่นๆ และแหล่งที่มาของ PGRs จากภายนอก เช่น ที่ใช้จากภายนอกและจากแหล่งกำเนิดในไรโซสเฟียร์[ 30 ]
การเปลี่ยนแปลงทางสัณฐานวิทยาในระหว่างการเจริญเติบโต
พืชแสดงความแปรผันตามธรรมชาติในรูปทรงและโครงสร้าง ในขณะที่สิ่งมีชีวิตทั้งหมดมีความแปรผันจากตัวหนึ่งไปยังอีกตัวหนึ่ง พืชยังแสดงความแปรผันอีกประเภทหนึ่งด้วย ภายในตัวเดียวกันนั้น ส่วนต่างๆ จะถูกทำซ้ำซึ่งอาจมีรูปทรงและโครงสร้างที่แตกต่างจากส่วนอื่นๆ ที่คล้ายคลึงกัน ความแปรผันนี้สามารถมองเห็นได้ง่ายที่สุดในใบของพืช แม้ว่าอวัยวะอื่นๆ เช่น ลำต้นและดอกอาจแสดงความแปรผันที่คล้ายกัน สาเหตุหลักสามประการของความแปรผันนี้ ได้แก่ ผลกระทบจากตำแหน่ง ผลกระทบจากสิ่งแวดล้อม และความอ่อนตัว[ 31 ]

มีความแตกต่างกันระหว่างส่วนต่างๆ ของพืชที่โตเต็มที่อันเนื่องมาจากตำแหน่งที่อวัยวะถูกสร้างขึ้น ตัวอย่างเช่น ใบตามกิ่งใหม่ อาจมีความแตกต่างกันในรูปแบบที่สม่ำเสมอตลอดกิ่ง รูปทรงของใบที่เกิดขึ้นใกล้โคนกิ่งจะแตกต่างจากใบที่เกิดขึ้นที่ปลายกิ่ง และความแตกต่างนี้จะสม่ำเสมอจากกิ่งหนึ่งไปยังอีกกิ่งหนึ่งในพืชชนิดเดียวกัน[ 32 ]
วิธีการที่โครงสร้างใหม่เจริญเติบโตนั้นอาจได้รับผลกระทบจากช่วงเวลาในวงจรชีวิตของพืชเมื่อเริ่มพัฒนา รวมถึงสภาพแวดล้อมที่โครงสร้างเหล่านั้นสัมผัส อุณหภูมิมีผลกระทบต่อพืชหลายด้าน ขึ้นอยู่กับปัจจัยหลายอย่าง รวมถึงขนาดและสภาพของพืช อุณหภูมิ และระยะเวลาที่สัมผัส พืชที่มีขนาดเล็กและอวบน้ำ มาก จะมีความเสี่ยงต่อความเสียหายหรือการตายจากอุณหภูมิที่สูงหรือต่ำเกินไปมากกว่า อุณหภูมิส่งผลต่ออัตราของกระบวนการทางชีวเคมีและสรีรวิทยา โดยทั่วไปอัตราเหล่านี้ (ภายในขอบเขตที่กำหนด) จะเพิ่มขึ้นตามอุณหภูมิ
ความเยาว์วัยหรือเฮเทอโรบลาสตีคือเมื่ออวัยวะและเนื้อเยื่อที่ผลิตโดยพืชอายุน้อย เช่นต้นกล้ามักจะแตกต่างจากที่ผลิตโดยพืชชนิดเดียวกันเมื่อโตขึ้น ตัวอย่างเช่น ต้นไม้อายุน้อยจะผลิตกิ่งที่ยาวและเรียวกว่าซึ่งเจริญเติบโตขึ้นด้านบนมากกว่ากิ่งที่จะผลิตเมื่อต้นไม้โตเต็มที่ นอกจากนี้ ใบที่ผลิตในช่วงการเจริญเติบโตในระยะแรกมักจะมีขนาดใหญ่กว่า บางกว่า และไม่สม่ำเสมอมากกว่าใบของพืชที่โตเต็มที่ ตัวอย่างของพืชอายุน้อยอาจมีลักษณะแตกต่างจากพืชที่โตเต็มที่ของสายพันธุ์เดียวกันอย่างสิ้นเชิงจนแมลงที่วางไข่ไม่รู้จักพืชนั้นว่าเป็นอาหารสำหรับลูกของมัน การเปลี่ยนจากรูปแบบการเจริญเติบโตในระยะแรกไปสู่ระยะหลังบางครั้งเรียกว่าการเปลี่ยนแปลงระยะการเจริญเติบโตทางพืช[ 33 ]
โครงสร้างที่เกิดขึ้นโดยบังเอิญ
โครงสร้างของพืช เช่นรากตาและหน่อ ที่เจริญเติบโตในตำแหน่ง ที่ไม่ปกติ เรียกว่าเนื้อเยื่อพิเศษ (adventitious )
รากและตาที่เกิดขึ้นเองมักจะพัฒนาใกล้กับเนื้อเยื่อหลอดเลือดที่มีอยู่เพื่อให้สามารถเชื่อมต่อกับไซเล็มและโฟลเอมได้อย่างไรก็ตาม ตำแหน่งที่แน่นอนจะแตกต่างกันอย่างมาก ในลำต้นอ่อน รากที่เกิดขึ้นเองมักจะเกิดขึ้นจากพาเรนไคมาระหว่างมัดหลอดเลือดในลำต้นที่มีการเจริญเติบโตทุติยภูมิ รากที่เกิดขึ้นเองมักจะเกิดขึ้นในพาเรนไคมาของโฟลเอมใกล้กับแคมเบียมหลอดเลือด ในการปักชำลำต้น บางครั้งรากที่เกิดขึ้นเองยังเกิดขึ้นใน เซลล์ แคลลัสที่เกิดขึ้นที่ผิวตัด การปักชำใบของCrassulaจะสร้างรากที่เกิดขึ้นเองในชั้นเอ พิเดอร์มิส [ 34 ]
ตาและหน่อ
ตาที่เกิดขึ้น เองตามธรรมชาติ (Adventitious buds)เกิดขึ้นจากบริเวณอื่นที่ไม่ใช่เนื้อเยื่อเจริญปลายยอด (shoot apical meristem ) ซึ่งอยู่บริเวณปลายลำต้น หรือบริเวณข้อของลำต้น หรือซอกใบ โดยตาเหล่านี้จะยังคงอยู่ ณ ที่นั้นระหว่างการเจริญเติบโตขั้นต้น ตาเหล่านี้อาจเกิดขึ้นที่ราก ใบ หรือลำต้นที่งอกใหม่ เนื้อเยื่อเจริญปลายยอดจะสร้างตาข้าง (axillary bud) หรือตาด้านข้าง (lateral bud) หนึ่งตาหรือมากกว่านั้นที่แต่ละข้อ เมื่อลำต้นมีการเจริญเติบโตขั้นทุติยภูมิมาก ตาข้างอาจถูกทำลายไป จากนั้นตาที่เกิดขึ้นเองตามธรรมชาติอาจเจริญเติบโตบนลำต้นที่มีการเจริญเติบโตขั้นทุติยภูมิ
ตาที่เกิดขึ้นเองตามธรรมชาติ มักเกิดขึ้นหลังจากลำต้นได้รับบาดเจ็บหรือถูกตัดแต่งกิ่ง ตาเหล่านี้ช่วยทดแทนกิ่งที่หายไป นอกจากนี้ ตาและหน่อที่เกิดขึ้นเองตามธรรมชาติยังอาจเกิดขึ้นบนลำต้นของต้นไม้ที่โตเต็มที่ เมื่อลำต้นที่อยู่ในที่ร่มถูกแสงแดดจัด เนื่องจากต้นไม้รอบข้างถูกตัดลง ต้นเรดวูด ( Sequoia sempervirens ) มักมีตาที่เกิดขึ้นเองตามธรรมชาติจำนวนมากที่ส่วนล่างของลำต้น หากลำต้นหลักตายไป ลำต้นใหม่มักจะแตกหน่อออกมาจากตาที่เกิดขึ้นเองตามธรรมชาติเหล่านั้น ชิ้นส่วนเล็กๆ ของลำต้นเรดวูดถูกนำมาขายเป็นของที่ระลึก เรียกว่า ปุ่มเรดวูด (redwood burls) โดยนำไปแช่ในถาดน้ำ แล้วตาที่เกิดขึ้นเองตามธรรมชาติจะงอกออกมาเป็นหน่อ
พืชบางชนิดมักสร้างตาพิเศษบนราก ซึ่งสามารถยื่นออกมาจากต้นได้ไกลพอสมควร หน่อที่งอกออกมาจากตาพิเศษบนรากเรียกว่า หน่ออ่อนหน่ออ่อนเป็นรูปแบบการขยายพันธุ์แบบไม่อาศัยเพศ ตามธรรมชาติ ในหลายชนิดเช่น หญ้าหลายชนิดต้นแอสเพนสั่นและต้นธิสเซิลแคนาดา ต้นแอสเพนสั่นแพนโดเติบโตจากลำต้นเดียวเป็น 47,000 ลำต้นโดยการสร้างตาพิเศษบนระบบรากเดียว
ใบของต้นไม้บางชนิดจะแตกหน่อพิเศษ ซึ่งต่อมาจะกลายเป็นรากพิเศษ เป็นส่วนหนึ่งของการขยายพันธุ์แบบไม่อาศัยเพศเช่น ต้นไม้เลื้อย ( Tolmiea menziesii ) และต้นไม้พันต้น ( Kalanchoe daigremontiana ) ต้นอ่อนที่แตกหน่อพิเศษเหล่านี้จะหลุดออกจากต้นแม่และเจริญเติบโตเป็นโคลน แยกต่างหาก จากต้นแม่
การตัดแต่งกิ่ง แบบโคปปิ้งคือการตัด ลำต้น ของต้นไม้ลงจนถึงพื้นดินเพื่อกระตุ้นการเจริญเติบโตอย่างรวดเร็วของหน่อใหม่ โดยทั่วไปแล้วจะใช้ในการผลิตเสา รั้ว หรือฟืน นอกจากนี้ยังใช้กับ พืช ชีวมวลที่ปลูกเพื่อเป็นเชื้อเพลิง เช่นต้นป็อปลาร์หรือต้นวิลโลว์ด้วย
ราก

การงอกรากพิเศษอาจเป็นการปรับตัวเพื่อหลีกเลี่ยงความเครียดสำหรับบางชนิด โดยได้รับแรงกระตุ้นจากปัจจัยต่างๆ เช่นภาวะขาดออกซิเจน[ 35 ]หรือการขาดสารอาหาร หน้าที่ทางนิเวศวิทยาที่สำคัญอีกประการหนึ่งของการงอกรากพิเศษคือการสืบพันธุ์แบบไม่อาศัยเพศของต้นไม้บางชนิด เช่นSalixและSequoiaในสภาพแวดล้อมริมแม่น้ำ[ 36 ]
ความสามารถของลำต้นพืชในการสร้างรากพิเศษถูกนำมาใช้ในการขยายพันธุ์เชิงพาณิชย์โดยการปักชำความเข้าใจเกี่ยวกับกลไกทางสรีรวิทยาเบื้องหลังการเกิดรากพิเศษทำให้มีความก้าวหน้าในการปรับปรุงการปักชำโดยการใช้สารออกซินสังเคราะห์เป็นผงเร่งรากและการใช้การบาดเจ็บที่โคนต้นแบบเลือกเฉพาะ[ 37 ]ความก้าวหน้าเพิ่มเติมสามารถเกิดขึ้นได้ในอนาคตโดยการนำงานวิจัยเกี่ยวกับกลไกการควบคุมอื่นๆ มาใช้ในการขยายพันธุ์เชิงพาณิชย์ และโดยการวิเคราะห์เปรียบเทียบการควบคุมระดับโมเลกุลและทางนิเวศวิทยาของการเกิดรากพิเศษในสายพันธุ์ที่ "ปักชำยาก" เทียบกับสายพันธุ์ที่ "ปักชำง่าย"
รากและตาที่เกิดขึ้นเองตามธรรมชาติมีความสำคัญมากเมื่อทำการขยายพันธุ์พืชด้วยการปักชำการตอนกิ่งหรือ การเพาะ เลี้ยงเนื้อเยื่อฮอร์โมนพืชที่เรียกว่าออกซินมักถูกนำมาใช้กับกิ่ง ลำต้นหรือใบที่ปักชำเพื่อกระตุ้นการเกิดรากที่เกิดขึ้นเองตามธรรมชาติ เช่น ใบของ แอฟริกันไวโอเล็ตและซีดัมและลำต้นของพอยน์เซตเทียและ โคเลีย ส การขยายพันธุ์ด้วยการปักชำรากต้องอาศัยการเกิดตาที่เกิดขึ้นเองตามธรรมชาติ เช่น ในฮอร์สแรดิชและแอปเปิลในการตอนกิ่ง รากที่เกิดขึ้นเองตามธรรมชาติจะเกิดขึ้นบนลำต้นเหนือดินก่อนที่จะตัดส่วนของลำต้นออกเพื่อทำเป็นต้นใหม่ ไม้ประดับขนาดใหญ่มักขยายพันธุ์ด้วยการตอนกิ่งแบบอากาศรากและตาที่เกิดขึ้นเองตามธรรมชาติจะต้องพัฒนาขึ้นในการขยายพันธุ์พืชด้วยการเพาะเลี้ยงเนื้อเยื่อ
แบบฟอร์มที่แก้ไขแล้ว
- รากหัวไม่มีรูปร่างที่แน่นอน ตัวอย่างเช่นมันเทศ
- รากรวมกลุ่ม (รากหัว) เกิดขึ้นเป็นกลุ่มๆ ที่โคนลำต้น ตัวอย่างเช่นหน่อไม้ฝรั่งดอกดาเลีย
- รากที่มีปุ่มนูนจะบวมขึ้นบริเวณปลายราก ตัวอย่างเช่นขมิ้น
- รากค้ำยันงอกออกมาจากข้อแรกๆ ของลำต้น รากเหล่านี้จะแทงลงไปในดินในแนวเฉียงและช่วยพยุงต้นพืชตัวอย่างเช่นข้าวโพดอ้อย
- รากค้ำยันช่วยพยุงกิ่งเหนือดิน กิ่งด้านข้างจะเจริญเติบโตลงไปในดินในแนวตั้งและทำหน้าที่เหมือนเสาหลัก ตัวอย่างเช่นต้นไทร
- รากเลื้อยที่งอกออกมาจากข้อจะยึดเกาะกับสิ่งรองรับและเลื้อยขึ้นไปตามนั้น ตัวอย่างเช่นEpipremnum aureum
- รากที่มีลักษณะเป็นลูกปัดหรือเป็นพวง รากอวบน้ำทำให้มี ลักษณะเป็นลูกปัด เช่นมะระและผักเบี้ยใหญ่
การพัฒนาของใบ
พันธุกรรมที่อยู่เบื้องหลังการพัฒนาของรูปทรงใบในArabidopsis thalianaถูกแบ่งออกเป็นสามขั้นตอน ได้แก่ การเริ่มต้นของใบอ่อนการสร้างลักษณะ ด้านบน และด้านล่าง ของใบ และการพัฒนาของเนื้อเยื่อเจริญ ขอบ ใบ การเริ่มต้นของใบอ่อนเกิดจากการยับยั้งยีนและโปรตีนในกลุ่มKNOX คลาส I (เช่นSHOOT APICAL MERISTEMLESS ) โปรตีน KNOX คลาส I เหล่านี้จะยับยั้ง การสังเคราะห์ จิบเบอเรลลินในใบอ่อนโดยตรง พบว่ามีปัจจัยทางพันธุกรรมหลายอย่างที่เกี่ยวข้องกับการยับยั้งยีนเหล่านี้ในใบอ่อน (เช่นASYMMETRIC LEAVES1, BLADE-ON-PETIOLE1 , SAWTOOTH1เป็นต้น) ดังนั้น ด้วยการยับยั้งนี้ ระดับของจิบเบอเรลลินจึงเพิ่มขึ้นและใบอ่อนจึงเริ่มเจริญเติบโต
การพัฒนาของดอกไม้

การพัฒนาของดอกไม้คือกระบวนการที่พืชดอกสร้างรูปแบบการแสดงออกของยีนในเนื้อเยื่อเจริญซึ่งนำไปสู่การปรากฏของอวัยวะที่มุ่งเน้นการสืบพันธุ์แบบอาศัยเพศ นั่น คือ ดอกไม้มีพัฒนาการทางสรีรวิทยา 3 ขั้นตอนที่ต้องเกิดขึ้นเพื่อให้กระบวนการนี้เกิดขึ้นได้: ประการแรก พืชต้องเปลี่ยนจากระยะที่ยังไม่เจริญพันธุ์ไปสู่ระยะที่เจริญพันธุ์แล้ว (กล่าวคือ การเปลี่ยนผ่านไปสู่การออกดอก) ประการที่สอง การเปลี่ยนแปลงหน้าที่ของเนื้อเยื่อเจริญปลายยอดจากเนื้อเยื่อเจริญส่วนลำต้นไปเป็นเนื้อเยื่อเจริญส่วนดอกหรือช่อ ดอก และประการสุดท้าย การเจริญเติบโตของอวัยวะแต่ละส่วนของดอกไม้ ขั้นตอนสุดท้ายนี้ได้รับการจำลองโดยใช้แบบจำลอง ABCซึ่งอธิบายพื้นฐานทางชีววิทยาของกระบวนการจากมุมมองของพันธุศาสตร์ ระดับโมเลกุลและ พันธุศาสตร์เชิง พัฒนาการ
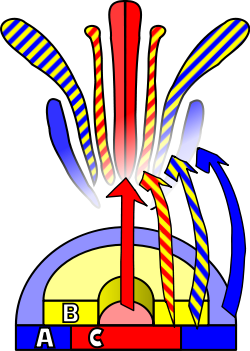
จำเป็นต้องมี สิ่งกระตุ้นภายนอกเพื่อกระตุ้นให้เนื้อเยื่อเจริญเปลี่ยน ไปเป็นเนื้อเยื่อเจริญของดอกไม้ สิ่งกระตุ้นนี้จะกระตุ้นการแบ่งเซลล์ แบบไมโทซิสในเนื้อเยื่อเจริญ โดยเฉพาะอย่างยิ่งบริเวณด้านข้างซึ่ง เป็นที่เกิดของ ปุ่มเนื้อเยื่อเจริญ ใหม่ สิ่งกระตุ้นเดียวกันนี้ยังจะทำให้เนื้อเยื่อเจริญพัฒนาไปตามแบบแผนที่นำไปสู่การเจริญเติบโตของเนื้อเยื่อเจริญดอกไม้ แทนที่จะเป็นเนื้อเยื่อเจริญส่วนลำต้น ความแตกต่างหลักระหว่างเนื้อเยื่อเจริญทั้งสองชนิดนี้ นอกเหนือจากความแตกต่างที่เห็นได้ชัดระหว่างอวัยวะเป้าหมายแล้ว คือการเรียง ตัวเป็นวง (หรือเป็นเกลียว) กล่าวคือ ไม่มี การยืด ตัวของลำต้น ระหว่าง วงหรือเกลียวที่ต่อเนื่องกันของปุ่มเนื้อเยื่อเจริญ เกลียวเหล่านี้จะพัฒนาจากล่างขึ้นบน ทำให้เกิดกลีบเลี้ยงกลีบดอกเกสรตัวผู้และเกสรตัวเมีย ความแตกต่างอีกประการหนึ่งจากเนื้อเยื่อเจริญข้างลำต้นของพืชคือเนื้อเยื่อเจริญของดอกนั้น "ถูกกำหนดแล้ว" ซึ่งหมายความว่าเมื่อเซลล์ของมันแยกตัวแล้ว เซลล์เหล่านั้นจะไม่แบ่งตัว อีกต่อ ไป[ 38 ]
เอกลักษณ์ของอวัยวะที่อยู่ในวงกลีบดอกทั้งสี่วงนั้น เป็นผลมาจากการทำงานร่วมกันของผลิตภัณฑ์ยีน อย่างน้อยสามชนิด ซึ่งแต่ละชนิดมีหน้าที่แตกต่างกัน ตามแบบจำลอง ABC หน้าที่ A และ C จำเป็นต่อการกำหนดเอกลักษณ์ของวงกลีบดอกและวงกลีบดอกสืบพันธุ์ตามลำดับ หน้าที่เหล่านี้เป็นหน้าที่เฉพาะ และหากขาดหน้าที่ใดหน้าที่หนึ่ง หน้าที่อีกหน้าที่หนึ่งจะกำหนดเอกลักษณ์ของวงกลีบดอกทั้งหมด หน้าที่ B ช่วยให้เกิดการแยกแยะกลีบดอกออกจากกลีบเลี้ยงในวงกลีบดอกรอง และการแยกแยะเกสรตัวผู้จากเกสรตัวเมียในวงกลีบดอกที่สาม
กลิ่นหอมของดอกไม้
พืชใช้รูปร่างดอก ดอกไม้ และกลิ่นเพื่อดึงดูดแมลงชนิดต่างๆ สำหรับการผสมเกสรสารประกอบบางชนิดในกลิ่นที่ปล่อยออกมาจะดึงดูดแมลง ผสมเกสรบางชนิด ในPetunia hybrida จะมีการผลิต เบนซีนอยด์ระเหยเพื่อให้เกิดกลิ่นดอกไม้ แม้ว่าส่วนประกอบของเส้นทางการสังเคราะห์เบนซีนอยด์จะเป็นที่รู้จัก แต่เอนไซม์ในเส้นทางดังกล่าวและการควบคุมเอนไซม์เหล่านั้นในภายหลังยังคงต้องได้รับการค้นพบ[ 39 ]
เพื่อศึกษาการควบคุมวิถีการสังเคราะห์สารระเหยจึงใช้ดอกP. hybrida Mitchell ใน ไมโครอาร์เรย์ เฉพาะกลีบดอก เพื่อเปรียบเทียบดอกที่กำลังจะปล่อยกลิ่นหอม กับ ดอก P. hybridaพันธุ์ W138 ที่ผลิตสารเบนซีนระเหยได้น้อย จากนั้นจึงทำการลำดับเบสของยีนในพืชทั้งสองชนิด ผลการศึกษาพบว่ามีปัจจัยการถอดรหัสที่ถูกกระตุ้นให้มีการแสดงออกเพิ่มขึ้นในดอก Mitchell แต่ไม่พบในดอก W138 ที่ไม่มีกลิ่นหอม จึงตั้งชื่อยีนนี้ว่า ODORANT1 (ODO1) เพื่อตรวจสอบการแสดงออกของ ODO1 ตลอดทั้งวัน จึง ทำการ วิเคราะห์ด้วยวิธี RNA gel blot ผลการวิเคราะห์แสดงให้เห็นว่าระดับการถอดรหัสของ ODO1 เริ่มเพิ่มขึ้นระหว่างเวลา 13:00 ถึง 16:00 น. สูงสุดที่ 22:00 น. และต่ำสุดที่ 10:00 น. ระดับการถอดรหัสของ ODO1 เหล่านี้สอดคล้องโดยตรงกับช่วงเวลาของการปล่อยสารเบนซีนระเหย นอกจากนี้ เจลยังสนับสนุนการค้นพบก่อนหน้านี้ที่ว่าดอกไม้ W138 ที่ไม่มีกลิ่นหอมมีระดับการถอดรหัส ODO1 เพียงหนึ่งในสิบของดอกไม้ Mitchell ดังนั้น ปริมาณของ ODO1 ที่สร้างขึ้นจึงสอดคล้องกับปริมาณของเบนซีนอยด์ระเหยที่ปล่อยออกมา ซึ่งบ่งชี้ว่า ODO1 ควบคุมการสังเคราะห์เบนซีนอยด์[ 39 ]
ยีนเพิ่มเติมที่มีส่วนช่วยในการสังเคราะห์สารประกอบกลิ่นหลัก ได้แก่ OOMT1 และ OOMT2 OOMT1 และ OOMT2 ช่วยในการสังเคราะห์เอนไซม์ออร์ซิโนล โอ-เมทิลทรานสเฟอเรส (OOMT) ซึ่งเป็นตัวเร่งปฏิกิริยาในสองขั้นตอนสุดท้ายของกระบวนการสังเคราะห์ DMT ทำให้เกิด 3,5-ไดเมทอกซีโทลูอีน (DMT) DMT เป็นสารประกอบกลิ่นที่ผลิตโดยกุหลาบหลายชนิด แต่กุหลาบบางสายพันธุ์ เช่นRosa gallicaและกุหลาบดามัสก์Rosa damasceneไม่ปล่อย DMT ออกมา มีการสันนิษฐานว่าสายพันธุ์เหล่านี้ไม่สร้าง DMT เนื่องจากไม่มีจีน OOMT อย่างไรก็ตาม จากการทดลองหาตำแหน่งทางภูมิคุ้มกัน พบว่า OOMT อยู่ในชั้นผิวของกลีบดอก เพื่อศึกษาเพิ่มเติม จึงนำกลีบกุหลาบไปปั่นเหวี่ยงด้วยความเร็วสูง จาก นั้นจึงตรวจสอบสารละลายส่วนบนและส่วนล่างด้วยวิธีเวสเทิร์นบลอต การตรวจพบโปรตีน OOMT ที่ 150,000g ในสารละลายส่วนบนและตะกอนทำให้ผู้วิจัยสรุปได้ว่าโปรตีน OOMT เกี่ยวข้องอย่างใกล้ชิดกับเยื่อหุ้มผิวกลีบดอก การทดลองดังกล่าวได้กำหนดว่ายีน OOMT มีอยู่ในกุหลาบRosa gallicaและกุหลาบดามัสก์Rosa damasceneแต่ยีน OOMT ไม่ได้แสดงออกในเนื้อเยื่อดอกไม้ที่สร้าง DMT [ 40 ]
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การพัฒนาพืช
โครงสร้างที่สำคัญในการ พัฒนาของพืช ได้แก่ ตา หน่อ ราก ใบ และ ดอก พืชสร้างเนื้อเยื่อและโครงสร้างเหล่านี้ตลอดช่วงชีวิตจาก เนื้อเยื่อเจริญ [ 1 ] ซึ่งตั้งอยู่ที่ปลาย อวัยวะ...
การเจริญเติบโต
พืชมีท่อลำเลียง เริ่ม ต้นจาก ไซโกต เซลล์เดียวที่เกิดจาก การปฏิสนธิ ของเซลล์ไข่กับเซลล์อสุจิ จากนั้นเซลล์ไซโกตจะเริ่มแบ่งตัวเพื่อสร้าง เอ็มบริโอ ของพืช ผ่านกระบวนการ เกิดเอ็มบริโอ เมื่อกระบวนการนี้เกิดขึ้น...
การสร้างอวัยวะของพืช
เมื่อเอ็มบริโอ งอกออกมา จากเมล็ดหรือต้นแม่ มันจะเริ่มสร้างอวัยวะเพิ่มเติม (ใบ ลำต้น และราก) ผ่านกระบวนการ สร้างอวัยวะ รากใหม่จะงอกออกมาจาก เนื้อเยื่อเจริญ ราก ที่อยู่ปลายราก และลำต้นและใบใหม่จะงอกออกมาจาก เนื้อเยื่อ เจริญยอด ที่อยู่ปลายยอด [ 9 ]...
การสร้างอวัยวะโดยตรง
การสร้างอวัยวะโดยตรง (Direct organogenesis) เป็นวิธี การเพาะเลี้ยง เนื้อเยื่อพืชชนิดหนึ่ง ที่อวัยวะต่างๆ เช่น รากและยอด พัฒนาขึ้นโดยตรงจากเซลล์เมริสเต็มหรือเซลล์ที่ไม่ใช่เมริสเต็ม โดยไม่ต้องผ่าน ขั้นตอนการสร้าง แคลลัส...