อ่าน 13 นาที
เมืองต่างๆ
Citipes ( / ˈ s ɪ t i p ɛ z / ) เป็นสกุลของเทโรพอดCaenagnathid ที่ สูญพันธุ์ไปแล้ว จากยุคครีเทเชียสตอนปลาย ใน Dinosaur Park...
เมืองต่างๆ
| เมืองต่างๆ ช่วงเวลา: ปลายยุคครีเทเชียสประมาณ 76.9 ถึง 75.8 ล้านปีที่แล้ว - | |
|---|---|
 | |
| แผนภาพแสดงวัสดุทั้งหมดที่ทราบจากแหล่งโบราณคดีซิติเปสโดยวัสดุที่ได้รับการอธิบายในปี 2020 แสดงด้วยสีชมพู | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลื้อยคลาน |
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ |
| กลุ่มสายพันธุ์ : | ซอริสเชีย |
| กลุ่มสายพันธุ์ : | เทโรโปดา |
| ตระกูล: | † Caenagnathidae |
| อนุวงศ์: | † เอลมิซอรีนา (?) |
| ประเภท: | † เมืองฟันสตัน, 2020 |
| ชนิดต้นแบบ | |
| † ออร์นิโทมิมัส อีเลแกนส์ สวนสาธารณะ , 1933 | |
| สายพันธุ์ | |
| |
| คำพ้องความหมาย | |
| |
Citipes ( / ˈ s ɪ t i p ɛ z / ) เป็นสกุลของเทโรพอดCaenagnathid ที่ สูญพันธุ์ไปแล้ว จากยุคครีเทเชียสตอนปลาย ใน Dinosaur Park Formationในอัลเบอร์ตาประเทศแคนาดาสกุลนี้มีเพียงชนิดเดียว คือชนิดต้นแบบC. elegansชื่อสกุล Citipes มาจากภาษาละติน แปลว่า "เท้าว่องไว" และ ชื่อ ชนิด " elegans " มาจาก ภาษาละติน แปลว่า "สง่างาม" [ 1 ] ตัวอย่างต้นแบบของ Citipesมี ประวัติ ทางอนุกรมวิธาน ที่ซับซ้อน และก่อนหน้า นี้เคยถูกจัดอยู่ในสกุล Ornithomimus [ 2 ] Macrophalangia [ 3 ] Elmisaurus [ 4 ] Chirostenotes [ 5 ]และ Leptorhynchos [ 6 ]ก่อนที่จะได้รับสกุลของตัวเองในปี 2020 [ 1 ]
การค้นพบและการตั้งชื่อ
การค้นพบเบื้องต้น

วัสดุที่ต่อมาได้รับการตั้งชื่อว่าCitipesถูกค้นพบในปี 1926 ระหว่าง การสำรวจของ มหาวิทยาลัยโทรอนโตในพื้นที่ Sand Creek ของแม่น้ำ Red Deerสถานที่แห่งนี้เป็นส่วนหนึ่งของกลุ่ม Belly Riverซึ่งปัจจุบันได้รับการยอมรับว่าเป็นDinosaur Park Formationอย่างไรก็ตาม ต้องใช้เวลาอีกเจ็ดปีกว่าที่คำอธิบายฉบับเต็มของวัสดุนี้จะได้รับการตีพิมพ์ใน วารสารธรณีวิทยาของ มหาวิทยาลัยโทรอนโตในปี 1933 โดย ดร. William A. Parks [ 6 ] วัสดุนี้ประกอบด้วยกระดูกฝ่าเท้า สามชิ้น ถูกเก็บรักษาไว้ที่พิพิธภัณฑ์ Royal Ontarioและได้รับการกำหนดหมายเลขROM 781 [ 7 ]
มันถูกตั้งชื่อเป็นสปีชีส์ใหม่ของสกุลOrnithomimus ทั่วไป — Ornithomimus elegansชื่อนี้ตั้งขึ้นโดยอิงจากกระดูกฝ่าเท้า สามชิ้น ซึ่งเชื่อว่าเป็นของออร์นิโทมิโมซอร์เนื่องจากไม่มีกระดูกฝ่าเท้าชิ้นที่ห้า[ 2 ]โอวิแรปโทโรซอเรียยังไม่ได้รับการยอมรับว่าเป็นกลุ่มสายพันธุ์เฉพาะ และโอวิแรปโทแรนหลายชนิด รวมถึงOviraptor [ 8 ] และ Chirostenotes [ 9 ] เชื่อว่าเป็นออร์นิโทมิมิด จนกระทั่งปี 1976 โอวิแรปโทโรซอรัสจึงได้รับการยอมรับว่าเป็นกลุ่มสายพันธุ์เฉพาะของตนเอง[ 10 ]
ประวัติการจำแนกประเภท
ROM 781 ถูกจัดให้อยู่ในสกุลMacrophalangia ที่น่าสงสัย โดย Dale Russell ในปี 1972 [ 3 ] อย่างไรก็ตาม สกุลนี้ถูกจัดให้เป็นชื่อพ้องรองของChirostenotesในอีกไม่กี่ปีต่อมา[ 11 ]
การตรวจสอบ ROM 781 อย่างละเอียดถี่ถ้วนยิ่งขึ้นได้ดำเนินการในปี 1988 โดยPhilip J. Currieซึ่งได้ตีพิมพ์คำอธิบายใหม่ของวัสดุในวารสาร Canadian Journal of Earth Sciences Currie จัดให้ตัวอย่างดังกล่าวเป็นสปีชีส์ใหม่ในสกุลElmisaurus ที่เพิ่งได้รับการอธิบายเมื่อไม่นานมานี้ คือElmisaurus elegansนอกจากนี้เขายังจัดให้ตัวอย่างใหม่สองตัวอย่างอยู่ในสปีชีส์นี้ด้วย ตัวอย่างใหม่ชิ้นแรกคือ ROM 37163 ซึ่งประกอบด้วยกระดูกฝ่าเท้า บางส่วน และถูกเก็บรวบรวมในช่วงระหว่างปี 1920 ถึง 1954 โดยคณะสำรวจของพิพิธภัณฑ์Royal Ontario Museumตัวอย่างที่สองคือ TMP 82.39.4 ซึ่งประกอบด้วยกระดูกฝ่าเท้าและกระดูกข้อเท้า บางส่วนอีกชิ้นหนึ่ง และถูกเก็บรวบรวมในปี 1982 โดย Linda Strong-Watson ตัวอย่างทั้งสองชิ้นนี้ถูกค้นพบในDinosaur Park Formationและมีลักษณะคล้ายกับตัวอย่างต้นแบบมากพอที่จะจัดให้อยู่ในส ปีชีส์เดียวกัน [ 4 ]
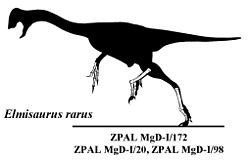

Elmisaurus elegansแตกต่างจากชนิดต้นแบบ E. rarusโดยมีสันที่อ่อนกว่าบนกระดูกฝ่าเท้าชิ้นที่สี่และมีส่วนยื่นที่ปลายสุดของกระดูกฝ่าเท้าชิ้นที่ II และ IV ในขณะที่ตีพิมพ์เอกสารนี้ มีการถกเถียงกันอย่างต่อเนื่องเกี่ยวกับความถูกต้องของสกุลCaenagnathid Chirostenotes , CaenagnathusและElmisaurusและ Currie ใช้การบรรยายลักษณะใหม่ของ ROM 781 เพื่อโต้แย้งว่าElmisaurusเป็นสกุลที่แตกต่างและถูกต้อง[ 4 ]
การจำแนกประเภทของ ROM 781 ได้รับการแก้ไขอีกครั้งในปี 1997 โดย Hans-Dieter Sues เมื่อเขาตีพิมพ์บทความวิจัยฉบับสมบูรณ์ที่อธิบายถึงตัวอย่างที่เพิ่งค้นพบใหม่ของสกุลChirostenotesในบทความวิจัยของเขา Sues ได้จัดให้Elmisaurus elegansอยู่ในสกุลChirostenotesในฐานะสปีชีส์ใหม่C. elegansและเขายังถือว่าสกุลCaenagnathusและElmisaurusเป็นชื่อพ้องรองของChirostenotesตัวอย่างบางส่วนถูกจัดให้อยู่ในC. pergracilisและบางส่วนอยู่ในC. elegans ที่ตั้งขึ้นใหม่ Sues แยกแยะC. elegansจากC. pergracilisโดยการมีกระดูกเชื่อมติดกันของกระดูกฝ่าเท้าที่สองและสี่ และมันถูกรวมอยู่ในสกุลนี้เพราะ Sues ถือว่าChirostenotes เป็นสกุล Caenagnathid ที่ถูกต้องเพียง สกุลเดียวในCampanian Laramidia [ 5 ]
ในปี 2013 พร้อมกับการบรรยายลักษณะของสกุลLeptorhynchosนั้น Nicholas Longrich, Ken Barnes, Scott Clark และ Larry Millar ได้จัดให้ ROM 781 อยู่ในสกุลใหม่ที่พวกเขาเพิ่งบรรยายขึ้น เหตุผลของการจัดกลุ่มใหม่นี้คือ การจัดกลุ่มตัวอย่างเพิ่มเติม ได้แก่ TMP 1992.36.390, TMP 1979.8.622, TMP 1991.144.1 (ชิ้นส่วนขากรรไกรล่างทั้งหมด) และ TMP 1982.39.4 (กระดูกฝ่าเท้าและกระดูกข้อเท้าบางส่วน) เข้ากับสปีชีส์ใหม่โดยพิจารณาจากขนาดที่เล็กกว่าเมื่อเทียบกับ Caenagnathids อื่นๆ ที่ได้รับการบรรยายไว้แล้ว Hagryphus , CaenagnathusและChirostenotesมีขนาดใหญ่กว่าLeptorhynchos ที่เพิ่งตั้งชื่อใหม่มาก ดังนั้นพวกเขาจึงย้ายChirostenotes pergracilisไปอยู่ในสกุลใหม่และเป็นสปีชีส์Leptorhynchos elegans Longrich และเพื่อนร่วมงานได้แยกL. elegans ออก จากชนิดต้นแบบL. gaddisiโดยพิจารณาจากปลายปากที่ยกขึ้นอย่างเห็นได้ชัดและคางที่มีรูปร่างเป็นสี่เหลี่ยมเมื่อมองจากด้านข้าง[ 6 ]
ความเข้าใจในปัจจุบัน

ในปี 2020 Gregory Funston ได้ทำการตรวจสอบ วัสดุ Caenagnathid ทั้งหมด ที่รู้จักจากDinosaur Park Formationเพื่อกำหนดความหลากหลายที่แท้จริงในสภาพแวดล้อมการสะสมตัวนั้น Funston ได้ตรวจสอบและให้คะแนนวัสดุในคอลเลกชันของพิพิธภัณฑ์ธรรมชาติแห่งแคนาดา พิพิธภัณฑ์ Royal Ontarioพิพิธภัณฑ์Royal TyrellมหาวิทยาลัยAlbertaและสถาบันวิทยาศาสตร์แห่งมองโกเลีย ด้วยตนเอง นอกจากนี้เขายังได้เก็บ ตัวอย่าง เนื้อเยื่อจากกระดูกหลายชิ้นเพื่อตรวจสอบ "เส้นการเจริญเติบโตที่หยุดชะงัก" ซึ่งสามารถใช้ประมาณอายุการเจริญเติบโตของตัวอย่างได้[ 1 ]
ในอดีตมีความไม่แน่นอนอย่างมากในเอกสารทางวิทยาศาสตร์เกี่ยวกับระยะการเจริญเติบโตของตัวอย่างแต่ละชิ้น และซากของ Caenagnathid ขนาดเล็กนั้นเป็นของตัวอ่อนหรือตัวเต็มวัยของสายพันธุ์ขนาดเล็กกว่าหรือไม่ การวิเคราะห์ของ Funston ส่งผลให้มีการวินิจฉัยใหม่สำหรับCaenagnathusและChirostenotesซึ่ง Funston โต้แย้งว่าเป็นสกุลที่แตกต่างและถูกต้อง การวิเคราะห์นี้ยังส่งผลให้ตัวอย่างที่จัดอยู่ในLeptorhynchos elegansรวมถึงตัวอย่างที่เพิ่งค้นพบใหม่บางส่วน ถูกจัดอยู่ในสกุลใหม่ที่ตั้งขึ้นคือCitipes [ 1 ]
วัสดุ Caenagnathid ส่วนใหญ่จากDinosaur Park Formationถูกจัดให้อยู่ในหนึ่งในสามสกุลนี้โดยพิจารณาจากขนาด และการวินิจฉัยได้รับการแก้ไขจากลักษณะเฉพาะของตัวอย่างเหล่านั้น ตัวอย่างบางส่วนไม่สามารถจัดอยู่ในสกุลใดสกุลหนึ่งได้อย่างแน่นอนเนื่องจากซากไม่สมบูรณ์[ 1 ]
คำอธิบาย
Citipes เป็นไดโนเสาร์กลุ่ม Caenagnathid ที่มีลำตัวขนาดเล็กในการตรวจสอบไดโนเสาร์ กลุ่ม Caenagnathid ใน Dinosaur Park Gregory Funston สรุปว่ามันมีขนาดตัวเต็มวัยที่เล็กกว่าสกุลCaenagnathusและChirostenotesที่ อยู่ในยุคเดียวกันอย่างมาก การวิเคราะห์ทางเนื้อเยื่อวิทยาของกระดูกฝ่าเท้า สองชิ้น ที่จัดอยู่ในสกุลนี้บ่งชี้ว่าไดโนเสาร์ที่นำกระดูกมานั้นใกล้จะโตเต็มวัยแล้ว แต่ยังไม่โตเต็มที่ Funston มั่นใจในการประเมินของเขาว่าCitipesมีขนาดค่อนข้างเล็กเมื่อยังมีชีวิตอยู่ โดยตัวเต็มวัยมีความยาว 1–2 เมตร (3.3–6.6 ฟุต) [ 1 ] Rubén Molina-Pérez และ Asier Larramendi เสนอการประมาณค่าที่เฉพาะเจาะจงมากขึ้น โดยมีความยาว 2.1 เมตร (6.9 ฟุต) และสูง 82 เซนติเมตร (2.69 ฟุต) ที่สะโพก โดยมีมวลโดยประมาณ 32 กิโลกรัม (71 ปอนด์) [ 12 ]
ในการบรรยายใหม่ที่ครอบคลุมของ caenagnathids จาก Dinosaur Park Formation Gregory Funston ได้ให้การวินิจฉัยที่แก้ไขแล้วสำหรับสกุลทั้งสามที่ระบุชื่อไว้ในการวิเคราะห์ของเขา สำหรับCitipesเขาได้ระบุลักษณะ เฉพาะหลายประการ ได้แก่ การเชื่อมติดกันของปลายสุดของกระดูกข้อเท้าที่ III และ IV การไม่มีส่วนยื่นด้านหลังที่ปลายด้านใกล้ของกระดูกฝ่าเท้าที่ II และ IV และสันนูนที่เด่นชัดบนพื้นผิวด้านหลังของกระดูกฝ่าเท้าที่ III ลักษณะเฉพาะเหล่านี้ไม่เพียงแต่ทำให้Citipes แตกต่างจากสัตว์ร่วมสมัยเท่านั้น แต่ยัง แตกต่างจากสกุลElmisaurusและLeptorhynchos ที่มีความสัมพันธ์ใกล้เคียงกันอีกด้วย [ 1 ]

- โฮโลไทป์
ตัวอย่างต้นแบบของCitipesได้รับการอธิบายในปี พ.ศ. 2476 โดย William Parks ว่าเป็นสายพันธุ์หนึ่งของOrnithomimusประกอบด้วยกระดูกฝ่าเท้าสามชิ้นซึ่งโค้งเข้าด้านในเล็กน้อย โดยกระดูกฝ่าเท้าชิ้นที่ II และ IV มาบรรจบกันที่ปลายด้านใกล้ลำตัว Parks ระบุว่าความยาวของกระดูกฝ่าเท้าอยู่ที่ประมาณ 160 มิลลิเมตร (6.3 นิ้ว) [ 2 ]
- เอกสารอ้างอิง (ปี 1989-2013)
ในปี 1989 ตัวอย่าง ROM 37163 (กระดูกฝ่าเท้าส่วนปลาย II) และ TMP 82.39.4 (กระดูกฝ่าเท้าและกระดูกข้อเท้าบางส่วน) ถูกจัดให้อยู่ในกลุ่มอนุกรมวิธาน เดียวกัน กับ ROM 781 และการวินิจฉัยของตัวอย่างต้นแบบก็ได้รับการแก้ไข การตรวจสอบตัวอย่างต้นแบบอีกครั้งนำไปสู่การค้นพบว่ากระดูกฝ่าเท้า II และ IV เชื่อมติดกัน และการแยกออกจากกันในตัวอย่างต้นแบบนั้นเกิดจากกระบวนการเกิด ซากดึกดำบรรพ์ ความคล้ายคลึงกันทางสัณฐานวิทยา โดยรวมกับ Elmisaurus rarusรวมถึงรูปร่างของหน้าตัดของกระดูกและการมีอยู่ของกระดูกฝ่าเท้าส่วนโค้งซึ่งเป็นสิ่งที่ทำให้Philip Currie จัดให้สปี ชี ส์นี้อยู่ใน สกุล Elmisaurus
ในปี 2013 Nicholas Longrich และเพื่อนร่วมงานได้อ้างอิงวัสดุเพิ่มเติมไปยังLeptorhynchos elegans ที่ได้รับการตั้งชื่อใหม่ ตัวอย่างเหล่านี้ประกอบด้วยกระดูกขากรรไกรล่างหลายชิ้น รวมถึงองค์ประกอบกระดูกฝ่าเท้าและ กระดูกข้อเท้าที่เพิ่งค้นพบ [ 6 ] พวกเขายังตั้งTMP 1992.36.290 (ครึ่งหน้าของกระดูกขากรรไกรล่างสองชิ้น ) เป็นพาราไทป์ของสปีชีส์[ 7 ] ในการจัดกลุ่มใหม่ Longrich และเพื่อนร่วมงานได้แยกL. elegans ออก จากสกุลที่เคยจัดไว้ก่อนหน้านี้ ( ElmisaurusและChirostenotes ) เนื่องจากขนาดที่เล็ก ซึ่งพวกเขาเชื่อว่าไม่สะท้อนถึงอายุตามพัฒนาการเนื่องจากการเชื่อมติดกันขององค์ประกอบกระดูกข้อเท้า นอกจากนี้พวกเขายังตั้งข้อสังเกตว่าขากรรไกรล่างของL. elegans — ซึ่งแสดงให้เห็นได้จากตัวอย่าง TMP 1992.36.390, TMP 1979.8.622 และ TMP 1991.144.1 — แตกต่างอย่างมากจาก caenagnathids อื่นๆ ในด้านความยาวที่ค่อนข้างสั้นและลักษณะที่ยกขึ้นของข้อต่อขากรรไกรล่าง[ 6 ]

- เนื้อหาใหม่ที่ได้รับมอบหมายในปี 2020
ระหว่างการวิเคราะห์อย่างครอบคลุมในปี 2020 Gregory Funston ได้อ้างอิงตัวอย่างสะโพกบางส่วนสองชิ้นไปยังCitipesอย่างไรก็ตาม การกำหนดนี้ค่อนข้างไม่แน่นอนเนื่องจากขาดวัสดุที่ทับซ้อนกับชุดต้นแบบ[ 1 ] Funston ยังโต้แย้งว่าการกำหนดพาราไทป์โดยอาศัยเพียงขนาดและสถานที่ของตัวอย่าง (เมื่อเปรียบเทียบกับ ROM 781) [ 7 ]นั้นไม่มีมูลความจริง และเขาระบุว่ามีหลักฐานไม่เพียงพอที่จะบ่งชี้ว่า TMP 1992.36.290 เป็นของกลุ่มอนุกรมวิธานเดียวกันกับ ROM 781 [ 1 ]
ตัวอย่างชิ้นแรกจากชุดตัวอย่างใหม่ TMP 1981.023.0039 เป็นกระดูกสันหลังส่วนกระเบนเหน็บ ด้านหน้า ซึ่งพบร่วมกับกระดูกเชิงกราน สองชิ้น (TMP 1981.023.0034-5) รูปร่างของกระดูกสันหลังส่วนกลางมีลักษณะเตี้ยและกว้าง ซึ่งเป็นลักษณะที่พบในโอวิแรปเตอร์ซอรัส อื่นๆ อีกหลายชนิด อย่างไรก็ตาม พื้นผิวข้อต่อด้านหน้าของกระดูกสันหลังส่วนกลางยังคงสมบูรณ์ ซึ่งฟันสตันสันนิษฐานว่าหมายความว่ากระดูกสันหลังส่วนกระเบนเหน็บยังไม่เชื่อมติดกันอย่างสมบูรณ์เมื่อสัตว์ตาย นอกจากนี้ยังพบร่องรอยของช่องว่างรอบกระดูกสันหลังส่วนกระเบนเหน็บบนพื้นผิวด้านข้างของกระดูกสันหลังส่วนกลาง แม้ว่ากระดูกเหล่านี้จะไม่ได้ถูกนำไปวิเคราะห์ทางจุลวิทยา แต่ฟันสตันเชื่อว่ากระดูกเหล่านี้เป็นของสัตว์ที่โตเต็มวัย เนื่องจากมีร่องรอยการยึดเกาะของกล้ามเนื้อขนาดใหญ่ที่มองเห็นได้บนกระดูก และความแข็งแรงของสันกระดูกเชิงกราน ขนาดที่เล็กในสัตว์ที่โตเต็มวัยทำให้ไม่สามารถจัด TMP 1981.023.0039 ให้กับCaenagnathusหรือChirostenotes ได้ดังนั้นจึงน่าจะเป็นของCitipes [ 1 ]
ตัวอย่างที่เหลือ ได้แก่ UALVP 59606, UALVP 55585 และ UALVP 59606 ประกอบด้วยกระดูกทาร์โซเมตาตาร์ซัล บางส่วน ที่มีความสมบูรณ์และสถานะการสร้างกระดูกที่แตกต่างกัน ตัวอย่างสุดท้ายนี้ถูกตัดเพื่อ การวิเคราะห์ ทางเนื้อเยื่อวิทยา พบ ความคล้ายคลึงกับองค์ประกอบเมตาตาร์ซัลที่รู้จักของElmisaurusในกระดูกเหล่านี้ ซึ่งสะท้อนให้เห็นในการวิเคราะห์ทางวิวัฒนาการที่ดำเนินการในเอกสารเดียวกัน[ 1 ]
Funston ได้บรรยายตัวอย่าง Caenagnathid อื่นๆ อีกหลายตัวอย่างจากคอลเลกชันของพิพิธภัณฑ์ Royal Tyrell แต่ไม่ได้อ้างอิงถึงสกุลใดสกุลหนึ่งโดยเฉพาะภายใน Caenagnathidae ซากเหล่านี้ซึ่งรวมถึงกระดูกขากรรไกรมีขนาดเล็ก แต่มีลักษณะทางสัณฐานวิทยาคล้ายกับ Chirostenotes Funston โต้แย้งว่าไม่สามารถอ้างอิงถึงสกุลใดสกุลหนึ่งได้หากไม่ได้ทำการตัดแบ่งและกำหนดอายุตามพัฒนาการ[ 1 ]
การจำแนกประเภท
วิวัฒนาการ

การวิเคราะห์ เชิงวิวัฒนาการอย่างเป็นระบบครั้งแรกที่รวม ROM 781 เข้าไปด้วยนั้น ดำเนินการโดย Nicholas Longrich, Ken Barnes, Scott Clark และ Larry Millar ในบทความของพวกเขาที่อธิบายสกุลLeptorhynchosในปี 2013 ในการวิเคราะห์ของพวกเขา พวกเขากำหนดให้ ROM 781 และตัวอย่างอื่นๆ อีกหลายตัวอย่างเป็นสปีชีส์ที่สองในสกุลLeptorhynchos — L. elegansการวิเคราะห์ของพวกเขาใช้ชุดข้อมูลจากสิ่งพิมพ์ก่อนหน้าของ Longrich ในปี 2010 โดยมีการเพิ่มแท็กซาใหม่หลายรายการ รวมเป็น 28 แท็กซาที่เข้ารหัสสำหรับ 205 ลักษณะ ชุดข้อมูลของพวกเขายังรวมถึงข้อมูลล่าสุดเกี่ยวกับNemegtomaiaและNomingiaซึ่งได้รับการตีพิมพ์ตั้งแต่การวิเคราะห์ในปี 2010 [ 6 ]
การวิเคราะห์ของพวกเขาสนับสนุนความเป็นกลุ่มเดียวของวงศ์ Caenagnathidae โดยอาศัยลักษณะเฉพาะดังต่อไปนี้: กระดูกขากรรไกรล่างเชื่อมติดกัน กระบวนการโค้งงอลงด้านล่างของกระดูกขากรรไกรล่างการไม่มีการสัมผัสกันระหว่างกระดูกขากรรไกรล่างและ ช่องเปิดของขากรรไกรล่าง และกระดูกซูแองกูลาร์ที่ ตื้น การวิเคราะห์นี้สามารถจำแนกกลุ่มอนุกรมวิธานหลายกลุ่มที่ฐานของวงศ์ Caenagnathidae ได้ รวมถึงGigantoraptorและMicrovenatorแต่การจำแนกในส่วนที่วิวัฒนาการสูงกว่าของแผนภูมิวิวัฒนาการนั้นทำได้ไม่ดีนัก โดยเฉพาะอย่างยิ่ง แผนภูมิวิวัฒนาการแบบฉันทามติที่เข้มงวดของพวกเขาไม่ได้สนับสนุนความเป็นกลุ่มเดียวที่ชัดเจนของสกุลLeptorhynchos ; สองชนิดที่พวกเขากำหนดให้เป็นสกุลนี้อยู่ในกลุ่มที่มีความสัมพันธ์กันหลายทางกับHagryphusและกลุ่มที่ประกอบด้วยCaenagnathus , Chirostenotesและตัวอย่าง BMNH 2033 (ซึ่งถูกระบุว่าเป็นชนิดที่ไม่สามารถระบุได้ของCaenagnathus ) ผู้เขียนตั้งข้อสังเกตว่าพวกเขาอ้างอิง ROM 781 ไปยังLeptorhynchosเนื่องจากขนาดของทั้งสองชนิดมีความคล้ายคลึงกัน แต่ตั้งข้อสังเกตว่าพวกมันอาจอยู่ในสกุลที่แยกจากกัน ขึ้นอยู่กับการวิจัยเพิ่มเติม[ 6 ]
Longrich และเพื่อนร่วมงานได้สรุปอย่างกว้างๆ ว่า สกุลของ caenagnathid ส่วนใหญ่ สามารถวินิจฉัยได้จากลักษณะของขากรรไกรล่างและมือซึ่งช่วยให้นักวิจัยสามารถวินิจฉัยสกุลและชนิดได้จากซากที่ไม่สมบูรณ์มาก แต่ก็หมายความว่าซากที่สมบูรณ์กว่านั้นจะไม่สามารถวินิจฉัยได้หากไม่มีขากรรไกรล่างหรือมือ อย่างไรก็ตาม ผู้เขียนตั้งข้อสังเกตว่ารูปร่างของจะงอยปากในนก สมัยใหม่ เป็นลักษณะการวินิจฉัยที่สำคัญ เช่นเดียวกับในไดโนเสาร์บางชนิด เช่นTriceratopsและEdmontosaurusดังนั้นพวกเขาจึงโต้แย้งว่าไม่ใช่เรื่องที่ไม่สมเหตุสมผลที่จะสันนิษฐานว่าสิ่งนี้อาจเพียงพอในการตั้งชื่อสกุลหรือชนิดใหม่ ยิ่งไปกว่านั้น ระดับการเชื่อมต่อของกระดูกขากรรไกร รวมถึงพื้นผิวของกระดูก สามารถบ่งชี้อายุการเจริญเติบโตในoviraptorosaurs หลายชนิด ซึ่งสามารถใช้เพื่อพิจารณาว่าตัวอย่างแต่ละตัวเป็น taxon ใหม่หรือเป็นเพียงระยะชีวิตที่แตกต่างกันของ taxon ที่มีอยู่แล้ว[ 6 ]
สำหรับตัวอย่างที่ไม่มีลักษณะการวินิจฉัยที่เกี่ยวข้องหรือข้อบ่งชี้ใด ๆ เกี่ยวกับอายุการเจริญเติบโต พวกมันจะถูกจัดให้อยู่ในกลุ่มอนุกรมวิธานที่มีอยู่ตามขนาดและสถานที่ Longrich และเพื่อนร่วมงานตั้งข้อสังเกตว่าวิธีการนี้มีข้อบกพร่อง แต่พวกเขาโต้แย้งว่าความอุดมสมบูรณ์ทางนิเวศวิทยาของสัตว์โตเต็มวัยในระบบนิเวศใด ๆ หมายความว่าสัตว์ที่กลายเป็นฟอสซิลส่วนใหญ่จะเป็นสัตว์โตเต็มวัยอยู่แล้ว ซึ่งพวกเขาแนะนำว่าเพียงพอสำหรับวัตถุประสงค์ของการวิเคราะห์ของพวกเขา แผนภูมิต้นไม้ที่ประหยัดที่สุด 116 ต้นในการวิเคราะห์ของพวกเขามีแสดงไว้ด้านล่าง[ 6 ]
| โอวิแรปเตอร์ซอเรีย |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

ในการทบทวนcaenagnathidsจากDinosaur Park Formation ของเขา Funston ได้ทำการ วิเคราะห์ เชิงวิวัฒนาการของoviraptorosauria [ 1 ] การ วิเคราะห์นี้ใช้ชุดข้อมูลเดียวกันกับที่เขาพัฒนาขึ้นเพื่ออธิบายสกุลใหม่Oksoko [ 13 ]และรวมถึง taxa oviraptorosaur ที่ได้รับการอธิบายทั้งหมดที่รู้จักในเวลานั้น รวมถึง taxa ที่ยังไม่ได้ตั้งชื่ออีกจำนวนหนึ่ง[ 1 ]
การวิเคราะห์นี้ได้ผลลัพธ์ที่คล้ายคลึงกับการวิเคราะห์ก่อนหน้านี้ส่วนใหญ่ กล่าวคือelmisaurinaeไม่ใช่กลุ่มโมโนฟิเลติก ตามธรรมชาติ ฟันสตันพบว่าสกุลที่จัดอยู่ในกลุ่ม "elmisaurinae" ดั้งเดิมหลายสกุล เช่นChirostenotesและApatoraptorมีการกระจายตัวอย่างกว้างขวางในแผนภูมิวิวัฒนาการของเขา มากกว่าที่จะรวมกลุ่มกันเป็นกลุ่มเดียวของการวิวัฒนาการของ caenagnathids ใน ภูมิภาคลารามิเดีย เขายังพบว่าCaenagnathasiaซึ่งเป็นหนึ่งใน caenagnathids ที่เก่าแก่ที่สุดและชัดเจนที่สุด อยู่ในตำแหน่งที่ค่อนข้างแตกต่างออกไป โดยเป็นสกุลพี่น้องของEpichirostenotesในภูมิภาคลารามิเดีย สกุล NomingiaและAnomalipesที่เป็นที่ถกเถียงกันในเอเชียก็ถูกจัดอยู่ในกลุ่ม caenagnathids ในการวิเคราะห์ของเขาเช่นกัน ซึ่งบ่งชี้ว่ากลุ่มนี้ยังคงมีอยู่ในเอเชียแม้หลังจากการวิวัฒนาการของoviraptoridaeอย่างไรก็ตาม ฟันสตันระบุอย่างระมัดระวังว่าโครงสร้างของแผนภูมิวิวัฒนาการของเขาไม่ได้รับการสนับสนุนอย่างแข็งแกร่ง และมีแนวโน้มที่จะได้รับการแก้ไขโดยการวิเคราะห์ในอนาคต[ 1 ]

ความหมายที่คล้ายคลึงกัน

ในการศึกษาสัณฐานวิทยา ของ กระดูกเชิงกราน ของวัสดุ Caenagnathid ทั้งหมดจากDinosaur Park Formationในปี 2019 Matthew Rhodes และเพื่อนร่วมงานได้จัดประเภทตัวอย่างแต่ละชิ้นออกเป็น 3 ประเภท สัณฐานวิทยาที่พวกเขาเชื่อว่าเป็นของกลุ่มอนุกรมวิธานใหม่[ 14 ] ซึ่งต่อมาได้รับการตั้งชื่อว่า Citipes [ 1 ] ถูกอธิบายว่าเป็น" สัณฐานวิทยาขนาดเล็กและแข็งแรง" และแตกต่างจากกระดูกเชิงกรานของ Caenagnathid อื่นๆ ทั้งหมดในชั้นหิน ความแตกต่างที่เห็นได้ชัดที่สุดคือกระดูกเชิงกรานของสัณฐานวิทยาขนาดเล็กและแข็งแรงนั้นโค้งงอเป็นรูปตัว S เล็กน้อยเมื่อมองจากด้านบนหรือด้านล่าง ความแตกต่างอื่นๆ ก็มีอยู่แต่ไม่ชัดเจนนัก เช่นก้านกระดูกหัวหน่าว ที่เล็กมาก และสันหลังที่ขยายใหญ่ขึ้นที่ส่วนหลังของกระดูกเชิงกราน[ 14 ]
โรดส์และเพื่อนร่วมงานแนะนำว่าความแตกต่างทางสัณฐานวิทยาในกระดูกสะโพกของซีนาแนทิดนั้นไม่น่าจะเป็นผลมาจากการเจริญเติบโต เนื่องจากระดับการเชื่อมต่อระหว่างกระดูกเชิงกรานและกระดูกศักระมีความคล้ายคลึงกันในตัวอย่างที่มีขนาดแตกต่างกันมาก[ 14 ] ฟันสตันได้ขยายความการค้นพบนี้ในปี 2020 ในคำอธิบายของCitipesอายุการเจริญเติบโตที่คล้ายคลึงกันของซากกระดูกส่วนลำตัวที่มีขนาดแตกต่างกันบ่งชี้ถึงความเป็นไปได้ที่จะมีซีนาแนทิดสามกลุ่มใน Dinosaur Park สองในกลุ่มอนุกรมวิธานเหล่านี้ได้รับการตั้งชื่อไปแล้ว และกลุ่มตัวอย่างที่เหลือ — ซึ่งประกอบด้วยรูปร่างเล็กและแข็งแรง — มีลักษณะทางสัณฐานวิทยาที่แตกต่างจากLeptorhynchos สายพันธุ์เท็กซัส ซึ่งได้รับการตั้งชื่อไว้ในปี 2013 [ 1 ] การวิเคราะห์ทางวิวัฒนาการไม่เคยพบว่า Leptorhynchos สอง "สายพันธุ์" เป็นกลุ่มพี่น้อง[ 6 ]แต่ยังไม่มีการทบทวนการวิเคราะห์ทางวิวัฒนาการของ Funston อย่างครอบคลุมจนถึงปัจจุบัน และยังไม่มีการวิเคราะห์ใหม่ใด ๆ ดำเนินการในขณะนี้
บรรพชีววิทยา
การเจริญเติบโตและเนื้อเยื่อวิทยา

การศึกษาพัฒนาการของซีนาแนทิดเป็นเรื่องยากเนื่องจากซากที่รู้จักไม่สมบูรณ์ ยิ่งไปกว่านั้นยังซับซ้อนขึ้นจากการขาดแคลนโครงกระดูกที่ทับซ้อนกันจากกลุ่มอนุกรมวิธานต่างๆ ที่ระบุชื่อไว้ในDinosaur Park Formation Gregory Funston และเพื่อนร่วมงานได้ทำการ ศึกษา ทางเนื้อเยื่อวิทยาของขากรรไกรล่างของซีนาแนทิด (ซากที่พบมากที่สุดใน Dinosaur Park) และสังเกตว่าข้อต่อขากรรไกรล่างจะกลายเป็นกระดูกแข็งอย่างสมบูรณ์ค่อนข้างเร็วในการเจริญเติบโตของซีนาแนทิด ดังนั้นจึงเป็นตัวบ่งชี้อายุที่แน่นอนของตัวอย่างที่ไม่ดี อุปสรรคอีกประการหนึ่งในการระบุอายุที่แน่นอนคือความไม่สม่ำเสมอในการสะสมของเนื้อเยื่อกระดูกเมื่อสัตว์เจริญเติบโต Funston และเพื่อนร่วมงานสังเกตว่าเส้นการเจริญเติบโตที่หยุดชะงัก (หรือ LAGs) ซึ่งมักจะสอดคล้องกับการเจริญเติบโตที่ช้าลงในช่วงฤดูหนาว อาจไม่ได้เกิดขึ้นในซีนาแนทิดที่มีอายุน้อยกว่า 1-2 ปี ซึ่งอาจนำไปสู่การประเมินอายุของตัวอย่างบางตัวอย่างต่ำกว่าความเป็นจริง[ 15 ]
พยาธิวิทยา
หนึ่งในตัวอย่างที่อ้างถึงCitipes (ในขณะนั้นคือLeptorhynchos elegans ) คือ TMP 1992.036.0476 ซึ่งเป็นกระดูกเชิงกรานที่ค่อนข้างสมบูรณ์แต่แตกหักในช่วงใดช่วงหนึ่งของชีวิตของสัตว์ รอยแตกขยายไปตามพื้นผิวระหว่าง ก้านกระดูก หัวหน่าวและ ก้าน กระดูกเชิงกรานและส่งผลให้กระดูกเชิงกรานบวมอย่างเห็นได้ชัด ซึ่งสามารถมองเห็นได้อย่างชัดเจนจากด้านบนและบดบังร่องเบรวิสเล็กน้อยการแตกหัก ของฟอสซิลยังเผยให้เห็นกระดูกพรุนในตัวอย่าง ซึ่งบ่งชี้ว่าสัตว์มีชีวิตรอดอย่างน้อยในช่วงเวลาสั้น ๆ หลังจากได้รับบาดเจ็บที่ทำให้เกิดรอยแตกนี้[ 16 ]
นิเวศวิทยาบรรพกาล
อาหารและการแบ่งกลุ่มเฉพาะถิ่น
ชั้นหินDinosaur Park Formationสามารถแบ่งออกเป็นกลุ่มสัตว์หลายกลุ่มที่แยกจากกัน ซึ่งไม่ได้เกิดขึ้นพร้อมกันทั้งหมด[ 17 ] อย่างไรก็ตาม ปัจจุบันยังไม่ชัดเจนว่าสกุล Caenagnathid ที่รู้จักนั้นจำกัดอยู่เฉพาะในเขตสัตว์หนึ่งเขตหรือมากกว่านั้นหรือไม่ ซากของ Caenagnathid ไม่ได้รับการค้นพบบ่อยเท่ากับซากของ Coelurosaur อื่นๆ แต่ส่วนหนึ่งเป็นเพราะขนาดที่ค่อนข้างเล็กและไม่มีฟัน ซึ่งอาจทำให้เกิดอคติในการสุ่มตัวอย่าง[ 1 ]
แม้ว่าซากดึกดำบรรพ์ของ Caenagnathid จะค่อนข้างหายาก แต่ก็กระจายอยู่ทั่ว Dinosaur Provincial Park โดยไม่มีรูปแบบที่ชัดเจนว่าสกุลใดพบได้ในสถานที่ใดโดยเฉพาะ ซึ่งบ่งชี้ว่าพวกมันมีอยู่ทั่วเขตสัตว์ของ Dinosaur Park Formation ซึ่งหมายความว่ามีการแบ่งแยกนิเวศวิทยาบางอย่างระหว่างสามกลุ่มอนุกรมวิธาน ความแตกต่างทางสัณฐานวิทยาที่เห็นได้ชัดที่สุดระหว่างCaenagnathus , ChirostenotesและCitipesคือความแตกต่างของขนาดตัวเต็มวัย นี่เป็นวิธีการหลักในการแยกแยะซากดึกดำบรรพ์ของทั้งสามสกุล แต่ยังมีความแตกต่างทางสัณฐานวิทยาที่ละเอียดอ่อนกว่าอีกหลายประการ เชื่อกันว่า Citipesมีการปรับตัวให้เหมาะกับการวิ่ง มากกว่า เนื่องจากการเชื่อมติดกันของกระดูกฝ่าเท้าและบริเวณยึดเกาะที่ขยายใหญ่ขึ้นสำหรับกล้ามเนื้อขาตามสันกระดูกเชิงกราน ยังไม่แน่ชัดว่าผลกระทบทางนิเวศวิทยาของการปรับตัวเหล่านี้จะเป็นอย่างไร แต่ดูเหมือนว่าจะเป็นเอกลักษณ์ในแง่นั้นในบรรดา Caenagnathid ของ Dinosaur Park Formation [ 1 ]
Caenagnathusมีขนาดใหญ่กว่า Citipesหรือ Chirostenotes อย่างมาก [ 1 ]และเชื่อกันว่าเป็นสัตว์กินพืชเป็นหลัก เนื่องจาก Anzu ซึ่งเป็นแท็กซอนที่เข้าใจได้ดีกว่าและมีความสัมพันธ์ใกล้ชิดกัน นั้นเชื่อกันว่าเป็นสัตว์กินพืช [ 18 ] ในทางกลับกัน Chirostenotesมีปลายขากรรไกรล่างที่งอนขึ้นอย่างเห็นได้ชัด ซึ่งอาจเป็นการปรับตัวเพื่อการดำรงชีวิตแบบกินเนื้อมากกว่า [ 19 ]แม้ว่าสิ่งนี้ยังคงเป็นเพียงการคาดเดาและไม่สามารถทดสอบได้เนื่องจากซากดึกดำบรรพ์เหล่านี้ไม่สมบูรณ์ [ 1 ]
การล่าเหยื่อ

ในปี 2023 François Therrien และเพื่อนร่วมงานได้อธิบาย TMP 2009.12.14 ซึ่งเป็นตัวอย่างลูกไดโนเสาร์Gorgosaurus ที่ได้รับการอนุรักษ์ไว้อย่างดีเยี่ยม โดยมีซากของไดโนเสาร์ Citipesอย่างน้อยสองตัวอยู่ภายในขาของไดโนเสาร์Citipesเหล่านั้นงอและบิดเป็นรูปทรงที่กะทัดรัดมาก ซึ่งบ่งชี้ว่าพวกมันอยู่ในกระเพาะของGorgosaurusมากกว่าที่จะถูกเก็บรักษาไว้ข้างๆ หรือถูกชะล้างมาด้วยกันหลังการตาย สิ่งนี้แสดงให้เห็นว่าGorgosaurusกินCitipesเป็นอาหารเป็นบางครั้ง[ 20 ]
กระดูกต้นขาของGorgosaurusถูกตัดแบ่งและนำไปวิเคราะห์ทางเนื้อเยื่อวิทยา ซึ่งเผยให้เห็นว่ามันมีอายุระหว่าง 5-7 ปีเมื่อมันตาย และมีมวลเพียงประมาณหนึ่งในแปดของGorgosaurus ตัวเต็มวัย เนื้อเยื่อวิทยาของ กระดูก Citipes แสดงให้เห็นว่าไม่มีร่องรอยการเจริญเติบโตเลย ทำให้ Therrien และเพื่อนร่วมงานเสนอแนะว่าบุคคลเหล่านี้มีอายุ อยู่ในปีแรกของชีวิตเมื่อถูกGorgosaurus กิน [ 20 ]
ที่น่าสังเกตคือ ดูเหมือนว่าซากของ Citipesจะไม่ได้ถูกกินไปทั้งหมด มีเพียงขาหลังและส่วนของกระดูกสันหลังส่วนหาง เท่านั้น ที่ยังคงอยู่ในกระเพาะอาหารของGorgosaurusซึ่งไม่น่าจะเกิดจากการย่อยอาหารเนื่องจากทราบกันว่าซากทั้งสองถูกกินในเวลาที่ต่างกันเนื่องจากตำแหน่งที่สัมพันธ์กันภายในทางเดินอาหารที่คาดการณ์ไว้ของสัตว์นักล่า และGorgosaurusต้องตายก่อนที่จะย่อยCitipes อย่างสมบูรณ์ มิฉะนั้นซากจะไม่ได้รับการเก็บรักษาไว้ เมื่อพิจารณาร่วมกันแล้ว สิ่งนี้ชี้ให้เห็นว่าสัตว์นักล่าอาจชอบกินขาหลังของCitipesซึ่งสันนิษฐานว่ามีกล้ามเนื้อมากกว่าและมีคุณค่าทางโภชนาการมากกว่า[ 20 ]
ในการบรรยาย TMP 2009.12.14 ของพวกเขา Therrien และเพื่อนร่วมงานได้ทำการวิเคราะห์การถดถอยของความสัมพันธ์ระหว่างมวลของสัตว์เลื้อยคลานและสัตว์เลี้ยงลูกด้วยนมที่เป็นผู้ล่าในปัจจุบันกับมวลของเหยื่อที่อาจอยู่ร่วมกัน พวกเขาใช้สิ่งนี้ร่วมกับมวลโดยประมาณของGorgosaurus วัยเยาว์และวัยผู้ใหญ่ เพื่อทำนายช่วงขนาดของเหยื่อที่เป็นไปได้สำหรับ Gorgosaurus วัยเยาว์และวัยผู้ใหญ่ ผลการค้นพบของพวกเขาบ่งชี้ว่าGorgosaurusทั้งสอง ช่วง การเจริญเติบโต หลัก คือ วัยเยาว์และวัยผู้ใหญ่ อยู่ในช่วงขนาดที่Citipes ทั้งวัยเยาว์และวัยผู้ใหญ่ สามารถถือเป็นเหยื่อได้[ 20 ]
สภาพแวดล้อมโบราณ

ไซติเปสถูกค้นพบที่บริเวณลิตเติลแซนด์ฮิลล์ครีก ซึ่งตั้งอยู่ในอุทยานไดโนเสาร์ประจำจังหวัดในอัลเบอร์ตาประเทศแคนาดาอายุของบริเวณนี้เมื่อเทียบกับส่วนอื่นๆ ของชั้นหินไดโนเสาร์พาร์คยังไม่เป็นที่ทราบแน่ชัด[ 1 ] อย่างไรก็ตาม ชั้นหินไดโนเสาร์พาร์คโดยรวมเป็นที่ทราบกันดีว่าถูกสะสมตัวในช่วงระหว่าง 77.03 ถึง 75.46 ล้านปีก่อน[ 21 ]
ชั้น หิน Dinosaur Park Formationประกอบด้วยตะกอนที่เกิดจากการกัดเซาะของภูเขาทางทิศตะวันตก โดยถูกทับถมบนที่ราบลุ่มน้ำชายฝั่งโดยระบบแม่น้ำที่ไหลไปทางทิศตะวันออกและทิศตะวันออกเฉียงใต้สู่ทะเล Bearpaw ซึ่งเป็นทะเลภายในขนาดใหญ่ที่เป็นส่วนหนึ่งของWestern Interior Seawayทะเลดังกล่าวค่อยๆ ท่วมที่ราบชายฝั่งที่อยู่ติดกัน ทำให้เกิดการสะสมของหินดินดาน ทะเล ของชั้น หิน Bearpaw Formationทับอยู่บนชั้นหิน Dinosaur Park Formation [ 22 ]
ชั้นหิน Dinosaur Park Formation มีความหนาประมาณ 75 เมตร (250 ฟุต) ส่วนล่างของชั้นหินนี้ถูกสะสมตัวในสภาพแวดล้อมของร่องน้ำและ ประกอบด้วย หินทรายเนื้อละเอียดถึงปานกลางที่มี การวางตัวแบบเฉียงเป็นหลัก ส่วนบนซึ่งถูกสะสมตัวใน สภาพแวดล้อม ของตลิ่งและที่ราบน้ำท่วมถึง ประกอบด้วยหินโคลน เนื้อแน่นถึงเป็นชั้นที่มีอินทรียวัตถุสูง มีร่องรอยรากจำนวนมาก และมีชั้นเบนโทไนต์ บางๆ เขตถ่านหิน Lethbridge ซึ่งประกอบด้วยชั้นถ่านหินคุณภาพต่ำ หลายชั้น สลับกับหินโคลนและหินทรายแป้งเป็นจุดสูงสุดของชั้นหินนี้[ 22 ]
ลักษณะทางธรณีวิทยาเหล่านี้ถูกตีความว่าเป็นภูมิประเทศที่มีความสูงต่ำของแม่น้ำและที่ราบน้ำท่วมถึง ซึ่งกลายเป็นพื้นที่ชื้นแฉะ มากขึ้น และได้รับอิทธิพลจาก สภาพแวดล้อม ทางทะเลเมื่อเวลาผ่านไป เนื่องจากทะเลภายในตะวันตกรุกคืบไปทางทิศตะวันตก[ 23 ]สภาพอากาศอบอุ่นกว่าอัลเบอร์ตาในปัจจุบัน ไม่มีน้ำค้างแข็งแต่มีฤดูที่เปียกชื้นและแห้งแล้ง สลับกันไป ต้นสน เป็น พืชเด่นในชั้นเรือนยอด โดยมี เฟิร์นเฟิร์นต้นไม้และพืชดอกเป็นพืชชั้นล่าง[ 24 ]
สัตว์ร่วมสมัย

แหล่งฟอสซิล Dinosaur Park Formation เป็นแหล่งรวม ไดโนเสาร์ที่ไม่ใช่นกที่มีความหลากหลายมากที่สุดแห่งหนึ่งเท่าที่เคยพบในบันทึกฟอสซิล อย่างน้อยหกชนิดของฮาโดรซอริเดส รวมถึง ParasaurolophusและCorythosaurusที่เป็นที่รู้จักกันดีพบได้ในชั้นต่างๆ นอกจากนี้ยังมีเซราทอปซิดอีกแปดชนิดขึ้นไปใน สกุลที่เป็นที่รู้จักกันดี เช่นStyracosaurus , ChasmosaurusและCentrosaurus แอนคิโล ซอรัส ก็มีอยู่มากมายเช่น กันโดยมีทั้งโนโดซอริเดสเช่นEdmontoniaและแอนคิโลซอริเดสเช่นEuoplocephalus [ 22 ]
ไดโนเสาร์กินเนื้อขนาดใหญ่หลากหลายชนิดเหล่านี้ได้แก่ไทแรนโนซอริเด อย่างกอร์โกซอรัส และดาสเปลโตซอรัสซึ่งอาศัยอยู่ร่วมกันในชั้นหินไดโนเสาร์พาร์ค ฟอร์เมชัน รวม ถึงชั้นหิน จูดิธริเวอร์และทูเมดิซีนฟอร์เมชัน[ 22 ]กอร์โกซอรัสยังเป็นที่รู้จักกันดีว่าล่าซิติเปสโดยตรง นอกเหนือจากไดโนเสาร์ขนาดใหญ่[ 20 ] ไทแรนโนซอริเดขนาดใหญ่เหล่านี้มาพร้อมกับเทโรพอด ขนาดเล็กหลายชนิด ได้แก่ออร์นิโทมิโม ซอรัส โทรโอโดนทิดและโดรเมโอซอรัสนอกเหนือจากซีนาแนทิด ที่กล่าวถึงข้างต้น เทโร พอดขนาดเล็กเหล่านี้เป็นที่รู้จักจากซากที่ไม่สมบูรณ์นัก โดยบางชนิด เช่นพารอนิโคดอนเป็นที่รู้จักจากฟันเท่านั้น เทโรพอดขนาดเล็กที่เป็นที่รู้จักกันดี ได้แก่โดรเมโอซอรัส ลาเทนิเวนาทริกซ์และซอโรร์นิโทเลสเตส[ 22 ]
ทางน้ำของ Dinosaur Park Formation ยังเป็นแหล่งอาศัยของเพลซิโอซอร์ หลายชนิด แม้ว่าจะมีเพียงชนิดเดียวคือFluvionectesที่ได้รับการตั้งชื่อ[ 25 ] ในบรรดาสัตว์เลื้อยคลานน้ำอื่นๆ ที่พบ ได้แก่Choristodere Champsosaurusและจระเข้AlbertochampsaและLeidyosuchus [ 22 ] Cryodrakon ซึ่ง เป็นAzhdarchid ที่เพิ่งได้รับการอธิบายเมื่อไม่นานมานี้ก็ได้รับการอนุรักษ์ไว้ร่วมกับซากของเทโรซอร์ ที่ไม่สามารถระบุชนิดได้ [ 26 ]
ชั้นหิน Dinosaur Park Formation ยังเก็บรักษาซากสัตว์ขนาดเล็กหลากหลายชนิดซึ่งเคยมีอยู่มากมายในทางน้ำของสภาพแวดล้อมการสะสมตัว ได้แก่ เต่าหลายชนิด เช่นBasilemys ขนาดใหญ่ ในวงศ์ Nanhsiungchelyidae และสกุลAdocus ที่แพร่หลาย นอกจากนี้ ยังมีกิ้งก่า จำนวนมาก ใน Dinosaur Park โดยมีสายพันธุ์จากวงศ์Helodermatidae , Varanidae , TeiidaeและXenosauridaeสัตว์ เลี้ยงลูกด้วยนมก็มีอยู่มากมายในพื้นที่ โดยมีตัวแทนจากกลุ่มสัตว์เลี้ยงลูกด้วยนม หลักทั้งสามกลุ่ม ในยุคครีเทเชียสตอนปลาย ได้แก่ eutherians , metatheriansและmultituberculatesจากหลายสกุลในแต่ละกลุ่ม ระบบน้ำจืดของภูมิภาคนี้ยังรองรับปลาหลากหลายชนิด รวมถึงฉลามพรมปลากระเบนปลากะพงปลาสเตอร์เจียนและปลาพาย[ 22 ]
ดูเพิ่มเติม
- ปี 2020 ในสาขาบรรพชีวินวิทยาของอาร์โคซอร์
- ปี 2023 ในสาขาบรรพชีวินวิทยาของอาร์โคซอร์
- ยุคครีเทเชียสของสัตว์มีกระดูกสันหลังบนบกในทวีปอเมริกาเหนือ ( ยุคจูดิเธียน )
- ชั้นหิน Judith River FormationและTwo Medicine Formation - ชั้นหินทางธรณีวิทยาที่เกิดขึ้นในช่วงเวลาใกล้เคียงกันในรัฐอัลเบอร์ตาและรัฐมอนแทนา
- รายชื่อไดโนเสาร์ในทวีปอเมริกาเหนือ
- รายชื่อชั้นหินที่มีซากดึกดำบรรพ์ของไดโนเสาร์
- ลำดับเหตุการณ์ของการวิจัยเกี่ยวกับโอวิแรปเตอร์ซอรัส
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เมืองต่างๆ
Citipes ( / ˈ s ɪ t i p ɛ z / ) เป็นสกุลของเทโรพอดCaenagnathid ที่ สูญพันธุ์ไปแล้ว จากยุคครีเทเชียสตอนปลาย ใน Dinosaur Park...
การค้นพบเบื้องต้น
วัสดุที่ต่อมาได้รับการตั้งชื่อว่า Citipes ถูกค้นพบในปี 1926 ระหว่าง การสำรวจของ มหาวิทยาลัยโทรอนโต ในพื้นที่ Sand Creek ของ แม่น้ำ Red Deer สถานที่แห่งนี้เป็นส่วนหนึ่งของ กลุ่ม Belly River ซึ่งปัจจุบันได้รับการยอมรับว่าเป็น Dinosaur Park Formation...
ประวัติการจำแนกประเภท
ROM 781 ถูกจัดให้อยู่ในสกุล Macrophalangia ที่น่าสงสัย โดย Dale Russell ในปี 1972 [ 3 ] อย่างไรก็ตาม สกุลนี้ถูกจัดให้เป็น ชื่อพ้องรอง ของ Chirostenotes ในอีกไม่กี่ปีต่อมา [ 11 ]
ความเข้าใจในปัจจุบัน
ในปี 2020 Gregory Funston ได้ทำการตรวจสอบ วัสดุ Caenagnathid ทั้งหมด ที่รู้จักจาก Dinosaur Park Formation เพื่อกำหนดความหลากหลายที่แท้จริงในสภาพแวดล้อมการสะสมตัวนั้น Funston ได้ตรวจสอบและให้คะแนนวัสดุในคอลเลกชันของพิพิธภัณฑ์ ธรรมชาติแห่งแคนาดา พิพิธภัณฑ์...