อ่าน 20 นาที
เทโรโปดา
Theropoda ( / θ ɪ ˈ r ɒ p ə d ə / ; จากภาษากรีกโบราณθηρίο- ποδός ) เป็นหนึ่งในสามกลุ่ม หลัก ของไดโนเสาร์ร่วมกับOrnithischiaและSauropodomorpha Theropods...
เทโรโปดา
| เทโรโปดา ช่วงเวลา: ปลายยุคไทรแอสสิก – ปัจจุบัน (รวมถึงนก) | |||||
|---|---|---|---|---|---|
แกลเลอรี่ภาพไดโนเสาร์เทอโรพอด (เรียงตามเข็มนาฬิกาจากซ้ายบน) คาร์โนทอรัส,โค เอโลฟิซิส , อิริเทเตอร์ , อาร์คีออปเทอริกซ์ , สตรูธิโอมิมัสและไทแรนโนซอรัส | |||||
| การจำแนกทางวิทยาศาสตร์ | |||||
| อาณาจักร: | แอนิมอลเลีย | ||||
| ไฟลัม: | คอร์ดาต้า | ||||
| ระดับ: | สัตว์เลื้อยคลาน | ||||
| กลุ่มสายพันธุ์ : | ไดโนเสาร์ | ||||
| กลุ่มสายพันธุ์ : | ซอริสเชีย | ||||
| กลุ่มสายพันธุ์ : | บึงเทโรโป ดา , 1881 | ||||
| กลุ่มย่อย[ 1 ] | |||||
| |||||







Theropoda ( / θ ɪ ˈ r ɒ p ə d ə / ; [ 2 ] จากภาษากรีกโบราณθηρίο- ποδός [ θηρίον , ( therion ) "สัตว์ป่า"; πούς , ποδός ( pous, podos ) "เท้า"]) เป็นหนึ่งในสามกลุ่ม หลัก ของไดโนเสาร์ร่วมกับOrnithischiaและSauropodomorpha Theropods ทั้งที่ยังมีชีวิตอยู่และที่สูญพันธุ์ไปแล้ว มีลักษณะเด่นคือกระดูกกลวงและมีนิ้วเท้าสามนิ้วและกรงเล็บในแต่ละขา โดยทั่วไปแล้วพวกมันถูกจัดอยู่ในกลุ่ม ไดโนเสาร์ ซอริ สเชียน ซึ่ง ทำให้พวกมันอยู่ใกล้กับซอโรโพโดมอร์ฟมากกว่าออร์นิธิสเชียน เดิมทีพวกมันเป็นสัตว์กินเนื้อแต่กลุ่มเทโรพอดหลายกลุ่มได้วิวัฒนาการมาเป็นสัตว์กินพืชและสัตว์กินทั้ง พืชและสัตว์ สมาชิกในกลุ่มย่อยโคเอลูโรซอเรีย ส่วนใหญ่ น่าจะมีขน ปกคลุมทั่วตัว และเป็นไปได้ว่าขนเหล่านี้อาจพบได้ในเทโรพอดกลุ่มอื่นๆ ด้วย ในยุคจูราสสิกนกได้วิวัฒนาการมาจากเทโรพอดโคเอลูโรซอเรียขนาดเล็กที่เชี่ยวชาญเฉพาะด้าน และปัจจุบันมีนกที่ยังมีชีวิตอยู่ประมาณ 11,000 ชนิด ทำให้เทโรพอดเป็นกลุ่มไดโนเสาร์เพียงกลุ่มเดียวที่ยังมีชีวิตอยู่ในปัจจุบัน
เทอโรพอดปรากฏตัวครั้งแรกในช่วง ยุค คาร์เนียนของยุคไทรแอสสิกตอนปลาย เมื่อ 231.4 ล้านปีก่อน ( Ma ) [ 3 ]และรวมถึงสัตว์กินเนื้อบนบกขนาดใหญ่ส่วนใหญ่ตั้งแต่ยุคจูราสสิกตอนต้นจนถึงสิ้นสุด ยุคครีเท เชียสเมื่อประมาณ 66 ล้านปีก่อน ซึ่งรวมถึงสัตว์กินเนื้อบนบกที่ใหญ่ที่สุดเท่าที่เคยมีมา เช่นไทแรนโนซอรัสและไจแกนโนโตซอรัสแม้ว่าเทอโรพอดที่ไม่ใช่นกจะมีขนาดที่หลากหลายมาก โดยเทอโรพอดที่ไม่ใช่นกบางชนิด เช่นสแกนโซริโอเทอริจิดมีขนาดไม่ใหญ่ไปกว่านกขนาดเล็ก
ชีววิทยา
ลักษณะเฉพาะ
มีการเสนอ synapomorphiesต่างๆสำหรับ Theropoda โดยอิงจาก taxa ที่รวมอยู่ในกลุ่ม ตัวอย่างเช่น บทความปี 1999 โดยPaul Serenoแนะนำว่า theropods มีลักษณะเฉพาะ เช่น ร่อง ectopterygoid (ร่องรอบกระดูก ectopterygoid) ข้อต่อ intramandibular ที่อยู่ในขากรรไกรล่างและโพรงภายในกระดูก ที่ รุนแรง[ 4 ]อย่างไรก็ตาม เนื่องจาก taxa เช่นHerrerasaurusอาจไม่ใช่ theropods ลักษณะเหล่านี้จึงอาจมีการกระจายอย่างกว้างขวางใน saurischians ยุคแรกมากกว่าที่จะเป็นเอกลักษณ์เฉพาะของ theropods
ในทางกลับกัน กลุ่มสิ่งมีชีวิตที่มีโอกาสอยู่ในกลุ่มเทโรโพดามากกว่า อาจมีลักษณะเฉพาะร่วมกันมากขึ้น เช่น ช่องเปิดขากรรไกร บนที่เด่นชัด กระดูกสันหลังส่วนคอที่มีช่องเปิดด้านข้างที่ส่วนหน้าของกระดูกสันหลังส่วนกลาง ทำให้คอมีลักษณะโปร่งมากขึ้น กระดูกสันหลังส่วน กระเบนเหน็บห้าชิ้นขึ้นไปกระดูกข้อมือที่ขยายใหญ่ขึ้น และส่วนปลายของกระดูกหน้าแข้งที่เว้าลงรวม ถึงลักษณะอื่นๆ อีกเล็กน้อยที่พบได้ทั่วทั้งโครงกระดูก เช่นเดียวกับซอโรโพโดมอร์ฟในยุคแรก นิ้วที่สองในมือของเทโรโพดาจะมีขนาดใหญ่ขึ้น เทโรโพดายังมี ข้อต่อแบบลูกบอลและเบ้าที่พัฒนามาอย่างดีใกล้กับคอและหัวอีกด้วย[ 5 ] [ 6 ]
เทอโรพอดส่วนใหญ่เป็นของกลุ่มนีโอเทอโรพอด (Neotheropoda) ซึ่งมีลักษณะเฉพาะคือการลดจำนวนกระดูกเท้าหลายชิ้น ทำให้เหลือรอยเท้าสามนิ้วบนพื้นเมื่อพวกมันเดิน (เท้าสามนิ้ว) นิ้วที่ 5 ลดจำนวนลงเหลือเพียงเศษซากในช่วงต้นของการวิวัฒนาการของเทอโรพอดและหายไปในช่วงปลายยุคไทรแอสสิกนิ้วที่ 1 ลดจำนวนลงและโดยทั่วไปจะไม่สัมผัสพื้น และลดจำนวนลงอย่างมากในบางสายพันธุ์[ 7 ]พวกมันยังขาดนิ้วที่ 5 บนมือและได้พัฒนากระดูกอก (furcula) ซึ่งเรียกอีกอย่างว่ากระดูกขอพร[ 5 ]นีโอเทอโรพอดในยุคแรก เช่น โคเอโลฟิซอยด์ (coelophysoids) มีรอยหยักที่เห็นได้ชัดในขากรรไกรบนที่เรียกว่าช่องว่างใต้จมูก (subnarial gap) อเวรอสทรานส์ (Averostrans) เป็นเทอโรพอดที่มีวิวัฒนาการมากที่สุดกลุ่มหนึ่งและประกอบด้วยเททานูรา (Tetanurae) และเซราโตซอเรีย (Ceratosauria) แม้ว่าบางคนเคยพิจารณาว่าโคเอโลฟิซอยด์และเซราโทซอร์อยู่ในกลุ่มเดียวกันเนื่องจากลักษณะต่างๆ เช่น สะโพกที่เชื่อมติดกัน แต่การศึกษาในภายหลังแสดงให้เห็นว่ามีความเป็นไปได้มากกว่าที่ลักษณะเหล่านี้เป็นลักษณะดั้งเดิมของนีโอเทอโรพอดและสูญหายไปในเททานูแรนพื้นฐาน[ 8 ]อเวรอสทรานส์และญาติใกล้ชิดของพวกมันมีความเชื่อมโยงกันโดยการสูญเสียส่วนที่เหลือของนิ้วที่ 5 อย่างสมบูรณ์ ฟันในขากรรไกรบนน้อยลง การเคลื่อนที่ของแถวฟันลงไปด้านล่างของขากรรไกรบน และช่องน้ำตา อเวรอสทรานส์ยังมีลักษณะร่วมกันในสะโพกและฟันอีกด้วย[ 9 ]
อาหารและฟัน

เทอโรพอดมีอาหารหลากหลาย ตั้งแต่กินแมลง กินพืช ไปจนถึงกินเนื้อ การกินเนื้ออย่างเดียวถือเป็นอาหารดั้งเดิมของเทอโรพอดมาโดยตลอด และในอดีต ความหลากหลายของอาหารถือเป็นลักษณะเฉพาะของเทอโรพอดที่เป็นนกเท่านั้น อย่างไรก็ตาม การค้นพบในช่วงปลายศตวรรษที่ 20 และต้นศตวรรษที่ 21 แสดงให้เห็นว่ามีอาหารหลากหลายแม้ในสายพันธุ์ดั้งเดิม[ 10 ]ฟอสซิลเทอโรพอดที่ค้นพบในยุคแรกๆ แสดงให้เห็นว่าพวกมันกินเนื้อ เป็นหลัก ตัวอย่างฟอสซิลของเทอโรพอดในยุคแรกๆ ที่นักวิทยาศาสตร์รู้จักในศตวรรษที่ 19 และต้นศตวรรษที่ 20 ล้วนมีฟันแหลมคมที่มีขอบหยักสำหรับตัดเนื้อ และบางตัวอย่างยังแสดงหลักฐานโดยตรงของพฤติกรรมการล่าเหยื่ออีกด้วย ตัวอย่างเช่น ฟอสซิล ของ Compsognathus longipesถูกพบโดยมีกิ้งก่าอยู่ในกระเพาะ และ ฟอสซิลของ Velociraptor mongoliensisถูกพบในสภาพกำลังต่อสู้กับProtoceratops andrewsi (ไดโนเสาร์ในกลุ่มออร์นิธิ สเชียนชนิดหนึ่ง) ซึ่งคาดว่าเกิดจากเนินทราย พัดทับ สัตว์ทั้งสองตัวขณะกำลังต่อสู้กัน ส่งผลให้เกิดการกลายเป็นฟอสซิล
ไดโนเสาร์เทอริซิโนซอร์เป็น ฟอสซิลกลุ่มแรกที่ได้รับการยืนยันว่าไม่กินเนื้อ สัตว์ เดิมทีรู้จักกันในชื่อ "เซกโนซอร์" ในตอนแรกคิดว่าเป็นโปรซอโรพอดแต่ต่อมาได้รับการพิสูจน์แล้วว่าเป็นเทอริซิโนซอร์ที่มีความเชี่ยวชาญสูง และกินพืชเป็นอาหาร เทอริ ซิโนซอร์มีท้องขนาดใหญ่สำหรับย่อยอาหารจากพืช และมีหัวขนาดเล็กที่มีจะงอยปากและฟันรูปใบไม้ การศึกษาเพิ่มเติมเกี่ยวกับเทอริซิโนซอร์ในกลุ่มมานิแรปทอแรนและความสัมพันธ์ของพวกมันแสดงให้เห็นว่าเทอริซิโนซอร์ไม่ใช่สมาชิกกลุ่มแรกๆ เพียงกลุ่มเดียวที่เลิกกินเนื้อสัตว์ สายพันธุ์อื่นๆ ของมานิแรปทอแรนในยุคแรกๆ อีกหลายสายพันธุ์แสดงให้เห็นถึงการปรับตัวเพื่อ กินอาหาร แบบกินทั้ง พืชและ สัตว์ รวมถึงการกินเมล็ดพืช ( โทรโอโดนทิด บางชนิด ) และการกินแมลง ( อะวิอาลันและอัลวาเรซซอร์ หลายชนิด ) โอวิแรปเตอร์ซอรัสออร์นิโทมิโมซอรัสและโทรโอโดนทิดขั้นสูงน่าจะเป็นสัตว์กินพืชและสัตว์กินเนื้อเช่นกัน และเทโรพอดบางชนิด (เช่นมาเซียกาซอรัส นอปเฟลรีและสไปโนซอริเด ) ดูเหมือนจะเชี่ยวชาญในการจับปลา[ 11 ] [ 12 ]
อาหารส่วนใหญ่สามารถอนุมานได้ จากลักษณะฟัน[ 13 ]รอยฟันบนกระดูกของเหยื่อ และเนื้อหาในลำไส้ เทอโรพอดบางชนิด เช่นBaryonyx , Lourinhanosaurus , ornithomimosaurs และนก เป็นที่ทราบกันว่าใช้gastrolithsหรือหินในกระเพาะ
ฟันของเทโรพอดส่วนใหญ่มีลักษณะคล้ายใบมีด มีรอยหยักที่ขอบ[ 14 ]เรียกว่าซิโฟดอนต์ ส่วนชนิดอื่น ๆ เป็นแบบแพคคีดอนต์หรือโฟลิดอนต์ ขึ้นอยู่กับรูปร่างของฟันหรือเดนทิเคิล[ 14 ]สัณฐานวิทยาของฟันมีความแตกต่างกันมากพอที่จะแยกแยะวงศ์หลัก ๆ ออกจากกันได้[ 13 ]ซึ่งบ่งชี้ถึงกลยุทธ์การกินที่แตกต่างกัน การตรวจสอบในเดือนกรกฎาคม 2015 พบว่าสิ่งที่ดูเหมือน "รอยแตก" ในฟันนั้น แท้จริงแล้วคือรอยพับที่ช่วยป้องกันฟันหักโดยการเสริมความแข็งแรงให้กับรอยหยักแต่ละรอยขณะที่พวกมันโจมตีเหยื่อ[ 15 ]รอยพับเหล่านี้ช่วยให้ฟันคงอยู่ในตำแหน่งได้นานขึ้น โดยเฉพาะอย่างยิ่งเมื่อเทโรพอดวิวัฒนาการให้มีขนาดใหญ่ขึ้นและมีแรงกัดมากขึ้น[ 16 ] [ 17 ]
เปลือกหุ้มร่างกาย (ผิวหนัง เกล็ด และขน)

เทโรพอ ดในยุคมี โซโซอิก มีความหลากหลายมากในแง่ของลักษณะพื้นผิวและการปกคลุมของผิวหนัง ขนหรือโครงสร้างคล้ายขน (เส้นใย) พบได้ในสายพันธุ์ส่วนใหญ่ของโคเอลูโรซอร์ (ดูไดโนเสาร์มีขน ) อย่างไรก็ตาม นอกเหนือจากโคเอลูโรซอร์แล้ว ขนอาจจำกัดอยู่เฉพาะในสายพันธุ์ที่อายุน้อยและมีขนาดเล็กกว่า หรือเฉพาะบางส่วนของร่างกายเท่านั้น เทโรพอดขนาดใหญ่หลายชนิดมีผิวหนังปกคลุมด้วยเกล็ดขนาดเล็กขรุขระ ในบางชนิด เกล็ดเหล่านี้จะแทรกด้วยเกล็ดขนาดใหญ่ ผิวหนังประเภทนี้เป็นที่รู้จักกันดีที่สุดในเซราโทซอร์คาร์โนทอรัสซึ่งได้รับการอนุรักษ์ไว้โดยมีร่องรอยของผิวหนังอย่างกว้างขวาง[ 18 ]ออสทีโอเดอร์ม ซึ่งเป็นเกล็ดที่มีแกนกระดูก พบได้ในเซราโทซอรัสซึ่งถูกค้นพบโดยมีส่วนของออสทีโอเดอร์มอยู่ด้านบนของคอและหาง ซึ่งอาจก่อตัวเป็นแถวต่อเนื่องกันในขณะที่ยังมีชีวิตอยู่[ 19 ]ในคาร์โนซอร์เกล็ดรูปสี่เหลี่ยมผืนผ้าพบได้ที่ด้านบนของเท้าและด้านล่างของหางในConcavenatorและที่ด้านล่างคอในAllosaurus [ 20 ]
มีหลักฐานว่าบางสายพันธุ์ของเทโรพอดมีขนเป็นบรรพบุรุษ แต่ได้สูญเสียขนไปและเปลี่ยนมาเป็นเกล็ดในสมาชิกรุ่นหลัง ในกลุ่มไทแรนโนซอรัสสมาชิกยุคแรก อย่าง DilongและYutyrannusมีหลักฐานของขน ในขณะที่ในกลุ่มไทแรนโนซอรัสรุ่นหลังเช่นTyrannosaurus , Tarbosaurus , Albertosaurus , GorgosaurusและDaspletosaurusมีหลักฐานของเกล็ด แม้ว่าจะไม่ทราบว่าพวกมันสูญเสียขนทั้งหมดไปหรือไม่[ 21 ]
สายพันธุ์โคเอลูโรซอร์ที่อยู่ห่างไกลจากนกมากที่สุดมีขนที่ค่อนข้างสั้นและประกอบด้วยเส้นใยที่เรียบง่าย อาจมีการแตกแขนง[ 22 ]เส้นใยที่เรียบง่ายยังพบได้ในเทอริซิโนซอร์ซึ่งมีขนขนาดใหญ่และแข็งคล้ายขนนก เทอโรพอดที่มีขนปกคลุมทั่วตัว เช่นดรอเมโอซอริเดมักจะเหลือเกล็ดไว้เฉพาะที่เท้าเท่านั้น บางชนิดอาจมีขนผสมอยู่บริเวณอื่นของร่างกายด้วยสแกนโซริโอเทอริกซ์มีเกล็ดอยู่ใกล้กับด้านล่างของหาง[ 23 ]และจูราเวเนเตอร์อาจมีเกล็ดเป็นส่วนใหญ่โดยมีเส้นใยที่เรียบง่ายแทรกอยู่บ้าง[ 24 ]ในทางกลับกัน เทอโรพอดบางชนิดมีขนปกคลุมทั่วตัว เช่น แอ นคิออร์ นิส ซึ่ง เป็นแอนคิออร์นิธิ ด ซึ่งมีขนแม้กระทั่งที่เท้าและนิ้วเท้า[ 25 ]
จากความสัมพันธ์ระหว่างขนาดฟันและความยาวกะโหลกศีรษะ รวมถึงการเปรียบเทียบระดับการสึกหรอของฟันของเทโรพอดที่ไม่ใช่นกและเลพิโดซอรัส ในปัจจุบัน สรุปได้ว่าเทโรพอดมีริมฝีปากที่ปกป้องฟันจากภายนอก เมื่อมองดูแล้ว จมูกของเทโรพอดอย่างDaspletosaurusมีความคล้ายคลึงกับกิ้งก่ามากกว่าจระเข้ ซึ่งไม่มีริมฝีปาก[ 26 ]
ขนาด


ไทแรนโนซอรัสเป็นไดโนเสาร์เทอโรพอดที่ใหญ่ที่สุดและเป็นที่รู้จักมากที่สุดในหมู่ประชาชนทั่วไปมานานหลายทศวรรษ อย่างไรก็ตาม นับตั้งแต่การค้นพบ ไดโนเสาร์กินเนื้อขนาดยักษ์อื่นๆ ที่มีขนาดใกล้เคียงกันก็ได้รับการอธิบายไว้เช่นกัน รวมถึงสไปโนซอรัส คาร์ชาโรดอนโตซอรัสและไจแกนโตซอรัส [ 27 ] ไดโนเสาร์เทอโรพอดขนาดใหญ่เหล่านี้คาดว่าจะมีความยาวมากกว่าไทแรนโนซอรัสโดยเฉพาะอย่างยิ่งในกรณีของสไปโนซอรัสแม้ว่าการศึกษาและการสร้างแบบจำลองล่าสุดจะแสดงให้เห็นว่าไทแรนโนซอรัสแม้จะสั้นกว่า แต่ก็มีโครงสร้างที่แข็งแรงกว่า ดังนั้นจึงยังคงเทียบได้กับพวกมันในแง่ของมวลและขนาดโดยรวม ตัวอย่าง ไทแรนโนซอรัส ที่ใหญ่ที่สุดที่รู้จัก เช่นซูและสก็อตตี้ปัจจุบันคาดว่าจะเป็นตัวอย่างเทอโรพอดแต่ละตัวที่มีมวลมากที่สุดเท่าที่วิทยาศาสตร์รู้จัก แม้ว่าจะเพียงเล็กน้อยก็ตาม ความลำเอียงอย่างมากในขนาดตัวอย่างที่เกิดจากจำนวนตัวอย่างผู้ใหญ่ใน Tyrannosaurus ที่มากกว่า เมื่อเทียบกับเทโรพอดตัวอื่นๆ ที่มีมวลใกล้เคียงกัน ประกอบกับเทโรพอดยักษ์ตัวเต็มวัยส่วนใหญ่ (แม้แต่กับ Tyrannosaurus ) ไม่เคยปรากฏในบันทึกฟอสซิล ทำให้ยากที่จะตัดสินว่าเทโรพอดตัวใดมีขนาดใหญ่ที่สุดตามมวลในความเป็นจริง [ 28 ]ยังไม่มีคำอธิบายที่ชัดเจนว่าทำไมเทโรพอดยักษ์จึงเติบโตจนมีขนาดใหญ่มากเมื่อเทียบกับสัตว์นักล่าบนบกอื่นๆ ที่มีมาก่อนและหลังพวกมัน
เทอโรพอดที่ใหญ่ที่สุดที่ยังมีชีวิตอยู่คือนกกระจอกเทศธรรมดาสูงถึง 2.74 เมตร (9 ฟุต) และมีน้ำหนักระหว่าง 90 ถึง 130 กิโลกรัม (200 – 290 ปอนด์) [ 29 ] เทอโรพอด ที่ ไม่ใช่นกที่เล็กที่สุดที่รู้จักจากตัวอย่างที่โตเต็มวัยคือAnchiornis huxleyiซึ่งมีน้ำหนัก 110 กรัมและยาว 34 เซนติเมตร (1 ฟุต) [ 25 ]เมื่อรวมนกในปัจจุบันเข้าไปด้วยนกฮัมมิงเบิร์ดผึ้ง ( Mellisuga helenae ) มีขนาดเล็กที่สุดที่ 1.9 กรัมและยาว 5.5 เซนติเมตร (2.2 นิ้ว) [ 30 ] [ 31 ]
ทฤษฎีล่าสุดเสนอว่าขนาดตัวของเทอโรพอดลดลงอย่างต่อเนื่องในช่วงระยะเวลา 50 ล้านปี จากเฉลี่ย 163 กิโลกรัม (359 ปอนด์) เหลือเพียง 0.8 กิโลกรัม (1.8 ปอนด์) และในที่สุดก็วิวัฒนาการเป็นนกสมัยใหม่กว่า 11,000 สายพันธุ์โดยอิงจากหลักฐานที่ว่าเทอโรพอดเป็นไดโนเสาร์เพียงชนิดเดียวที่มีขนาดเล็ลงอย่างต่อเนื่อง และโครงกระดูกของพวกมันเปลี่ยนแปลงเร็วกว่าไดโนเสาร์ชนิดอื่นถึงสี่เท่า[ 32 ] [ 33 ]
อัตราการเติบโต
เพื่อประมาณอัตราการเติบโตของเทโรพอด นักวิทยาศาสตร์จำเป็นต้องคำนวณทั้งอายุและมวลร่างกายของไดโนเสาร์ การวัดทั้งสองนี้สามารถคำนวณได้จากกระดูกและเนื้อเยื่อ ที่กลายเป็นฟอสซิลเท่านั้น ดังนั้นจึง ใช้ การวิเคราะห์การถดถอยและอัตราการเติบโตของสัตว์ที่ยังมีชีวิตอยู่เป็นตัวแทนในการคาดการณ์ กระดูกที่กลายเป็นฟอสซิลแสดงวงแหวนการเติบโตที่ปรากฏขึ้นอันเป็นผลมาจากการเติบโตหรือการเปลี่ยนแปลงตามฤดูกาล ซึ่งสามารถใช้เพื่อประมาณอายุ ณ เวลาที่เสียชีวิต[ 34 ]อย่างไรก็ตาม จำนวนวงแหวนในโครงกระดูกอาจแตกต่างกันไปในแต่ละกระดูก และวงแหวนเก่าๆ ก็อาจหายไปเมื่ออายุมากขึ้น ดังนั้นนักวิทยาศาสตร์จึงจำเป็นต้องควบคุมตัวแปรที่อาจทำให้เกิดความสับสนทั้งสองนี้อย่างเหมาะสม
การกำหนดมวลร่างกายทำได้ยากกว่า เนื่องจากมวลกระดูกคิดเป็นเพียงสัดส่วนเล็กน้อยของมวลร่างกายทั้งหมดของสัตว์ วิธีหนึ่งคือการวัดเส้นรอบวงของกระดูกต้นขา ซึ่งในไดโนเสาร์เทอโรพอดที่ไม่ใช่นกนั้นพบว่ามีสัดส่วนค่อนข้างใกล้เคียงกับสัตว์เลี้ยงลูกด้วยนมสี่ขา[ 35 ]และใช้การวัดนี้เป็นฟังก์ชันของน้ำหนักตัว เนื่องจากสัดส่วนของกระดูกยาวเช่นกระดูกต้นขาจะเติบโตตามสัดส่วนของมวลร่างกาย[ 35 ]วิธีการใช้สัดส่วนกระดูกของสัตว์ที่ยังมีชีวิตอยู่ต่ออัตราส่วนมวลร่างกายเพื่อทำนายเกี่ยวกับสัตว์ที่สูญพันธุ์เรียกว่าวิธีการปรับขนาดตามสัตว์ที่ยังมีชีวิตอยู่ (ES) [ 36 ]วิธีที่สองที่เรียกว่าวิธีการความหนาแน่นเชิงปริมาตร (VD) ใช้แบบจำลองโครงกระดูกขนาดเต็มเพื่ออนุมานเกี่ยวกับมวลที่เป็นไปได้ วิธีการ ES เหมาะสมกว่าสำหรับการศึกษาในวงกว้างซึ่งรวมถึงตัวอย่างจำนวนมาก และไม่จำเป็นต้องใช้โครงกระดูกที่สมบูรณ์เท่ากับวิธีการ VD แต่วิธีการ VD ช่วยให้นักวิทยาศาสตร์สามารถตอบคำถามทางสรีรวิทยาเกี่ยวกับสัตว์ได้ดียิ่งขึ้น เช่นการเคลื่อนไหวและจุดศูนย์ถ่วง[ 36 ]
ปัจจุบันเป็นที่ยอมรับกันว่าเทโรพอดที่ไม่ใช่นกไม่ได้แสดงอัตราการเติบโตที่เท่ากันทั้งกลุ่ม แต่มีอัตราที่แตกต่างกันไปตามขนาด อย่างไรก็ตาม เทโรพอดที่ไม่ใช่นกทุกตัวมีอัตราการเติบโตเร็วกว่าสัตว์เลื้อยคลานในปัจจุบัน แม้กระทั่งเมื่อปรับขนาดสัตว์เลื้อยคลานในปัจจุบันให้มีขนาดใหญ่เท่ากับเทโรพอดที่ไม่ใช่นกบางชนิดก็ตาม เมื่อมวลร่างกายเพิ่มขึ้น อัตราการเติบโตสัมพัทธ์ก็เพิ่มขึ้นด้วย แนวโน้มนี้อาจเกิดจากความจำเป็นในการไปถึงขนาดที่จำเป็นสำหรับการเจริญเติบโตเต็มที่เพื่อการสืบพันธุ์ [ 37 ] ตัวอย่างเช่น เทโรพอดที่เล็กที่สุดที่รู้จักตัวหนึ่งคือMicroraptor zhaoianusซึ่งมีมวลร่างกาย 200 กรัม เติบโตในอัตราประมาณ 0.33 กรัมต่อวัน[ 38 ]สัตว์เลื้อยคลานที่มีขนาดใกล้เคียงกันจะเติบโตในอัตราครึ่งหนึ่งของอัตรานี้ อัตราการเติบโตของเทโรพอดที่ไม่ใช่นกขนาดกลาง (100–1000 กก.) ใกล้เคียงกับนกที่โตเต็มวัย ซึ่งช้ากว่านกที่ยังไม่โตเต็มวัยมาก เทโรพอดขนาดใหญ่ (1500–3500 กก.) เติบโตเร็วขึ้นไปอีก คล้ายกับอัตราที่พบในสัตว์เลี้ยงลูก ด้วยนมยูเทอเรียน [ 38 ]เทโรพอดที่ไม่ใช่นกที่ใหญ่ที่สุด เช่นไทแรนโนซอรัส เร็กซ์ มีพลวัตการเติบโตคล้ายกับสัตว์บกที่ใหญ่ที่สุดที่ยังมีชีวิตอยู่ในปัจจุบัน คือ ช้างแอฟริกาซึ่งมีลักษณะเด่นคือช่วงการเติบโตอย่างรวดเร็วจนถึงวัยเจริญพันธุ์ จากนั้นจึงเติบโตช้าลงในวัยผู้ใหญ่[ 39 ]
ท่าทางและการเดิน

เนื่องจากเป็นกลุ่มสัตว์ที่มีความหลากหลายอย่างมาก ท่าทางที่เทโรพอดใช้จึงน่าจะแตกต่างกันอย่างมากระหว่างสายพันธุ์ต่างๆ ตลอดช่วงเวลา[ 40 ]เทโรพอดที่รู้จักทั้งหมดเป็นสัตว์สองขาโดยมีแขนขาหน้าสั้นลงและมีความเชี่ยวชาญสำหรับงานที่หลากหลาย (ดูด้านล่าง) ในนกสมัยใหม่ ร่างกายมักจะอยู่ในท่าที่ค่อนข้างตั้งตรง โดยขาบน (กระดูกต้นขา) ขนานกับกระดูกสันหลัง และแรงผลักดันไปข้างหน้าของการเคลื่อนที่เกิดขึ้นที่หัวเข่า นักวิทยาศาสตร์ยังไม่แน่ใจว่าท่าทางและการเคลื่อนที่แบบนี้มีมาแต่โบราณในวงศ์เทโรพอดนานแค่ไหน[ 40 ]
ไดโนเสาร์เทอโรพอดที่ไม่ใช่นกได้รับการยอมรับว่าเป็นสัตว์สองขาเป็นครั้งแรกในช่วงศตวรรษที่ 19 ก่อนที่ความสัมพันธ์ของพวกมันกับนกจะเป็นที่ยอมรับกันอย่างกว้างขวาง ในช่วงเวลานั้น ไดโนเสาร์เทอโรพอด เช่นคาร์โนซอร์และไทแรนโนซอริเดถูกคิดว่าเดินโดยใช้กระดูกต้นขาและกระดูกสันหลังในแนวตั้งในท่าทางที่ยืนตรงเกือบตั้งตรง โดยใช้หางที่ยาวและมีกล้ามเนื้อเป็นส่วนช่วยพยุงเพิ่มเติมในท่าทางสามขาคล้ายจิงโจ้[ 40 ]ตั้งแต่ทศวรรษที่ 1970 การศึกษาทางชีวกลศาสตร์ของไดโนเสาร์เทอโรพอดขนาดยักษ์ที่สูญพันธุ์ไปแล้วทำให้เกิดข้อสงสัยเกี่ยวกับการตีความนี้ การศึกษาเกี่ยวกับการเชื่อมต่อของกระดูกแขนขาและการขาดหลักฐานร่องรอยการลากหางชี้ให้เห็นว่า เมื่อเดิน ไดโนเสาร์เทอโรพอดขนาดยักษ์ที่มีหางยาวจะใช้ท่าทางในแนวนอนมากขึ้นโดยให้หางขนานกับพื้น[ 40 ] [ 41 ]อย่างไรก็ตาม การวางแนวของขาในสายพันธุ์เหล่านี้ขณะเดินยังคงเป็นที่ถกเถียงกันอยู่ การศึกษาบางชิ้นสนับสนุนการวางแนวกระดูกต้นขาในแนวตั้งแบบดั้งเดิม อย่างน้อยก็ในเทโรพอดหางยาวที่ใหญ่ที่สุด[ 41 ]ในขณะที่การศึกษาอื่นๆ ชี้ให้เห็นว่าโดยปกติแล้วหัวเข่าจะงออย่างมากในเทโรพอดทุกตัวขณะเดิน แม้แต่ยักษ์อย่างไทแรนโนซอริเด[ 42 ] [ 43 ]เป็นไปได้ว่าท่าทาง การยืน และการเดินที่หลากหลายมีอยู่ในกลุ่มเทโรพอดที่สูญพันธุ์ไปแล้วหลายกลุ่ม[ 40 ] [ 44 ]
ระบบประสาทและประสาทสัมผัส
แม้ว่าจะหายาก แต่ก็พบแบบจำลองกะโหลกศีรษะชั้นในของเทโรพอดที่สมบูรณ์จากฟอสซิล นอกจากนี้ยังสามารถสร้างกะโหลกศีรษะชั้นในของเทโรพอดขึ้นใหม่จากกะโหลกสมองที่เก็บรักษาไว้ได้โดยไม่ทำให้ตัวอย่างที่มีค่าเสียหาย โดยใช้การสแกนเอกซเรย์คอมพิวเตอร์และ ซอฟต์แวร์ สร้างภาพสามมิติการค้นพบเหล่านี้มีความสำคัญทางวิวัฒนาการเพราะช่วยบันทึกการเกิดขึ้นของระบบประสาทของนกสมัยใหม่จากสัตว์เลื้อยคลานในยุคก่อน การเพิ่มขึ้นของสัดส่วนของสมองที่ถูกครอบครองโดยสมองใหญ่ดูเหมือนจะเกิดขึ้นพร้อมกับการกำเนิดของโคเอลูโรซอเรียและ "ดำเนินต่อไปตลอดวิวัฒนาการของมานิแรปทอแรนและนกยุคแรก" [ 45 ]
การศึกษาแสดงให้เห็นว่าเทโรพอดมีจมูกที่ไวมาก มีการเสนอแนะว่าจมูกเหล่านี้อาจใช้สำหรับการตรวจจับอุณหภูมิ พฤติกรรมการกินอาหาร และการตรวจจับคลื่น[ 46 ] [ 47 ]
สัณฐานวิทยาของแขนขาหน้า

แขนขาหน้าที่สั้นกว่าเมื่อเทียบกับขาหลังเป็นลักษณะทั่วไปในกลุ่มเทอโรพอด โดยเฉพาะอย่างยิ่งในกลุ่มอะเบลิซอริเด (เช่นคาร์โนทอรัส ) และกลุ่มไทแรนโนซอริเด (เช่นไทแรนโนซอรัส ) อย่างไรก็ตาม ลักษณะนี้ไม่ได้เป็นลักษณะทั่วไปทั้งหมด สไปโนซอริเดมีแขนขาหน้าที่พัฒนาดี เช่นเดียวกับโคเอลูโรซอรัสหลายชนิด แขนขาหน้าที่แข็งแรงของสกุลหนึ่งคือซวนหานซอรัสทำให้ดง จื้อหมิงเสนอว่าสัตว์ชนิดนี้อาจเดินสี่ขา[ 48 ]อย่างไรก็ตาม ปัจจุบันไม่คิดว่าจะเป็นไปได้อีกต่อไป[ 49 ]
มือก็แตกต่างกันมากในกลุ่มต่างๆ รูปแบบที่พบได้บ่อยที่สุดในเทโรพอดที่ไม่ใช่นกคืออวัยวะที่ประกอบด้วยนิ้วสามนิ้ว ได้แก่ นิ้วที่ 1, 2 และ 3 (หรืออาจจะเป็นนิ้วที่2, 3 และ 4 ) พร้อมกรงเล็บแหลมคม เทโรพอดพื้นฐานบางชนิด เช่นเซราโทซอเรียน ส่วนใหญ่ มีสี่นิ้ว และยังมี กระดูก ฝ่ามือที่ 5 ที่ลดขนาดลง (เช่นไดโลโฟซอรัส ) เททานูรันส่วนใหญ่มีสามนิ้ว[ a ] [ 50 ]แต่บางชนิดมีน้อยกว่านั้น[ 51 ]
เชื่อกันว่าขอบเขตการใช้งานของแขนขาหน้าก็แตกต่างกันไปในแต่ละวงศ์เช่นกัน สไปโนซอริเดอาจใช้แขนขาหน้าที่แข็งแรงในการจับปลา แมนิแรปทอแรนขนาดเล็กบางชนิด เช่นสแกนโซริโอเทอริจิดเชื่อกันว่าใช้แขนขาหน้าใน การปี นต้นไม้[ 23 ]ปีกของนกในปัจจุบันส่วนใหญ่ใช้สำหรับการบิน แม้ว่าจะมีการปรับเปลี่ยนเพื่อวัตถุประสงค์อื่นในบางกลุ่มก็ตาม ตัวอย่างเช่น นกน้ำ เช่นเพนกวินใช้ปีกของพวกมันเป็นครีบ
การเคลื่อนไหวของแขนขาหน้า
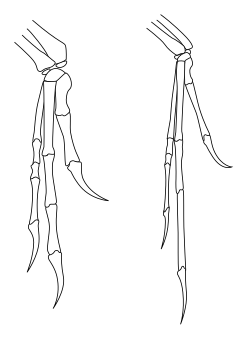
ตรงกันข้ามกับวิธีที่เทโรพอดมักถูกสร้างขึ้นใหม่ในงานศิลปะและสื่อยอดนิยม ช่วงการเคลื่อนไหวของแขนขาหน้าของเทโรพอดนั้นถูกจำกัดอย่างมาก โดยเฉพาะอย่างยิ่งเมื่อเทียบกับความคล่องแคล่วของแขนขาหน้าของมนุษย์และไพรเมตอื่น ๆ [ 52 ]ที่สำคัญที่สุด เทโรพอดและไดโนเสาร์ซอริสเชียนที่เดินสองขาอื่นๆ (รวมถึงโปรซอโร พอดที่เดินสองขา ) ไม่สามารถคว่ำมือได้ นั่นคือ พวกมันไม่สามารถหมุนแขนท่อนล่างเพื่อให้ฝ่ามือหันลงพื้นหรือหันไปทางขาได้ ในมนุษย์ การคว่ำมือเกิดขึ้นจากการเคลื่อนไหวของกระดูกเร เดียส เทียบกับกระดูกอัลนา (กระดูกสองชิ้นของแขนท่อนล่าง) อย่างไรก็ตาม ในไดโนเสาร์ซอริสเชียน ปลายของกระดูกเรเดียสใกล้ข้อศอกนั้นถูกล็อกไว้ในร่องของกระดูกอัลนา ทำให้ไม่สามารถเคลื่อนไหวได้ การเคลื่อนไหวที่ข้อมือก็ถูกจำกัดในหลายๆ สปีชีส์ ทำให้แขนท่อนล่างและมือทั้งหมดต้องเคลื่อนไหวเป็นหน่วยเดียวโดยมีความยืดหยุ่นน้อย[ 53 ]ในเทอโรพอดและโปรซอโรพอด วิธีเดียวที่ฝ่ามือจะหันลงพื้นได้คือการกางแขนขาหน้าทั้งหมดออกไปด้านข้าง เหมือนกับนกที่ยกปีกขึ้น[ 52 ]
ในคาร์โนซอร์ เช่นอะโครแคนโทซอรัสมือยังคงมีความยืดหยุ่นค่อนข้างสูง โดยมีนิ้วที่เคลื่อนไหวได้ สิ่งนี้เป็นจริงสำหรับเทโรพอดดั้งเดิม เช่นเฮอร์เรราซอร์โคเอลูโรซอร์แสดงให้เห็นถึงการเปลี่ยนแปลงในการใช้แขนท่อนล่าง โดยมีความยืดหยุ่นมากขึ้นที่ไหล่ทำให้สามารถยกแขนขึ้นไปในระนาบแนวนอนได้ และยิ่งมากขึ้นในนกที่บินได้ อย่างไรก็ตาม ในโคเอลูโรซอร์ เช่น ออร์นิโทมิโมซอร์ และโดยเฉพาะอย่างยิ่งดรอเมโอซอริเด มือได้สูญเสียความยืดหยุ่นไปมาก โดยมีนิ้วที่ไม่ยืดหยุ่นอย่างมาก ดรอเมโอซอริเดและมานิแรปทอแรนอื่นๆ ยังแสดงให้เห็นถึงการเคลื่อนไหวที่เพิ่มขึ้นที่ข้อมือซึ่งไม่พบในเทโรพอดอื่นๆ เนื่องจากการมีกระดูกข้อมือรูปครึ่งวงกลม (กระดูกข้อมือรูปครึ่งวงกลม) ที่ช่วยให้มือทั้งหมดพับไปด้านหลังเข้าหาแขนท่อนล่างในลักษณะเดียวกับนกในปัจจุบัน[ 53 ]
พยาธิวิทยาโบราณ
ในปี 2001 ราล์ฟ อี. มอลนาร์ได้ตีพิมพ์ผลการสำรวจพยาธิสภาพในกระดูกไดโนเสาร์เทอโรพอด เขาพบพยาธิสภาพใน 21 สกุลจาก 10 วงศ์ พยาธิสภาพเหล่านี้พบในเทอโรพอดทุกขนาดลำตัว แม้ว่าจะพบได้น้อยในฟอสซิลของเทอโรพอดขนาดเล็ก ซึ่งอาจเป็นผลจากกระบวนการเก็บรักษา พยาธิสภาพเหล่านี้กระจายอยู่ทั่วส่วนต่างๆ ของร่างกายเทอโรพอดอย่างกว้างขวาง บริเวณที่พบร่องรอยการบาดเจ็บและโรคในไดโนเสาร์เทอโรพอดมากที่สุดคือซี่โครงและกระดูกสันหลังส่วนหางแม้ว่าจะมีร่องรอยการบาดเจ็บมากมายในซี่โครงและกระดูกสันหลัง แต่ดูเหมือนว่าการบาดเจ็บจะ "ไม่มี...หรือพบได้น้อยมาก" ในกระดูกหลักที่รองรับน้ำหนักของร่างกาย เช่นกระดูกเชิงกรานกระดูกต้นขาและกระดูกหน้าแข้งการที่ไม่มีร่องรอยการบาดเจ็บในกระดูกเหล่านี้บ่งชี้ว่ากระดูกเหล่านี้ได้รับการคัดเลือกโดยวิวัฒนาการให้มีความต้านทานต่อการแตกหัก บริเวณที่พบร่องรอยการบาดเจ็บน้อยที่สุดคือกะโหลกศีรษะและแขนขาหน้า โดยพบการบาดเจ็บในความถี่ที่ใกล้เคียงกันในแต่ละบริเวณ พยาธิสภาพส่วนใหญ่ที่พบในฟอสซิลของเทโรพอดคือร่องรอยของการบาดเจ็บ เช่น กระดูกหัก รอยบุ๋ม และรอยเจาะ ซึ่งมักเกิดจากการกัด พยาธิสภาพโบราณของเทโรพอดบางอย่างดูเหมือนจะเป็นหลักฐานของการติดเชื้อซึ่งมักจำกัดอยู่เฉพาะบริเวณเล็กๆ ของร่างกายสัตว์เท่านั้น นอกจากนี้ยังพบหลักฐานของความผิดปกติแต่กำเนิดในซากของเทโรพอด การค้นพบดังกล่าวสามารถให้ข้อมูลที่เป็นประโยชน์สำหรับการทำความเข้าใจประวัติวิวัฒนาการของกระบวนการพัฒนาทางชีวภาพการเชื่อมติดกัน ที่ผิดปกติ ในส่วนประกอบของกะโหลกศีรษะหรือความไม่สมมาตรในส่วนประกอบเหล่านั้น อาจเป็นหลักฐานว่ากำลังตรวจสอบฟอสซิลของสิ่งมีชีวิตที่มีอายุมากเป็นพิเศษมากกว่าสิ่งมีชีวิตที่เป็นโรค[ 54 ]
การว่ายน้ำ
ร่องรอยการว่ายน้ำของเทโรพอด ซึ่งเป็นร่องรอยแรกในประเทศจีนของสกุลร่องรอยที่ชื่อว่าCharacichnosถูกค้นพบที่ชั้นหินเฟยเทียนซานในมณฑลเสฉวน[ 55 ]ร่องรอยการว่ายน้ำเหล่านี้สนับสนุนสมมติฐานที่ว่าเทโรพอดปรับตัวให้ว่ายน้ำได้และสามารถเคลื่อนที่ในน้ำที่มีความลึกปานกลางได้ ร่องรอยการว่ายน้ำของไดโนเสาร์ถือเป็นฟอสซิลร่องรอยที่หายาก และอยู่ในกลุ่มร่องรอยการว่ายน้ำของสัตว์มีกระดูกสันหลัง ซึ่งรวมถึงร่องรอยของเทโรซอร์และจระเข้ด้วย การศึกษานี้ได้อธิบายและวิเคราะห์แม่พิมพ์ธรรมชาติที่สมบูรณ์สี่แม่พิมพ์ของรอยเท้าเทโรพอด ซึ่งปัจจุบันเก็บรักษาไว้ที่ศูนย์วิจัยและพัฒนาร่องรอยไดโนเสาร์ Huaxia (HDT) รอยเท้าไดโนเสาร์เหล่านี้แท้จริงแล้วเป็นรอยกรงเล็บ ซึ่งบ่งชี้ว่าเทโรพอดตัวนี้ว่ายน้ำอยู่ใกล้ผิวน้ำ และมีเพียงปลายนิ้วเท้าและกรงเล็บเท่านั้นที่สัมผัสพื้นด้านล่าง ร่องรอยเหล่านี้แสดงให้เห็นถึงการเคลื่อนที่แบบประสานกัน ซ้าย-ขวา ซ้าย-ขวา ซึ่งสนับสนุนข้อเสนอที่ว่าเทโรพอดเป็นนักว่ายน้ำที่มีการประสานงานที่ดี[ 55 ]
ประวัติวิวัฒนาการ

ในช่วงปลายยุคไทรแอสสิก ไดโนเสาร์กลุ่มโปรโตเทโรพอดและเทโรพอดดั้งเดิมจำนวนมากดำรงอยู่และวิวัฒนาการควบคู่กันไป
ไดโนเสาร์เทอโรพอดที่เก่าแก่และดั้งเดิมที่สุดคือEodromaeus ซึ่ง เป็นไดโนเสาร์กินเนื้อ และอาจจะเป็นเฮอร์เรรา ซอริเดส จากอาร์เจนตินาเฮอร์เรราซอรัสมีชีวิตอยู่ในช่วงต้นยุคไทรแอสสิกตอนปลาย (ปลายยุคคาร์เนียนถึงต้นยุคโนเรียน ) พบได้ในทวีปอเมริกาเหนือและอเมริกาใต้ และอาจพบในอินเดียและแอฟริกาใต้ด้วย เฮอร์เรราซอรัสมีลักษณะผสมผสานระหว่างลักษณะดั้งเดิมและลักษณะที่ก้าวหน้า นักบรรพชีวินวิทยาบางคนในอดีตเคยพิจารณาว่าเฮอร์เรราซอรัสเป็นสมาชิกของเทอโรพอด ในขณะที่คนอื่นๆ ตั้งทฤษฎีว่ากลุ่มนี้เป็น ซอริสเชียน ขั้นพื้นฐานและอาจวิวัฒนาการมาก่อนการแยกตัวของซอริสเชียน-ออร์นิธิสเชียน การวิเคราะห์ทางคลัดิสติกส์หลังจากการค้นพบทาวา ซึ่ง เป็นไดโนเสาร์ไทรแอสสิกอีกชนิดหนึ่ง ชี้ให้เห็นว่าเฮอร์เรราซอรัสน่าจะเป็นเทอโรพอดในยุคแรก[ 56 ]
ไดโนเสาร์เทอโรพอดที่เก่าแก่ที่สุดและดั้งเดิมที่สุดอย่างชัดเจนคือกลุ่ม โคเอโลฟิซอยด์ ( Coelophysoidea ) โคเอโลฟิซอยด์เป็นกลุ่มสัตว์ที่มีการกระจายตัวอย่างกว้างขวาง มีโครงสร้างเบา และอาจอยู่รวมกันเป็นฝูง พวกมันรวมถึงนักล่าขนาดเล็ก เช่นโคเอโลฟิซิส (Coelophysis ) และแคมโปซอรัส (Camposaurus ) สัตว์ที่ประสบความสำเร็จเหล่านี้ดำรงอยู่ตั้งแต่ยุคคาร์เนียนตอนปลาย (ต้นยุคไทรแอสสิกตอนปลาย) ไปจนถึงยุคทัวร์เซียน (ปลายยุคจูราสสิกตอนต้น ) แม้ว่าในการจำแนกประเภทคลัดิสติกในยุคแรก พวกมันจะถูกรวมอยู่ภายใต้ กลุ่มเซราโตซอเรี ย (Ceratosauria ) และถือว่าเป็นสาขาย่อยของเทอโรพอดที่ก้าวหน้ากว่า[ 57 ]แต่พวกมันอาจเป็นบรรพบุรุษของเทอโรพอดอื่นๆ ทั้งหมด (ซึ่งจะทำให้พวกมันเป็น กลุ่ม พาราไฟเลติก ) [ 58 ] [ 59 ]
นีโอเธอโรโพดา (หมายถึง "เทโรพอดใหม่") เป็นกลุ่มที่รวมถึงโคเอโลฟิซอยด์ และ ไดโนเสาร์เทโรพอ ด ที่ก้าวหน้ากว่าและเป็นกลุ่มเทโรพอดเพียงกลุ่มเดียวที่รอดชีวิต จาก เหตุการณ์การสูญพันธุ์ในยุคไทรแอสสิก-จูราสสิก นีโอเธอโรโพดาได้รับการตั้งชื่อโดยRT Bakkerในปี 1986 โดยเป็นกลุ่มที่รวมถึงกลุ่มย่อยเทโรพอดที่พัฒนาแล้วอย่างCeratosauria และ Tetanurae และไม่รวมโคเอโลฟิซอยด์ [ 60 ]อย่างไรก็ตามนักวิจัยส่วนใหญ่ในภายหลังได้ใช้ชื่อนี้เพื่อหมายถึงกลุ่มที่กว้างกว่า นีโอเธอโรโพดาได้รับการกำหนดให้เป็นกลุ่ม ครั้งแรก โดยPaul Serenoในปี 1998 โดยเป็นโคเอโลฟิซิส บวกกับ นกสมัยใหม่ซึ่งรวมถึงเทโรพอดเกือบทั้งหมด ยกเว้นสายพันธุ์ที่ดั้งเดิมที่สุด[ 61 ]เดิมที Dilophosauridae ถูกพิจารณาว่าเป็นกลุ่มเล็กๆ ภายในนีโอเธอโรโพดา แต่ต่อมาถูกพิจารณาว่าเป็นกลุ่มพาราไฟเลติก เมื่อถึงยุคจูราสสิกตอนต้นนีโอเทอโรพอดที่ไม่ใช่อะเวอรอสแตรนทั้งหมดก็สูญพันธุ์ไป[ 62 ]
Averostra (หรือ "จมูกนก") เป็นกลุ่มย่อยภายใน Neotheropoda ซึ่งรวมถึงไดโนเสาร์เทอโรพอด ส่วนใหญ่ ได้แก่CeratosauriaและTetanuraeเป็นกลุ่มเดียวของเทอโรพอดหลังยุคจูราสสิกตอนต้น คุณลักษณะสำคัญอย่างหนึ่งของ Averostra คือการไม่มีกระดูกฝ่ามือชิ้นที่ห้า ซอริสเชียนอื่นๆยังคงมีกระดูกชิ้นนี้อยู่ แม้ว่าจะลดขนาดลงอย่างมากก็ตาม[ 63 ]
ไดโนเสาร์กลุ่มเซราโทซอร์ที่มีวิวัฒนาการค่อนข้างสูงกว่า(รวมถึงเซราโทซอรัสและคาร์โนทอรัส ) ปรากฏตัวขึ้นในช่วงต้นยุคจูราสสิกและดำรงอยู่จนถึงปลายยุคจูราสสิกในทวีปโลราเซียพวกมันแข่งขันกับญาติของพวกมันในกลุ่มเททานูรานที่มีโครงสร้างทางกายวิภาคที่ก้าวหน้ากว่า และในรูปแบบของ สายพันธุ์ อะเบลิซอร์ก็ดำรงอยู่จนถึงสิ้นสุดยุคครีเทเชียสในทวีปก็อนด์วานา
ไดโนเสาร์กลุ่ม Tetanuraeมีความเชี่ยวชาญเฉพาะด้านมากกว่าไดโนเสาร์กลุ่ม Ceratosauras พวกมันถูกแบ่งย่อยออกเป็นกลุ่มMegalosauroidea (หรือSpinosauroidea ) ซึ่งเป็นกลุ่มพื้นฐาน และกลุ่ม Avetheropoda ซึ่งเป็นกลุ่มที่พัฒนาขึ้นมามากกว่าMegalosauridae เป็นไดโนเสาร์นักล่าในยุคจูราสสิกตอนกลางถึงยุคครีเทเชียสตอนต้นเป็นหลัก และซากดึกดำบรรพ์ของญาติๆ อย่าง Spinosaurid ส่วนใหญ่มาจากหินในยุคครีเทเชียสตอนต้นและตอนกลาง ส่วน Avetheropoda นั้น ตามชื่อของมันบ่งบอกว่ามีความสัมพันธ์ใกล้ชิดกับนกมากกว่า และถูกแบ่งออกเป็นAllosauroidea ( ไดโนเสาร์กลุ่ม Carcharodontosaurs ที่มีความ หลากหลาย) และ Coelurosauria (กลุ่มไดโนเสาร์ขนาดใหญ่และมีความหลากหลายมาก ซึ่งรวมถึงนกด้วย)
ดังนั้น ในช่วงปลายยุคจูราสสิก จึงมีไดโนเสาร์เทอโรพอดอย่างน้อยสี่สายพันธุ์ที่แตกต่างกัน ได้แก่ เซราโทซอรัส เมกาโลซอรัส อัลโลซอรัส และโคเอลูโรซอรัส ซึ่งล่าไดโนเสาร์กินพืชขนาดเล็กและขนาดใหญ่จำนวนมาก ทั้งสี่กลุ่มนี้มีชีวิตรอดมาจนถึงยุคครีเทเชียส และสองกลุ่มนั้น ได้แก่ เซราโทซอรัสและโคเอลูโรซอรัส มีชีวิตรอดจนถึงสิ้นสุดยุคนั้น โดยพวกมันแยกกันอยู่ตามภูมิศาสตร์ เซราโทซอรัสอยู่ในกอนด์วานาและยุโรป ส่วนโคเอลูโรซอรัสอยู่ในลอราเซีย
ในบรรดากลุ่มเทโรพอดทั้งหมด โคเอลูโรซอร์เป็นกลุ่มที่มีความหลากหลายมากที่สุด กลุ่มโคเอลูโรซอร์บางกลุ่มที่เจริญรุ่งเรืองในช่วงยุคครีเทเชียส ได้แก่ ไทแรนโนซอริเด ( รวมถึง ไทแรนโนซอรัส ) ดรอเมโอซอริเด (รวมถึงเวโลซิแรปเตอร์และไดโนนิคัสซึ่งมีรูปร่างคล้ายกับนกที่เก่าแก่ที่สุดชนิดหนึ่งที่รู้จักกันคืออาร์คีออปเทอริกซ์ ) [ 64 ] [ 65 ]โทรโอโดนทิดและโอวิแรปโทซอรัสที่มีลักษณะคล้ายนก ออร์นิโทมิโมซอรัส (หรือ "ไดโนเสาร์นกกระจอกเทศ") เทริซิโนซอรัสกินพืชที่มีกรงเล็บยักษ์ที่แปลกประหลาด และเอวิอาลัน ซึ่งรวมถึงนกในปัจจุบันและเป็นสายพันธุ์ไดโนเสาร์เพียงสายพันธุ์เดียวที่รอดชีวิตจากเหตุการณ์การสูญพันธุ์ในยุคครีเทเชียส-พาลีโอจีน [ 66 ] แม้ว่ารากเหง้าของกลุ่มต่างๆ เหล่านี้จะพบได้ในยุคจูราสสิกตอนกลาง แต่พวกมันกลับมีจำนวนมากในยุคครีเทเชียสตอนต้นเท่านั้น นักบรรพชีวินวิทยาบางคน เช่นGregory S. Paulได้เสนอแนะว่าเทโรพอดขั้นสูงเหล่านี้บางส่วนหรือทั้งหมดสืบเชื้อสายมาจากไดโนเสาร์บินได้หรือนกยุคแรก เช่นArchaeopteryxที่สูญเสียความสามารถในการบินและกลับมาอาศัยอยู่ในถิ่นที่อยู่บนบก[ 67 ]
มีการรายงานถึงวิวัฒนาการของนกจากไดโนเสาร์เทอโรพอดชนิดอื่น โดยมีลักษณะเชื่อมโยงบางประการ ได้แก่กระดูกอก (กระดูกอกรูปตัววี) กระดูกที่มีโพรงอากาศ การกกไข่และ (อย่างน้อยในโคเอลูโรซอร์ ) ขน[ 32 ] [ 33 ] [ 68 ]
การจำแนกประเภท
ประวัติการจำแนกประเภท

OC Marshเป็นผู้บัญญัติศัพท์ชื่อ Theropoda (หมายถึง "เท้าสัตว์ร้าย") ในปี 1881 [ 69 ]ในตอนแรก Marsh ตั้งชื่อ Theropoda เป็นอันดับย่อยเพื่อรวมวงศ์Allosauridaeแต่ต่อมาได้ขยายขอบเขตและจัดลำดับใหม่เป็นอันดับเพื่อรวมวงศ์ไดโนเสาร์ "กินเนื้อ" หลากหลายวงศ์ รวมถึงMegalosauridae , Compsognathidae , Ornithomimidae , PlateosauridaeและAnchisauridae (ปัจจุบันทราบกันว่าเป็นซอโรโพโดมอร์ ฟกินพืช ) และHallopodidae (ต่อมาเปิดเผยว่าเป็นญาติของจระเข้) เนื่องจากขอบเขตของอันดับ Theropoda ของ Marsh ทำให้มันเข้ามาแทนที่กลุ่มอนุกรมวิธานก่อนหน้านี้ที่ED Cope คู่แข่งของ Marsh สร้างขึ้นในปี 1866 สำหรับไดโนเสาร์กินเนื้อ: Goniopoda ("เท้าเหลี่ยม") [ 49 ]
ในช่วงต้นศตวรรษที่ 20 นักบรรพชีวินวิทยาบางคน เช่นฟรีดริช ฟอน ฮูเอเนไม่ได้มองว่าไดโนเสาร์กินเนื้อเป็นกลุ่มธรรมชาติอีกต่อไป ฮูเอเนละทิ้งชื่อ "เทโรโพดา" และหันมาใช้ชื่อ อันดับซอ ริสเคีย (Saurischia ) ของแฮร์รี ซีลีย์ แทน ซึ่งฮูเอเนได้แบ่งออกเป็นอันดับย่อย โคเอ ลูโรซอเรีย (Coelurosauria ) และ แพคีโพโดซอเรีย ( Pachypodosauria ) ฮูเอเนจัดกลุ่มเทโรโพดาขนาดเล็กส่วนใหญ่ไว้ในโคเอลูโรซอเรีย และจัดเทโรโพดาขนาดใหญ่และโปรซอโรโพดาไว้ในแพคีโพโดซอเรีย ซึ่งเขาถือว่าเป็นบรรพบุรุษของซอโรโพดา (ในเวลานั้น โปรซอโรโพดายังคงถูกมองว่าเป็นสัตว์กินเนื้อ เนื่องจากการเชื่อมโยงที่ไม่ถูกต้องระหว่างกะโหลกและฟันของราอุยซูเคียนกับลำตัวของโปรซอโรโพดา ในสัตว์เช่น เทราโตซอรัส ) [ 49 ]เมื่อมีการบรรยายถึงไดโนเสาร์ดรอเมโอซอริเดตัวแรกที่รู้จัก ( Dromaeosaurus albertensis ) ในปี พ.ศ. 2465 [ 70 ] WD MatthewและBarnum Brownกลายเป็นนักบรรพชีวินวิทยาคนแรกที่แยกโปรซอโรพอดออกจากไดโนเสาร์กินเนื้อ และพยายามที่จะนำชื่อ "Goniopoda" กลับมาใช้สำหรับกลุ่มนั้น แต่นักวิทยาศาสตร์คนอื่นๆ ไม่ยอมรับข้อเสนอทั้งสองนี้[ 49 ]

ในปี 1956 คำว่า "Theropoda" กลับมาใช้อีกครั้งในฐานะกลุ่มสิ่งมีชีวิตที่ประกอบด้วยไดโนเสาร์กินเนื้อและลูกหลานของพวกมัน เมื่ออัลเฟรด โรเมอร์จัดจำแนกอันดับ Saurischia ใหม่เป็นสองอันดับย่อย คือ Theropoda และ Sauropoda การแบ่งพื้นฐานนี้ยังคงใช้มาจนถึงปัจจุบันในสาขาบรรพชีวินวิทยา ยกเว้น Prosauropoda ซึ่งโรเมอร์รวมไว้เป็นอันดับย่อยของ Theropoda โรเมอร์ยังคงรักษาการแบ่งระหว่าง Coelurosauria และCarnosauria (ซึ่งเขาก็จัดอันดับเป็นอันดับย่อยเช่นกัน) การแบ่งแยกนี้ถูกล้มล้างโดยการค้นพบDeinonychusและDeinocheirusในปี 1969 ซึ่งทั้งสองชนิดนี้ไม่สามารถจัดอยู่ในกลุ่ม "carnosaurs" หรือ "coelurosaurs" ได้อย่างง่ายดาย จากผลการค้นพบเหล่านี้และการค้นพบอื่นๆ ในช่วงปลายทศวรรษ 1970 Rinchen Barsboldได้สร้างกลุ่มย่อยของเทโรพอดชุดใหม่ ได้แก่ Coelurosauria, Deinonychosauria , Oviraptorosauria , Carnosauria , Ornithomimosauria และDeinocheirosauria [ 49 ]
ด้วยการเกิดขึ้นของการจัดกลุ่มตามวิวัฒนาการและการตั้งชื่อตามวิวัฒนาการในช่วงทศวรรษ 1980 และการพัฒนาในช่วงทศวรรษ 1990 และ 2000 ทำให้ภาพความสัมพันธ์ของเทโรพอดชัดเจนขึ้นJacques Gauthierได้ตั้งชื่อกลุ่มเทโรพอดหลักหลายกลุ่มในปี 1986 รวมถึงกลุ่มTetanuraeสำหรับสาขาหนึ่งของเทโรพอดพื้นฐานที่แยกตัวออกจากกลุ่มอื่นคือ Ceratosauria เมื่อมีข้อมูลเพิ่มเติมเกี่ยวกับความเชื่อมโยงระหว่างไดโนเสาร์และนกมากขึ้น เทโรพอดที่มีลักษณะคล้ายนกจึงถูกจัดกลุ่มไว้ในกลุ่มManiraptora (ซึ่ง Gauthier ตั้งชื่อไว้ในปี 1986 เช่นกัน[ 57 ] ) การพัฒนาใหม่เหล่านี้ยังมาพร้อมกับการยอมรับในหมู่นักวิทยาศาสตร์ส่วนใหญ่ว่านกเกิดขึ้นโดยตรงจากเทโรพอดกลุ่มมานิแรปทอแรน และเมื่อมีการยกเลิกการจัดลำดับชั้นในการจำแนกประเภทคลัดิสติก ก็มีการประเมินนกใหม่ในฐานะกลุ่มย่อยของไดโนเสาร์เทโรพอดที่รอดชีวิตจากการสูญพันธุ์ในยุคมีโซโซอิกและมีชีวิตอยู่มาจนถึงปัจจุบัน[ 49 ]
กลุ่มหลัก





ต่อไปนี้เป็นการจำแนกกลุ่มเทโรพอดแบบง่ายโดยอิงตามความสัมพันธ์ทางวิวัฒนาการ และจัดเรียงตามรายการสายพันธุ์ไดโนเสาร์ยุคมีโซโซอิกที่จัดทำโดย Holtz [ 1 ]สามารถดูเวอร์ชันที่มีรายละเอียดมากขึ้นได้ที่การจำแนกไดโนเสาร์เครื่องหมายกริช (†) ใช้เพื่อระบุกลุ่มที่ไม่มีสมาชิกที่ยังมีชีวิตอยู่
- † โคเอโลฟิโซอิเดีย (เทโรพอดนักล่าขนาดเล็กยุคแรก; รวมถึงโคเอโลฟิซิสและญาติใกล้เคียง)
- † เซราโตซอเรีย (มักมีเขาที่ซับซ้อน เป็นสัตว์กินเนื้อที่โดดเด่นทางตอนใต้ของยุคครีเทเชียสตอนปลาย ประกอบด้วยเซราโตซอรัสอะเบลิซอริเด โน อาซอริเดและญาติๆ ของพวกมัน)
- Tetanurae ("หางแข็ง"; รวมถึงเทอโรพอดส่วนใหญ่)
- † เมกาโลซอโรอิเดีย (กลุ่มแรกของสัตว์กินเนื้อขนาดใหญ่ ซึ่งรวมถึงเมกาโลซอริเดและสไปโนซอริเดที่อาศัย อยู่กึ่งน้ำกึ่งบก )
- † อัลโลซอโรอิเดีย (กลุ่มสัตว์กินเนื้อขนาดใหญ่บนบกที่ประสบความสำเร็จอย่างสูง ครองโลกในช่วงปลายยุคจูราสสิก ต้นยุคครีเทเชียส และต้นยุคครีเทเชียสตอนปลาย; รวมถึงอัลโลซอรัสและญาติๆ เช่นคาร์คาโรดอนโตซอริเดีย )
- † เมการัปทอรา (กลุ่มไดโนเสาร์เทอโรพอดนักล่าขนาดกลางถึงใหญ่ มีแขนยาวและกรงเล็บขนาดใหญ่ ซึ่งยังไม่ทราบความสัมพันธ์ที่แน่ชัด เป็นสัตว์กินเนื้อที่ครองอำนาจในซีกโลกใต้ในช่วงปลายยุคครีเทเชียส)
- โคเอลูโรซอเรีย (ไดโนเสาร์เทอโรพอดมีขน มีขนาดตัวและถิ่นที่อยู่หลากหลาย)
- † คอมป์โซกนาธิเด (โคเอลูโรซอร์ยุคแรกที่มีแขนขาหน้าลดขนาดลง)
- † ไทแรนโนซอโรอิเดีย ( ไดโนเสาร์วงศ์ไทแรน โนซอรัส ซึ่งเป็นสัตว์กินเนื้อที่ครองอำนาจในแถบซีกโลกเหนือเป็นส่วนใหญ่ในช่วงปลายยุคครีเทเชียส และญาติใกล้ชิด มักมีแขนขาหน้าลดขนาดลง)
- † ออร์นิโทมิโมซอเรีย (" สัตว์เลียน แบบนกกระจอกเทศ "; ส่วนใหญ่ไม่มีฟัน; กินได้ทั้งพืชและสัตว์ หรืออาจกินพืชเป็นอาหาร)
- มานิรัปทอรา ("ผู้ฉกฉวยมือ"; มีแขนและนิ้วเรียวยาว)
- † อัลวาเรซซอโรอิเดีย (สัตว์กินแมลงขนาดเล็กที่มีแขนหน้าลดขนาดลง โดยแต่ละแขนมีกรงเล็บขนาดใหญ่หนึ่งอัน)
- † เทริซิโนซอเรีย (สัตว์กินพืชสองขาที่มีกรงเล็บมือขนาดใหญ่และหัวเล็ก)
- † สแกนโซริโอปเทอริกิเด (แมนิแรปเตอร์ขนาดเล็ก อาศัยอยู่บนต้นไม้ มีนิ้วที่สามยาว)
- † โอวิแรปเตอร์ซอเรีย (ส่วนใหญ่ไม่มีฟัน อาหารและวิถีชีวิตยังไม่เป็นที่แน่ชัด)
- พาราเวส ("นกใกล้เคียง"; โดยทั่วไปกินเนื้อหรือกินทั้งพืชและสัตว์ มีกรงเล็บรูปเคียว)
- † Anchiornithidae (นกดึกดำบรรพ์ขนาดเล็กมีปีก)
- † ดรอเมโอซอริเด (ไดโนเสาร์เทอโรพอดนักล่าขนาดเล็กถึงขนาดกลาง)
- † วงศ์ Troodontidae (ไดโนเสาร์เทอโรพอดขนาดเล็ก รูปร่างเพรียวบาง กินเนื้อเป็นอาหารหรือกินทั้งพืชและสัตว์)
- Avialae (นกและญาติที่สูญพันธุ์ไปแล้ว)
- † Omnivoropterygidae (นกขนาดใหญ่ หางสั้น ยุคแรก)
- † Confuciusornithidae (นกขนาดเล็กไม่มีฟัน)
- † เอนันติออร์นิเธส (นกโบราณที่อาศัยอยู่บนต้นไม้และบินได้)
- นก ยูออร์นิเทส (นกบินขั้นสูง)
- † Yanornithiformes (นกฟันยุคครีเทเชียสของจีน)
- † เฮสเปอโรนนิเทส (นกน้ำที่เชี่ยวชาญด้านการดำน้ำ)
- นก (นกปากจงอยในปัจจุบันและญาติที่สูญพันธุ์ไปแล้ว)
ความสัมพันธ์
แผนผังวงศ์ตระกูลต่อไปนี้แสดงให้เห็นถึงการสังเคราะห์ความสัมพันธ์ของกลุ่มเทโรพอดหลักโดยอิงจากการศึกษาต่างๆ ที่ดำเนินการในช่วงทศวรรษ 2010 [ 72 ]
| เทโรโปดา | |


Averostra ได้รับการตั้งชื่อโดยGS Paul ในปี 2002 โดย กำหนดเป็นกลุ่มตามลักษณะเฉพาะ (apomorphy-based clade) ซึ่งหมายถึงกลุ่มที่รวมถึง DromaeosauridaeและAvepoda อื่นๆ ที่มี (บรรพบุรุษที่มี) ช่องเปิด promaxillary fenestra ( fenestra promaxillaris ) ซึ่งอาจเรียกได้ว่าเป็นช่องเปิด maxillary fenestra [ 73 ]ซึ่งเป็นช่องเปิดพิเศษที่ด้านหน้าด้านนอกของ กระดูกขา กรรไกรบน[ 74 ]ต่อมาได้รับการกำหนดนิยามใหม่โดยMartin EzcurraและGilles Cunyในปี 2007 โดยกำหนดเป็นกลุ่มตามจุดเชื่อมต่อ (node-based clade) ซึ่งประกอบด้วยCeratosaurus nasicornis , Allosaurus fragilisบรรพบุรุษร่วมสุดท้ายของพวกมันและลูกหลานทั้งหมด[ 75 ] Mickey Mortimer แสดงความคิดเห็นว่าคำจำกัดความดั้งเดิมของ Paul ที่อิงตามลักษณะเฉพาะอาจทำให้ Averostra เป็นกลุ่มที่กว้างกว่า จุดเชื่อมต่อ Ceratosaurus + Allosaurus มาก ซึ่งอาจรวมถึง Avepoda ทั้งหมดหรือมากกว่านั้น[ 76 ]
การศึกษาขนาดใหญ่เกี่ยวกับไดโนเสาร์ยุคแรกโดย ดร. Matthew G. Baron, David Normanและ Paul M. Barrett (2017) ที่ตีพิมพ์ในวารสารNatureชี้ให้เห็นว่า Theropoda มีความสัมพันธ์ใกล้ชิดกับOrnithischia มากกว่า โดยเป็นกลุ่มพี่น้องภายในกลุ่มOrnithoscelidaสมมติฐานใหม่นี้ยังพบว่าHerrerasauridaeเป็นกลุ่มพี่น้องกับSauropodomorphaในSaurischia ที่ได้รับการกำหนดนิยามใหม่ และชี้ให้เห็นว่าลักษณะทางกายภาพของสัตว์กินเนื้ออย่างสุดขั้วที่พบในตัวอย่างของ theropods และ herrerasaurids นั้นได้มา จากการวิวัฒนาการ แบบลู่เข้า [ 77 ] [ 78 ] อย่างไรก็ตาม แผนภูมิ วิวัฒนาการนี้ยังคงเป็นที่ถกเถียงกันอยู่ และกำลังมีการดำเนินการเพิ่มเติมเพื่อชี้แจงความสัมพันธ์เหล่านี้[ 79 ]
เชิงอรรถ
- ^ อย่างไรก็ตาม บางสกุลใน Avetheropodaมีสี่นิ้ว ดู ""Theropoda I" เกี่ยวกับAvetheropoda ภาควิชาธรณีวิทยามหาวิทยาลัยแมริแลนด์ 14 กรกฎาคม 2549
ดูเพิ่มเติม
- ไดโนเสาร์มีขน
- ที่มาของนก
- กลุ่มสายพันธุ์หลักอื่นๆ ของไดโนเสาร์
ลิงก์ภายนอก
 สื่อที่เกี่ยวข้องกับเทโรโปดาในวิกิมีเดียคอมมอนส์
สื่อที่เกี่ยวข้องกับเทโรโปดาในวิกิมีเดียคอมมอนส์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เทโรโปดา
Theropoda ( / θ ɪ ˈ r ɒ p ə d ə / ; จากภาษากรีกโบราณθηρίο- ποδός ) เป็นหนึ่งในสามกลุ่ม หลัก ของไดโนเสาร์ร่วมกับOrnithischiaและSauropodomorpha Theropods...
ลักษณะเฉพาะ
มีการเสนอ synapomorphies ต่างๆสำหรับ Theropoda โดยอิงจาก taxa ที่รวมอยู่ในกลุ่ม ตัวอย่างเช่น บทความปี 1999 โดย Paul Sereno แนะนำว่า theropods มีลักษณะเฉพาะ เช่น ร่อง ectopterygoid (ร่องรอบกระดูก ectopterygoid) ข้อต่อ intramandibular ที่อยู่ใน ขากรรไกรล่าง และ...
อาหารและฟัน
เทอโรพอดมีอาหารหลากหลาย ตั้งแต่กินแมลง กินพืช ไปจนถึงกินเนื้อ การกินเนื้ออย่างเดียวถือเป็นอาหารดั้งเดิมของเทอโรพอดมาโดยตลอด และในอดีต ความหลากหลายของอาหารถือเป็นลักษณะเฉพาะของเทอโรพอดที่เป็นนกเท่านั้น อย่างไรก็ตาม การค้นพบในช่วงปลายศตวรรษที่ 20...
เปลือกหุ้มร่างกาย (ผิวหนัง เกล็ด และขน)
เทโรพอ ดในยุคมี โซโซอิก มีความหลากหลายมากในแง่ของลักษณะพื้นผิวและการปกคลุมของผิวหนัง ขนหรือโครงสร้างคล้ายขน (เส้นใย) พบได้ในสายพันธุ์ส่วนใหญ่ของ โคเอลูโรซอร์ (ดู ไดโนเสาร์มีขน ) อย่างไรก็ตาม นอกเหนือจากโคเอลูโรซอร์แล้ว...