อ่าน 15 นาที
ครีโอดอนต้า
Creodonta ("ฟันเนื้อ") เป็นอันดับ ของ สัตว์ เลี้ยงลูกด้วยนมกิน เนื้อที่ สูญพันธุ์ไปแล้ว ซึ่งมีชีวิตอยู่ตั้งแต่ ยุค Paleocene ตอนต้น (หรือ Cretaceous ตอนปลาย ) จนถึง ยุค Miocene...
ครีโอดอนต้า
| ครีโอดอนต้า ช่วงเวลา: ต้นพาลีโอซีนถึงปลายไมโอซีน (คาดว่าHyaenodontaมีต้นกำเนิดในช่วงปลายครีเทเชียสแต่ยังไม่ได้รับการยืนยันจากฟอสซิล) [ 1 ] [ 2 ] | |
|---|---|
 | |
| ฟอสซิลครีโอดอนต์หลากหลายชนิดจากยุคอีโอซีนรัฐโคโลราโดสหรัฐอเมริกาจากบนลงล่าง: ไทรเทมโนดอน , แพทริโอเฟลิส , มาเคอรอยด์และซิโนปา | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | คอร์ดาต้า |
| ระดับ: | สัตว์เลี้ยงลูกด้วยนม |
| อินฟราคลาส: | รก |
| มิเรอร์เดอร์: | เฟราเอ |
| คำสั่ง: | † Creodonta Cope , 1875 [ 3 ] |
| ครอบครัว | |
| คำพ้องความหมาย | |
รายชื่อคำพ้องความหมาย: | |
Creodonta ("ฟันเนื้อ") เป็นอันดับของสัตว์เลี้ยงลูกด้วยนมกินเนื้อที่สูญพันธุ์ไปแล้วซึ่งมีชีวิตอยู่ตั้งแต่ยุค Paleocene ตอนต้น (หรือCretaceous ตอนปลาย ) จนถึง ยุค Miocene ตอนปลายในทวีปอเมริกาเหนือยูเรเซียและแอฟริกาเดิมทีคิดว่าเป็นกลุ่มสัตว์กลุ่มเดียวที่เป็นบรรพบุรุษของสัตว์กินเนื้อ ในปัจจุบัน แต่ปัจจุบันอันดับนี้มักถูกพิจารณาว่าเป็น กลุ่มที่มี บรรพบุรุษร่วมกันหลายสายจากสองกลุ่มที่แตกต่างกัน คือOxyaenidsและHyaenodontsไม่ใช่กลุ่มตามธรรมชาติ Oxyaenids เป็นที่รู้จักครั้งแรกจากยุค Paleocene ตอนต้นของทวีปอเมริกาเหนือ[ 12 ]ในขณะที่ Hyaenodonts มาจากยุค Paleocene ตอนปลาย หรือ Cretaceous ตอนปลายของทวีปยุโรป[ 13 ] [ 14 ] [ 15 ]
ครีโอโดนต์เป็นสัตว์เลี้ยงลูกด้วยนมกินเนื้อที่เด่นที่สุดในช่วง55 ถึง 35ล้านปีก่อนโดยมีความหลากหลายและแพร่หลายสูงสุดในช่วงยุคอีโอซีน [ 16 ] สัตว์เลี้ยงลูกด้วยนมกินเนื้อขนาดใหญ่กลุ่มแรกปรากฏขึ้นพร้อมกับการแพร่กระจายของออกซีเอนิดในช่วงปลายยุคพาลีโอซีน[ 17 ]ในช่วง ยุค พาลีโอจีนสัตว์ในกลุ่ม "ครีโอโดนต์" เป็นสัตว์กินเนื้อบนบกที่อุดมสมบูรณ์ที่สุดในโลกเก่า[ 18 ]ใน แอฟริกาใน ยุคโอลิโกซีนไฮยาโนดอนต์เป็นกลุ่มสัตว์กินเนื้อขนาดใหญ่ที่เด่นที่สุด และดำรงอยู่จนถึงกลางยุคไมโอซีน
กลุ่ม "ครีโอดอนต์" มีช่วงการกระจายตัวที่กว้างขวางทั้งทางภูมิศาสตร์และช่วงเวลา พวกมันเป็นที่รู้จักตั้งแต่ปลายยุคพาลีโอซีนจนถึงปลายยุคโอลิโกซีนในอเมริกาเหนือต้นยุคอีโอซีนจนถึงปลายยุคโอลิโกซีนในยุโรปตั้งแต่ปลายยุคพาลีโอซีนจนถึงปลายยุคไมโอซีนในเอเชียและตั้งแต่ปลายยุคพาลีโอซีนจนถึงปลายยุคไมโอซีนในแอฟริกา[ 19 ] ในขณะที่ส่วนใหญ่เป็นสัตว์เลี้ยงลูกด้วยนมขนาดเล็กถึงขนาดกลาง แต่ในจำนวนนั้นมีMegistotheriumและSarkastodonซึ่งทั้งสองชนิดเป็นสัตว์กินเนื้อขนาดใหญ่ที่สุดบนบก[ 20 ] [ 21 ]สกุลสุดท้ายDissopsalisสูญพันธุ์ไปเมื่อประมาณ10 ล้านปีก่อน[ 22 ]
นักบรรพชีวินวิทยาสมัยใหม่ส่วนใหญ่เห็นพ้องกันว่าทั้งสองตระกูล "ครีโอโดนต์" เกี่ยวข้องกับสัตว์กินเนื้อแต่ไม่ใช่บรรพบุรุษโดยตรง ยังไม่ชัดเจนว่าทั้งสองตระกูลมีความสัมพันธ์ใกล้ชิดกันมากน้อยเพียงใด โดยทั่วไป การจำแนกประเภทมีความซับซ้อนเนื่องจากความสัมพันธ์ระหว่างสัตว์เลี้ยงลูกด้วยนมฟอสซิลมักตัดสินจากความคล้ายคลึงกันของฟัน แต่ฟันของ สัตว์ กินเนื้ออาจวิวัฒนาการเป็นรูปทรงที่คล้ายกันผ่านวิวัฒนาการแบบลู่เข้าเพื่อรับมือกับกลไกการกินเนื้อ[ 23 ]
สัตว์ในอันดับ "ครีโอดอนต์" มีลักษณะร่วมกับสัตว์กินเนื้อ (Carnivora) และสัตว์เลี้ยงลูกด้วยนมกินเนื้อกลุ่มอื่นๆ อีกหลายกลุ่ม คือฟันกรามบนและล่างที่ดัดแปลงให้มีลักษณะคล้ายกรรไกร ใช้สำหรับตัดเนื้อเยื่อกล้ามเนื้อ การปรับตัวนี้ยังพบได้ในสัตว์เลี้ยงลูกด้วยนมกินเนื้อกลุ่มอื่นๆ ด้วย
อนุกรมวิธานและประวัติศาสตร์
คำว่า "Creodonta" ถูกบัญญัติโดยEdward Drinker Copeในปี 1875 [ 3 ] Cope รวมoxyaenidsและviverravid Didymictisแต่ละเว้นHyaenodontidaeในปี 1880 เขาขยายคำนี้ให้รวมถึงวงศ์Miacidae (รวมถึง Viverravidae), Arctocyonidae , Leptictidae (ปัจจุบันคือ Pseudorhyncocyonidae ), Oxyaenidae , AmbloctonidaeและMesonychidae [ 24 ] เดิมที Cope จัดให้ creodonts อยู่ในInsectivora อย่างไรก็ตาม ในปี 1884 เขาถือว่าพวกมันเป็นกลุ่ม พื้นฐานที่ทั้ง carnivorans และ insectivorans ถือกำเนิดขึ้น[ 25 ]
Hyaenodontidae ไม่ได้ถูกรวมอยู่ในกลุ่มครีโอโดนต์จนกระทั่งปี พ.ศ. 2452 [ 26 ] William Diller Matthewถือว่า Creodonta เป็นอันดับย่อยของอันดับCarnivoraซึ่งแบ่งออกเป็นสามกลุ่ม:
- "ครีโอดอนตาที่ไม่ปรับตัว" (Creodonta inadaptiva) กลุ่มที่รวมถึง "ซูโดครีโอดี" (ออกซิเอนิดและไฮยาโนดอนทิด) และ เมโซ นิคิด
- "Adaptive Creodonta" (Creodonta adaptiva) ประกอบด้วยไมแอซิดและแท็กซ่าที่รวมอยู่ในตะกร้าขยะ " Arctocyonidae "
- และ "ดั้งเดิม Creodonta" (Creodonta primitiva) ประกอบด้วย Oxyclaenidae
เมื่อเวลาผ่านไป กลุ่มและสายพันธุ์ต่างๆ ถูกแยกออกจากอันดับนี้ อันดับนี้มีเสถียรภาพในช่วงกลางศตวรรษที่ 20 โดยประกอบด้วยออกซิเอนิด ไฮเอโนดอนต์ เมโซนิคิด และอาร์คโตไซโอนิด[ 27 ]ซึ่งเข้าใจกันว่าเป็นกลุ่มหลักของสัตว์เลี้ยงลูกด้วยนมที่มีรกที่กินเนื้อเป็นอาหารที่ไม่ใช่สมาชิกของกลุ่มสัตว์กินเนื้อ เริ่มชัดเจนมากขึ้นว่าอาร์คโตไซโอนิดเป็นกลุ่มที่รวมเอาสิ่งที่ไม่จำเป็นไว้ด้วยกันและเมโซนิคิดอาจมีความสัมพันธ์ใกล้ชิดกับสัตว์กีบ มากกว่า ในปี 1969 ครีโอโดนตาประกอบด้วยออกซิเอนิดและไฮเอโนดอนต์เท่านั้น[ 28 ]
เมื่อไม่นานมานี้ "Creodonta" ถูกพิจารณาว่าเป็นกลุ่มสัตว์เลี้ยงลูกด้วยนมกินเนื้อที่มีรก ซึ่งไม่มี บรรพบุรุษร่วม กัน (และไม่ใช่กลุ่มตามธรรมชาติ) และสมาชิกของ Creodonta เป็น กลุ่มพี่น้องกับ Carnivoramorpha (สัตว์กินเนื้อและบรรพบุรุษ) ภายในกลุ่มPan-Carnivora (ในอันดับย่อยFerae ) โดยแบ่งออกเป็นสองกลุ่ม: อันดับOxyaenodontaเป็นกลุ่มหนึ่ง และอันดับHyaenodontaบวกกับบรรพบุรุษ (สกุลAltacreodusและSimidectes ) อยู่ในอีกกลุ่มหนึ่ง[ 29 ] [ 30 ] [ 31 ] [ 32 ] [ 33 ]อย่างไรก็ตาม การวิเคราะห์ทางวิวัฒนาการบางอย่างได้จัดให้พวกมันเป็นกลุ่มตามธรรมชาติ เช่น การวิเคราะห์ทางวิวัฒนาการของสัตว์เลี้ยงลูกด้วยนมในยุค Paleocene ที่ตีพิมพ์ในปี 2015 ซึ่งสนับสนุนความเป็นกลุ่มเดียวของ Creodonta และจัดให้กลุ่มนี้เป็นญาติของกลุ่มPholidotamorpha ( ตัวนิ่มและบรรพบุรุษ) [ 34 ]
พอลลี่ได้โต้แย้งว่าลักษณะร่วม ที่มีอยู่เพียงอย่างเดียว ระหว่างออกซิยาอินิดส์และไฮยาโนดอนทิดส์คือใบมีดเมตาสไตลาร์ขนาดใหญ่บนฟันกรามซี่แรก (M1) แต่เขาเชื่อว่าลักษณะดังกล่าวเป็นลักษณะทั่วไปของยูเทอเรียพื้นฐานทั้งหมด[ 35 ]การแยกออกซิยาอินิดส์ออกจากไฮยาโนดอนทิดส์จะสอดคล้องกับหลักฐานทางชีวภูมิศาสตร์ด้วย เนื่องจากออกซิยาอินิดส์ตัวแรกเป็นที่รู้จักจากยุคพาลีโอซีนตอนต้นของอเมริกาเหนือ และไฮยาโนดอนทิดส์ตัวแรกมาจากยุคพาลีโอซีนตอนปลายมากของแอฟริกาเหนือ[ 17 ]
สิ่ง ที่ทำให้การจัดเรียงนี้ซับซ้อนขึ้นคือการรับรองเบื้องต้นของ Gunnell [ 19 ]ในการสร้างวงศ์ที่สามLimnocyonidae [ 36 ] กลุ่มนี้รวมถึงแท็ก ซาที่เคยถูกพิจารณาว่าเป็น oxyaenids เช่นLimnocyon , Thinocyon [ 26 ]และProlimnocyon [ 37 ] Wortman ถึงกับสร้างวงศ์ย่อย Limnocyoninae ภายใน oxyaenids [ 38 ] Van Valen จัดวงศ์ย่อยเดียวกัน (รวมถึงOxyaenodon ) ไว้ใน Hyaenodontidae [ 28 ] Gunnell ไม่แน่ใจว่า Limnocyonidae เป็นกลุ่มภายใน Hyaenodontidae (แม้จะเป็นกลุ่มพี่น้องกับ hyaenodontids ที่เหลือ) หรือแยกออกไปโดยสิ้นเชิง[ 19 ]
ตามที่ Gunnell กล่าว คุณลักษณะเด่นของ oxyaenids ได้แก่: กะโหลกสมองขนาดเล็กที่อยู่ต่ำในกะโหลกศีรษะ กระดูกท้ายทอยกว้างที่ฐานและแคบลงทางด้านบน (เพื่อให้มีรูปร่างเป็นสามเหลี่ยม) กระดูกน้ำตาเป็นส่วนขยายครึ่งวงกลมบนใบหน้า ขากรรไกรล่างมีส่วนเชื่อมต่อที่แข็งแรง ฟันกรามM1และ M2 ประกอบเป็นฟันกรามเนื้อ ในขณะที่ฟันกราม M3/m3 ไม่มีมือและเท้าอยู่ในท่าราบหรือกึ่งราบ กระดูกน่องเชื่อมต่อกับกระดูกส้นเท้าและกระดูกข้อเท้าเชื่อมต่อกับกระดูกลูกบาศก์ กระดูก นิ้วมือและ นิ้วเท้าถูกบีบอัดและมีรอยแตกที่ปลาย[ 19 ]
ในทำนองเดียวกัน รายการลักษณะเด่นของไฮยาโนดอนทิดที่ Gunnell ระบุไว้ ได้แก่ กะโหลกยาวและแคบ มีฐานกะโหลก แคบ และท้ายทอยสูงแคบกระดูกหน้าผากเว้าระหว่างบริเวณเบ้าตา ฟันกราม M2 และ M3 ประกอบเป็นฟันกรามใหญ่ M3 มีอยู่ในสายพันธุ์ส่วนใหญ่ ในขณะที่ M3 มีอยู่เสมอ มือและเท้ามีตั้งแต่แบบวางฝ่าเท้าไปจนถึงแบบวางนิ้ว กระดูกน่องเชื่อมต่อกับกระดูกส้นเท้า ในขณะที่ข้อต่อระหว่างกระดูกข้อเท้าและกระดูกลูกบาศก์ลดลงหรือไม่มีเลย กระดูกนิ้วเท้าส่วนปลายถูกบีบอัดและมีรอยแตกที่ปลาย[ 19 ]
ตามข้อมูลของ Gunnell สัตว์ในกลุ่ม Limnocyonids มีลักษณะดังต่อไปนี้: M3/m3 ลดลงหรือไม่มีอยู่ ฟันอื่นๆ ไม่ลดลงจะงอยปากยาว สัตว์เหล่านี้มีขนาดเล็กถึงปานกลาง[ 19 ]
สัณฐานวิทยา
ฟัน
ในกลุ่มครีโอดอนท์ดั้งเดิมสูตรทางทันตกรรมคือ3.1.4.33.1.4.3แต่รูปแบบในภายหลังมักมีจำนวนฟันตัด ฟันกรามน้อย และ/หรือฟันกรามลดลง[ 39 ]ฟันเขี้ยวมีขนาดใหญ่และแหลมเสมอ ฟันตัดข้างมีขนาดใหญ่ ในขณะที่ฟันตัดตรงกลางมักมีขนาดเล็ก[ 19 ]ฟันกรามน้อยมีลักษณะดั้งเดิม โดยมีปุ่มหลักหนึ่งปุ่มและปุ่มรองต่างๆ[ 18 ]
ครีโอดอนต์มี ฟันตัดเนื้อสองหรือสามคู่โดยคู่หนึ่งทำหน้าที่ตัดที่ใหญ่ที่สุด (ไม่ว่าจะเป็น M1/m2 หรือ M2/m3) [ 19 ]การจัดเรียงนี้แตกต่างจากสัตว์กินเนื้อในปัจจุบัน ซึ่งใช้ P4 และ m1 เป็นฟันตัดเนื้อ[ 40 ]ความแตกต่างนี้ชี้ให้เห็นถึงวิวัฒนาการแบบลู่เข้าในกลุ่มสัตว์กินเนื้อ โดยมีประวัติวิวัฒนาการที่แยกจากกันและการแบ่งระดับอันดับ[ 41 ]เนื่องจากฟันที่แตกต่างกันได้วิวัฒนาการเป็นฟันตัดเนื้อทั้งระหว่างครีโอดอนต์และสัตว์กินเนื้อ และระหว่าง ออก ซิเอนิดส์และไฮยาเอโนดอนต์ ฟันตัดเนื้อยังพบได้ในกลุ่มสัตว์เลี้ยงลูกด้วยนมกินเนื้ออื่นๆ เช่นค้างคาว ที่สูญพันธุ์ไปแล้วอย่าง เนโครแมนติสรวมถึงกลุ่มสิ่งมีชีวิตที่ไม่เกี่ยวข้องกันอย่างมาก เช่นไทลาโคเลโอ สัตว์มีถุง หน้าท้องกินเนื้อ
ฟันกรามที่แตกต่างกันมีส่วนเกี่ยวข้องในสองกลุ่มหลักของครีโอโดนต์ ใน Oxyaenidae ฟันกราม M1 และ m2 เป็นฟันตัดเนื้อ ในกลุ่มไฮยาโนดอนทิด ฟันกราม M2 และ m3 เป็นฟันตัดเนื้อ แตกต่างจากสัตว์กินเนื้อส่วนใหญ่ในปัจจุบันที่ฟันตัดเนื้อเป็นฟันตัดเพียงอย่างเดียว ฟันกรามครีโอโดนต์อื่นๆ มีหน้าที่ตัดเป็นรอง[ 26 ]ความแตกต่างในฟันที่เป็นฟันตัดเนื้อเป็นข้อโต้แย้งสำคัญสำหรับความเป็นพหุวงศ์ของครีโอโดนต์



กะโหลก
ครีโอโดนต์มีกะโหลกยาวและแคบ มีสมองขนาดเล็ก กะโหลกจะแคบลงอย่างเห็นได้ชัดด้านหลังดวงตา ทำให้เกิดส่วนกะโหลกที่เรียกว่าสปลานโนคราเนียมและนิวโรคราเนียม อย่างชัดเจน พวกเขามี สันกระดูกซาจิตัล ขนาดใหญ่ และกระดูกมาสตอยด์ ที่กว้าง (ซึ่งอาจเป็นลักษณะเฉพาะของกลุ่ม) [ 19 ] ครีโอ โดนต์หลายตัวมีหัวขนาดใหญ่เมื่อเทียบกับสัดส่วน[ 18 ]ในรูปแบบพื้นฐาน กระดูกหูจะไม่แข็งตัว โดยทั่วไปโพรงขมับจะกว้างมาก[ 19 ]


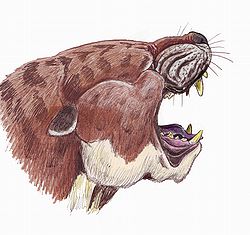



โครงกระดูกส่วนลำตัว
ครีโอดอนต์มีโครงกระดูกส่วนหลังกะโหลกแบบทั่วไป แขนขาของพวกมันเป็นแบบเมซาโซนิก (โดยมีแกนของเท้าอยู่ที่กลางนิ้วทั้งห้า) วิธีการเคลื่อนที่ของพวกมันมีตั้งแต่แบบเดินด้วยฝ่าเท้าไปจนถึงแบบเดินด้วยนิ้วปลายนิ้วเป็นกรงเล็บที่เชื่อมติดกัน[ 19 ]
ภายในกลุ่มออกซีเอนิดส์ กลุ่มอนุกรมวิธานจำนวนมากในวงศ์นี้ยังคงเป็นการปรับตัว ในทางกลับกัน ไฮยาโนดอนต์แสดงให้เห็นถึงการเปลี่ยนผ่านจากแบบเดินด้วยฝ่าเท้าไปเป็นแบบเดินด้วยนิ้วเท้าตลอดประวัติศาสตร์วิวัฒนาการของพวกมัน[ 42 ] [ 43 ]

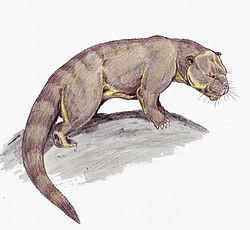

ขนาด

ครีโอโดนต์มีขนาดตั้งแต่เท่าแมวตัวเล็กไปจนถึงซาร์คาสโตดอนและไฮยาไนลูรีนขนาด ใหญ่ [ 20 ]ขนาดของมันถูกอธิบายว่าใหญ่กว่าแพทริโอเฟลิส ถึง 50% ซึ่งมันมีลักษณะคล้ายคลึงกันหลายอย่าง[ 44 ]การศึกษาหนึ่งประเมินว่ามันอาจมีน้ำหนักถึง 800 กิโลกรัม (1,800 ปอนด์) แม้ว่าอาจเป็นการประเมินที่สูงเกินไปก็ตาม[ 20 ]ไฮยาไนลูรีนขนาดใหญ่ได้แก่เมกิสโตเทอเรียม ไฮยาไนลูรอสและซิมบาคูบวา [ 45 ] [ 46 ] [ 20 ] [ 21 ] ครีโอโดนต์ที่ใหญ่ที่สุดในอเมริกาเหนือคือเฮมิปซาโลดอน แกรนดิสซึ่งอาจมีน้ำหนัก 430–760 กิโลกรัม (950–1,680 ปอนด์) [ 47 ]
อย่างไรก็ตาม สัตว์ในกลุ่มออกซิเอนิดขนาดใหญ่ยังไม่เป็นที่รู้จักจนกระทั่งช่วงปลายยุคพาลีโอซีน ซึ่งมีการแพร่กระจายของออกซิเอนิด[ 17 ]เช่นDipsalidictis ที่มีขนาดเท่าเสือพูมา และ Dipsalodon ที่น่าจะเป็น สัตว์กินซากที่บดกระดูก[ 48 ]ในทางกลับกัน สัตว์ในกลุ่มไฮยาโนดอนต์ที่ใหญ่ที่สุดเท่าที่รู้จักไม่ได้ปรากฏขึ้นจนกระทั่งช่วงต้นยุคไมโอซีน[ 45 ]
ครีโอโดนต์บางชนิด ( Arfia , ProlimnocyonและPalaeonictis ) ดูเหมือนจะประสบกับปรากฏการณ์แคระแกร็นในช่วงPaleocene-Eocene Thermal Maximumที่พบในสกุลสัตว์เลี้ยงลูกด้วยนมอื่นๆ คำอธิบายที่เสนอสำหรับปรากฏการณ์นี้คือ ระดับคาร์บอนไดออกไซด์ที่เพิ่มขึ้นในบรรยากาศส่งผลกระทบโดยตรงต่อสัตว์กินเนื้อผ่านอุณหภูมิและความแห้งแล้งที่เพิ่มขึ้น และยังส่งผลกระทบทางอ้อมต่อพวกมันด้วยการลดขนาดของเหยื่อที่เป็นสัตว์กินพืชผ่านแรงกดดันการคัดเลือกแบบเดียวกัน[ 49 ]
ชีววิทยา
อาหารและการให้อาหาร
ครีโอดอนต์ยุคแรก (ทั้งออกซีเอนิดและไฮยาเอโนดอนทิด) แสดงให้เห็นฟันกรามแบบไตรโบสเฟนิกซึ่งพบได้ทั่วไปในเทอเรียน ยุคแรก รูปแบบขนาดเล็กมีสันหลังเมตาคริสตา-เมตาสเตลลาร์ที่ค่อนข้างแข็งแรง[ 50 ]ซึ่งบ่งชี้ว่าพวกมันน่าจะเป็นสัตว์กินอาหารแบบฉวยโอกาส โดยกินสิ่งต่างๆ เช่น ไข่ นก สัตว์เลี้ยงลูกด้วยนมขนาดเล็ก แมลง และอาจรวมถึงพืชด้วย[ 19 ]ซึ่งอาจคล้ายกับวิเวอร์ริดใน ปัจจุบัน [ 39 ]รูปแบบขนาดใหญ่มีกำลังในการเฉือนที่มากกว่า และกำลังนี้เพิ่มขึ้นเมื่อเวลาผ่านไปอาร์เฟียซึ่งเป็นสัตว์เลี้ยงลูกด้วยนมกินเนื้อที่พบได้ทั่วไปในอเมริกาเหนือยุคอีโอซีนตอนต้น ได้พัฒนาไตรโกนิดที่เปิดกว้างมากขึ้นบน M3 ตลอดช่วงยุคอีโอซีนตอนต้น ทำให้ความสามารถในการเฉือนของฟันกรามเพิ่มขึ้น[ 50 ]การพัฒนาที่คล้ายกันนี้สามารถเห็นได้จากการเปรียบเทียบออกซีเอนาโปรโตโตมัสและลิมโนไซออ น กับสัตว์กินอาหารทั่วไปที่มีขนาดเล็กกว่าในกลุ่มครีโอดอนต์[ 19 ]ที่น่าสนใจคือ ออกซิเอนิดส์กลุ่มสุดท้ายบางกลุ่ม เช่นPatriofelisและMachaeroidinaeแสดงให้เห็นถึงการปรับตัวไปสู่การกินเนื้อสัตว์ เป็นหลัก [ 51 ] [ 52 ]ในทำนองเดียวกัน ไฮยาโนดอนต์กลุ่มที่อายุน้อยที่สุดในยุคไมโอซีนบางกลุ่มก็แสดงให้เห็นถึงความเชี่ยวชาญอย่างมากในการกินเนื้อสัตว์เป็นหลัก[ 53 ] [ 54 ]
การสูญพันธุ์
โดยรวมแล้ว Creodonts เริ่มลดจำนวนลงในช่วงยุคอีโอซีน [ 55 ]โดยOxyaenidsสูญพันธุ์ไปในช่วงกลางยุคอีโอซีน[ 56 ] [ 57 ] ผู้เชี่ยวชาญบางคนเสนอว่าการสูญพันธุ์ของพวกมันเกิดจากการแข่งขันกับNimravids [ 57 ]อย่างไรก็ตาม ผู้เชี่ยวชาญคนอื่นๆ ไม่เห็นด้วยกับสมมติฐานนี้[ 58 ] [ 56 ]บันทึกฟอสซิลที่ยอดเยี่ยมในอเมริกาเหนือแสดงให้เห็นว่า Oxyaenids ลดจำนวนลงก่อนการปรากฏตัวของกลุ่มสิ่งมีชีวิตทดแทน เช่น Nimravids [ 59 ] นอกจากนี้ บันทึกสุดท้ายของ Machaeroidines มีอายุเก่ากว่าบันทึกแรกสุดของ Nimravids ในอเมริกาเหนือ[ 57 ] [ 60 ]ในทางกลับกัน การเปลี่ยนแปลงสภาพภูมิอากาศในช่วงปลายยุคอีโอซีนมีบทบาทในการสูญพันธุ์ของพวกมัน เนื่องจากในช่วงปลายยุคอีโอซีน สภาพภูมิอากาศของโลกเริ่มเย็นลง ส่งผลให้สภาพแวดล้อมแห้งแล้งและเปิดโล่งมากขึ้น[ 59 ] [ 56 ]เนื่องจากการเคลื่อนที่ต่ำ ออกซิเอนิดจึงสูญพันธุ์เพราะไม่สามารถปรับตัวให้เข้ากับถิ่นที่อยู่อาศัยในป่าโปร่งเขตอบอุ่นได้[ 56 ]
ไฮยาโนดอนต์ซึ่งเป็นกลุ่มสุดท้ายของครีโอดอนต์ ก็ประสบกับภาวะลดจำนวนลงในยูเรเซียและอเมริกาเหนือในช่วงยุคอีโอซีนเช่นกัน[ 58 ]ในอเมริกาเหนือ มี ไฮยาโนดอน เพียงไม่กี่ชนิดเท่านั้น ที่ยังคงอยู่รอดมาจนถึงยุคโอลิโกซีน[ 55 ]ในช่วงยุคไมโอซีน ไฮยาไนลูริดส์ประสบกับภาวะลดจำนวนลงอย่างมากหลังจากต้นยุคไมโอซีนและสูญพันธุ์ไปในช่วงปลายยุคไมโอซีน[ 54 ]ผู้เชี่ยวชาญหลายคนโต้แย้งว่าการสูญพันธุ์ของไฮยาโนดอนต์เกิดจากการแข่งขันกับ สัตว์ กินเนื้อ[ 54 ] [ 55 ] [ 58 ] [ 45 ] [ 61 ] [ 62 ]ผู้เชี่ยวชาญบางคนโต้แย้งว่าสัตว์กินเนื้อสามารถแข่งขันกับไฮยาโนดอนต์ได้ดีกว่าในกลุ่มสัตว์กินเนื้อขนาดกลาง ซึ่งบังคับให้ไฮยาโนดอนต์ต้องกลายเป็นสัตว์กินเนื้อขนาดใหญ่และมีความเชี่ยวชาญเฉพาะด้านมากขึ้น[ 55 ] [ 54 ] Lang et al. (2021) แนะนำว่าสัตว์กินเนื้ออาจมีบทบาทในการสูญพันธุ์ของไฮยาโนดอนต์เนื่องจากศักยภาพในการปรับตัวของฟันกรามของพวกมัน[ 61 ] Serio et al. (2024) พบว่าความแตกต่างของครีโอโดนต์แสดงให้เห็นถึงความแตกต่างทางสัณฐานวิทยาในระดับหนึ่งจนถึงยุคอีโอซีนตอนกลาง เมื่อความแตกต่างจากสัตว์กินเนื้อเพิ่มขึ้น พวกเขาโต้แย้งว่าสิ่งนี้ชี้ให้เห็นว่าสัตว์กินเนื้อเข้ามาแทนที่ไฮยาโนดอนต์ในการแข่งขัน[ 62 ] Borths และ Stevens (2019) แนะนำว่าไฮยาไนลูรีน ขนาดใหญ่ อาจสูญพันธุ์ไปเนื่องจากการสูญเสียความหลากหลายของสัตว์กินพืชขนาดใหญ่และการแข่งขันกับสัตว์กินเนื้อที่อยู่รวมกันเป็นฝูงซึ่งมีสมองที่ใหญ่กว่าและซับซ้อนกว่า ทำให้พวกมันมีความชำนาญมากขึ้นในการขโมยซากสัตว์จากไฮยาไนลูรีน[ 45 ]
อย่างไรก็ตาม สมมติฐานนี้ถูกตั้งคำถามโดยผู้เชี่ยวชาญ[ 63 ] [ 64 ] [ 65 ]การค้นพบSimbakubwaชี้ให้เห็นว่าขนาดที่ใหญ่ของพวกมันเกิดจากการเปลี่ยนแปลงของสัตว์กินพืชมากกว่าการแข่งขันกับสัตว์กินเนื้อ[ 45 ]นอกจากนี้ ขนาดสมองโดยรวมแทบไม่มีความสัมพันธ์กับพฤติกรรมทางสังคมของสัตว์กินเนื้อ[ 66 ] [ 67 ] [ 68 ]ในยุโรปHyaenodonและ amphicyonids ชอบที่อยู่อาศัยที่แตกต่างกัน โดย Hyaenodon ล่าในสภาพแวดล้อมที่เปิดโล่งมากกว่า[ 69 ]ในทางกลับกัน ปัจจุบันเชื่อกันว่าไฮยาโนดอนในยุโรปสูญพันธุ์ไปเนื่องจากการเปลี่ยนแปลงสภาพภูมิอากาศแทนที่จะเป็นการแข่งขันกับสัตว์กินเนื้อ[ 54 ] Christison et al. (2021) ได้ทำการศึกษาเกี่ยวกับแหล่งอาหารของสัตว์กินเนื้อและไฮยาโนดอนในCypress Hills Formation ผลการศึกษาของพวกเขาแสดงให้เห็นว่าไฮยาโนดอนต์ที่มีขนาดเล็กที่สุดในกลุ่มสัตว์เท่านั้นที่มีอาหารทับซ้อนกับสัตว์กินเนื้อขนาดเล็กกว่าอย่างมีนัยสำคัญ ในขณะที่สัตว์กินเนื้อขนาดใหญ่และไฮยาโนดอนต์มีความแตกต่างกันอย่างมาก โดยการศึกษาสรุปว่าไม่น่าเป็นไปได้สูงที่การสูญพันธุ์ของไฮยาโนดอนต์ในอเมริกาเหนือจะเกิดจากการแข่งขันกับสัตว์กินเนื้อ แต่พวกเขาโต้แย้งว่าการสูญพันธุ์ของไฮยาโนดอนต์อาจเกิดจากความชอบเหยื่อขนาดใหญ่ เช่น บรอนโทเธอเรสรวมถึงความไม่สามารถปรับตัวให้เข้ากับสภาพแวดล้อมที่เปิดโล่งมากขึ้นเนื่องจากขาที่ค่อนข้างสั้นของพวกมัน[ 63 ]
อ่านเพิ่มเติม
- แมคโดนัลด์, เดวิด (มกราคม 1992). กรงเล็บกำมะหยี่: ประวัติศาสตร์ธรรมชาติของสัตว์กินเนื้อ . สำนักพิมพ์บีบีซี . ISBN 978-0-563-20844-0.
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ครีโอดอนต้า
Creodonta ("ฟันเนื้อ") เป็นอันดับ ของ สัตว์ เลี้ยงลูกด้วยนมกิน เนื้อที่ สูญพันธุ์ไปแล้ว ซึ่งมีชีวิตอยู่ตั้งแต่ ยุค Paleocene ตอนต้น (หรือ Cretaceous ตอนปลาย ) จนถึง ยุค Miocene...
อนุกรมวิธานและประวัติศาสตร์
คำว่า "Creodonta" ถูกบัญญัติโดย Edward Drinker Cope ในปี 1875 [ 3 ] Cope รวม oxyaenids และ viverravid Didymictis แต่ละเว้น Hyaenodontidae ในปี 1880 เขาขยายคำนี้ให้รวมถึงวงศ์ Miacidae (รวมถึง Viverravidae), Arctocyonidae , Leptictidae (ปัจจุบัน คือ...
ฟัน
ในกลุ่มครีโอดอนท์ดั้งเดิม สูตรทางทันตกรรม คือ 3.1.4.3 3.1.4.3 แต่รูปแบบในภายหลังมักมีจำนวนฟันตัด ฟันกรามน้อย และ/หรือฟันกรามลดลง [ 39 ] ฟันเขี้ยวมีขนาดใหญ่และแหลมเสมอ ฟันตัดข้างมีขนาดใหญ่ ในขณะที่ฟันตัดตรงกลางมักมีขนาดเล็ก [ 19 ] ฟันกรามน้อยมีลักษณะดั้งเดิม...
กะโหลก
ครีโอโดนต์มีกะโหลกยาวและแคบ มีสมองขนาดเล็ก กะโหลกจะแคบลงอย่างเห็นได้ชัดด้านหลังดวงตา ทำให้เกิดส่วนกะโหลกที่เรียกว่า สปลานโนคราเนียม และ นิวโรคราเนียม อย่างชัดเจน พวกเขามี สันกระดูกซาจิตัล ขนาดใหญ่ และ กระดูกมาสตอยด์ ที่กว้าง (ซึ่งอาจเป็นลักษณะเฉพาะของกลุ่ม) [...