อ่าน 26 นาที
ยูริปเทอริด
ยูริปเทอริด (Eurypterids ) หรือที่เรียกกันอย่างไม่เป็นทางการว่าแมงป่องทะเลเป็นกลุ่มของสัตว์ขาปล้อง ในทะเลที่สูญพันธุ์ไปแล้ว ซึ่งอยู่ในอันดับยูริปเทอริดา (Eurypterida )
ยูริปเทอริด
| ยูริปเทอริด | |
|---|---|
 | |
| ตัวอย่างฟอสซิลของEurypterus remipesที่จัดแสดงอยู่ที่พิพิธภัณฑ์ประวัติศาสตร์ธรรมชาติแห่งรัฐคาร์ลสรูห์ในเมืองคาร์ลสรูห์ประเทศเยอรมนี | |
| การจำแนกทางวิทยาศาสตร์ | |
| อาณาจักร: | แอนิมอลเลีย |
| ไฟลัม: | อาร์โทรโปดา |
| ไฟลัมย่อย: | เชลิเซราตา |
| กลุ่มสายพันธุ์ : | สเคลอโรโฟราตา |
| คำสั่ง: | † ยูริพเทริดาฮักซ์ลีย์ , 1857 [ 1 ] [ a ] |
| คำสั่งย่อย | |
| |
| คำพ้องความหมาย | |
ยูริปเทอริด (Eurypterids ) หรือที่เรียกกันอย่างไม่เป็นทางการว่าแมงป่องทะเลเป็นกลุ่มของสัตว์ขาปล้อง ในทะเลที่สูญพันธุ์ไปแล้ว ซึ่งอยู่ในอันดับยูริปเทอริดา (Eurypterida ) ยูริปเทอริดที่เก่าแก่ที่สุดที่รู้จักกันนั้นมีอายุย้อนไปถึงยุคเทรมาโดเซียน (Tremadocian) ในยุค ออร์ โดวิเชียน (Ordovician) เมื่อ 480 ล้านปีก่อนกลุ่มนี้น่าจะปรากฏตัวครั้งแรกในช่วงปลายยุคแคมเบรียน (Late Cambrian) ด้วยจำนวนประมาณ 250 ชนิด ยูริปเทอริดาจึงเป็นอันดับของสัตว์มีก้าม (Celicerata) ที่มีความหลากหลายมากที่สุดในยุคพาลี โอโซอิก (Paleozoic ) หลังจากปรากฏตัวในยุคออร์โดวิเชียน ยูริปเทอริดก็กลายเป็นองค์ประกอบสำคัญของสัตว์ทะเลในยุค ไซลูเรียน (Silurian)ซึ่งเป็นยุคที่พบยูริปเทอริดส่วนใหญ่ สกุลยูริปเทอรัส (Eurypterus) ในยุคไซลูเรียน คิดเป็นมากกว่า 90% ของตัวอย่างยูริปเทอริดที่รู้จักทั้งหมด แม้ว่ากลุ่มนี้จะยังคงมีความหลากหลายมากขึ้นใน ยุค เดวอน (Devonian)แต่ยูริปเทอริดก็ได้รับผลกระทบอย่างหนักจากเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในปลายยุคเดวอน จำนวนและความหลากหลายของพวกมันลดลงในช่วงยุคเพอร์เมียน แม้ว่าช่วงเวลาที่พวกมันสูญพันธุ์ยังคงเป็นที่ถกเถียงกันอยู่ โดยมีการกล่าวอ้างว่าอาจเกิดขึ้นในช่วงต้นยุคเพอร์เมียนตอนกลางหรือเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในยุคเพอร์เมียน-ไทรแอสสิก เมื่อ 251.9 ล้านปีก่อน
แม้ว่าโดยทั่วไปจะเรียกกันว่า "แมงป่องทะเล" แต่ยูริปเทอริด (Eurypterid) ในยุคแรกๆ เท่านั้นที่อาศัยอยู่ในทะเล ยูริปเทอริด ในยุคหลังๆ หลายชนิดอาศัยอยู่ใน น้ำ กร่อยหรือน้ำจืดและพวกมันก็ไม่ใช่แมงป่อง แท้ๆ บางการศึกษาชี้ให้เห็นว่า พวกมันมี ระบบหายใจ แบบคู่ ซึ่งจะช่วยให้พวกมันสามารถอยู่ในสภาพแวดล้อมบนบกได้ในช่วงเวลาสั้นๆ ชื่อ ยูริปเทอริด ( Eurypterida)มาจากภาษากรีกโบราณεὐρύς (eurús) ซึ่งหมายถึง "กว้าง" และπτερόν (pterón) ซึ่งหมายถึง "ปีก" โดยหมายถึงระยางค์ว่ายน้ำขนาดใหญ่คู่หนึ่งที่พบในสมาชิกหลายชนิดของกลุ่มนี้
อันดับยูริปเทอริด (Eurypterid) ประกอบด้วยสัตว์ขาปล้องที่ใหญ่ที่สุดเท่าที่เคยมีชีวิตอยู่ ตัวที่ใหญ่ที่สุดคือ เจ เคิลอปเทอรัส (Jaekelopterus ) มีความยาวถึง 2.5 เมตร (8.2 ฟุต) ยูริปเทอริดไม่ได้มีขนาดใหญ่เท่ากันทั้งหมด และส่วนใหญ่มีความยาวน้อยกว่า 20 เซนติเมตร (8 นิ้ว) ยูริปเทอริดที่เล็กที่สุดคือ อัลเคนอปเทอรัส (Alkenopterus ) มีความยาวเพียง 2.03 เซนติเมตร (0.80 นิ้ว) มีการค้นพบฟอสซิลของยูริปเทอริดในทุกทวีป ฟอสซิลส่วนใหญ่มาจากแหล่งฟอสซิลในอเมริกาเหนือและยุโรปเนื่องจากกลุ่มนี้อาศัยอยู่เป็นหลักในน่านน้ำรอบๆ และภายในทวีปโบราณลอราเซีย (Laurasia ) มีเพียงไม่กี่กลุ่มของยูริปเทอริดที่แพร่กระจายออกไปนอกขอบเขตของยูราเมอริกา และมีเพียงไม่กี่สกุล เช่นอะเดโลฟทัลมัส (Adelophthalmus)และ เทอริโกตัส (Pterygotus ) ที่มีการกระจายตัวไปทั่วโลก โดยพบฟอสซิลอยู่ทั่วโลก
คำอธิบาย
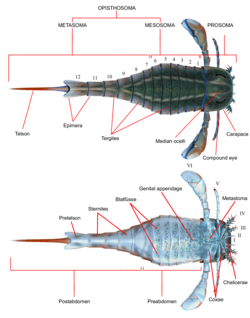
เช่นเดียวกับ สัตว์ขาปล้องอื่นๆยูริปเทอริดมี ลำ ตัวเป็นปล้องและระยางค์ (แขนขา) ที่มีข้อต่อปกคลุม ด้วย คิว ติเคิล ซึ่งประกอบด้วยโปรตีนและไคตินเช่นเดียวกับในเชลิเซอเรต อื่นๆ ลำตัว แบ่งออกเป็นสองส่วน (แท็กมาตา) คือโพรโซมา ส่วนหน้า (หัว) และโอพิสโทโซมา ส่วนหลัง (ท้อง) [ 5 ]โพรโซมาถูกปกคลุมด้วยกระดอง (บางครั้งเรียกว่า "เกราะโพรโซมา") ซึ่ง มีทั้ง ตาประกอบและโอเซลลี (อวัยวะรับความรู้สึกคล้ายตาแบบง่ายๆ) ตั้งอยู่[ 6 ]
ส่วนหัว (prosoma) ยังมีระยางค์หกคู่ ซึ่งโดยทั่วไปเรียกว่าระยางค์คู่ที่ 1 ถึง 6 ระยางค์คู่แรก ซึ่งเป็นคู่เดียวที่อยู่ด้านหน้าปาก เรียกว่า คีลิเซรา ( chelicerae) ( ซึ่งเทียบเคียงได้กับเขี้ยวของแมงมุม) คีลิเซรามีก้ามเล็กๆ ใช้สำหรับจับเศษอาหารและดันเข้าไปในปาก[ 6 ]ในสายพันธุ์หนึ่งคือPterygotidaeคีลิเซรามีขนาดใหญ่และยาว มีฟันที่แข็งแรงและพัฒนาดีบนคีเล (กรงเล็บ) ที่มีลักษณะเฉพาะ [ 7 ]ระยางค์คู่ถัดไป คือคู่ที่ 2 ถึง 6 มีฐาน ฟัน ( gnathobasesหรือ "แผ่นฟัน") บนโคกซา (ส่วนของแขนขา) ใช้สำหรับกินอาหาร ระยางค์เหล่านี้โดยทั่วไปเป็นขาเดินที่มีรูปร่างทรงกระบอกและปกคลุมด้วยหนามในบางชนิด ในสายพันธุ์ส่วนใหญ่ แขนขาจะมีขนาดใหญ่ขึ้นเมื่ออยู่ด้านหลังมากขึ้น ในอันดับย่อยEurypterina ซึ่งเป็นอันดับย่อยที่ใหญ่กว่าในสองอันดับย่อยของ Eurypterid ระยางค์คู่ที่หกยังถูกดัดแปลงเป็นใบพายว่ายน้ำเพื่อช่วยในการเดินทางในสภาพแวดล้อมทางน้ำ[ 6 ]
ส่วนท้องประกอบด้วย 12 ปล้องและส่วนหางซึ่งเป็นส่วนท้ายสุดของร่างกาย ซึ่งในสายพันธุ์ส่วนใหญ่มีรูปร่างคล้ายใบมีด[ 6 ]ในบางสายพันธุ์ โดยเฉพาะอย่างยิ่งPterygotioidea , HibbertopteridaeและMycteroptidaeส่วนหางจะแบนราบและอาจใช้เป็นหางเสือขณะว่ายน้ำ สกุลบางสกุลในวงศ์ใหญ่Carcinosomatoideaโดยเฉพาะอย่างยิ่งEusarcana มีส่วนหางที่คล้ายกับ แมงป่องแท้และอาจสามารถใช้ฉีดพิษ ได้ เช่นเดียวกับแมงป่อง[ 8 ] [ 9 ]โคนขาของระยางค์คู่ที่หกถูกคลุมด้วยแผ่นที่เรียกว่าเมตาสโตมา ซึ่งเดิมทีมาจากส่วนของโครงกระดูกภายนอกที่สมบูรณ์ ส่วนท้องสามารถแบ่งออกได้เป็น " ส่วนท้องกลาง " (ประกอบด้วยปล้องที่ 1 ถึง 6) และ " ส่วนท้องท้าย " (ประกอบด้วยปล้องที่ 7 ถึง 12) หรือเป็น "ส่วนท้องก่อน" (โดยทั่วไปประกอบด้วยปล้องที่ 1 ถึง 7) และ "ส่วนท้องหลัง" (โดยทั่วไปประกอบด้วยปล้องที่ 8 ถึง 12) [ 6 ]
ด้านล่างของออพิสโทโซมาถูกปกคลุมด้วยโครงสร้างที่พัฒนามาจากรยางค์ออพิสโทโซมาที่ดัดแปลงแล้ว ตลอดทั้งออพิสโทโซมา โครงสร้างเหล่านี้ก่อตัวเป็นโครงสร้างคล้ายแผ่นที่เรียกว่าBlattfüsse ( แปลว่า' เท้าใบไม้'ในภาษาเยอรมัน) [ 10 ]สิ่งเหล่านี้สร้างช่องเหงือก (ทางเดินเหงือก) ระหว่างBlattfüsse ก่อนหน้า กับ พื้นผิว ด้านล่างของออพิสโทโซมาเอง ซึ่งมีอวัยวะหายใจอยู่ ปล้องออพิสโทโซมาที่สองถึงหกยังมีอวัยวะรูปไข่หรือรูปสามเหลี่ยมซึ่งถูกตีความว่าเป็นอวัยวะที่ช่วยในการหายใจ อวัยวะเหล่านี้เรียกว่าKiemenplatten หรือ "ทางเดินเหงือก" ซึ่งอาจช่วยให้ยูริ ปเทอริดหายใจอากาศเหนือน้ำได้ ในขณะที่Blattfüssenคล้ายกับอวัยวะในปูม้า ในปัจจุบัน จะปกคลุมส่วนที่ทำหน้าที่ในการหายใจใต้น้ำ [ 6 ]
ส่วนยื่นของปล้องโอพิสโทโซมัลที่ 1 และ 2 (ปล้องที่ 7 และ 8 โดยรวม) เชื่อมรวมกันเป็นโครงสร้างที่เรียกว่าฝาปิดอวัยวะสืบพันธุ์ ซึ่งครอบคลุมพื้นที่ส่วนใหญ่ของด้านล่างของปล้องโอพิสโทโซมัลที่ 2 ใกล้กับ ขอบ ด้านหน้าของโครงสร้างนี้ ส่วนยื่นอวัยวะสืบพันธุ์ (เรียกอีกอย่างว่าZipfelหรือส่วนยื่นช่องท้องตรงกลาง) ยื่นออกมา ส่วนยื่นนี้มักจะได้รับการอนุรักษ์ไว้อย่างเด่นชัด และได้รับการตีความอย่างสม่ำเสมอว่าเป็นส่วนหนึ่งของระบบสืบพันธุ์ และมีอยู่ 2 ประเภทที่ได้รับการยอมรับ ซึ่งสันนิษฐานว่าสอดคล้องกับเพศผู้และเพศเมีย[ 6 ]
ชีววิทยา
ขนาด

ยูริปเทอริดมีขนาดที่หลากหลายมาก ขึ้นอยู่กับปัจจัยต่างๆ เช่น วิถีชีวิต สภาพแวดล้อมในการดำรงชีวิต และความสัมพันธ์ทางอนุกรมวิธานขนาดประมาณ 100 เซนติเมตร (3.3 ฟุต) เป็นเรื่องปกติในกลุ่มยูริปเทอริดส่วนใหญ่[ 11 ]ยูริปเทอริดที่เล็กที่สุดคือAlkenopterus burglahrensisมีความยาวเพียง 2.03 เซนติเมตร (0.80 นิ้ว) [ 12 ]
สัตว์ในวงศ์ Eurypteridae ที่ใหญ่ที่สุด และเป็นสัตว์ขาปล้องที่ใหญ่ที่สุดเท่าที่เคยมีชีวิตอยู่ คือJaekelopterus rhenaniaeก้ามของมันจากชั้นหินEmsian Klerf Formationใน Willwerath ประเทศเยอรมนีมีความยาว 36.4 เซนติเมตร (14.3 นิ้ว) แต่ขาดไปหนึ่งในสี่ของความยาวทั้งหมด ซึ่งบ่งชี้ว่าก้ามที่สมบูรณ์น่าจะมีความยาว 45.5 เซนติเมตร (17.9 นิ้ว) หากสัดส่วนระหว่างความยาวลำตัวและก้ามตรงกับญาติสนิทของมัน ซึ่งอัตราส่วนระหว่างขนาดก้ามและความยาวลำตัวค่อนข้างคงที่ ตัวอย่างของJaekelopterusที่มีก้ามชิ้นนี้จะมีความยาวระหว่าง 233 ถึง 259 เซนติเมตร (7.64 ถึง 8.50 ฟุต) โดยเฉลี่ย 2.5 เมตร (8.2 ฟุต) เมื่อยืดก้ามออก ความยาวจะเพิ่มขึ้นอีก 1 เมตร (3.28 ฟุต) การประมาณค่านี้เกินขนาดตัวสูงสุดของสัตว์ขาปล้องยักษ์ที่รู้จักทั้งหมดไปเกือบครึ่งเมตร (1.64 ฟุต) แม้ว่าจะไม่รวมก้ามที่ยื่นออกมาก็ตาม[ 13 ]ยูริปเทอริดอีกสองชนิดก็ได้รับการประมาณค่าว่ามีความยาวถึง 2.5 เมตรเช่นกัน ได้แก่Erettopterus grandis (มีความสัมพันธ์ใกล้ชิดกับJaekelopterus ) และHibbertopterus wittebergensisแต่E. grandisนั้นมีซากที่แตกหักมาก และ การประมาณขนาดของ H. wittebergensisนั้นอิงจากหลักฐานร่องรอย ไม่ใช่ซากดึกดำบรรพ์[ 14 ]
วงศ์Jaekelopterusหรือ Pterygotidae มีชื่อเสียงในเรื่องสายพันธุ์ที่มีขนาดใหญ่ผิดปกติหลายชนิด ทั้งAcutiramusซึ่งสมาชิกที่ใหญ่ที่สุดคือA. bohemicusมีความยาว 2.1 เมตร (6.9 ฟุต) และPterygotusซึ่งสายพันธุ์ที่ใหญ่ที่สุดคือ P. grandidentatusมีความยาว 1.75 เมตร (5.7 ฟุต) ต่างก็มีขนาดใหญ่โตมโหฬาร[ 13 ]มีการเสนอปัจจัยหลายประการที่ทำให้ Pterygotidae มีขนาดใหญ่ รวมถึงพฤติกรรมการเกี้ยวพาราสี การล่าเหยื่อ และการแข่งขันแย่งชิงทรัพยากรสิ่งแวดล้อม[ 15 ]
ยูริปเทอริดขนาดยักษ์ไม่ได้จำกัดอยู่เฉพาะในวงศ์ Pterygotidae เท่านั้น ฟอสซิลเมตาสโตมาที่แยกออกมายาว 12.7 เซนติเมตร (5.0 นิ้ว) ของยูริปเทอริดคาร์ซิโนโซมาตอยด์Carcinosoma punctatumบ่งชี้ว่าสัตว์ชนิดนี้จะมีขนาดความยาวถึง 2.2 เมตร (7.2 ฟุต) ในช่วงมีชีวิต ซึ่งมีขนาดเทียบเท่ากับพเทอริโกทิด[ 16 ]อีกหนึ่งสัตว์ยักษ์คือPentecopterus decorahensisซึ่งเป็นคาร์ซิโนโซมาตอยด์ดั้งเดิม ซึ่งคาดว่ามีความยาวถึง 1.7 เมตร (5.6 ฟุต) [ 17 ]
ลักษณะทั่วไปของยูริปเทอริดขนาดใหญ่คือโครงสร้างที่เบา ปัจจัยต่างๆ เช่น การเคลื่อนที่ ต้นทุนพลังงานในการลอกคราบและการหายใจ รวมถึงคุณสมบัติทางกายภาพของโครงกระดูกภายนอกล้วนจำกัดขนาดที่อาร์โทรพอดสามารถมีได้ โครงสร้างที่เบาช่วยลดอิทธิพลของปัจจัยเหล่านี้ได้อย่างมาก พเทอริโกทิดมีน้ำหนักเบาเป็นพิเศษ โดยส่วนลำตัวขนาดใหญ่ที่กลายเป็นฟอสซิลส่วนใหญ่ยังคงมีลักษณะบางและไม่มีแร่ธาตุ[ 13 ]การปรับตัวให้เบายังพบได้ในอาร์โทรพอดขนาดยักษ์ในยุคพาลีโอโซอิกอื่นๆ เช่น ตะขาบยักษ์อาร์โทรพลูราและอาจมีความสำคัญต่อวิวัฒนาการของขนาดยักษ์ในอาร์โทรพอด[ 13 ] [ 18 ]
นอกจากยูริปเทอริดขนาดยักษ์ที่มีน้ำหนักเบาแล้ว สัตว์บางชนิดในวงศ์ Hibbertopteridae ที่มีลำตัวลึกก็มีขนาดใหญ่มากเช่นกัน กระดองจากยุคคาร์บอนิเฟอรัสของสกอตแลนด์ที่ระบุว่าเป็นสายพันธุ์Hibbertoperus scouleriมีความกว้าง 65 ซม. (26 นิ้ว) เนื่องจากHibbertopterusมีความกว้างมากเมื่อเทียบกับความยาว สัตว์ชนิดนี้จึงอาจมีความยาวเกือบ 2 เมตร (6.6 ฟุต) แข็งแรงกว่าเทอริโกทิดส์Hibbertopterus ขนาดยักษ์นี้ อาจมีน้ำหนักเทียบเท่ากับเทอริโกทิดส์ที่ใหญ่ที่สุด หรืออาจจะมากกว่า และด้วยเหตุนี้จึงจัดอยู่ในกลุ่มสัตว์ขาปล้องที่มีน้ำหนักมากที่สุด[ 19 ]
การเคลื่อนที่


กลุ่มย่อยยูริปเทอริดสองกลุ่ม ได้แก่ ยูริปเทอรินาและสไตโลนูรินาแตกต่างกันหลักๆ ที่รูปร่างของระยางค์คู่สุดท้าย ในสไตโลนูรินา ระยางค์นี้มีลักษณะเป็นขาเดินที่ยาวและเรียว ในขณะที่ในยูริปเทอรินา ขาจะถูกดัดแปลงและขยายออกเป็นพายสำหรับว่ายน้ำ[ 20 ]นอกเหนือจากพายสำหรับว่ายน้ำแล้ว ขาของยูริปเทอรีนจำนวนมากมีขนาดเล็กเกินไปที่จะทำอะไรได้มากไปกว่าการคลานไปบนพื้นทะเลในทางตรงกันข้าม สไตโลนูริรีนจำนวนหนึ่งมีขาที่ยาวและแข็งแรงซึ่งอาจช่วยให้พวกมันเดินบนบกได้ (คล้ายกับปู ในปัจจุบัน ) [ 21 ]
มีการค้นพบ ร่องรอยฟอสซิลในแหล่งฟอสซิลยุคคาร์บอนิเฟอรัสของสกอตแลนด์ในปี 2548 ร่องรอยดังกล่าวถูกระบุว่าเป็นของฮิบเบอร์ทอปเทอรัส ( Hibbertopterus ) ซึ่งเป็นสัตว์ในกลุ่มยูริปเทอริดในกลุ่มสไตโลนูรีน (Stylonurine Eurypterid) เนื่องจากมีขนาดที่ตรงกัน (คาดว่าผู้สร้างร่องรอยมีความยาวประมาณ 1.6 เมตร (5.2 ฟุต)) และลักษณะทางกายวิภาคของขาที่อนุมานได้ นี่คือร่องรอยบนบกที่ใหญ่ที่สุดที่สร้างโดยสัตว์ขาปล้องจนถึงปัจจุบัน โดยมีความยาว 6 เมตร (20 ฟุต) และกว้างเฉลี่ย 95 เซนติเมตร (3.12 ฟุต) นอกจากนี้ยังเป็นบันทึกแรกของการเคลื่อนที่บนบกของยูริปเทอริด ร่องรอยนี้เป็นหลักฐานที่แสดงว่ายูริปเทอริดบางชนิดสามารถอยู่รอดได้ในสภาพแวดล้อมบนบก อย่างน้อยก็ในช่วงเวลาสั้นๆ และยังเผยข้อมูลเกี่ยวกับลักษณะการเดินของสไตโลนูรีนอีกด้วย ในHibbertopterusเช่นเดียวกับยูริปเทอริดส่วนใหญ่ คู่ของระยางค์จะมีขนาดแตกต่างกัน (เรียกว่าภาวะระยางค์เฮเทอโรโพดัส) ระยางค์คู่ที่มีขนาดแตกต่างกันเหล่านี้จะเคลื่อนที่ไปพร้อมกัน และระยะก้าวที่สั้นแสดงให้เห็นว่าHibbertopterusคลานด้วยความเร็วที่ช้าเป็นพิเศษ อย่างน้อยก็บนบก เทลสันขนาดใหญ่ถูกลากไปตามพื้นและทิ้งร่องกลางขนาดใหญ่ไว้ด้านหลังสัตว์ ความลาดชันในรอยเท้าในช่วงเวลาสุ่มบ่งชี้ว่าการเคลื่อนไหวนั้นกระตุก[ 22 ]การเดินของสไตโลนูรีนขนาดเล็กกว่า เช่นParastylonurusน่าจะเร็วกว่าและแม่นยำกว่า[ 23 ]
การทำงานของใบพายว่ายน้ำของยูริปเทอรีนแตกต่างกันไปในแต่ละกลุ่ม ในยูริปเทอรอยเดียใบพายมีรูปร่างคล้ายกับไม้พาย สภาพของข้อต่อในระยางค์ทำให้ใบพายสามารถเคลื่อนที่ได้เฉพาะในระนาบเกือบแนวนอนเท่านั้น ไม่สามารถเคลื่อนที่ขึ้นหรือลงได้ กลุ่มอื่นๆ เช่น พเทอริโกทิโอเดีย อาจไม่มีสภาพเช่นนี้และอาจว่ายน้ำได้เร็วกว่า[ 24 ]โดยทั่วไปแล้วยูริปเทอรีนส่วนใหญ่เห็นพ้องกันว่าใช้การขับเคลื่อนแบบพายคล้ายกับปูและด้วงน้ำตัวที่ใหญ่กว่าอาจสามารถบินใต้น้ำได้ (หรือบินใต้น้ำ ) ซึ่งการเคลื่อนไหวและรูปร่างของใบพายเพียงพอที่จะสร้างแรงยกคล้ายกับการว่ายน้ำของเต่าทะเลและสิงโตทะเลการเคลื่อนไหวประเภทนี้มีอัตราเร่งที่ค่อนข้างช้ากว่าแบบพาย โดยเฉพาะอย่างยิ่งเนื่องจากตัวเต็มวัยมีใบพายที่เล็กกว่าเมื่อเทียบกับตัวอ่อน อย่างไรก็ตาม เนื่องจากขนาดที่ใหญ่ขึ้นของผู้ใหญ่หมายถึงค่าสัมประสิทธิ์แรงต้าน ที่สูงขึ้น การใช้ระบบขับเคลื่อนประเภทนี้จึงมีประสิทธิภาพด้านพลังงานมากกว่า[ 25 ]

ยูริปเทอรีนบางชนิด เช่นมิกซอปเทอรัส (ตามที่อนุมานจากร่องรอยฟอสซิล) อาจไม่ได้เป็นนักว่ายน้ำที่ดีนัก มันน่าจะอาศัยอยู่บริเวณก้นทะเลเป็นส่วนใหญ่ โดยใช้ครีบว่ายน้ำเพื่อเคลื่อนที่ขึ้นลงเป็นครั้งคราว โดยมีระยางค์คู่ที่สี่และห้าวางอยู่ด้านหลังเพื่อสร้างการเคลื่อนที่ไปข้างหน้าเล็กน้อย ในขณะเดิน มันอาจใช้ท่าทางการเดินคล้ายกับแมลงสมัยใหม่ส่วนใหญ่ น้ำหนักของท้องที่ยาวของมันจะถูกถ่วงดุลด้วยระยางค์ด้านหน้าที่หนักและเฉพาะทางสองอัน และจุดศูนย์ถ่วงอาจปรับได้โดยการยกและจัดตำแหน่งหาง[ 26 ]
ร่องรอยเท้าของยูริปเทอริดที่กลายเป็นฟอสซิลมักมีขนาดใหญ่และมีลักษณะแตกต่างกัน และมักมีรอยลากของเทลสันอยู่ตามแนวกลางลำตัว (เช่นเดียวกับ ร่องรอยเท้า ของฮิบเบอร์ทอปเทอ รัสในสกอตแลนด์ ) ร่องรอยเท้าดังกล่าวถูกค้นพบในทุกทวีปยกเว้นอเมริกาใต้ ในบางสถานที่ที่ซากฟอสซิลของยูริปเทอริดหายาก เช่น ในแอฟริกาใต้และส่วนที่เหลือของอดีตมหาทวีปก็อนด์วานาการค้นพบร่องรอยเท้าเกิดขึ้นก่อนและมีจำนวนมากกว่าฟอสซิลตัวของยูริปเทอริด[ 27 ]ร่องรอยเท้าของยูริปเทอริดถูกจัดอยู่ในหลายสกุลร่องรอยเท้า โดยเฉพาะอย่างยิ่งPalmichnium (ซึ่งกำหนดเป็นชุดของร่องรอยเท้าสี่ร่องรอยที่มักมีรอยลากอยู่ตามแนวกลางลำตัว) [ 28 ]ซึ่งตัวอย่างต้นแบบของชนิดร่องรอยเท้าP. kosinkiorumเก็บรักษาร่องรอยเท้าของยูริปเทอริดที่ใหญ่ที่สุดเท่าที่รู้จักในปัจจุบัน โดยร่องรอยเท้าแต่ละร่องรอยมีเส้นผ่านศูนย์กลางประมาณ 7.6 เซนติเมตร (3.0 นิ้ว) [ 29 ]สกุลร่องรอยยูริปเทอริดอื่นๆ ได้แก่เมโรสโตมิคนิทส์ (แม้ว่าตัวอย่างจำนวนมากอาจแสดงถึงร่องรอยของสัตว์จำพวกครัสเตเชียน) และอาร์คิวส์ (ซึ่งเก็บรักษาร่องรอยที่เกิดจากระยางค์ว่ายน้ำ) [ 28 ] [ 30 ] [ 31 ]
การหายใจ

ในยูริปเทอริด อวัยวะระบบหายใจตั้งอยู่บนผนังลำตัวด้านท้อง (ด้านล่างของออพิสโทโซมา) บลัตต์ฟัสเซซึ่งวิวัฒนาการมาจากรยางค์ออพิสโทโซมา ปกคลุมด้านล่างและสร้างช่องเหงือกซึ่งเป็นที่ตั้งของ "ทางเดินเหงือก" [ 6 ]ขึ้นอยู่กับชนิด ทางเดินเหงือกของยูริปเทอริดอาจมีรูปร่างเป็นสามเหลี่ยมหรือรูปไข่ และอาจยกขึ้นเป็นรูปทรงคล้ายเบาะ พื้นผิวของทางเดินเหงือกนี้มีหนามเล็กๆ หลายอัน ทำให้มีพื้นที่ผิวเพิ่มขึ้น ประกอบด้วยเนื้อเยื่อฟองน้ำเนื่องจากการเว้าเข้าไป จำนวนมาก ในโครงสร้าง[ 32 ]
แม้ว่าKiemenplatteจะถูกเรียกว่า "ทางเดินเหงือก" แต่ก็อาจไม่ได้ทำหน้าที่เป็นเหงือกจริงๆ ในสัตว์ชนิดอื่นๆ เหงือกทำหน้าที่ดูดซับออกซิเจนจากน้ำและเป็นส่วนที่ยื่นออกมาจากผนังลำตัว ถึงแม้ว่ายูริปเทอริดจะเป็นสัตว์น้ำเป็นหลักและวิวัฒนาการใต้น้ำอย่างแน่นอน (ยูริปเทอริดบางชนิด เช่น เทอริโกทิด อาจไม่สามารถเดินบนบกได้ด้วยซ้ำ) แต่ก็ไม่น่าเป็นไปได้ที่ "ทางเดินเหงือก" จะมีเหงือกที่ใช้งานได้ เมื่อเปรียบเทียบกับเหงือกในสัตว์ไม่มีกระดูกสันหลังชนิดอื่นๆ หรือแม้แต่ปลา การตีความก่อนหน้านี้มักระบุว่า "เหงือก" ของยูริปเทอริดมีความคล้ายคลึงกับเหงือกของกลุ่มอื่นๆ (จึงเป็นที่มาของคำศัพท์นี้) โดยมีการแลกเปลี่ยนก๊าซเกิดขึ้นภายในทางเดินที่เป็นฟองน้ำ และมีรูปแบบของเส้นเลือด branchio-cardiac และ dendritic (เช่นเดียวกับในกลุ่มที่เกี่ยวข้อง) ที่นำเลือดที่มีออกซิเจนเข้าสู่ร่างกาย การเปรียบเทียบหลักที่ใช้ในการศึกษาครั้งก่อนๆ คือปูม้า แม้ว่าโครงสร้างเหงือกของปูม้าและยูริปเทอริดจะแตกต่างกันอย่างมากก็ตาม ในปูม้า เหงือกมีความซับซ้อนกว่าและประกอบด้วยแผ่นบางๆ จำนวนมาก ซึ่งให้พื้นที่ผิวที่ใช้ในการแลกเปลี่ยนก๊าซมากขึ้น นอกจากนี้ ทางเดินเหงือกของยูริปเทอริดมีขนาดเล็กเกินไปเมื่อเทียบกับสัดส่วนที่จะรองรับพวกมันได้ หากเปรียบเทียบกับเหงือกของกลุ่มอื่นๆ เหงือกที่ใช้งานได้จะต้องมีประสิทธิภาพสูงและต้องมีระบบไหลเวียนโลหิตที่มีประสิทธิภาพสูง อย่างไรก็ตาม ถือว่าไม่น่าเป็นไปได้ที่ปัจจัยเหล่านี้จะเพียงพอที่จะอธิบายความแตกต่างอย่างมากระหว่างขนาดของทางเดินเหงือกและขนาดของร่างกาย[ 33 ]
มีการเสนอแนะว่า "ทางเดินเหงือก" อาจเป็นอวัยวะสำหรับหายใจเอาอากาศเข้าไป ซึ่งอาจจะเป็นปอดกระดองหรือท่อหายใจเทียม [ 34 ] กระดอง เป็นอวัยวะที่สัตว์ขาปล้องบางชนิดวิวัฒนาการขึ้นมาเพื่อหายใจเอาอากาศใต้น้ำ ซึ่งถือว่าเป็นคำอธิบายที่ไม่น่าเป็นไปได้ เนื่องจากยูริ ปเทอริดวิวัฒนาการอยู่ในน้ำตั้งแต่แรกเริ่ม และพวกมันจะไม่มีอวัยวะที่วิวัฒนาการมาจากอวัยวะหายใจเอาอากาศ นอกจากนี้ กระดองมักจะอยู่ด้านนอกของร่างกาย ในขณะที่ทางเดินเหงือกของยูริปเทอริดตั้งอยู่ด้านหลังBlattfüssen [ 35 ]ในทางกลับกัน ในบรรดาอวัยวะหายใจของสัตว์ขาปล้อง ทางเดินเหงือกของยูริปเทอริดมีความคล้ายคลึงกับท่อหายใจเทียมที่พบในไอโซพอดในปัจจุบัน มากที่สุด อวัยวะเหล่านี้เรียกว่า pseudotracheae เนื่องจากมีความคล้ายคลึงกับtracheae (ท่อลม) ของสิ่งมีชีวิตที่หายใจด้วยอากาศ มีลักษณะคล้ายปอดและอยู่ภายในpleopods (ขาหลัง) ของ isopods โครงสร้างของ pseudotracheae ได้รับการเปรียบเทียบกับโครงสร้างที่เป็นฟองน้ำของทางเดินเหงือกของ eurypterid เป็นไปได้ว่าอวัยวะทั้งสองทำหน้าที่ในลักษณะเดียวกัน[ 36 ]
นักวิจัยบางคนเสนอว่ายูริปเทอริดอาจปรับตัวให้เข้ากับวิถีชีวิตแบบสะเทินน้ำสะเทินบก โดยใช้โครงสร้างทางเดินเหงือกทั้งหมดเป็นเหงือก และใช้ส่วนที่เว้าเข้าไปภายในเป็นหลอดลมเทียม อย่างไรก็ตาม รูปแบบการดำรงชีวิตนี้อาจเป็นไปไม่ได้ในทางสรีรวิทยา เนื่องจากแรงดันน้ำจะดันน้ำเข้าไปในส่วนที่เว้าเข้าไป ทำให้เกิดการขาดอากาศหายใจนอกจากนี้ ยูริปเทอริดส่วนใหญ่จะอาศัยอยู่ในน้ำตลอดชีวิต ไม่ว่าพวกมันจะใช้เวลาอยู่บนบกมากน้อยเพียงใด อวัยวะสำหรับการหายใจในสภาพแวดล้อมใต้น้ำก็ต้องมีอยู่ เหงือกที่แท้จริง ซึ่งคาดว่าจะอยู่ภายในช่องเหงือกในBlattfüssenยังคงไม่เป็นที่รู้จักในยูริปเทอริด[ 36 ]
พัฒนาการ

เช่นเดียวกับสัตว์ขาปล้องทั้งหมด ยูริปเทอริดเจริญเติบโตและพัฒนาผ่านขั้นตอนการพัฒนาแบบคงที่ที่เรียกว่าอินสตาร์อินสตาร์เหล่านี้ถูกคั่นด้วยช่วงเวลาที่ยูริปเทอริดลอกคราบ(การลอกคราบของคิวติเคิล) หลังจากนั้นพวกมันจะเจริญเติบโตอย่างรวดเร็วและทันที สัตว์ขาปล้องบางชนิด เช่น แมลงและกุ้งหลายชนิด มีการเปลี่ยนแปลงอย่างมากในช่วงการเจริญเติบโต เชลิเซอเรต รวมถึงยูริปเทอริด โดยทั่วไปถือว่าเป็นสัตว์ที่พัฒนาโดยตรง โดยไม่มีการเปลี่ยนแปลงอย่างมากหลังจากการฟักไข่ (แม้ว่าอาจมีส่วนลำตัวและแขนขาเพิ่มขึ้นในระหว่างการเจริญเติบโตในบางสายพันธุ์ เช่นซิโฟซูแรนและแมงมุมทะเล ) ในอดีตมีการถกเถียงกันว่ายูริปเทอริดเป็นสัตว์ที่พัฒนาโดยตรงอย่างแท้จริง (โดยที่ลูกอ่อนมีลักษณะเหมือนกับตัวเต็มวัย) หรือเป็นสัตว์ที่พัฒนาโดยตรงแบบเฮมิอานามอร์ฟิก (โดยอาจมีส่วนลำตัวและแขนขาเพิ่มขึ้นในระหว่างการเจริญเติบโต) [ 37 ]
การพัฒนาโดยตรงแบบเฮมิอานามอร์ฟิกได้รับการสังเกตในกลุ่มสัตว์ขาปล้องหลายกลุ่ม เช่นไทรโลไบต์เมกาไคแรนครัสเตเชียนพื้นฐานและไมริอาพอดพื้นฐานการพัฒนาโดยตรงที่แท้จริงบางครั้งถูกกล่าวถึงว่าเป็นลักษณะเฉพาะของแมงมุมมีการศึกษาเกี่ยวกับการเจริญเติบโตของยูริปเทอริดน้อยมาก เนื่องจากโดยทั่วไปแล้วมีตัวอย่างในบันทึกฟอสซิลน้อยมากที่สามารถระบุได้อย่างมั่นใจว่าเป็นตัวแทนของตัวอ่อน[ 37 ]เป็นไปได้ว่ายูริปเทอริดหลายชนิดที่คิดว่าแตกต่างกันนั้น แท้จริงแล้วอาจเป็นตัวอย่างตัวอ่อนของสายพันธุ์อื่น โดยที่นักบรรพชีวินวิทยาไม่ค่อยพิจารณาอิทธิพลของการเจริญเติบโตเมื่ออธิบายสายพันธุ์ใหม่[ 38 ]
การศึกษาฟอสซิลยูริปเทอริดที่ได้รับการอนุรักษ์ไว้อย่างดีจากแหล่งฟอสซิลBeartooth Butte Formation ใน ยุคPragianในCottonwood Canyonรัฐไวโอมิงซึ่งประกอบด้วยตัวอย่างหลายชิ้นจากระยะการพัฒนาต่างๆ ของยูริปเทอริดJaekelopterusและStrobilopterusเผยให้เห็นว่าการเจริญเติบโตของยูริปเทอริดนั้นค่อนข้างขนานและคล้ายคลึงกับการเจริญเติบโตของซิโฟซูแรนที่สูญพันธุ์และยังมีชีวิตอยู่ โดยมีข้อแตกต่างที่สำคัญที่สุดคือยูริปเทอริดฟักออกมาพร้อมกับระยางค์และปล้องโอพิสโทโซมัลครบชุด ดังนั้นยูริปเทอริดจึงไม่ใช่สิ่งมีชีวิตที่พัฒนาโดยตรงแบบเฮมิอานามอร์ฟิก แต่เป็นสิ่งมีชีวิตที่พัฒนาโดยตรงอย่างแท้จริงเช่นเดียวกับแมงมุมในปัจจุบัน[ 39 ]
การเปลี่ยนแปลงที่สังเกตได้บ่อยที่สุดที่เกิดขึ้นตลอดการเจริญเติบโต (ยกเว้นบางสกุล เช่นEurypterusซึ่งดูเหมือนจะคงที่) คือ เมตาสโตมามีความกว้างน้อยลงตามสัดส่วน การเปลี่ยนแปลงการเจริญเติบโตนี้ได้รับการสังเกตในสมาชิกของวงศ์ใหญ่หลายวงศ์ เช่น Eurypteroidea, Pterygotioidea และMoselopteroidea [ 40 ]
การให้อาหาร

ไม่พบซากดึกดำบรรพ์ของอาหารในกระเพาะของยูริปเทอริด ดังนั้นจึงไม่มีหลักฐานโดยตรงเกี่ยวกับอาหารของพวกมัน ชีววิทยาของยูริปเทอริดบ่งชี้อย่างชัดเจนถึงวิถีชีวิตแบบกินเนื้อเป็นอาหาร ไม่เพียงแต่พวกมันจะมีขนาดใหญ่ (โดยทั่วไปแล้ว ผู้ล่าส่วนใหญ่มักจะมีขนาดใหญ่กว่าเหยื่อ) แต่พวกมันยังมีการมองเห็นแบบสามมิติ (ความสามารถในการรับรู้ความลึก) [ 41 ]ขาของยูริปเทอริดหลายตัวถูกปกคลุมด้วยหนามบางๆ ซึ่งใช้ทั้งในการเคลื่อนที่และการหาอาหาร ในบางกลุ่ม รยางค์ที่มีหนามเหล่านี้มีความเชี่ยวชาญเฉพาะด้านอย่างมาก ในยูริปเทอริดบางชนิดในกลุ่ม Carcinosomatoidea รยางค์ที่หันไปข้างหน้ามีขนาดใหญ่และมีหนามที่ยาวมาก (เช่นในMixopterusและMegalograptus ) ใน สมาชิก ที่พัฒนาแล้วของกลุ่ม Pterygotioidea รยางค์ไม่มีหนามเลย แต่มีกรงเล็บที่เชี่ยวชาญแทน[ 42 ]ยูริปเทอริดอื่นๆ ที่ไม่มีระยางค์พิเศษเหล่านี้ น่าจะกินอาหารในลักษณะที่คล้ายกับปูม้าในปัจจุบัน โดยการจับและฉีกอาหารด้วยระยางค์ก่อนที่จะดันเข้าไปในปากโดยใช้ก้าม[ 43 ]
มีการรายงานฟอสซิลที่เก็บรักษาระบบทางเดินอาหาร จากฟอสซิลของยูริปเทอริดหลายชนิด รวมถึง คาร์ซิโนโซมาอะคูติรามัสและยูริปเทอรัสแม้ว่าจะมีการรายงานช่องทวารหนักที่อาจเกิดขึ้นได้จากเทลสันของตัวอย่างบัฟฟาโลปเทอรัส แต่ ก็มีความเป็นไปได้มากกว่าที่ทวารหนักจะเปิดผ่านคิวติเคิลบาง ๆ ระหว่างปล้องสุดท้ายก่อนเทลสันและเทลสันเอง เช่นเดียวกับในปูม้าในปัจจุบัน[ 41 ]
อุจจาระฟอสซิลของยูริปเทอริดที่ค้นพบในแหล่งสะสมยุคออร์โดวิเชียนในโอไฮโอ ซึ่งประกอบด้วยชิ้นส่วนของไทรโลไบต์และยูริปเทอริดMegalograptus ohioensisร่วมกับตัวอย่างที่สมบูรณ์ของยูริปเทอริดชนิดเดียวกันนั้น ได้รับการเสนอแนะว่าเป็นหลักฐานของ การกินพวกเดียวกัน อุจจาระฟอสซิลที่คล้ายกันซึ่งจัดอยู่ในสายพันธุ์Lanarkopterus dolichoschelusจากยุคออร์โดวิเชียนของโอไฮโอ ประกอบด้วยชิ้นส่วนของปลาที่ไม่มีขากรรไกรและชิ้นส่วนของตัวอย่างLanarkopterus ขนาดเล็ก กว่า[ 41 ]
แม้ว่า บทบาท การล่าสูงสุดจะจำกัดอยู่เฉพาะยูริปเทอริดขนาดใหญ่ที่สุดเท่านั้น แต่ยูริปเทอริดขนาดเล็กก็อาจเป็นนักล่าที่น่าเกรงขามได้เช่นเดียวกับญาติที่มีขนาดใหญ่กว่า[ 11 ]
ชีววิทยาการสืบพันธุ์


เช่นเดียวกับกลุ่มที่สูญพันธุ์ไปอย่างสิ้นเชิงอื่นๆ การทำความเข้าใจและวิจัยเกี่ยวกับการสืบพันธุ์และความแตกต่างทางเพศของยูริปเทอริดเป็นเรื่องยาก เนื่องจากเรารู้จักพวกมันจากเปลือกและกระดองฟอสซิลเท่านั้น ในบางกรณี อาจไม่มีความแตกต่างที่ชัดเจนเพียงพอที่จะแยกเพศโดยอาศัยสัณฐานวิทยาเพียงอย่างเดียว[ 21 ]บางครั้งเพศสองเพศของสายพันธุ์เดียวกันถูกตีความว่าเป็นสองสายพันธุ์ที่แตกต่างกัน ดังเช่นกรณีของสองสายพันธุ์ของDrepanopterus ( D. bembycoidesและD. lobatus ) [ 44 ]
โพรโซมาของยูริปเทอริดประกอบด้วยปล้องโครงกระดูกภายนอกหกปล้องแรกที่เชื่อมติดกันเป็นโครงสร้างขนาดใหญ่ ปล้องที่เจ็ด (ดังนั้นจึงเป็นปล้องออพิสโทโซมัลแรก) เรียกว่าเมตาสโตมาและปล้องที่แปด (มีลักษณะเป็นแผ่นอย่างชัดเจน) เรียกว่าโอเปอร์คูลัมและมีช่องเปิดอวัยวะสืบพันธุ์ ด้านล่างของปล้องนี้เป็นที่ตั้งของโอเปอร์คูลัมอวัยวะสืบพันธุ์ ซึ่งเป็นโครงสร้างที่วิวัฒนาการมาจากรยางค์คู่ที่เจ็ดและแปดของบรรพบุรุษ ตรงกลางของมัน เช่นเดียวกับในปูม้าในปัจจุบัน มีรยางค์อวัยวะสืบพันธุ์ รยางค์นี้เป็นแท่งยาวที่มีท่อภายใน พบได้ในสองรูปแบบที่แตกต่างกัน โดยทั่วไปเรียกว่า "แบบ A" และ "แบบ B" [ 21 ]รยางค์อวัยวะสืบพันธุ์เหล่านี้มักจะได้รับการอนุรักษ์ไว้อย่างเด่นชัดในฟอสซิล และเป็นหัวข้อของการตีความต่างๆ เกี่ยวกับการสืบพันธุ์และภาวะเพศสภาพที่แตกต่างกันของยูริปเทอริด[ 45 ]
รยางค์ประเภท A โดยทั่วไปจะยาวกว่ารยางค์ประเภท B ในบางสกุล รยางค์ประเภทนี้จะแบ่งออกเป็นจำนวนส่วนที่แตกต่างกัน เช่น ในสกุล Eurypterusรยางค์ประเภท A จะแบ่งออกเป็นสามส่วน แต่รยางค์ประเภท B จะแบ่งออกเป็นเพียงสองส่วน[ 46 ]การแบ่งรยางค์อวัยวะสืบพันธุ์เช่นนี้เป็นเรื่องปกติในยูริปเทอริด แต่จำนวนส่วนนั้นไม่เหมือนกันทั้งหมด ตัวอย่างเช่น รยางค์ทั้งสองประเภทในวงศ์ Pterygotidae นั้นไม่มีการแบ่งส่วน[ 47 ]รยางค์ประเภท A ยังมีหนามโค้งสองอันที่เรียกว่าfurca (แปลว่า 'ส้อม' ในภาษาละติน) การมีfurcaในรยางค์ประเภท B ก็เป็นไปได้เช่นกัน และโครงสร้างนี้อาจแสดงถึงปลายรยางค์ที่ยังไม่เชื่อมติดกัน อวัยวะชุดหนึ่งที่อยู่ระหว่าง พื้นผิว ด้านบนและด้านล่างของBlattfüsseที่เกี่ยวข้องกับรยางค์ประเภท A นั้นถูกอธิบายตามธรรมเนียมว่าเป็น "อวัยวะรูปท่อ" หรือ "อวัยวะรูปเขา" อวัยวะเหล่านี้มักถูกตีความว่าเป็นสเปิร์มมาเทกา (อวัยวะสำหรับเก็บสเปิร์ม ) แม้ว่าหน้าที่นี้ยังไม่ได้รับการพิสูจน์อย่างแน่ชัดก็ตาม[ 48 ]ในอาร์โทรพอด สเปิร์มมาเทกาใช้สำหรับเก็บสเปิร์มมาโทฟอร์ที่ได้รับจากตัวผู้ ซึ่งหมายความว่าระยางค์ประเภท A เป็นลักษณะของตัวเมีย และระยางค์ประเภท B เป็นลักษณะของตัวผู้[ 21 ]หลักฐานเพิ่มเติมที่แสดงว่าระยางค์ประเภท A เป็นลักษณะของตัวเมียของระยางค์อวัยวะสืบพันธุ์ มาจากโครงสร้างที่ซับซ้อนกว่า (ซึ่งเป็นแนวโน้มทั่วไปของอวัยวะสืบพันธุ์ตัวเมียของอาร์โทรพอด) เป็นไปได้ว่าความยาวที่มากกว่าของระยางค์ประเภท A หมายความว่ามันถูกใช้เป็นอวัยวะวางไข่ (ใช้สำหรับวางไข่) [ 49 ]ระยางค์อวัยวะสืบพันธุ์ประเภทต่างๆ ไม่จำเป็นต้องเป็นคุณลักษณะเดียวที่แยกแยะเพศของยูริปเทอริดได้ ขึ้นอยู่กับสกุลและชนิดที่กล่าวถึง คุณลักษณะอื่นๆ เช่น ขนาด ปริมาณการตกแต่ง และความกว้างตามสัดส่วนของร่างกาย อาจเป็นผลมาจากความแตกต่างทางเพศ[ 6 ]โดยทั่วไป ยูริปเทอริดที่มีระยางค์ประเภท B (ตัวผู้) ดูเหมือนจะมีสัดส่วนที่กว้างกว่ายูริปเทอริดที่มีระยางค์ประเภท A (ตัวเมีย) ในสกุลเดียวกัน[ 50 ]

หน้าที่หลักของระยางค์ประเภท A ที่ยาวและสันนิษฐานว่าเป็นของตัวเมีย น่าจะทำหน้าที่รับสเปิร์มมาโทฟอร์จากพื้นผิวเข้าไปในระบบสืบพันธุ์มากกว่าที่จะทำหน้าที่เป็นอวัยวะวางไข่ เนื่องจากอวัยวะวางไข่ของอาร์โทรพอดโดยทั่วไปจะยาวกว่าระยางค์ประเภท A ของยูริปเทอริด การหมุนด้านข้างของฝาปิดจะทำให้สามารถลดระยางค์ลงจากตัวได้ เนื่องจากลักษณะการซ้อนทับกันของแผ่นต่างๆ ในตำแหน่งนั้น ระยางค์จึงไม่สามารถเคลื่อนที่ได้หากไม่มีการหดตัวของกล้ามเนื้อรอบๆ ฝาปิด มันจะถูกยึดไว้ในตำแหน่งเดิมเมื่อไม่ได้ใช้งานส่วนที่ยื่นออกมาของระยางค์ประเภท A อาจช่วยในการเปิดสเปิร์มมาโทฟอร์เพื่อปล่อยสเปิร์มอิสระภายในออกมาให้ดูดซับ อวัยวะที่เรียกว่า "อวัยวะเขา" ซึ่งอาจเป็นสเปิร์มมาเทกา เชื่อว่าเชื่อมต่อโดยตรงกับระยางค์ผ่านทางท่อ แต่ท่อที่สันนิษฐานเหล่านี้ยังไม่ได้รับการเก็บรักษาไว้ในวัสดุฟอสซิลที่มีอยู่[ 51 ]
ส่วนประกอบประเภท B ซึ่งสันนิษฐานว่าเป็นของเพศผู้ จะผลิต จัดเก็บ และอาจสร้างสเปิร์มมาโทฟอร์ในโครงสร้างรูปหัวใจบนพื้นผิวด้านหลังของส่วนประกอบ ช่องเปิดอวัยวะสืบพันธุ์ที่กว้างจะช่วยให้สามารถปล่อยสเปิร์มมาโทฟอร์จำนวนมากได้ในคราวเดียวฟอร์กา ที่ยาว ซึ่งเกี่ยวข้องกับส่วนประกอบประเภท B อาจสามารถลดระดับลงได้เหมือนส่วนประกอบประเภท A และอาจใช้ในการตรวจจับว่าพื้นผิวเหมาะสมสำหรับการวางสเปิร์มมาโทฟอร์หรือไม่[ 52 ]
ประวัติวิวัฒนาการ
ต้นกำเนิด

จนกระทั่งปี 1882 ยังไม่มีการค้นพบยูริปเทอริดจากยุคก่อนไซลูเรียน การค้นพบร่วมสมัยตั้งแต่ปี 1880 เป็นต้นมาได้ขยายความรู้เกี่ยวกับยูริปเทอริดยุคแรกจากยุคออร์โดวิเชียน[ 53 ]ยูริปเทอริดที่เก่าแก่ที่สุด (ในอดีต) ที่รู้จักในปัจจุบันคือเมกะโลกราปทิดเพนเทคอปเทอรัสซึ่งมีอายุย้อนไปถึง ยุค ดาร์ริวิเลียนของยุคออร์โดวิเชียนตอนกลาง เมื่อ 467.3 ล้านปีก่อน [ 54 ] อย่างไรก็ตามส่วนประกอบจากชั้นหินเฟซูอาตา ที่เก่ากว่านั้น ได้รับการอธิบายว่าเป็นสายพันธุ์ ? Carcinosoma auroraeซึ่งเป็นหลักฐานที่ไม่อาจปฏิเสธได้ชิ้นแรกของยูริปเทอริดในยุคเทรมาโดเซียน[ 55 ]
Pentecopterusเป็นยูริปเทอริดที่มีวิวัฒนาการค่อนข้างสูง เป็นส่วนหนึ่งของวงศ์เมกะโลกราปทิดภายในวงศ์ใหญ่คาร์ซิโนโซมาทอยด์ เคียงข้างกับคาร์ซิโนโซมาออโรรา ซึ่งเป็น คา ร์ซิโนโซมาทอยด์ ตำแหน่งวิวัฒนาการของพวกมันบ่งชี้ว่ากลุ่มยูริปเทอริดส่วนใหญ่ อย่างน้อยภายในอันดับย่อยยูริปเทอรีน ได้ถูกก่อตั้งขึ้นแล้วในช่วงเวลานี้ในยุคออร์โดวิเชียนตอนต้น[ 56 ]ยูริปเทอริดสไตโลนูรีนที่เก่าแก่ที่สุดที่รู้จัก คือ Brachyopterus [ 11 ]ซึ่งมีอายุอยู่ในยุคออร์โดวิเชียนตอนกลางเช่นกัน การมีสมาชิกของทั้งสองอันดับย่อยบ่งชี้ว่ายูริปเทอริดต้นกำเนิดดั้งเดิมน่าจะมีมาก่อนพวกมัน แม้ว่าสิ่งเหล่านี้จะยังไม่เป็นที่รู้จักในบันทึกฟอสซิลก็ตาม การมีกลุ่มยูริปเทอริดหลายกลุ่มในช่วงยุคออร์โดวิเชียนตอนกลางบ่งชี้ว่ายูริปเทอริดมีต้นกำเนิดในช่วงยุคแคมเบรียน[ 55 ] [ 56 ]
ด้วยเหตุนี้ เวลากำเนิดที่แน่นอนของยูริปเทอริดจึงยังไม่เป็นที่ทราบแน่ชัด แม้ว่าฟอสซิลที่เรียกว่า "ยูริปเทอริดดั้งเดิม" จะได้รับการอธิบายเป็นครั้งคราวจากแหล่งสะสมของยุคแคมเบรียนหรือแม้แต่ยุคพรีแคม เบรียน [ 57 ] แต่ในปัจจุบันฟอสซิลเหล่านั้นไม่ได้รับการยอมรับว่าเป็นยูริปเทอริด และบางครั้งก็ไม่ได้รับการ ยอมรับว่าเป็นรูปแบบที่เกี่ยวข้องด้วยซ้ำ สัตว์บางชนิดที่เคยถูกมองว่าเป็นยูริปเทอริดดั้งเดิม เช่น สกุลStrabopsจากยุคแคมเบรียนของมิสซูรี [ 58 ]ปัจจุบันถูกจัดประเภทเป็นอะกลาสพิดิดหรือสตรโบพิด อะกลาสพิดิดซึ่งครั้งหนึ่งเคยถูกมองว่าเป็นคีลิเซอเรตดั้งเดิม ปัจจุบันถูกมองว่าเป็นกลุ่มที่มีความสัมพันธ์ใกล้ชิดกับไทรโลไบต์มากกว่า[ 59 ]
บันทึกฟอสซิลของยูริปเทอริดในยุคออร์โดวิเชียนค่อนข้างไม่สมบูรณ์ ยูริปเทอริดส่วนใหญ่ที่เคยมีรายงานว่ารู้จักจากยุคออร์โดวิเชียนนั้นได้รับการพิสูจน์แล้วว่าเป็นการระบุผิดพลาดหรือฟอสซิลปลอมปัจจุบันมีเพียง 11 ชนิดเท่านั้นที่สามารถระบุได้อย่างมั่นใจว่าเป็นตัวแทนของยูริปเทอริดในยุคออร์โดวิเชียน กลุ่มอนุกรมวิธานเหล่านี้แบ่งออกเป็นสองประเภททางนิเวศวิทยาที่แตกต่างกัน ได้แก่ สัตว์นักล่าขนาดใหญ่และว่องไวจากทวีปโบราณลอเรนเทียและสัตว์ที่อาศัยอยู่บนพื้นทะเล และสัตว์พื้นฐานจากทวีปอะวาโลเนีย และกอนด์วานา[ 54 ]สัตว์นักล่าจากลอเรนเทีย ซึ่งจัดอยู่ในวงศ์ Megalograptidae (ประกอบด้วยสกุลEchinognathus , MegalograptusและPentecopterus ) น่าจะเป็นตัวแทนของกลุ่มยูริปเทอริดกลุ่มแรกที่ประสบความสำเร็จอย่างแท้จริง โดยมีการแพร่กระจายเล็กน้อยในช่วงปลายยุคออร์โดวิเชียน[ 60 ]
ไซลูเรียน

ยูริปเทอริดมีความหลากหลายและอุดมสมบูรณ์มากที่สุดระหว่างยุคไซลูเรียนตอนกลางและยุคเดโวเนียนตอนต้น โดยมีความหลากหลายสูงสุดในช่วงยุคพริโดลีซึ่งเกิดขึ้นเมื่อ 423 ถึง 419.2 ล้านปีก่อน ในช่วงปลายยุคไซลูเรียน[ 20 ]ความหลากหลายสูงสุดนี้ได้รับการยอมรับมาตั้งแต่ต้นศตวรรษที่ 20 โดยจากจำนวนยูริปเทอริดประมาณ 150 ชนิดที่รู้จักในปี พ.ศ. 2459 มากกว่าครึ่งหนึ่งมาจากยุคไซลูเรียน และหนึ่งในสามมาจากยุคไซลูเรียนตอนปลายเพียงอย่างเดียว[ 53 ]
แม้ว่ายูริปเทอริดกลุ่มสไตโลนูรีนโดยทั่วไปจะยังคงหายากและมีจำนวนน้อย เช่นเดียวกับในช่วงยุคออร์โดวิเชียนก่อนหน้านี้ แต่ยูริปเทอริดกลุ่มยูริปเทอรีนกลับมีความหลากหลายและจำนวนเพิ่มขึ้นอย่างรวดเร็ว[ 61 ]ในแหล่งฟอสซิลยุคไซลูเรียนส่วนใหญ่ ยูริปเทอริดกลุ่มยูริปเทอรีนคิดเป็น 90% ของยูริปเทอริดทั้งหมดที่มีอยู่[ 62 ]แม้ว่าบางกลุ่มอาจมีอยู่แล้วตั้งแต่ปลายยุคออร์โดวิเชียน (เพียงแต่ยังไม่ปรากฏในบันทึกฟอสซิลจนถึงปัจจุบัน) [ 56 ]แต่กลุ่มยูริปเทอริดส่วนใหญ่ถูกบันทึกไว้เป็นครั้งแรกในชั้นหินยุคไซลูเรียน ซึ่งรวมถึงทั้งกลุ่มสไตโลนูรีน เช่นStylonuroidea , KokomopteroideaและMycteropoideaรวมถึงกลุ่มยูริปเทอรีน เช่น Pterygotioidea, Eurypteroidea และWaeringopteroidea [ 63 ]
ยูริปเทอริดที่ประสบความสำเร็จมากที่สุดคือยูริปเทอรัส ในช่วงกลางถึงปลาย ยุคไซลูเรียน ซึ่งเป็นสัตว์กินเนื้อทั่วไป มีแนวโน้มที่จะล่าเหยื่อหรือกินซากสัตว์เท่าๆ กัน เชื่อกันว่าพวกมันล่าสัตว์ไม่มีกระดูกสันหลังขนาดเล็กและอ่อนนุ่มเป็นหลัก เช่น หนอน [ 64 ] สปีชีส์ในสกุลนี้( ซึ่งสปีชีส์ที่พบมากที่สุดคือสปีชีส์ต้นแบบE. remipes ) คิดเป็นมากกว่า 90% (อาจมากถึง 95%) ของตัวอย่างฟอสซิลยูริปเทอริดที่รู้จักทั้งหมด[ 62 ]แม้จะมีจำนวนมาก แต่ ยูริปเทอรัสเป็นที่รู้จักในช่วงเวลาที่ค่อนข้างสั้น โดยปรากฏตัวครั้งแรกในช่วงปลายยุคแลนโดเวอรี (ประมาณ 432 ล้านปีก่อน) และสูญพันธุ์ไปเมื่อสิ้นสุดยุคพริโดลี[ 65 ] Eurypterusยังถูกจำกัดอยู่ในทวีปยูราเมริกา (ประกอบด้วย ทวีปเขต ร้อน Avalonia, Balticaและ Laurentia) ซึ่งถูกสกุลนี้เข้ามาตั้งถิ่นฐานอย่างสมบูรณ์ในช่วงที่ทวีปนี้รวมตัวกัน และไม่สามารถข้ามมหาสมุทรอันกว้างใหญ่ที่แยกทวีปนี้ออกจากส่วนอื่นๆ ของโลก เช่น มหาทวีปทางใต้ Gondwana ได้ ดังนั้นEurypterusจึงถูกจำกัดทางภูมิศาสตร์อยู่เฉพาะบริเวณชายฝั่งและทะเลภายในแผ่นดินที่ตื้นของยูราเมริกา[ 62 ] [ 66 ]
ในช่วงปลายยุคไซลูเรียน สัตว์ปีกดึกดำบรรพ์กลุ่มยูริปเทอริด (Pterygotid eurypterids) ซึ่งมีขนาดใหญ่และมีลักษณะเฉพาะ พร้อมด้วยการปรับตัวใหม่หลายอย่าง เช่น เทลสัน (telson) ขนาดใหญ่และแบนที่สามารถใช้เป็นหางเสือได้ และคีลิเซรา (chelicerae) ขนาดใหญ่และมีลักษณะเฉพาะ พร้อมก้ามหนีบที่ขยายใหญ่ขึ้นสำหรับจับ (และอาจฆ่าในบางกรณี) เหยื่อได้ ปรากฏขึ้น[ 7 ] [ 8 ] แม้ว่าสมาชิกที่ใหญ่ที่สุดของวงศ์นี้จะปรากฏขึ้นใน ยุคเดโวเนียนแต่สัตว์ปีกดึกดำบรรพ์ขนาดใหญ่สองเมตร (6.5+ ฟุต) เช่นAcutiramus ก็มีอยู่แล้วในช่วงปลายยุคไซลูเรียน [ 14 ]นิเวศวิทยาของพวกมันมีตั้งแต่พฤติกรรมการล่าเหยื่อทั่วไปไปจนถึงการล่าเหยื่อแบบซุ่มโจมตีและบางชนิด เช่นPterygotusเอง เป็นผู้ล่าสูงสุดที่กระตือรือร้นในระบบนิเวศทางทะเลของยุคไซลูเรียนตอนปลาย[ 67 ]เห็นได้ชัดว่าสัตว์ปีกดึกดำบรรพ์เหล่านี้สามารถข้ามมหาสมุทรได้ กลายเป็นหนึ่งในสองกลุ่มยูริปเทอริดเท่านั้นที่มี การกระจายตัว ไปทั่วโลก[ 68 ]
ยุคดีโวเนียน

แม้ว่ายูริปเทอริดจะยังคงมีจำนวนมากและมีความหลากหลายในช่วงต้นยุคดีโวเนียน (ตัวอย่างเช่น นำไปสู่การวิวัฒนาการของเจคเคโลปเทอรัส ซึ่งเป็นสัตว์ขาปล้องที่ใหญ่ที่สุด) แต่กลุ่มนี้ก็เป็นหนึ่งในหลายกลุ่มที่ได้รับผลกระทบอย่างหนักจากการสูญพันธุ์ในช่วงปลายยุคดีโวเนียนเหตุการณ์การสูญพันธุ์ดังกล่าวเป็นที่ทราบกันว่าส่งผลกระทบต่อสิ่งมีชีวิตในทะเลเท่านั้น (โดยเฉพาะไทรโลไบต์แบรคิโอพอดและ สิ่งมีชีวิตที่สร้าง แนวปะการัง ) ทำให้ความอุดมสมบูรณ์และความหลากหลายที่เคยมีอยู่ในยูริปเทอริดลดลงอย่างมาก[ 69 ]
ความหลากหลายที่ลดลงอย่างมากได้เริ่มขึ้นแล้วในช่วงต้นยุคดีโวเนียน และยูริปเทอริดก็หายากในสภาพแวดล้อมทางทะเลในช่วงปลายยุคดีโวเนียน ในช่วง ยุค ฟราสเนียนมีสี่วงศ์สูญพันธุ์ไป และในยุคฟาเมนเนียน ตอนปลาย มีอีกห้าวงศ์สูญพันธุ์ไป[ 69 ]เนื่องจากกลุ่มสิ่งมีชีวิตในทะเลได้รับผลกระทบมากที่สุด ยูริปเทอริดจึงได้รับผลกระทบเป็นหลักภายในอันดับย่อยยูริปเทอรีน มีเพียงกลุ่มเดียวของสไตโลนูรีน (วงศ์Parastylonuridae ) ที่สูญพันธุ์ไปในช่วงต้นยุคดีโวเนียน มีเพียงสองวงศ์ของยูริปเทอรีนเท่านั้นที่รอดชีวิตมาได้จนถึงปลายยุคดีโวเนียน ( Adelophthalmidaeและ Waeringopteridae) ยูริปเทอรีนประสบกับการลดลงครั้งใหญ่ที่สุดในช่วงต้นยุคดีโวเนียน ซึ่งความหลากหลายของพวกมันหายไปมากกว่า 50% ในเวลาเพียง 10 ล้านปี ในทางกลับกัน Stylonurines ยังคงดำรงอยู่ตลอดช่วงเวลาดังกล่าวด้วยความหลากหลายและความอุดมสมบูรณ์ที่ค่อนข้างคงที่ แต่ได้รับผลกระทบในช่วงปลายยุคดีโวเนียน เมื่อกลุ่มเก่าๆ หลายกลุ่มถูกแทนที่ด้วยรูปแบบใหม่ในวงศ์ Mycteroptidae และ Hibbertopteridae [ 70 ]
เป็นไปได้ว่ารูปแบบการสูญพันธุ์ครั้งใหญ่ที่พบในอันดับย่อยยูริปเทอรีนนั้นเกี่ยวข้องกับการเกิดขึ้นของปลาที่มีวิวัฒนาการมากขึ้น การลดลงของยูริปเทอรีนเริ่มต้นขึ้น ณ จุดที่ปลาไม่มีขากรรไกรเริ่มมีวิวัฒนาการมากขึ้น และตรงกับการเกิดขึ้นของ ปลา แพลโคเดอร์ม (ปลาเกราะ) ทั้งในอเมริกาเหนือและยุโรป[ 71 ]
สไตโลนูรีนจากตระกูลฮิบเบอร์ทอปเทอริดและไมคเทอรอปทิดที่ยังมีชีวิตอยู่ได้หลีกเลี่ยงการแข่งขันกับปลาโดยสิ้นเชิงด้วยการวิวัฒนาการไปสู่นิเวศวิทยาเฉพาะถิ่นใหม่และแตกต่างออกไป ตระกูลเหล่านี้ประสบกับการแพร่กระจายและการแตกแขนงออกไปในช่วงปลายยุคดีโวเนียนและต้นยุคคาร์บอนิเฟอรัส ซึ่งเป็นการแพร่กระจายครั้งสุดท้ายในกลุ่มยูริปเทอริด ทำให้เกิดรูปแบบใหม่หลายรูปแบบที่สามารถ "กินอาหารแบบกวาด" (กวาดไปทั่วพื้นผิวเพื่อค้นหาเหยื่อ) [ 72 ]
ยุคคาร์บอนิเฟอรัสและยุคเพอร์เมียน

มีเพียงสามวงศ์ของยูริปเทอริด ได้แก่ Adelophthalmidae, Hibbertopteridae และ Mycteroptidae เท่านั้นที่รอดพ้นจากเหตุการณ์การสูญพันธุ์ทั้งหมด สันนิษฐานว่าสัตว์เหล่านี้ล้วนเป็นสัตว์น้ำจืด ซึ่งจะทำให้ยูริปเทอริดสูญพันธุ์ในสภาพแวดล้อมทางทะเล[ 69 ]และเมื่อผู้ล่าของยูริปเทอริดในทะเลหายไปซาร์โคปเทอริเจียนเช่นไรโซดอนต์จึงกลายเป็นผู้ล่าสูงสุดตัวใหม่ในสภาพแวดล้อมทางทะเล[ 71 ]อย่างไรก็ตาม การค้นพบใหม่ๆ หลายอย่างทำให้เกิดข้อสงสัยเกี่ยวกับเรื่องนี้ และชี้ให้เห็นว่ายูริปเทอริดเหล่านี้เป็นรูปแบบที่ทนต่อความเค็มได้ ซึ่งอาศัยอยู่ในสภาพแวดล้อมทางทะเลชายขอบ เช่น ปากแม่น้ำ ดินดอนสามเหลี่ยมปากแม่น้ำ ทะเลสาบ และบ่อชายฝั่ง ข้อโต้แย้งหนึ่งคือด้านบรรพชีวภูมิศาสตร์ การกระจายตัวของเทอริโกทอยด์ดูเหมือนจะต้องอาศัยการแพร่กระจายในมหาสมุทร[ 73 ]การทบทวน Adelophthalmoidea ครั้งล่าสุดยอมรับว่า "มีอิทธิพลทางทะเลมากกว่าที่เคยยอมรับกัน ในหลายส่วนที่พบ Adelophthalmus " [ 74 ]ในทำนองเดียวกัน การศึกษาเกี่ยวกับHibbertopterus ซึ่งเป็นยูริปเทอริด จากยุคคาร์บอนิเฟอรัสของนิวเม็กซิโก สรุปได้ว่าถิ่นที่อยู่ของยูริปเทอริดบางชนิด "อาจจำเป็นต้องได้รับการประเมินใหม่" [ 75 ]วงศ์ยูริปเทอรีนที่ยังคงอยู่รอดเพียงวงศ์เดียวคือ Adelophthalmidae มีเพียงสกุลเดียวคือAdelophthalmusส่วนฮิบเบอร์ท็อปเทอริด ไมคเทอรอปทิด และAdelophthalmusยังคงอยู่รอดมาจนถึงยุคเพอร์เมียน[ 76 ]
Adelophthalmusกลายเป็นยูริปเทอริดที่พบได้ทั่วไปมากที่สุดในยุคพาลีโอโซอิกตอนปลาย โดยมีจำนวนและความหลากหลายมากกว่าสไตโลนูรีนที่ยังมีชีวิตอยู่ และมีความหลากหลายมากขึ้นเมื่อไม่มียูริปเทอรีนชนิดอื่น[ 77 ]จาก 33 สปีชีส์ที่อ้างถึงAdelophthalmusมี 23 สปีชีส์ (69%) มาจากยุคคาร์บอนิเฟอรัสเพียงอย่างเดียว[ 78 ] [ 79 ]สกุลนี้มีความหลากหลายสูงสุดในยุคคาร์บอนิเฟอรัสตอนปลาย แม้ว่าAdelophthalmusจะแพร่หลายและมีตัวแทนอยู่ทั่วแผ่นดินใหญ่ในยุคเดโวเนียนตอนปลายแล้วก็ตาม การรวมตัวของแพนเจียเข้าเป็นมหาทวีปทั่วโลกในช่วงสองยุคสุดท้ายของยุคพาลีโอโซอิกทำให้Adelophthalmusมีการกระจายตัวเกือบจะทั่วโลก[ 62 ]
ในช่วงปลายยุคคาร์บอนิเฟอรัสและต้นยุคเพอร์เมียนAdelophthalmusแพร่หลาย โดยอาศัยอยู่เป็นหลักในสภาพแวดล้อมน้ำกร่อยและน้ำจืดที่อยู่ติดกับที่ราบชายฝั่ง สภาพแวดล้อมเหล่านี้ได้รับการรักษาไว้ด้วยสภาพภูมิอากาศที่เอื้ออำนวย แต่ไม่สามารถคงอยู่ได้เนื่องจากการเปลี่ยนแปลงสภาพภูมิอากาศอันเนื่องมาจากการก่อตัวของแพนเจีย ซึ่งเปลี่ยนแปลงรูปแบบการสะสมและการเจริญเติบโตของพืชทั่วโลก เมื่อที่อยู่อาศัยของพวกมันหายไป จำนวนของ Adelophthalmusก็ลดลงและสูญพันธุ์ไปแล้วในช่วง ยุค ลีโอนาร์เดียนของต้นยุคเพอร์เมียน[ 80 ]
Mycteroptids และ hibbertopterids ยังคงมีชีวิตอยู่ต่อไปอีกระยะหนึ่ง โดยมีสกุลหนึ่งของแต่ละกลุ่มที่รู้จักจากชั้นหินยุคเพอร์เมียน ได้แก่HastimimaและCampylocephalusตามลำดับ[ 81 ] Hastimimaสูญพันธุ์ไปในช่วงต้นยุคเพอร์เมียน[ 82 ]เช่นเดียวกับAdelophthalmusในขณะที่Campylocephalusยังคงมีชีวิตอยู่ได้นานกว่า เปลือกแข็งขนาดใหญ่ที่ไม่สมบูรณ์จากแหล่งสะสมยุคเพอร์เมียนในรัสเซียเป็นซากดึกดำบรรพ์เพียงชิ้นเดียวของสายพันธุ์C. permianusซึ่งอาจมีความยาวถึง 1.4 เมตร (4.6 ฟุต) ในขณะที่เดิมทีถือว่ามีอายุอยู่ในช่วงปลายยุคเพอร์เมียน[ 14 ]แหล่งข้อมูลอื่น ๆ แนะนำว่า ตัวอย่างนี้มีอายุอยู่ในช่วง Kungurian - Roadian (~283-267 ล้านปีก่อน) [ 83 ]ชิ้นส่วนของคิวติเคิลของสัตว์ขาปล้องจากปลายยุคเพอร์เมียน (ประมาณ 254-252 ล้านปีก่อน) ของออสเตรเลีย ซึ่งได้รับชื่อสายพันธุ์ว่าWoodwardopterus freemanorumนั้น ในตอนแรกถูกเสนอว่าเป็นยูริปเทอริดที่อายุน้อยที่สุดที่รู้จัก[ 83 ]ต่อมาในปี 2025 เจมส์ แลมส์เดลล์ ได้เสนอว่ารูปร่างของมันผิดปกติสำหรับยูริปเทอริด และอาจเป็นแมงป่องที่แท้จริงแทน แม้ว่าเขาจะเสนอว่าเป็นไปไม่ได้ที่จะจัดมันให้อยู่ในกลุ่มใดกลุ่มหนึ่งได้อย่างมั่นใจ เนื่องจากซากที่รู้จักในปัจจุบันนั้นเป็นเพียงชิ้นส่วน[ 84 ]ไม่พบยูริปเทอริดจากแหล่งฟอสซิลที่สูงกว่ายุคเพอร์เมียน ซึ่งบ่งชี้ว่ายูริปเทอริดสุดท้ายตายไปในเหตุการณ์การสูญพันธุ์ครั้งใหญ่ในช่วงปลายยุคเพอร์เมียน หรือในช่วงเวลาก่อนหน้านั้นไม่นาน เหตุการณ์การสูญพันธุ์ครั้งนี้ ซึ่งก็คือเหตุการณ์การสูญพันธุ์ในยุคเพอร์เมียน-ไทรแอสสิก (ซึ่งเกิดขึ้นเมื่อประมาณ 251.9 ล้านปีก่อน) ถือเป็นการ สูญพันธุ์ครั้งใหญ่ที่สุดที่บันทึกไว้ และทำให้กลุ่มสิ่งมีชีวิตในยุคพาลีโอโซอิกที่ประสบความสำเร็จอื่นๆ อีกมากมาย เช่น ไทรโลไบต์ สูญพันธุ์ไป[ 85 ]
ประวัติการศึกษา

ซากดึกดำบรรพ์ยูริปเทอริดชิ้นแรกที่รู้จักกันถูกค้นพบในหินยุคไซลูเรียนของรัฐนิวยอร์กซึ่งจนถึงทุกวันนี้ยังคงเป็นหนึ่งในแหล่งซากดึกดำบรรพ์ยูริปเทอริดที่อุดมสมบูรณ์ที่สุดแห่ง หนึ่ง ซามูเอล แอล. มิตชิลล์ ได้บรรยายลักษณะของ ซากดึกดำบรรพ์ที่ค้นพบใกล้กับเวสต์มอร์แลนด์ในเขตโอไนดาในปี 1818 เขาเข้าใจผิดว่าซากดึกดำบรรพ์นี้เป็นตัวอย่างของปลาสกุล Silurusซึ่งอาจเป็นเพราะลักษณะกระดองที่แปลก ประหลาดคล้าย ปลาแคทฟิช เจ็ดปีต่อมา ในปี 1825 เจมส์ อี. เดอเคย์ได้ตรวจสอบซากดึกดำบรรพ์และพบว่ามันเป็นของสัตว์ขาปล้องอย่างชัดเจน เขาคิดว่าซากดึกดำบรรพ์นี้ ซึ่งเขาตั้งชื่อว่าEurypterus remipesเป็นตัวแทนของสัตว์จำพวกครัสเตเชียนในอันดับBranchiopodaและเสนอแนะว่ามันอาจเป็นห่วงโซ่ที่ขาดหายไประหว่างไทรโลไบต์และแบรนชิโอพอดที่วิวัฒนาการสูงกว่า[ 86 ]ชื่อEurypterusมาจากภาษากรีกeurús ( εὐρύς 'กว้าง') และpteron ( πτερόν 'ปีก') [ 87 ]
ในปี พ.ศ. 2386 เฮอร์มันน์ บูร์ไมสเตอร์ได้ตีพิมพ์มุมมองของเขาเกี่ยวกับการจำแนกประเภทของไทรโลไบต์และความสัมพันธ์ของกลุ่มนี้กับสิ่งมีชีวิตอื่นๆ ทั้งที่ยังมีชีวิตอยู่และที่สูญพันธุ์ไปแล้ว ในงาน เขียนชื่อ Die Organisation der Trilobiten aus ihren lebenden Verwandten entwickeltเขาถือว่าไทรโลไบต์เป็นสัตว์จำพวกครัสเตเชียน เช่นเดียวกับที่ผู้เขียนคนก่อนๆ เคยกล่าวไว้ และจัดจำแนกพวกมันไว้ร่วมกับสิ่งที่เขาคิดว่าเป็นญาติใกล้ชิดที่สุด ได้แก่EurypterusและสกุลCytherinaภายในกลุ่มที่เขาตั้งชื่อว่า "Palaeadae" ภายใน Palaeadae บูร์ไมสเตอร์ได้ตั้งวงศ์ขึ้น 3 วงศ์ ได้แก่ "Trilobitae" (ประกอบด้วยไทรโลไบต์ทั้งหมด) "Cytherinidae" (ประกอบด้วยCytherina เท่านั้น ซึ่งปัจจุบันสัตว์ชนิดนี้ถูกมองว่าเป็นโอสทราโคด ) และEurypteridae (ประกอบด้วยEurypterusซึ่งในขณะนั้นมี 3 ชนิด) [ 88 ]
สกุลยูริปเทอริดที่สี่ที่ได้รับการอธิบาย (ต่อจากHibbertopterusในปี 1836 และCampylocephalusในปี 1838 ซึ่งไม่ได้รับการระบุว่าเป็นยูริปเทอริดจนกระทั่งภายหลัง) จากสกุลที่ยังคงถือว่าถูกต้องตามหลักอนุกรม วิธาน ในยุคปัจจุบัน คือPterygotus ( แปลว่า' ผู้มีปีก' ) ซึ่งได้รับการอธิบายโดยLouis Agassizในปี 1839 [ 89 ] Pterygotusมีขนาดใหญ่กว่าEurypterus อย่างมาก และเมื่อคนงานเหมืองหินใน สกอตแลนด์ค้นพบฟอสซิลชิ้นแรกพวกเขาเรียกฟอสซิลเหล่านั้นว่า " Seraphims " Agassiz คิดว่าฟอสซิลเหล่านั้นเป็นซากของปลาในตอนแรก และเพิ่งตระหนักว่าฟอสซิลเหล่านั้นเป็นซากของสัตว์ขาปล้องในอีกห้าปีต่อมาในปี 1844 [ 90 ]
ในปี พ.ศ. 2492 เฟรเดอริค แมคคอยได้จัดจำแนกPterygotusร่วมกับEurypterusและBelinurus (สกุลที่ปัจจุบันถือว่าเป็น xiphosuran) ภายใน Eurypteridae ของ Burmeister แมคคอยถือว่า Eurypteridae เป็นกลุ่มของสัตว์จำพวกครัสเตเชียนในอันดับEntomostracaซึ่งมีความสัมพันธ์ใกล้ชิดกับปูม้า[ 91 ]สกุลที่สี่Slimoniaซึ่งอิงจากซากดึกดำบรรพ์ที่เคยถูกกำหนดให้เป็นPterygotus ชนิดใหม่ ได้ถูกจัดให้อยู่ใน Eurypteridae ในปี พ.ศ. 2499 โดยเดวิดเพจ[ 92 ]

หนังสือ De Euryptero Remipede (1858) ของ Jan Nieszkowski นำเสนอคำอธิบายอย่างละเอียดเกี่ยวกับEurypterus fischeri (ปัจจุบันถือว่าเป็นชื่อพ้องกับEurypterus อีกชนิดหนึ่ง คือE. tetragonophthalmus ) ซึ่งร่วมกับหนังสือOn the Genus PterygotusโดยThomas Henry HuxleyและJohn William Salterและคำอธิบายอย่างละเอียดเกี่ยวกับยูริปเทอริดชนิดต่างๆ ในนิวยอร์กในเล่มที่ 3 ของPalaeontology of New York (1859) โดยJames Hallได้มีส่วนช่วยอย่างมากต่อความเข้าใจเกี่ยวกับความหลากหลายและชีววิทยาของยูริปเทอริด สิ่งพิมพ์เหล่านี้เป็นสิ่งพิมพ์แรกที่อธิบายกายวิภาคทั้งหมดของยูริปเทอริดอย่างครบถ้วน โดยระบุจำนวนระยางค์ส่วนหัวและจำนวนปล้องก่อนและหลังช่องท้อง ทั้ง Nieszkowski และ Hall ต่างยอมรับว่ายูริปเทอริดมีความสัมพันธ์ใกล้ชิดกับสัตว์ในกลุ่ม Chelicerata ในปัจจุบัน เช่น ปูม้า[ 93 ]
ในปี พ.ศ. 2408 เฮนรี วูดเวิร์ดได้บรรยายถึงสกุลStylonurus (ซึ่งได้รับการตั้งชื่อและวาดภาพ แต่ไม่ได้บรรยายอย่างละเอียดโดยเดวิด เพจ ในปี พ.ศ. 2499) และยกระดับ Eurypteridae ให้เป็นอันดับ ทำให้เกิด Eurypterida ขึ้นมาเป็นหน่วยอนุกรมวิธานอย่างที่เห็นในปัจจุบัน[ 94 ]ในงานAnatomy and Relations of the Eurypterida (พ.ศ. 2436) มัลคอล์ม ลอรีได้เพิ่มพูนความรู้และการอภิปรายเกี่ยวกับกายวิภาคศาสตร์และความสัมพันธ์ของยูริปเทอริดอย่างมาก เขาเน้นไปที่ความสัมพันธ์ระหว่างยูริปเทอริดด้วยกันเองและกับไทรโลไบต์ ครัสเตเชียน แมงป่อง แมงมุมชนิดอื่นๆ และปูม้า คำอธิบายของEurypterus fischeriโดย Gerhard Holm ในปี พ.ศ. 2449 นั้นละเอียดมากจนทำให้สายพันธุ์นี้กลายเป็นหนึ่งในสัตว์ที่สูญพันธุ์ไปแล้วที่เป็นที่รู้จักอย่างสมบูรณ์ที่สุด ความรู้เกี่ยวกับE. fischeriเทียบได้กับความรู้เกี่ยวกับญาติของมันในปัจจุบัน (เช่นปูม้าแอตแลนติก ) คำอธิบายนี้ยังช่วยยืนยันความสัมพันธ์ที่ใกล้ชิดระหว่างยูริปเทอริดและคีลิเซอเรตอื่นๆ โดยแสดงให้เห็นถึงความเหมือนกัน หลายประการ ระหว่างสองกลุ่มนี้[ 95 ]
ในปี ค.ศ. 1912 จอห์น เมสัน คลาร์กและรูดอล์ฟ รูเดมันน์ได้ตีพิมพ์หนังสือ ชื่อ The Eurypterida of New Yorkซึ่งกล่าวถึงสัตว์ในอันดับยูริปเทอริดทั้งหมดที่ค้นพบจากแหล่งฟอสซิลในบริเวณนั้น คลาร์กและรูเดมันน์ได้สร้างแผนภูมิวิวัฒนาการของยูริปเทอริดเป็นครั้งแรก โดยแบ่งอันดับนี้ออกเป็นสองวงศ์ คือ Eurypteridae (มีลักษณะเด่นคือตาเรียบ และรวมถึงEurypterus , Anthraconectes , Stylonurus , Eusarcus , Dolichopterus , OnychopterusและDrepanopterus ) และ Pterygotidae (มีลักษณะเด่นคือตาเป็นเหลี่ยม และรวมถึงPterygotus , Erettopterus , SlimoniaและHughmilleria ) ทั้งสองวงศ์นี้ถือว่าสืบเชื้อสายมาจากบรรพบุรุษร่วมกันคือStrabops [ 96 ]สอดคล้องกับผู้เขียนก่อนหน้านี้ คลาร์กและรูเดมันน์ยังสนับสนุนความสัมพันธ์ที่ใกล้ชิดระหว่างยูริปเทอริดและปูม้า (รวมกันอยู่ในชั้นเมโรสโตมาตา) แต่ยังได้อภิปรายสมมติฐานทางเลือกอื่นๆ เช่น ความสัมพันธ์ที่ใกล้ชิดกับแมงมุม[ 97 ]
การจำแนกประเภท


ในอดีต นักวิจัยส่วนใหญ่สันนิษฐานว่ายูริปเทอริดและซิโฟซูแรน (เช่น ปูม้าแอตแลนติกในปัจจุบัน) มีความสัมพันธ์ใกล้ชิดกัน ความคล้ายคลึงกันหลายประการสนับสนุนมุมมองนี้ เช่น ความสัมพันธ์กันของส่วนต่างๆ ของระยางค์และส่วนหัว นอกจากนี้ การมีระยางค์รูปแผ่นที่มี "ทางเดินเหงือก" บนระยางค์ของส่วนท้อง ( Blattfüssen ) ก็ถูกอ้างถึงตั้งแต่แรกว่าเป็นความคล้ายคลึงกันที่สำคัญ ในช่วงไม่กี่ทศวรรษสุดท้ายของศตวรรษที่สิบเก้า มีการค้นพบความคล้ายคลึงกันเพิ่มเติม เช่น โครงสร้างที่คล้ายคลึงกันของดวงตารวมของPterygotusและปูม้า (ซึ่งถือว่ามีความสำคัญอย่างยิ่ง เนื่องจากดวงตาของปูม้ามีโครงสร้างที่เกือบจะเป็นเอกลักษณ์) และความคล้ายคลึงกันในพัฒนาการของทั้งสองกลุ่ม[ 98 ]ความคล้ายคลึงกันทางด้านพัฒนาการเหล่านี้เห็นได้ชัดเจนที่สุดเมื่อศึกษาในระยะเนพิออนิก (ระยะพัฒนาการถัดจากระยะเอ็มบริโอนิกทันที) ในทั้งสองกลุ่ม ซึ่งในระยะนี้ทั้งซิโฟซูแรนและยูริปเทอริดจะมีกระดองที่ใหญ่กว่าตัวเต็มวัยอย่างเห็นได้ชัด มีความกว้างมากกว่า มีสันที่เห็นได้ชัดเจนตรงกลาง มีจำนวนปล้องน้อยกว่าซึ่งขาดการแบ่งแยก และมีเทลสันที่พัฒนาไม่เต็มที่[ 99 ]
เนื่องจากความคล้ายคลึงกันเหล่านี้ ซิโฟซูแรนและยูริปเทอริดจึงมักถูกรวมไว้ภายใต้ชั้นหรือชั้นย่อย เดียวกัน ที่เรียกว่า เมโรสโตมาตา (ซึ่งเฮนรี วูดเวิร์ดตั้งขึ้นเพื่อรวมทั้งสองกลุ่มในปี 1866) แม้ว่าซิโฟซูแรน (เช่นเดียวกับยูริปเทอริด) จะถูกมองว่าเป็นสัตว์จำพวกครัสเตเชียนในอดีตเนื่องจากระบบหายใจและวิถีชีวิตในน้ำ แต่สมมติฐานนี้ก็ถูกหักล้างหลังจากมีการค้นพบความคล้ายคลึงกันมากมายระหว่างปูม้าและแมงมุม[ 99 ]ผู้เขียนบางคน เช่นจอห์น สเตอร์ลิง คิงส์ลีย์ในปี 1894 ได้จัดประเภทเมโรสโตมาตาเป็นกลุ่มพี่น้องกับอาราคนิดภายใต้ชั้น "อะเซราตา" ภายในไฟลัมย่อย "แบรนคิอาตา" คนอื่นๆ เช่นเรย์ แลนเคสเตอร์ในปี 1909 ได้ไปไกลกว่านั้นและจัดประเภทเมโรสโตมาตาเป็นชั้นย่อยภายในอาราคนิด โดยยกระดับขึ้นเป็นชั้น[ 100 ]
ในปี พ.ศ. 2409 Ernst Haeckelได้จัดจำแนก Merostomata (ซึ่งแทบจะประกอบด้วย Eurypterida เท่านั้น) และ Xiphosura ไว้ในกลุ่มที่เขาตั้งชื่อว่า Gigantostraca ภายในกลุ่มครัสเตเชียน แม้ว่า Haeckel จะไม่ได้กำหนดลำดับชั้นทางอนุกรมวิธานใดๆ ให้กับกลุ่มนี้ แต่นักวิจัยรุ่นหลัง เช่น John Sterling Kinssley ได้ตีความว่ากลุ่มนี้เทียบเท่ากับลำดับชั้นย่อย เช่น Malacostraca และ Entomostraca [ 101 ]ในการวิจัยต่อมา Gigantostraca ได้รับการพิจารณาว่าเป็นคำพ้องความหมายกับ Merostomata (พบได้น้อย) และ Eurypterida เอง (พบได้บ่อยกว่า) [ 102 ] [ 103 ] การวิเคราะห์เชิงวิวัฒนาการ (ผลลัพธ์ที่นำเสนอในแผนภูมิวิวัฒนาการด้านล่าง) ที่ดำเนินการโดย James Lamsdell ในปี 2013 เกี่ยวกับความสัมพันธ์ภายใน Xiphosura และความสัมพันธ์กับกลุ่มอื่น ๆ ที่เกี่ยวข้องอย่างใกล้ชิด (รวมถึงยูริปเทอริด ซึ่งแสดงในการวิเคราะห์โดยสกุลEurypterus , Parastylonurus , RhenopterusและStoermeropterus ) สรุปได้ว่า Xiphosura ตามที่เข้าใจในปัจจุบัน เป็น กลุ่ม พาราไฟเลติก (กลุ่มที่มีบรรพบุรุษร่วมกันสุดท้ายแต่ไม่รวมลูกหลานทั้งหมดของบรรพบุรุษนี้) และดังนั้นจึงไม่ใช่กลุ่มวิวัฒนาการที่ถูกต้อง[ 104 ]ยูริปเทอริดถูกค้นพบว่ามีความสัมพันธ์ใกล้ชิดกับแมงมุมแทนที่จะเป็นซิโฟซูราน โดยก่อตัวเป็นกลุ่มSclerophorataภายในกลุ่มDekatriata (ประกอบด้วยสเคลอโรโฟเรตและชาสมาทาสพิดิด ) Lamsdell ตั้งข้อสังเกตว่าเป็นไปได้ที่ Dekatriata จะเป็นชื่อพ้องกับ Sclerophorata เนื่องจากระบบสืบพันธุ์ ซึ่งเป็นลักษณะเด่นหลักของ sclerophorates ยังไม่ได้รับการศึกษาอย่างละเอียดใน chasmataspidids Dekatriata เป็นส่วนหนึ่งของProsomapodaซึ่งเป็นกลุ่มที่รวมถึงXiphosurida (กลุ่ม xiphosuran ที่เป็นโมโนฟิเลติกเพียงกลุ่มเดียว) และสกุลต้นกำเนิดอื่นๆ[ 105 ]
ความสัมพันธ์ภายใน


การจำแนกประเภทภายในของยูริปเทอริดภายในกลุ่มยูริปเทอริดาขึ้นอยู่กับลักษณะที่กำหนดไว้ 11 ประการเป็นหลัก ลักษณะเหล่านี้ถูกนำมาใช้ตลอดประวัติศาสตร์การวิจัยยูริปเทอริดเพื่อสร้างกลุ่มและสกุล ลักษณะเหล่านี้ได้แก่ รูปร่างของโพรโซมา รูปร่างของเมตาสโตมา รูปร่างและตำแหน่งของดวงตา ประเภทของระยางค์โพรโซมา ประเภทของพายขาว่ายน้ำ โครงสร้างของดูบลูร์ (ขอบของโครงกระดูกภายนอกด้านหลัง) โครงสร้างของโอพิสโทโซมา โครงสร้างของระยางค์อวัยวะสืบพันธุ์ รูปร่างของเทลสัน และประเภทของเครื่องประดับที่มีอยู่ ควรสังเกตว่าลักษณะเหล่านี้ไม่ได้มีความสำคัญทางอนุกรมวิธานเท่ากันทั้งหมด[ 106 ]และไม่ได้ใช้ได้กับยูริปเทอริดทั้งหมดด้วย ยูริปเทอริดกลุ่มสไตโลนูรีนไม่มีพายขาว่ายน้ำเลย[ 20 ]ลักษณะบางอย่าง เช่น รูปร่างของโพรโซมาและเมตาสโตมา รวมถึงตำแหน่งและรูปร่างของดวงตา ถือว่ามีความสำคัญเฉพาะในการแยกแยะระหว่างสกุลต่างๆ เท่านั้น[ 107 ]วงศ์ใหญ่และวงศ์ส่วนใหญ่ถูกกำหนดโดยอาศัยสัณฐานวิทยาของรยางค์[ 108 ]
ลักษณะสำคัญที่สุดที่ใช้ในการจำแนกอนุกรมวิธานของยูริปเทอริดคือประเภทของระยางค์โปรโซมา เนื่องจากลักษณะนี้ใช้ในการกำหนดอันดับย่อยทั้งหมด กายวิภาคของขาโดยทั่วไปยังสามารถใช้ในการกำหนดวงศ์ใหญ่และวงศ์ได้อีกด้วย ในอดีต ก้ามคีลิเซราถือเป็นระยางค์ที่สำคัญที่สุดจากมุมมองทางอนุกรมวิธาน เนื่องจากมีเพียงสองประเภทหลักเท่านั้น คือ ประเภทยูริปเทอริดที่มีก้ามคีลิเซราขนาดเล็กและไม่มีฟัน และประเภทเทอริโกติดที่มีก้ามคีลิเซราขนาดใหญ่และมีฟัน การแบ่งแยกนี้ถูกนำมาใช้ในอดีตเพื่อแบ่งยูริปเทอริดาออกเป็นสองอันดับย่อย คือ ยูริปเทอรินา (ก้ามคีลิเซราขนาดเล็ก) และ "เทอริโกตินา" (ก้ามคีลิเซราขนาดใหญ่และแข็งแรง) [ 109 ]แผนการจำแนกประเภทนี้ไม่ได้ปราศจากปัญหา ในการแก้ไขอนุกรมวิธานของ Victor Tollerton ในปี 1989 เกี่ยวกับ Eurypterida ซึ่งได้รับการยอมรับเป็นอันดับย่อย Eurypterina และ Pterygotina กลุ่มย่อยของ eurypterids หลายกลุ่มที่ปัจจุบันได้รับการยอมรับว่าเป็น stylonurines (รวมถึง hibbertopterids และ mycteroptids) ได้รับการจัดประเภทใหม่เป็น non-eurypterids ในอันดับใหม่ที่แยกต่างหาก "Cyrtoctenida" โดยพิจารณาจากความไม่สอดคล้องกันที่รับรู้ได้ในส่วนประกอบ prosomal [ 110 ]
งานวิจัยสมัยใหม่สนับสนุนการจำแนกประเภทเป็นอันดับย่อย Eurypterina และ Stylonurina แทน โดยได้รับการสนับสนุนจากการวิเคราะห์ทางวิวัฒนาการ[ 111 ] [ 40 ]โดยเฉพาะอย่างยิ่ง ยูริปเทอริดกลุ่ม pterygotid มีความคล้ายคลึงกันหลายประการกับยูริปเทอริดกลุ่ม eurypterine ที่พัฒนาแล้ว เช่น adelophthalmids ดังนั้นจึงจัดอยู่ในกลุ่มสมาชิกที่พัฒนาแล้วของอันดับย่อยเดียวกัน[ 112 ]ใน Stylonurina ระยางค์คู่ที่หกเป็นขาเดินที่ยาวและเรียว และไม่มีกระดูกสันหลังที่ดัดแปลง (เรียกว่า podomere 7a) ในยูริปเทอริดส่วนใหญ่ใน Eurypterina ระยางค์คู่ที่หกจะขยายออกเป็นใบพายว่ายน้ำและมี podomere 7a เสมอ 75% ของสายพันธุ์ยูริปเทอริดเป็นยูริปเทอรีน และคิดเป็น 99% ของตัวอย่างฟอสซิลยูริปเทอริดทั้งหมด[ 20 ]ในบรรดากลุ่มยูริปเทอริดทั้งหมด กลุ่ม Pterygotioidea เป็นกลุ่มที่มีจำนวนชนิดมากที่สุด โดยมีมากกว่า 50 ชนิด กลุ่มที่มีจำนวนชนิดมากเป็นอันดับสองคือกลุ่ม Adelophthalmoidea โดยมีมากกว่า 40 ชนิด[ 62 ]
การวิเคราะห์อนุกรมวิธานและวิวัฒนาการของยูริปเทอริดส์อย่างสมบูรณ์ได้รับการตีพิมพ์ในปี 2025 โดย Lamsdell โดยอิงจากการวิจัยทั้งหมดของเขาเกี่ยวกับกลุ่มนี้ โดยยอมรับความสัมพันธ์ที่คล้ายคลึงกับการศึกษาครั้งก่อนๆ แต่ยังเพิ่มวงศ์ใหญ่ใหม่หนึ่งวงศ์ วงศ์ใหม่สิบวงศ์ สกุลใหม่หกสกุล และชนิดใหม่หนึ่งชนิด ทุกชนิดที่ได้รับการอธิบายได้รับการประเมิน แม้ว่าจะไม่ได้รวมไว้ในการวิเคราะห์ทั้งหมด โดยDorfopterusและMarsupipterusถูกแยกออกจาก Eurypterida เนื่องจากเป็นอาร์โทรพอดที่ไม่สามารถระบุชนิดได้ ความสัมพันธ์ระหว่าง Eurypterida ที่ได้สามารถดูได้ด้านล่าง ซึ่งแสดงอันดับ วงศ์ใหญ่ และวงศ์[ 113 ]
ดูเพิ่มเติม
- รายชื่อสกุลของยูริปเทอริด
- Nepidae —วงศ์แมลงที่ไม่เกี่ยวข้องกับวงศ์อื่น ๆ ซึ่งโดยทั่วไปรู้จักกันในชื่อ "แมงป่องน้ำ"
- Cottidae —วงศ์ปลาที่มีสมาชิกบางชนิดมีคำว่า "แมงป่องทะเล" อยู่ในชื่อสามัญ
- ประวัติชีวิต
หมายเหตุ
ลิงก์ภายนอก
- Eurypterids.co.uk – แหล่งข้อมูลและงานวิจัยเกี่ยวกับยูริปเทอริดทางออนไลน์
- eurypterid.net
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ยูริปเทอริด
ยูริปเทอริด (Eurypterids ) หรือที่เรียกกันอย่างไม่เป็นทางการว่าแมงป่องทะเลเป็นกลุ่มของสัตว์ขาปล้อง ในทะเลที่สูญพันธุ์ไปแล้ว ซึ่งอยู่ในอันดับยูริปเทอริดา (Eurypterida )
คำอธิบาย
เช่นเดียวกับ สัตว์ขาปล้อง อื่นๆยูริปเทอริดมี ลำ ตัวเป็นปล้อง และระยางค์ (แขนขา) ที่มีข้อต่อปกคลุม ด้วย คิว ติเคิล ซึ่งประกอบด้วย โปรตีน และ ไคติน เช่นเดียวกับใน เชลิเซอเรต อื่นๆ ลำตัว แบ่งออกเป็นสอง ส่วน (แท็กมาตา) คือ โพรโซมา ส่วนหน้า (หัว) และ โอพิสโทโซมา...
ขนาด
ยูริปเทอริดมีขนาดที่หลากหลายมาก ขึ้นอยู่กับปัจจัยต่างๆ เช่น วิถีชีวิต สภาพแวดล้อมในการดำรงชีวิต และ ความสัมพันธ์ทางอนุกรมวิธาน ขนาดประมาณ 100 เซนติเมตร (3.
การเคลื่อนที่
กลุ่มย่อยยูริปเทอริดสองกลุ่ม ได้แก่ ยูริปเทอรินาและ สไตโลนูรินา แตกต่างกันหลักๆ ที่รูปร่างของระยางค์คู่สุดท้าย ในสไตโลนูรินา ระยางค์นี้มีลักษณะเป็นขาเดินที่ยาวและเรียว ในขณะที่ในยูริปเทอรินา ขาจะถูกดัดแปลงและขยายออกเป็นพายสำหรับว่ายน้ำ [ 20 ]...