อ่าน 7 นาที
ห้องสมุดจีโนม
ห้องสมุด จีโนม คือชุดของชิ้นส่วน DNA ที่ซ้อนทับกันซึ่งประกอบกันเป็น DNA จีโนมทั้งหมด ของ สิ่งมีชีวิต หนึ่งเดียว DNA จะถูกเก็บไว้ในกลุ่มของ เวกเตอร์ ที่เหมือนกัน...
ห้องสมุดจีโนม
ห้องสมุดจีโนมคือชุดของชิ้นส่วน DNA ที่ซ้อนทับกันซึ่งประกอบกันเป็นDNA จีโนมทั้งหมด ของสิ่งมีชีวิต หนึ่งเดียว DNA จะถูกเก็บไว้ในกลุ่มของเวกเตอร์ ที่เหมือนกัน โดยแต่ละเวกเตอร์จะมีชิ้นส่วน DNA ที่แตกต่างกัน เพื่อสร้างห้องสมุดจีโนม DNA ของสิ่งมีชีวิตจะถูกสกัดจากเซลล์แล้วย่อยด้วยเอนไซม์ตัดจำเพาะเพื่อตัด DNA ออกเป็นชิ้นส่วนที่มีขนาดเฉพาะ จากนั้นชิ้นส่วนเหล่านี้จะถูกแทรกเข้าไปในเวกเตอร์โดยใช้DNA ไลเกส [ 1 ] ต่อมา DNA ของเวกเตอร์สามารถถูกดูดซึมโดยสิ่งมีชีวิตเจ้าบ้าน ซึ่งโดยทั่วไปคือประชากรของEscherichia coliหรือยีสต์โดยแต่ละเซลล์จะมีโมเลกุลเวกเตอร์เพียงหนึ่งโมเลกุล การใช้เซลล์เจ้าบ้านในการนำพาเวกเตอร์ทำให้สามารถขยาย และดึง โคลนเฉพาะจากห้องสมุดเพื่อการวิเคราะห์ ได้ง่าย [ 2 ]
มีเวกเตอร์หลายประเภทที่มีความจุในการแทรกที่แตกต่างกัน โดยทั่วไปแล้ว ไลบรารีที่สร้างจากสิ่งมีชีวิตที่มีจีโนม ขนาดใหญ่กว่า จะต้องใช้เวกเตอร์ที่มีตัวแทรกขนาดใหญ่กว่า ดังนั้นจึงต้องการโมเลกุลเวกเตอร์น้อยลงในการสร้างไลบรารี นักวิจัยสามารถเลือกเวกเตอร์โดยพิจารณาจากขนาดของตัวแทรกที่เหมาะสมเพื่อหาจำนวนโคลนที่ต้องการซึ่งจำเป็นสำหรับการครอบคลุมจีโนมทั้งหมด[ 3 ]
ห้องสมุดจีโนมมักใช้สำหรับ การ ประยุกต์ใช้การจัดลำดับมีบทบาทสำคัญในการจัดลำดับจีโนมทั้งหมดของสิ่งมีชีวิตหลายชนิด รวมถึงจีโนมของมนุษย์และสิ่งมีชีวิตต้นแบบ หลายชนิด [ 4 ] [ 5 ]
ประวัติศาสตร์
จีโนมที่ใช้ดีเอ็นเอเป็นครั้งแรก ที่ได้รับการจัดลำดับอย่างสมบูรณ์นั้นสำเร็จได้โดย เฟรเดอริก แซงเกอร์ผู้ได้รับรางวัลโนเบลสองครั้งในปี 1977 แซงเกอร์และทีมงานนักวิทยาศาสตร์ของเขาได้สร้างคลังของแบคทีริโอเฟจphi X 174เพื่อใช้ในการจัดลำดับดีเอ็นเอ[ 6 ]ความสำคัญของความสำเร็จนี้ส่งผลให้ความต้องการในการจัดลำดับจีโนมเพื่อการวิจัยการบำบัดด้วยยีน เพิ่มมากขึ้นเรื่อยๆ ปัจจุบันทีมงานสามารถจัดทำรายการโพลีมอร์ฟิซึมในจีโนมและตรวจสอบยีนที่อาจเป็นสาเหตุของโรคต่างๆ เช่นโรคพาร์กินสัน โรคอัลไซเมอร์โรคปลอกประสาทเสื่อมแข็ง โรคข้ออักเสบรูมาต อยด์ และโรคเบาหวานชนิดที่ 1 [ 7 ]สิ่งเหล่านี้เป็นผลมาจากความก้าวหน้าของการศึกษาความสัมพันธ์ทั่วทั้งจีโนมจากความสามารถในการสร้างและจัดลำดับคลังจีโนม ก่อนหน้านี้ การศึกษาการเชื่อมโยงและยีนเป้าหมายเป็นเพียงวิธีการบางส่วนเท่านั้น[ 8 ]
การสร้างคลังข้อมูลจีโนม
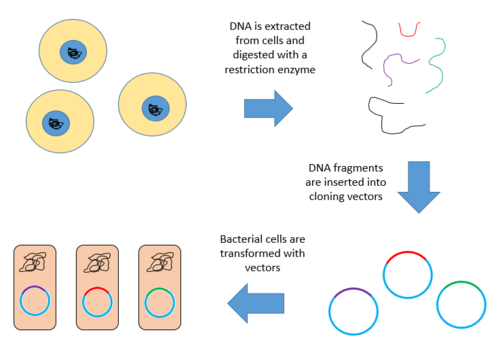
การสร้างไลบรารีจีโนมเกี่ยวข้องกับการสร้าง โมเลกุล DNA ลูกผสม จำนวนมาก DNAจีโนมของสิ่งมีชีวิตจะถูกสกัดและย่อยด้วยเอนไซม์ตัดจำเพาะสำหรับสิ่งมีชีวิตที่มีจีโนมขนาดเล็กมาก(~10 kb)ชิ้นส่วนที่ถูกย่อยสามารถแยกออกจากกันได้ด้วยเจลอิเล็กโทรโฟเรซิสจากนั้นชิ้นส่วนที่แยกออกมาสามารถตัดและโคลนลงในเวกเตอร์แยกกันได้ อย่างไรก็ตาม เมื่อจีโนมขนาดใหญ่ถูกย่อยด้วยเอนไซม์ตัดจำเพาะ จะมีชิ้นส่วนมากเกินไปที่จะตัดออกทีละชิ้น ชิ้นส่วนทั้งหมดจะต้องถูกโคลนร่วมกับเวกเตอร์ และการแยกโคลนสามารถเกิดขึ้นได้ในภายหลัง ไม่ว่าในกรณีใด ชิ้นส่วนจะถูกเชื่อมต่อเข้ากับเวกเตอร์ที่ถูกย่อยด้วยเอนไซม์ตัดจำเพาะเดียวกัน เวกเตอร์ที่มีชิ้นส่วน DNA จีโนมที่แทรกเข้าไปสามารถนำเข้าสู่สิ่งมีชีวิตเจ้าบ้านได้[ 1 ]
ด้านล่างนี้คือขั้นตอนการสร้างคลังข้อมูลจีโนมจากจีโนมขนาดใหญ่
- สกัดและทำให้ดีเอ็นเอบริสุทธิ์
- ย่อยดีเอ็นเอด้วยเอนไซม์ตัดจำเพาะ กระบวนการนี้จะสร้างชิ้นส่วนที่มีขนาดใกล้เคียงกัน โดยแต่ละชิ้นส่วนประกอบด้วยยีนหนึ่งตัวหรือมากกว่านั้น
- นำชิ้นส่วน DNA เข้าไปในเวกเตอร์ที่ถูกตัดด้วยเอนไซม์ตัดจำเพาะชนิดเดียวกัน จากนั้นใช้เอนไซม์ DNA ไลเกสเชื่อมต่อชิ้นส่วน DNA เข้ากับเวกเตอร์ วิธีนี้จะสร้างโมเลกุลลูกผสมจำนวนมาก
- โมเลกุลรีคอมบิแนนท์เหล่านี้จะถูกดูดซึมโดยแบคทีเรียเจ้าบ้านโดยการเปลี่ยนแปลงทำให้เกิดคลัง DNA [ 9 ] [ 10 ]
ด้านล่างนี้คือแผนภาพแสดงขั้นตอนต่างๆ ที่ได้กล่าวไว้ข้างต้น
การหาค่าไทเตอร์ของไลบรารี
หลังจากสร้างไลบรารีจีโนมด้วยเวกเตอร์ไวรัส เช่นแลมบ์ดาฟาจ แล้ว สามารถกำหนด ไทเตอร์ของไลบรารีได้ การคำนวณไทเตอร์ช่วยให้นักวิจัยสามารถประมาณจำนวนอนุภาคไวรัสที่ติดเชื้อที่สร้างขึ้นในไลบรารีได้สำเร็จ โดยใช้วิธีเจือจางไลบรารีเพื่อเปลี่ยนวัฒนธรรมของ E. coli ที่มีความเข้มข้นที่ทราบ จากนั้นนำวัฒนธรรมไปเพาะบนจานวุ้นและบ่มไว้ข้ามคืน นับจำนวนพลาคไวรัสและสามารถใช้คำนวณจำนวนอนุภาคไวรัสที่ติดเชื้อทั้งหมดในไลบรารีได้ เวกเตอร์ไวรัสส่วนใหญ่ยังมีเครื่องหมายที่ช่วยให้สามารถแยกแยะโคลนที่มีชิ้นส่วนแทรกออกจากโคลนที่ไม่มีชิ้นส่วนแทรกได้ ซึ่งช่วยให้นักวิจัยสามารถกำหนดเปอร์เซ็นต์ของอนุภาคไวรัสที่ติดเชื้อที่บรรจุชิ้นส่วนของไลบรารีได้จริง[ 11 ]
สามารถใช้วิธีการที่คล้ายกันในการหาค่าความเข้มข้นของไลบรารีจีโนมที่สร้างด้วยเวกเตอร์ที่ไม่ใช่ไวรัส เช่นพลาสมิดและBAC สามารถใช้ การเชื่อมต่อทดสอบของไลบรารีเพื่อแปลง E. coli จากนั้นจึงกระจายการแปลงลงบนจานวุ้นและบ่มไว้ข้ามคืน ค่าความเข้มข้นของการแปลงจะถูกกำหนดโดยการนับจำนวนโคโลนีที่ปรากฏบนจาน เวกเตอร์เหล่านี้โดยทั่วไปจะมีเครื่องหมายที่เลือกได้ซึ่งช่วยให้สามารถแยกแยะโคลนที่มีชิ้นส่วนแทรกออกจากโคลนที่ไม่มีได้ โดยการทำการทดสอบนี้ นักวิจัยยังสามารถกำหนดประสิทธิภาพของการเชื่อมต่อและปรับเปลี่ยนตามความจำเป็นเพื่อให้แน่ใจว่าพวกเขาได้รับจำนวนโคลนที่ต้องการสำหรับไลบรารี[ 12 ]
ห้องสมุดฉายภาพ

เพื่อแยกโคลนที่มีบริเวณที่สนใจออกจากไลบรารี ไลบรารีจะต้องได้รับการคัดกรอง ก่อน วิธีการคัดกรองวิธีหนึ่งคือการไฮบริดไดเซชัน เซลล์โฮสต์ที่แปลงสภาพแต่ละเซลล์ของไลบรารีจะมีเวกเตอร์เพียงตัวเดียวที่มีดีเอ็นเอแทรกเพียงชิ้นเดียว ไลบรารีทั้งหมดสามารถเพาะลงบนแผ่นกรองเหนืออาหารเลี้ยงเชื้อได้ แผ่นกรองและโคโลนีจะถูกเตรียมสำหรับการไฮบริดไดเซชัน จากนั้นจึงติดฉลากด้วยโพรบ [ 13 ] ดีเอ็นเอเป้าหมาย - ดีเอ็นเอแทรกที่สนใจ - สามารถระบุได้โดยการตรวจจับ เช่นออโตเรดิโอกราฟีเนื่องจากการไฮบริดไดเซชันกับโพรบดังที่เห็นด้านล่าง
วิธีการคัดกรองอีกวิธีหนึ่งคือการใช้ปฏิกิริยาลูกโซ่พอลิเมอเรส (PCR) ไลบรารีบางส่วนถูกจัดเก็บในรูปแบบของกลุ่มโคลน และการคัดกรองด้วย PCR เป็นวิธีที่มีประสิทธิภาพในการระบุกลุ่มที่มีโคลนเฉพาะ[ 2 ]
ประเภทของเวกเตอร์
ขนาด จีโนมแตกต่างกันไปในสิ่งมีชีวิตต่าง ๆ และต้องเลือกเวกเตอร์โคลน ให้เหมาะสม สำหรับจีโนมขนาดใหญ่ ควรเลือกเวกเตอร์ที่มีความจุสูงเพื่อให้ โคลน จำนวนน้อยเพียงพอต่อการครอบคลุมจีโนมทั้งหมด อย่างไรก็ตาม การระบุลักษณะของ อินเสิร์ตที่อยู่ในเวกเตอร์ที่มีความจุสูงกว่ามักจะทำได้ยากกว่า[ 3 ]
ด้านล่างนี้คือตารางแสดงเวกเตอร์หลายชนิดที่นิยมใช้สำหรับสร้างไลบรารีจีโนม และขนาดของชิ้นส่วนดีเอ็นเอที่แต่ละชนิดสามารถบรรจุได้โดยทั่วไป
| ประเภท เวกเตอร์ | ขนาด แทรก (หลายพันฐาน ) |
|---|---|
| พลาสมิด | มากถึง 10 |
| ฟาจแลมบ์ดา (λ) | มากถึง 25 |
| คอสมิด | สูงสุด 45 |
| แบคทีริโอเฟจ P1 | 70 ถึง 100 |
| โครโมโซมเทียม P1 (PACs) | 130 ถึง 150 |
| โครโมโซมเทียมของแบคทีเรีย (BACs) | 120 ถึง 300 |
| โครโมโซมเทียมของยีสต์ (YACs) | 250 ถึง 2000 |
พลาสมิด
พลาสมิด เป็นโมเลกุล DNAแบบวงกลมสองสายที่ใช้กันทั่วไปในการโคลนนิ่งระดับโมเลกุลโดยทั่วไปพลาสมิดมีความยาว 2 ถึง 4 กิโลเบสแพร์ (kb) และสามารถบรรจุสารแทรกได้ถึง 15kb พลาสมิดมีต้นกำเนิดการจำลองแบบที่ช่วยให้สามารถจำลองแบบภายในแบคทีเรียได้อย่างอิสระจากโครโมโซม ของโฮสต์ พลาสมิดมักมียีนต้านทานยาปฏิชีวนะที่ช่วยให้สามารถคัดเลือกเซลล์แบคทีเรียที่มีพลาสมิดได้ พลาสมิดจำนวนมากยังมียีนรายงานที่ช่วยให้นักวิจัยสามารถแยกแยะโคลนที่มีสารแทรกออกจากโคลนที่ไม่มีสารแทรกได้[ 3 ]
ฟาจแลมบ์ดา (λ)
ฟาจ λเป็นไวรัสดีเอ็นเอสายคู่ที่ติดเชื้อE. coliโครโมโซม λ มีความยาว 48.5 กิโลเบส และสามารถบรรจุชิ้นส่วนแทรกได้ถึง 25 กิโลเบส ชิ้นส่วนแทรกเหล่านี้จะแทนที่ลำดับไวรัสที่ไม่จำเป็นในโครโมโซม λ ในขณะที่ยีนที่จำเป็นสำหรับการสร้างอนุภาคไวรัสและการติดเชื้อยังคงอยู่ครบถ้วน ดีเอ็นเอชิ้นส่วนแทรกจะถูกจำลองแบบพร้อมกับดีเอ็นเอไวรัส ดังนั้นจึงถูกบรรจุรวมกันเป็นอนุภาคไวรัส อนุภาคเหล่านี้มีประสิทธิภาพสูงในการติดเชื้อและการเพิ่มจำนวน ส่งผลให้มีการผลิตโครโมโซม λ ลูกผสมมากขึ้น[ 3 ]อย่างไรก็ตาม เนื่องจากขนาดของชิ้นส่วนแทรกที่เล็กกว่า ไลบรารีที่สร้างด้วยฟาจ λ อาจต้องใช้โคลนจำนวนมากเพื่อให้ครอบคลุมจีโนมทั้งหมด[ 14 ]
คอสมิด
เวกเตอร์ คอสมิดเป็นพลาสมิดที่มีดีเอ็นเอของแบคทีริโอเฟจ λ ขนาดเล็กที่เรียกว่าลำดับคอส ลำดับนี้ทำให้คอสมิดสามารถบรรจุลงในอนุภาคแบคทีริโอเฟจ λ ได้ อนุภาคเหล่านี้ซึ่งมีคอสมิดแบบเส้นตรงอยู่ภายใน จะถูกนำเข้าสู่เซลล์โฮสต์โดยการถ่ายทอด เมื่ออยู่ภายในโฮสต์แล้ว คอสมิดจะกลายเป็นวงกลมด้วยความช่วยเหลือของ ดีเอ็นเอไลเกสของโฮสต์จากนั้นจึงทำหน้าที่เป็นพลาสมิด คอสมิดสามารถบรรจุสารแทรกที่มีขนาดได้ถึง 40kb [ 2 ]
เวกเตอร์แบคทีริโอเฟจ P1
เวกเตอร์แบค ทีริโอเฟจ P1สามารถบรรจุอินเสิร์ตขนาด 70 – 100kb ได้ โดยเริ่มต้นจากโมเลกุล DNA แบบเส้นตรงที่บรรจุอยู่ในอนุภาคแบคทีริโอเฟจ P1 อนุภาคเหล่านี้จะถูกฉีดเข้าไปในสายพันธุ์ E. coli ที่แสดงออกถึงCre recombinaseเวกเตอร์ P1 แบบเส้นตรงจะกลายเป็นวงกลมโดยการรีคอมบิเนชันระหว่างไซต์ loxP สองไซต์ในเวกเตอร์ เวกเตอร์ P1 โดยทั่วไปจะมียีนต้านทานยาปฏิชีวนะและเครื่องหมายการคัดเลือกเชิงบวกเพื่อแยกแยะโคลนที่มีอินเสิร์ตออกจากโคลนที่ไม่มี เวกเตอร์ P1 ยังมีรีพลิคอน พลาสมิด P1 ซึ่งทำให้มั่นใจได้ว่าจะมีเวกเตอร์เพียงสำเนาเดียวในเซลล์ อย่างไรก็ตาม ยังมีรีพลิคอน P1 ตัวที่สองที่เรียกว่ารีพลิคอนไลติก P1 ซึ่งถูกควบคุมโดยโปรโมเตอร์ ที่เหนี่ยวนำได้ โปรโมเตอร์นี้ช่วยให้สามารถขยายเวกเตอร์ ได้มากกว่าหนึ่งสำเนาต่อเซลล์ก่อนการสกัด DNA [ 2 ]

โครโมโซมเทียม P1
โครโมโซมเทียม P1 (PACs) มีคุณสมบัติทั้งของเวกเตอร์ P1 และโครโมโซมเทียมแบคทีเรีย (BACs) คล้ายกับเวกเตอร์ P1 พวกมันประกอบด้วยพลาสมิดและรีพลิคอนไลติกดังที่กล่าวไว้ข้างต้น แตกต่างจากเวกเตอร์ P1 ตรงที่พวกมันไม่จำเป็นต้องบรรจุลงในอนุภาคแบคทีริโอเฟจเพื่อการถ่ายทอด แต่จะถูกนำเข้าสู่ E. coli ในรูปของโมเลกุล DNA วงกลมผ่านอิเล็กโทรโพเรชันเช่นเดียวกับ BACs [ 2 ]นอกจากนี้ยังคล้ายกับ BACs การเตรียม PACs ค่อนข้างยากกว่าเนื่องจากมีต้นกำเนิดการจำลองแบบเพียงจุดเดียว[ 14 ]
โครโมโซมเทียมของแบคทีเรีย
โครโมโซมเทียมของแบคทีเรีย (BACs) เป็นโมเลกุล DNA วงกลม โดยทั่วไปมีความยาวประมาณ 7 กิโลเบส ซึ่งสามารถบรรจุชิ้นส่วนแทรกที่มีขนาดสูงสุด 300 กิโลเบส เวกเตอร์ BAC ประกอบด้วยรีพลิคอนที่ได้มาจากแฟกเตอร์ F ของ E. coli ซึ่งทำให้มั่นใจได้ว่าจะมีสำเนาเพียงหนึ่งชุดต่อเซลล์[ 4 ]เมื่อชิ้นส่วนแทรกถูกเชื่อมเข้ากับ BAC แล้ว BAC จะถูกนำเข้าสู่ สายพันธุ์ E. coli ที่ขาดความสามารถ ในการรวมตัวใหม่โดยการใช้กระแสไฟฟ้า เวกเตอร์ BAC ส่วนใหญ่มียีนต้านทานยาปฏิชีวนะและเครื่องหมายการคัดเลือกเชิงบวก[ 2 ]ภาพด้านขวาแสดงให้เห็นเวกเตอร์ BAC ที่ถูกตัดด้วยเอนไซม์จำกัด ตามด้วยการแทรก DNA ต่างประเทศที่ถูกเชื่อมต่อใหม่โดยไลเกส โดยรวมแล้ว เวกเตอร์นี้มีความเสถียรมาก แต่การเตรียมอาจทำได้ยากเนื่องจากมีต้นกำเนิดการจำลองแบบเพียงจุดเดียวเช่นเดียวกับ PACs [ 14 ]
โครโมโซมเทียมของยีสต์
โครโมโซมเทียมของยีสต์ (YACs) เป็นโมเลกุล DNA เชิงเส้นที่มีคุณสมบัติที่จำเป็นของโครโมโซมยีสต์ ที่แท้จริง รวมถึง เทโลเมียร์เซนโทรเมียร์และจุดเริ่มต้นของการจำลองแบบสามารถเชื่อมต่อชิ้นส่วน DNA ขนาดใหญ่เข้ากับตรงกลางของ YAC ได้ ทำให้มี "แขน" ของ YAC อยู่ทั้งสองด้านของชิ้นส่วนที่แทรกเข้าไป YAC ที่สร้างขึ้นใหม่จะถูกนำเข้าสู่ยีสต์โดยการแปลงสภาพ เครื่องหมายที่เลือกได้ที่มีอยู่ใน YAC ช่วยให้สามารถระบุทรานส์ฟอร์แมนต์ที่ประสบความสำเร็จได้ YAC สามารถบรรจุชิ้นส่วนได้ถึง 2000kb แต่ไลบรารี YAC ส่วนใหญ่มีชิ้นส่วนขนาด 250-400kb ในทางทฤษฎีแล้วไม่มีขีดจำกัดสูงสุดของขนาดชิ้นส่วนที่ YAC สามารถบรรจุได้ คุณภาพในการเตรียม DNA ที่ใช้สำหรับชิ้นส่วนที่แทรกจะเป็นตัวกำหนดขีดจำกัดขนาด[ 2 ]แง่มุมที่ท้าทายที่สุดของการใช้ YAC คือความจริงที่ว่าพวกมันมีแนวโน้มที่จะเกิดการจัดเรียงใหม่[ 14 ]
วิธีการเลือกเวกเตอร์
การเลือกเวกเตอร์จำเป็นต้องตรวจสอบให้แน่ใจว่าไลบรารีที่สร้างขึ้นเป็นตัวแทนของจีโนมทั้งหมด ชิ้นส่วนจีโนมใดๆ ที่ได้จากเอนไซม์จำกัดควรมีโอกาสเท่ากันที่จะอยู่ในไลบรารีเมื่อเทียบกับชิ้นส่วนอื่นๆ นอกจากนี้ โมเลกุลรีคอมบิแนนท์ควรมีชิ้นส่วนที่มีขนาดใหญ่พอที่จะทำให้ขนาดของไลบรารีสามารถจัดการได้อย่างสะดวก[ 14 ]โดยเฉพาะอย่างยิ่งสิ่งนี้ถูกกำหนดโดยจำนวนโคลนที่จำเป็นต้องมีในไลบรารี จำนวนโคลนที่จะได้ตัวอย่างของยีนทั้งหมดถูกกำหนดโดยขนาดของจีโนมของสิ่งมีชีวิตรวมถึงขนาดเฉลี่ยของชิ้นส่วน สิ่งนี้แสดงโดยสูตร (หรือที่รู้จักกันในชื่อสูตรคาร์บอนและคลาร์ก): [ 15 ]

ที่ไหน,
คือจำนวนรีคอมบิแนนท์ที่จำเป็น[ 16 ]

คือความน่าจะเป็นที่ต้องการว่าชิ้นส่วนใดๆ ในจีโนมจะปรากฏอย่างน้อยหนึ่งครั้งในไลบรารีที่สร้างขึ้น

คือสัดส่วนเศษส่วนของจีโนมในรีคอมบิแนนท์เดี่ยว

สามารถแสดงให้เห็นเพิ่มเติมได้ว่า:

ที่ไหน,
ขนาดของแผ่นแทรกคือ

ขนาดของจีโนม

ดังนั้น การเพิ่มขนาดของชิ้นส่วนแทรก (โดยการเลือกเวกเตอร์) จะทำให้ต้องใช้โคลนน้อยลงในการแสดงจีโนม สัดส่วนของขนาดชิ้นส่วนแทรกเทียบกับขนาดจีโนมแสดงถึงสัดส่วนของจีโนมแต่ละส่วนในโคลนเดียว[ 14 ]นี่คือสมการที่พิจารณาทุกส่วนแล้ว:

ตัวอย่างการเลือกเวกเตอร์
สูตรข้างต้นสามารถใช้เพื่อกำหนดระดับความเชื่อมั่น 99% ว่าลำดับทั้งหมดในจีโนมนั้นถูกแสดงโดยใช้เวกเตอร์ที่มีขนาดแทรก 20,000 คู่เบส (เช่น เวกเตอร์ของฟาจแลมบ์ดา) ขนาดจีโนมของสิ่งมีชีวิตในตัวอย่างนี้คือ 3,000 คู่เบส
![{\displaystyle N={\frac {ln(1-0.99)}{ln[1-{\frac {2.0\times 10^{4}basepairs}{3.0\times 10^{9}basepairs}}]}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/053b596ba7adfc402824b91ceae3dd24973b2fec)

โคลน

ดังนั้น จึงจำเป็นต้องใช้โคลนประมาณ 688,060 ตัว เพื่อให้มั่นใจได้ว่ามีความน่าจะเป็น 99% ที่ลำดับดีเอ็นเอที่กำหนดจากจีโนมขนาดสามพันล้านเบสแพร์นี้ จะปรากฏอยู่ในไลบรารีโดยใช้เวกเตอร์ที่มีขนาดแทรกสองหมื่นเบสแพร์
แอปพลิเคชัน
หลังจากสร้างไลบรารีแล้ว จีโนมของสิ่งมีชีวิตสามารถถูกจัดลำดับเพื่ออธิบายว่ายีนมีผลต่อสิ่งมีชีวิตอย่างไร หรือเพื่อเปรียบเทียบสิ่งมีชีวิตที่คล้ายคลึงกันในระดับจีโนมการศึกษาการเชื่อมโยงทั่วทั้งจีโนม ที่กล่าวถึงข้างต้น สามารถระบุยีนเป้าหมายที่มาจากลักษณะการทำงานหลายอย่าง ยีนสามารถแยกได้จากไลบรารีจีโนมและนำไปใช้กับสายเซลล์ของมนุษย์หรือแบบจำลองสัตว์เพื่อการวิจัยเพิ่มเติม[ 17 ]นอกจากนี้ การสร้างโคลนที่มีความแม่นยำสูงด้วยการแสดงจีโนมที่ถูกต้องและไม่มีปัญหาเรื่องความเสถียรจะช่วยได้เป็นอย่างดีในฐานะตัวกลางสำหรับการจัดลำดับแบบช็อตกันหรือการศึกษายีนทั้งหมดในการวิเคราะห์การทำงาน[ 10 ]
การจัดลำดับชั้น

การใช้งานหลักอย่างหนึ่งของไลบรารีจีโนมคือการจัดลำดับช็อตกันแบบลำดับชั้นซึ่งเรียกอีกอย่างว่าการจัดลำดับแบบท็อปดาวน์ แบบอิงแผนที่ หรือแบบโคลนต่อโคลน กลยุทธ์นี้ได้รับการพัฒนาขึ้นในช่วงทศวรรษ 1980 สำหรับการจัดลำดับจีโนมทั้งหมดก่อนที่จะมีเทคนิคการจัดลำดับแบบความเร็วสูง โคลนแต่ละตัวจากไลบรารีจีโนมสามารถถูกตัดเป็นชิ้นส่วนเล็กๆ ซึ่งโดยทั่วไปจะมีขนาด 500bp ถึง 1000bp ซึ่งจัดการได้ง่ายกว่าสำหรับการจัดลำดับ[ 4 ]เมื่อโคลนจากไลบรารีจีโนมได้รับการจัดลำดับแล้ว ลำดับนั้นสามารถนำมาใช้คัดกรองไลบรารีเพื่อหาโคลนอื่นๆ ที่มีชิ้นส่วนแทรกที่ทับซ้อนกับโคลนที่จัดลำดับแล้ว โคลนที่ทับซ้อนใหม่ใดๆ ก็สามารถจัดลำดับได้เพื่อสร้างคอนติ๊กเทคนิคนี้เรียกว่าการเดินโครโมโซมสามารถนำมาใช้เพื่อจัดลำดับโครโมโซมทั้งหมดได้[ 2 ]
การจัดลำดับจีโนมแบบช็อตกัน (Whole genome shotgun sequencing)เป็นอีกวิธีหนึ่งในการจัดลำดับจีโนมที่ไม่ต้องใช้ไลบรารีของเวกเตอร์ความจุสูง แต่จะใช้อัลกอริธึมคอมพิวเตอร์ในการประกอบลำดับสั้นๆ เพื่อครอบคลุมจีโนมทั้งหมด ด้วยเหตุนี้ ไลบรารีจีโนมจึงมักใช้ร่วมกับการจัดลำดับจีโนมแบบช็อตกัน สามารถสร้างแผนที่ความละเอียดสูงได้โดยการจัดลำดับปลายทั้งสองข้างของชิ้นส่วนแทรกจากโคลนหลายๆ ตัวในไลบรารีจีโนม แผนที่นี้ให้ลำดับที่มีระยะห่างที่ทราบ ซึ่งสามารถนำมาใช้ช่วยในการประกอบลำดับที่ได้จากการจัดลำดับแบบช็อตกัน[ 4 ]ลำดับจีโนมมนุษย์ ซึ่งได้รับการประกาศว่าสมบูรณ์ในปี 2546 ได้รับการประกอบโดยใช้ทั้งไลบรารี BAC และการจัดลำดับแบบช็อตกัน[ 18 ] [ 19 ]
การศึกษาความสัมพันธ์ทั่วทั้งจีโนม
การศึกษาการเชื่อมโยงทั่วทั้งจีโนมเป็นการประยุกต์ใช้ทั่วไปเพื่อค้นหาเป้าหมายยีนและโพลีมอร์ฟิซึมที่เฉพาะเจาะจงภายในเผ่าพันธุ์มนุษย์ อันที่จริง โครงการ International HapMap ถูกสร้างขึ้นผ่านความร่วมมือของนักวิทยาศาสตร์และหน่วยงานจากหลายประเทศเพื่อจัดทำแคตตาล็อกและใช้ข้อมูลนี้[ 20 ]เป้าหมายของโครงการนี้คือการเปรียบเทียบลำดับทางพันธุกรรมของบุคคลต่างๆ เพื่อชี้แจงความคล้ายคลึงและความแตกต่างภายในบริเวณโครโมโซม[ 20 ]นักวิทยาศาสตร์จากทุกประเทศที่เข้าร่วมกำลังจัดทำแคตตาล็อกคุณลักษณะเหล่านี้ด้วยข้อมูลจากประชากรที่มีเชื้อสายแอฟริกัน เอเชีย และยุโรป การประเมินทั่วทั้งจีโนมดังกล่าวอาจนำไปสู่การวินิจฉัยและการบำบัดด้วยยาเพิ่มเติม ในขณะเดียวกันก็ช่วยให้ทีมในอนาคตมุ่งเน้นไปที่การวางแผนการรักษาโดยคำนึงถึงคุณลักษณะทางพันธุกรรม แนวคิดเหล่านี้กำลังถูกนำไปใช้ประโยชน์แล้วในวิศวกรรมพันธุกรรม[ 20 ]ตัวอย่างเช่น ทีมวิจัยได้สร้างเวกเตอร์ PAC shuttle ที่สร้างไลบรารีที่แสดงถึงการครอบคลุมจีโนมมนุษย์สองเท่า[ 17 ]สิ่งนี้สามารถทำหน้าที่เป็นแหล่งข้อมูลที่น่าทึ่งในการระบุยีน หรือชุดของยีน ที่ก่อให้เกิดโรค ยิ่งไปกว่านั้น การศึกษาเหล่านี้สามารถใช้เป็นวิธีที่มีประสิทธิภาพในการตรวจสอบการควบคุมการถอดรหัส ดังที่เห็นได้จากการศึกษาไวรัสบาคุโล[ 21 ]โดยรวมแล้ว ความก้าวหน้าในการสร้างคลังจีโนมและการจัดลำดับดีเอ็นเอทำให้สามารถค้นพบเป้าหมายโมเลกุลต่างๆ ได้อย่างมีประสิทธิภาพ[ 5 ]การผสมผสานคุณสมบัติเหล่านี้ผ่านวิธีการที่มีประสิทธิภาพดังกล่าวสามารถเร่งการใช้ตัวยาใหม่ๆ ได้
อ่านเพิ่มเติม
Klug, Cummings, Spencer, Palladino (2010). สาระสำคัญของพันธุศาสตร์ . เพียร์สัน. หน้า 355–264 . ISBN 978-0-321-61869-6.{{cite book}}: CS1 maint: multiple names: authors list ( link )
ลิงก์ภายนอก
- การสร้างไลบรารี BAC จีโนม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ ห้องสมุดจีโนม
ห้องสมุด จีโนม คือชุดของชิ้นส่วน DNA ที่ซ้อนทับกันซึ่งประกอบกันเป็น DNA จีโนมทั้งหมด ของ สิ่งมีชีวิต หนึ่งเดียว DNA จะถูกเก็บไว้ในกลุ่มของ เวกเตอร์ ที่เหมือนกัน...
ประวัติศาสตร์
จีโนม ที่ใช้ดีเอ็นเอเป็นครั้งแรก ที่ได้รับการจัดลำดับอย่างสมบูรณ์นั้นสำเร็จได้โดย เฟรเดอริก แซงเกอร์ ผู้ได้รับรางวัลโนเบลสองครั้งในปี 1977 แซงเกอร์และทีมงานนักวิทยาศาสตร์ของเขาได้สร้างคลังของ แบคทีริโอเฟ จ phi X 174 เพื่อใช้ใน การจัดลำดับ ดีเอ็นเอ [ 6 ]...
การสร้างคลังข้อมูลจีโนม
การสร้างไลบรารีจีโนมเกี่ยวข้องกับการสร้าง โมเลกุล DNA ลูกผสม จำนวนมาก DNA จีโนมของสิ่งมีชีวิตจะถูกสกัดและย่อยด้วย เอนไซม์ตัดจำเพาะ สำหรับสิ่งมีชีวิตที่มีจีโนมขนาดเล็กมาก (~10 kb) ชิ้นส่วนที่ถูกย่อยสามารถแยกออกจากกันได้ด้วย เจลอิเล็กโทรโฟเรซิส...
การหาค่าไทเตอร์ของไลบรารี
หลังจากสร้างไลบรารีจีโนมด้วยเวกเตอร์ไวรัส เช่น แลมบ์ดาฟาจ แล้ว สามารถกำหนด ไท เตอร์ ของไลบรารีได้ การคำนวณไทเตอร์ช่วยให้นักวิจัยสามารถประมาณจำนวนอนุภาคไวรัสที่ติดเชื้อที่สร้างขึ้นในไลบรารีได้สำเร็จ โดยใช้วิธีเจือจางไลบรารีเพื่อ เปลี่ยน วัฒนธรรม ของ E.