อ่าน 10 นาที
กลุ่มแฮปโลไทป์
แฮพลอไทป์คือกลุ่มของอัลลีลในสิ่งมีชีวิตที่ได้รับการถ่ายทอดมาด้วยกันจากพ่อแม่เพียงคนเดียว และแฮพลอกรุ๊ป ( haploidมาจากภาษากรีก : ἁπλοῦς , haploûs , "หนึ่งเท่า, ง่าย" และภาษาอังกฤษ:.
กลุ่มแฮปโลไทป์
| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| ลำดับวงศ์ตระกูลทางพันธุกรรม |
|---|
| แนวคิด |
| หัวข้อที่เกี่ยวข้อง |
แฮพลอไทป์คือกลุ่มของอัลลีลในสิ่งมีชีวิตที่ได้รับการถ่ายทอดมาด้วยกันจากพ่อแม่เพียงคนเดียว[ 1 ] [ 2 ]และแฮพลอกรุ๊ป ( haploidมาจากภาษากรีก : ἁπλοῦς , haploûs , "หนึ่งเท่า, ง่าย" และภาษาอังกฤษ: กลุ่ม ) คือกลุ่มของแฮพลอไทป์ที่คล้ายคลึงกันซึ่งมีบรรพบุรุษร่วมกันที่ระบุโดยการกลายพันธุ์ของโพลีมอร์ฟิซึมแบบนิวคลีโอไทด์เดี่ยว (SNP) ที่เฉพาะเจาะจง [ 3 ] โดยเฉพาะอย่างยิ่ง แฮพลอไทป์คือชุดของอัลลีลที่เชื่อมโยงกันอย่างใกล้ชิดซึ่งมีแนวโน้มที่จะได้รับการถ่ายทอดมาด้วยกัน เนื่องจากแฮพลอกรุ๊ปประกอบด้วยแฮพลอไทป์ที่คล้ายคลึงกัน จึงมักจะสามารถทำนายแฮพลอกรุ๊ปจากแฮพลอไทป์ได้ แฮพลอกรุ๊ปแสดงถึงสายสืบเชื้อสายทางพ่อหรือแม่เพียงสายเดียว ตัวอย่างเช่น การเป็นสมาชิกในแฮพลอกรุ๊ป R-M269 ของบุคคลใด ๆ ขึ้นอยู่กับสัดส่วนที่ค่อนข้างน้อยของสารพันธุกรรมของบุคคลนั้นจากสารพันธุกรรมทั้งหมดที่บุคคลนั้นมีอยู่

แต่ละแฮปโลกรุ๊ปมีต้นกำเนิดมาจาก และยังคงเป็นส่วนหนึ่งของแฮปโลกรุ๊ป (หรือพารากรุ๊ป ) ก่อนหน้าเพียงกลุ่มเดียว ดังนั้น กลุ่มแฮปโลกรุ๊ปที่เกี่ยวข้องใดๆ ก็ตาม สามารถจำลองได้อย่างแม่นยำในรูปแบบลำดับชั้นแบบซ้อนกันโดยที่แต่ละชุด (แฮปโลกรุ๊ป) ก็เป็นส่วนย่อยของชุดที่กว้างกว่าเพียงชุดเดียว (ตรงข้ามกับแบบจำลองแบบสองฝ่าย เช่น แผนผังครอบครัวของมนุษย์) แฮปโลกรุ๊ปยังสามารถแบ่งย่อยออกเป็นกลุ่มย่อยได้อีกด้วย
โดยปกติแล้วกลุ่มแฮพลอจะถูกระบุด้วยตัวอักษรตัวแรกของตัวอักษร และการปรับปรุงจะประกอบด้วยการรวมกันของตัวเลขและตัวอักษรเพิ่มเติม เช่น (ตัวอย่างเช่น) A → A1 → A1aระบบการตั้งชื่อตามตัวอักษรได้รับการเผยแพร่ในปี 2545 โดยY Chromosome Consortium [ 4 ]
ในพันธุศาสตร์ของมนุษย์กลุ่มแฮปโลกรุ๊ปที่ศึกษากันมากที่สุดคือกลุ่มแฮปโลกรุ๊ปของโครโมโซม Y (Y-DNA)และกลุ่มแฮปโลกรุ๊ปของดีเอ็นเอไมโทคอนเดรีย (mtDNA)ซึ่งแต่ละกลุ่มสามารถใช้กำหนดลักษณะทางพันธุกรรมของประชากรได้ Y-DNA ถ่ายทอดทางสายพ่อสู่ลูกเท่านั้น ในขณะที่ mtDNA ถ่ายทอดทางสายแม่สู่ลูกทั้งสองเพศ ทั้งสองชนิดไม่เกิดการรวมตัวกันใหม่ดังนั้น Y-DNA และ mtDNA จึงเปลี่ยนแปลงได้เฉพาะจากการกลายพันธุ์โดยบังเอิญในแต่ละรุ่น โดยไม่มีการผสมผสานกันของสารพันธุกรรมจากพ่อแม่
การก่อตัวของแฮพลอกรุ๊ป
ไมโทคอนเดรียเป็นออร์แกเนลล์ ขนาดเล็ก ที่อยู่ในไซโทพลาสซึมของเซลล์ยูคาริโอติกเช่น เซลล์ของมนุษย์ หน้าที่หลักของมันคือการให้พลังงานแก่เซลล์ เชื่อกันว่าไมโทคอนเดรียเป็นลูกหลานที่ลดขนาดลงของแบคทีเรียที่ อยู่ ร่วมกันแบบ พึ่งพาอาศัยกัน ซึ่งครั้งหนึ่งเคยดำรงชีวิตอย่างอิสระ ข้อบ่งชี้อย่างหนึ่งว่าไมโทคอนเดรียเคยดำรงชีวิตอย่างอิสระคือ แต่ละตัวมีดีเอ็นเอ แบบวงกลม เรียกว่าดีเอ็นเอไมโทคอนเดรีย (mtDNA) ซึ่งมีโครงสร้างคล้ายกับแบคทีเรียมากกว่าสิ่งมีชีวิตยูคาริโอติก (ดูทฤษฎีเอนโดซิมไบ โอติก ) ดีเอ็นเอส่วนใหญ่ของมนุษย์บรรจุอยู่ในโครโมโซมในนิวเคลียสของเซลล์ แต่ mtDNA เป็นข้อยกเว้น บุคคลจะได้รับไซโทพลาสซึมและออร์แกเนลล์ที่อยู่ในไซโทพลาสซึมนั้นจากไข่ของมารดาเท่านั้นอสุจิจะส่งต่อเฉพาะดีเอ็นเอของโครโมโซมเท่านั้น ไมโทคอนเดรียทั้งหมดจากบิดาจะถูกย่อยสลายในโอโอไซต์ เมื่อ เกิด การกลายพันธุ์ในโมเลกุล mtDNA การกลายพันธุ์นั้นจึงถูกส่งต่อทางสายเลือดหญิงโดยตรง การกลายพันธุ์คือการเปลี่ยนแปลงในเบสไนโตรเจนของลำดับดีเอ็นเอการเปลี่ยนแปลงเพียงจุดเดียวจากลำดับเดิมเรียกว่าโพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยว (SNPs)
โครโมโซม Yของมนุษย์ เป็น โครโมโซมเพศเฉพาะเพศชายเกือบทุกคนที่มีโครโมโซม Y จะมีลักษณะทางกายภาพเป็นเพศชาย แม้ว่าโครโมโซม Y จะอยู่ในนิวเคลียสของเซลล์และจับคู่กับโครโมโซม Xแต่จะเกิดการรวมตัวใหม่กับโครโมโซม X เฉพาะที่ปลายของโครโมโซม Yเท่านั้น ส่วนที่เหลืออีก 95% ของโครโมโซม Y จะไม่เกิดการรวมตัวใหม่ ดังนั้น โครโมโซม Y และการกลายพันธุ์ใดๆ ที่เกิดขึ้นในโครโมโซม Y จึงถูกส่งต่อทางสายเลือดเพศชายโดยตรง
โครโมโซมอื่นๆ เช่นออโตโซมและโครโมโซม X (เมื่อมีโครโมโซม X อีกตัวพร้อมที่จะจับคู่) จะเกิดการไขว้กันและแลกเปลี่ยนสารพันธุกรรมระหว่างกระบวนการ แบ่งเซลล์ แบบไมโอซิส ซึ่ง เป็นกระบวนการแบ่งเซลล์ที่สร้างเซลล์สืบพันธุ์โดยหลักแล้วหมายความว่าสารพันธุกรรมจากโครโมโซมเหล่านี้จะผสมปนเปกันในทุกๆ รุ่น และการกลายพันธุ์ใหม่ๆ จะถูกส่งต่อจากพ่อแม่ไปยังลูกหลานแบบสุ่ม อย่างไรก็ตามการเชื่อมโยงทางพันธุกรรมยังคงมีอยู่: การกลายพันธุ์ที่อยู่ใกล้กันมักจะถูกส่งต่อร่วมกันเนื่องจากกลไกการไขว้กัน ดังนั้นจึงยังคงเป็นไปได้ที่จะ"เดา" แฮพลอไทป์ในไม่กี่รุ่น แต่การจัดหมวดหมู่ "กลุ่ม" ในหลายรุ่นนั้นเป็นไปไม่ได้
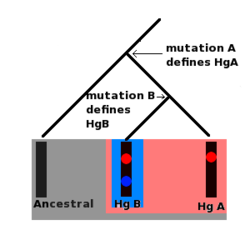
ลักษณะพิเศษที่ทั้งโครโมโซม Y และ mtDNA แสดงให้เห็นคือ การกลายพันธุ์สามารถสะสมได้ตามส่วนใดส่วนหนึ่งของโมเลกุลทั้งสอง และการกลายพันธุ์เหล่านี้จะคงที่อยู่บนดีเอ็นเอ นอกจากนี้ ยังสามารถอนุมานลำดับทางประวัติศาสตร์ของการกลายพันธุ์เหล่านี้ได้อีกด้วย ตัวอย่างเช่น หากชุดโครโมโซม Y จำนวนสิบตัว (ที่ได้มาจากบุคคลสิบคน) มีการกลายพันธุ์ A แต่มีเพียงห้าตัวเท่านั้นที่มีการกลายพันธุ์ B ก็มีความเป็นไปได้สูงมากที่การกลายพันธุ์ B เกิดขึ้นหลังจากการกลายพันธุ์ A
นอกจากนี้ บุคคลทั้งสิบคนที่มียีนกลายพันธุ์ A ล้วนเป็นลูกหลานทางสายเลือดชายโดยตรงจากชายคนเดียวกัน ซึ่งเป็นบุคคลแรกที่มียีนกลายพันธุ์นี้ ชายคนแรกที่มียีนกลายพันธุ์ B ก็เป็นลูกหลานทางสายเลือดชายโดยตรงจากชายคนนี้เช่นกัน แต่เขายังเป็นบรรพบุรุษทางสายเลือดชายโดยตรงของชายทุกคนที่มียีนกลายพันธุ์ B ด้วย การกลายพันธุ์หลายชุดเช่นนี้ก่อให้เกิดสายเลือดระดับโมเลกุล ยิ่งไปกว่านั้น การกลายพันธุ์แต่ละครั้งจะกำหนดชุดของโครโมโซม Y ที่เฉพาะเจาะจง เรียกว่า แฮพลอกรุ๊ป
มนุษย์ทุกคนที่มียีนกลายพันธุ์ A จะรวมกันเป็นแฮปโลกรุ๊ปเดียว และมนุษย์ทุกคนที่มียีนกลายพันธุ์ B ก็เป็นส่วนหนึ่งของแฮปโลกรุ๊ปนี้เช่นกัน แต่ยีนกลายพันธุ์ B ยังกำหนดแฮปโลกรุ๊ปที่เกิดขึ้นใหม่กว่า (ซึ่งเป็นกลุ่มย่อยหรือสายย่อย ) ของตัวเอง ซึ่งมนุษย์ที่มียีนกลายพันธุ์ A เพียงอย่างเดียวจะไม่เป็นสมาชิก ทั้งดีเอ็นเอไมโทคอนเดรียและโครโมโซม Y จะถูกจัดกลุ่มเป็นสายเลือดและแฮปโลกรุ๊ป ซึ่งมักแสดงในรูปแผนภาพคล้ายต้นไม้
กลุ่มแฮปโลไทป์ของดีเอ็นเอโครโมโซม Y ของมนุษย์

กลุ่มแฮพลอของดีเอ็นเอ โครโมโซม Yของมนุษย์(Y-DNA) ได้รับการตั้งชื่อตั้งแต่ A ถึง T และยังแบ่งย่อยเพิ่มเติมโดยใช้ตัวเลขและตัวอักษรพิมพ์เล็ก การกำหนดกลุ่มแฮพลอของโครโมโซม Y ได้รับการกำหนดโดย Y Chromosome Consortium [ 5 ]
Y-chromosomal Adamคือชื่อที่นักวิจัยตั้งให้กับเพศชายที่เป็นบรรพบุรุษร่วมสายเลือดชายที่ใกล้ที่สุดของมนุษย์ทุกคนที่ยังมีชีวิตอยู่
กลุ่มแฮปโลไทป์หลักของโครโมโซม Y และภูมิภาคทางภูมิศาสตร์ที่พบ (ก่อนการเข้ามาล่าอาณานิคมของชาวยุโรปในยุคหลัง) ได้แก่:
กลุ่มที่ไม่มีการกลายพันธุ์ M168
- กลุ่มแฮปโลกรุ๊ป A (M91) (แอฟริกา โดยเฉพาะชาวโคยซานและชาวนิโลต )
- กลุ่มแฮปโลกรุ๊ป B (M60) (แอฟริกา โดยเฉพาะชาวปิกมีและชาวฮัดซาเบ )
กลุ่มที่มีการกลายพันธุ์ M168
(การกลายพันธุ์ M168 เกิดขึ้นที่ระยะประมาณ 50,000 bp)
- กลุ่มแฮปโลกรุ๊ป C (M130) (โอเชียเนีย เอเชียเหนือ/กลาง/ตะวันออก อเมริกาเหนือ และพบเล็กน้อยในอเมริกาใต้ เอเชียตะวันออกเฉียงใต้ เอเชียใต้ เอเชียตะวันตก และยุโรป)
- กลุ่มแฮปโลไทป์ YAP+
- กลุ่มแฮปโลไทป์ DE (M1, M145, M203)
- กลุ่มแฮปโลกรุ๊ป D (CTS3946) (ทิเบต เนปาล ญี่ปุ่น หมู่เกาะอันดามัน เอเชียกลาง และพบประปรายในไนจีเรีย ซีเรีย และซาอุดีอาระเบีย)
- กลุ่มแฮปโลไทป์ E (M96)
- กลุ่มแฮปโลไทป์ E1b1a (V38) แอฟริกาตะวันตกและภูมิภาคโดยรอบ เดิมรู้จักกันในชื่อ E3a
- กลุ่มแฮปโลไทป์ E1b1b (M215) เกี่ยวข้องกับการแพร่กระจายของภาษาแอฟริกาเอเชีย ปัจจุบันกระจุกตัวอยู่ในแอฟริกาเหนือและแอฟริกาตะวันออกเฉียงเหนือ รวมถึงบางส่วนของตะวันออกกลาง เมดิเตอร์เรเนียน และบอลข่าน เดิมรู้จักกันในชื่อ E3b
- กลุ่มแฮปโลไทป์ DE (M1, M145, M203)
กลุ่มที่มีการกลายพันธุ์ M89
(การกลายพันธุ์ M89 เกิดขึ้นที่ระยะประมาณ 45,000 bp)
- กลุ่มแฮปโลกรุ๊ป F (M89) โอเชียเนีย ยุโรป เอเชีย อเมริกาเหนือและอเมริกาใต้
- กลุ่มแฮปโลกรุ๊ป FT (P14, M213) (จีน, เวียดนาม, [ 6 ]สิงคโปร์[ 7 ] )
- กลุ่มแฮปโลไทป์ G (M201) (พบได้ในกลุ่มชาติพันธุ์หลายกลุ่มในยูเรเซีย โดยปกติมีความถี่ต่ำ พบมากที่สุดในเทือกเขาคอเคซัสที่ราบสูงอิหร่านและอนาโตเลีย ในยุโรปส่วนใหญ่พบในกรีซ อิตาลีไอบีเรียไทโรล และโบฮีเมีย พบได้น้อยในยุโรปเหนือ)
- กลุ่มแฮปโลไทป์ H (L901/M2939)
- H1'3 (Z4221/M2826, Z13960)
- H1 (L902/M3061)
- H1a (M69/Page45) อินเดีย ศรีลังกา เนปาล ปากีสถาน อิหร่าน เอเชียกลาง
- H1b (B108) พบใน บุคคล ชาวพม่าในประเทศเมียนมาร์[ 7 ]
- H3 (Z5857) อินเดีย ศรีลังกา ปากีสถาน บาห์เรน กาตาร์
- H1 (L902/M3061)
- H2 (P96) เดิมรู้จักกันในชื่อแฮปโลกรุ๊ป F3 พบได้ในความถี่ต่ำในยุโรปและเอเชียตะวันตก
- H1'3 (Z4221/M2826, Z13960)
- กลุ่มแฮปโลไทป์ IJK (L15, L16)
กลุ่มที่มีการกลายพันธุ์ L15 และ L16
- กลุ่มแฮปโลไทป์ IJK (L15, L16)
- กลุ่มแฮปโลไทป์ IJ (S2, S22)
- กลุ่มแฮปโลกรุ๊ป I (M170, P19, M258) (แพร่หลายในยุโรป พบไม่บ่อยในบางส่วนของตะวันออกกลาง และแทบไม่มีในที่อื่น[ 8 ] )
- กลุ่มแฮปโลไทป์ I1 (M253, M307, P30, P40) (ยุโรปเหนือ พบมากในสแกนดิเนเวีย)
- กลุ่มแฮปโลไทป์ I2 (S31) (ยุโรปกลางและตะวันออกเฉียงใต้, ซาร์ดิเนีย, บอลข่าน)
- กลุ่มแฮปโลไทป์ J (M304) (ตะวันออกกลาง ตุรกี คอเคซัส อิตาลี กรีซ บัลแกเรีย แอฟริกาเหนือ)
- กลุ่มแฮปโล กรุ๊ปJ* (พบมากในเกาะโซโคตราและพบบ้างในปากีสถานโอมานกรีซสาธารณรัฐเช็กและในกลุ่มชนชาติเติร์ก )
- กลุ่มแฮปโลไทป์ J1 (M267) (ส่วนใหญ่เกี่ยวข้องกับชนชาติเซมิติกในตะวันออกกลาง แต่ก็พบได้ในยุโรปเมดิเตอร์เรเนียน เอธิโอเปีย แอฟริกาเหนือ ปากีสถาน อินเดีย และชนชาติคอเคซัสตะวันออกเฉียงเหนือ ใน ดาเกสถาน ; J1 ที่มี DYS388=13 เกี่ยวข้องกับอนาโตเลียตะวันออก)
- กลุ่มแฮปโลไทป์ J2 (M172) (พบมากในเอเชียตะวันตก เอเชียกลาง อิหร่าน อิตาลี กรีซ บัลแกเรีย และแอฟริกาเหนือ)
- กลุ่มแฮปโลกรุ๊ป I (M170, P19, M258) (แพร่หลายในยุโรป พบไม่บ่อยในบางส่วนของตะวันออกกลาง และแทบไม่มีในที่อื่น[ 8 ] )
- กลุ่มแฮปโลกรุ๊ป K (M9, P128, P131, P132)
- กลุ่มแฮปโลไทป์ IJ (S2, S22)
กลุ่มที่มีการกลายพันธุ์ M9
(การกลายพันธุ์ M9 เกิดขึ้นที่ระยะประมาณ 40,000 bp )
- กลุ่มแฮปโลกรุ๊ป K
- กลุ่มแฮปโลไทป์ LT (L298/P326)
- กลุ่มแฮปโลไทป์ L (M11, M20, M22, M61, M185, M295) (เอเชียใต้, เอเชียกลาง, เอเชียตะวันตกเฉียงใต้, เมดิเตอร์เรเนียน)
- กลุ่มแฮปโลไทป์ T (M70, M184/USP9Y+3178, M193, M272) (แอฟริกาเหนือ, แอฟริกาตะวันออก, เอเชียตะวันตกเฉียงใต้, เมดิเตอร์เรเนียน, เอเชียใต้); เดิมรู้จักกันในชื่อกลุ่มแฮปโลไทป์ K2
- กลุ่มแฮปโลไทป์ K(xLT) (rs2033003/M526)
- กลุ่มแฮปโลไทป์ LT (L298/P326)
กลุ่มที่มีการกลายพันธุ์ M526
- กลุ่มแฮปโลกรุ๊ป M (P256) (นิวกินี เมลานีเซีย อินโดนีเซียตะวันออก)
- กลุ่มแฮปโลกรุ๊ป NO (M214)
- กลุ่มแฮปโลไทป์ N (M231) (ตอนเหนือสุดของทวีปยูเรเซีย)
- กลุ่มแฮปโลกรุ๊ป O (M175) (เอเชียตะวันออก เอเชียตะวันออกเฉียงใต้ แปซิฟิกใต้ เอเชียใต้ เอเชียกลาง)
- กลุ่มแฮปโลกรุ๊ป O1 (F265)
- กลุ่มแฮปโลไทป์ O1a (M119)
- กลุ่มแฮปโลไทป์ O1b (P31, M268)
- กลุ่มแฮปโลไทป์ O2 (M122)
- กลุ่มแฮปโลกรุ๊ป O1 (F265)
- กลุ่มแฮปโลไทป์ P-M45 (M45) (M45 ปรากฏขึ้นเมื่อประมาณ 35,000 ปีก่อน)
- กลุ่มแฮปโลไทป์ Q-M242 (M242) (เกิดขึ้นเมื่อประมาณ 15,000–20,000 ปีก่อน พบในเอเชียและอเมริกา)
- กลุ่มแฮปโลกรุ๊ป Q-M3 (M3) (อเมริกาเหนือ อเมริกากลาง และอเมริกาใต้)
- กลุ่มแฮปโลกรุ๊ป R (M207)
- กลุ่มแฮปโลไทป์ R1 (M173)
- กลุ่มแฮปโลไทป์ R1a (M17) (เอเชียกลาง เอเชียใต้ และยุโรปกลาง ยุโรปเหนือ และยุโรปตะวันออก)
- กลุ่มแฮปโลไทป์ R1b (M343) (ยุโรป, คอเคซัส, เอเชียกลาง, เอเชียใต้, แอฟริกาเหนือ, แอฟริกาตอนกลาง)
- กลุ่มแฮปโลไทป์ R2 (M124) (เอเชียใต้ คอเคซัส เอเชียกลาง)
- กลุ่มแฮปโลไทป์ R1 (M173)
- กลุ่มแฮปโลไทป์ Q-M242 (M242) (เกิดขึ้นเมื่อประมาณ 15,000–20,000 ปีก่อน พบในเอเชียและอเมริกา)
- กลุ่มแฮปโลไทป์ S (M230, P202, P204) (นิวกินี, เมลานีเซีย, อินโดนีเซียตะวันออก)
กลุ่มแฮปโลไทป์ของดีเอ็นเอไมโทคอนเดรียของมนุษย์


กลุ่มแฮ ป โลไทป์ ของ mtDNAในมนุษย์จะถูกกำหนดด้วยตัวอักษรดังนี้: A , B , C , CZ , D , E , F , G , H , HV , I , J , pre -JT , JT , K , L0 , L1 , L2 , L3 , L4 , L5 , L6 , M , N , O , P , Q , R , R0 , S , T , U , V , W , X , Yและ Z
เวอร์ชันของแผนภูมิ mtDNA ได้รับการดูแลโดย Mannis van Oven บนเว็บไซต์ PhyloTree จนถึงปี 2016 [ 9 ] เมื่อจำนวนการทดสอบ mtDNA ใหม่เริ่มเพิ่มขึ้นอย่างมาก บริษัทอื่นๆ ก็เริ่มพัฒนาแผนภูมิแฮปโลไทป์ mtDNA บริษัทแรกคือ YFull ได้เปิดตัว MTree ของตน[ 10 ]ในปี 2025 FamilyTreeDNAได้เปิดตัว MitoTree(Beta) ของตน[ 11 ]
แผนภูมิวิวัฒนาการของกลุ่มแฮปโลไทป์ของดีเอ็นเอไมโทคอนเดรีย (mtDNA) ในมนุษย์ | |||||||||||||||||||||||||||||||||||||||
| ไมโตคอนเดรียล อีฟ ( L ) | |||||||||||||||||||||||||||||||||||||||
| แอล0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| แอล1 | แอล2 | แอล3 | แอล4 | แอล5 | แอล6 | ||||||||||||||||||||||||||||||||||
| เอ็ม | เอ็น | ||||||||||||||||||||||||||||||||||||||
| ซีซี | ดี | อี | จี | คิว | โอ | เอ | เอส | อาร์ | ฉัน | ว | X | วาย | |||||||||||||||||||||||||||
| ซี | ซ | บี | เอฟ | อาร์0 | ก่อน JT | พี | ยู | ||||||||||||||||||||||||||||||||
| เอชวี | เจที | เค | |||||||||||||||||||||||||||||||||||||
| ชม | วี | เจ | ที | ||||||||||||||||||||||||||||||||||||
Mitochondrial Eveคือชื่อที่นักวิจัยตั้งให้กับผู้หญิงที่เป็นบรรพบุรุษร่วมสายเลือดหญิงที่ใกล้ที่สุดของมนุษย์ทุกคนที่ยังมีชีวิตอยู่
การกำหนดประชากร
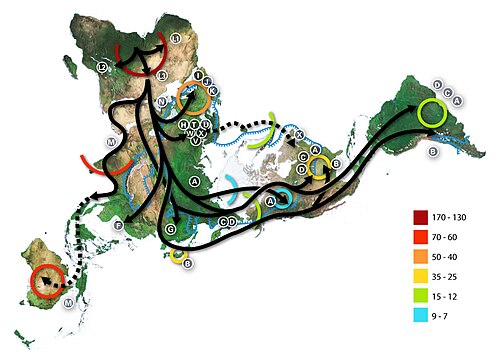
กลุ่มแฮปโลไทป์สามารถใช้ในการกำหนด ประชากร ทางพันธุกรรมและมักจะสัมพันธ์กับพื้นที่ทางภูมิศาสตร์ ตัวอย่างเช่น การแบ่งกลุ่มแฮปโลไทป์ของ mtDNA ที่พบได้ทั่วไปมีดังนี้:
- แอฟริกัน: L0 , L1 , L2 , L3 , L4 , L5 , L6
- ยูเรเซียตะวันตก: H , T , U , V , X , K , I , J , W (แฮปโลกรุ๊ปยูเรเซียตะวันตกทั้งหมดที่ระบุไว้มาจากมาโครแฮปโลกรุ๊ปN ) [ 12 ]
- ยูเรเซียตะวันออก: A , B , C , D , E , F , G , Y , Z (หมายเหตุ: C, D, E, G และ Z อยู่ในกลุ่มแฮปโลกรุ๊ปขนาดใหญ่M )
- ชนพื้นเมืองอเมริกัน: A , B , C , D , X
- ออสตราโล-เมลานีเซียน : P , Q , S
กลุ่มแฮปโลไทป์ของไมโทคอนเดรียแบ่งออกเป็นสามกลุ่มหลัก ซึ่งกำหนดโดยตัวอักษรเรียงลำดับ L, M, N มนุษยชาติแยกตัวออกเป็นกลุ่มแรกภายในกลุ่ม L ระหว่าง L0 และ L1-6 กลุ่ม L1-6 ก่อให้เกิดกลุ่ม L อื่นๆ ซึ่งหนึ่งในนั้นคือ L3 ได้แยกตัวออกเป็นกลุ่ม M และ N
กลุ่ม M ประกอบด้วยคลื่นการอพยพของมนุษย์ระลอกแรก ซึ่งเชื่อกันว่าวิวัฒนาการเกิดขึ้นนอกทวีปแอฟริกา โดยเคลื่อนตัวไปทางทิศตะวันออกตามแนวชายฝั่งทางใต้ ปัจจุบันพบสายเลือดของผู้สืบเชื้อสายจากกลุ่มแฮปโลกรุ๊ป M ทั่วเอเชีย อเมริกา และเมลานีเซีย รวมถึงบางส่วนของแอฟริกาตะวันออกและแอฟริกาเหนือ แทบไม่พบในยุโรปเลย กลุ่มแฮปโลกรุ๊ป N อาจเป็นตัวแทนของสายเลือดขนาดใหญ่อีกสายหนึ่งที่วิวัฒนาการเกิดขึ้นนอกทวีปแอฟริกา โดยมุ่งหน้าไปทางเหนือแทนที่จะไปทางตะวันออก ไม่นานหลังจากนั้น กลุ่ม R ขนาดใหญ่ก็แยกตัวออกจากกลุ่ม N
กลุ่มแฮปโลกรุ๊ป R ประกอบด้วยสองกลุ่มย่อยที่กำหนดตามการกระจายทางภูมิศาสตร์ กลุ่มหนึ่งพบในเอเชียตะวันออกเฉียงใต้และโอเชียเนีย และอีกกลุ่มหนึ่งครอบคลุมประชากรยุโรปสมัยใหม่เกือบทั้งหมด กลุ่มแฮปโลกรุ๊ป N(xR) คือ mtDNA ที่อยู่ในกลุ่ม N แต่ไม่อยู่ในกลุ่มย่อย R เป็นลักษณะเฉพาะของประชากรอะบอริจินออสเตรเลีย ในขณะเดียวกันก็พบได้ในความถี่ต่ำในประชากรหลายกลุ่มในยูเรเซียและอเมริกา
กลุ่มประเภท Lประกอบด้วยชาวแอฟริกันเกือบทั้งหมด
ประเภท Mประกอบด้วย:
M1 – ประชากรชาวเอธิโอเปีย โซมาเลีย และอินเดีย น่าจะเป็นผลมาจากการถ่ายเทยีนจำนวนมากระหว่างแอฟริกาตะวันออกและคาบสมุทรอาหรับ (ซาอุดีอาระเบีย เยเมน โอมาน) ซึ่งถูกคั่นด้วยช่องแคบแคบๆ ระหว่างทะเลแดงและอ่าวเอเดน
CZ – ชาวไซบีเรียจำนวนมาก; สาขา C – ชาวอเมริกันพื้นเมืองบางส่วน; สาขา Z – ชาวซามีจำนวนมาก ชาวเกาหลีบางส่วน ชาวจีนตอนเหนือบางส่วน และประชากรจากเอเชียกลางบางส่วน
D – ชาวอเมริกันพื้นเมืองบางส่วน ชาวไซบีเรียจำนวนมาก และชาวเอเชียตะวันออกตอนเหนือ
E – มาเลย์, บอร์เนียว, ฟิลิปปินส์, ชนพื้นเมืองไต้หวัน , ปาปัวนิวกินี
G – ชาวไซบีเรียตะวันออกเฉียงเหนือ ชาวเอเชียตะวันออกตอนเหนือ และชาวเอเชียกลางจำนวนมาก
Q – ประชากรชาวเมลานีเซีย โพลินีเซีย และปาปัวนิวกินี
ประเภท Nประกอบด้วย:
A – พบได้ในชนพื้นเมืองอเมริกันหลายกลุ่ม รวมถึงชาวเอเชียตะวันออกและชาวไซบีเรียบางกลุ่ม
I – ความถี่ 10% ในยุโรปเหนือและยุโรปตะวันออก
S – ชาวพื้นเมืองออสเตรเลียบางส่วน (ชนพื้นเมืองกลุ่มแรกของออสเตรเลีย)
W – ชาวยุโรปตะวันออกบางส่วน ชาวเอเชียใต้ และชาวเอเชียตะวันออกตอนใต้
X – ชนพื้นเมืองอเมริกันบางกลุ่ม ชาวไซบีเรียตอนใต้ ชาวเอเชียตะวันตกเฉียงใต้ และชาวยุโรปตอนใต้
Y – พบมากใน ชาว นิฟค์และชาวเกาะเนียส เป็นส่วนใหญ่ พบในชาวไอนุ ชาวตังกูสิกและชาวออสโตรเน เซียน จำนวนมากนอกจากนี้ยังพบได้ในความถี่ต่ำในประชากรบางกลุ่มในไซบีเรีย เอเชียตะวันออก และเอเชียกลาง
R – กลุ่มขนาดใหญ่ที่พบภายในกลุ่มประเภท N ประชากรที่อยู่ในกลุ่มนี้สามารถแบ่งตามภูมิศาสตร์ได้เป็นเอเชียตะวันตกและเอเชียตะวันออก เกือบทั้งหมดของประชากรยุโรปและประชากรจำนวนมากในตะวันออกกลางในปัจจุบันอยู่ในกลุ่มนี้ มีเพียงส่วนน้อยที่อยู่ในกลุ่มประเภท N อื่นๆ (ดูด้านบน) ด้านล่างนี้คือกลุ่มย่อยของ R :
B – ชาวจีน ชาวทิเบต ชาวมองโกล ชาวเอเชียกลาง ชาวเกาหลี ชาวอเมริกันพื้นเมือง ชาวไซบีเรียใต้ ชาวญี่ปุ่น และชาวออสโตรเนเซียนบางส่วน
F – พบมากในเอเชียตะวันออกเฉียงใต้ โดยเฉพาะเวียดนาม 8.3% ในเกาะฮวาร์ในโครเอเชีย[ 13 ]
R0 – พบในอาระเบียและในหมู่ชาวเอธิโอเปียและโซมาเลีย; สาขา HV (สาขา H; สาขา V) – ยุโรป เอเชียตะวันตก แอฟริกาเหนือ;
กลุ่ม Pre-JT – เกิดขึ้นในแถบเลแวนต์ (บริเวณประเทศเลบานอนในปัจจุบัน) พบในประชากรเบดูอินประมาณ 25%; กลุ่มย่อย JT (กลุ่มย่อย J; กลุ่มย่อย T) – พบในยุโรปเหนือ ยุโรปตะวันออก ลุ่มแม่น้ำสินธุ และทะเลเมดิเตอร์เรเนียน
U – พบได้บ่อยในเอเชียตะวันตก อนุทวีปอินเดีย และแอลจีเรีย พบได้ตั้งแต่ประเทศอินเดียไปจนถึงทะเลเมดิเตอร์เรเนียนและส่วนอื่นๆ ของยุโรป โดยเฉพาะอย่างยิ่ง U5 พบได้บ่อยในสแกนดิเนเวียและประเทศแถบทะเลบอลติก และพบมากที่สุดในกลุ่มชาวซามิ
การกำหนดกลุ่มแฮปโลไทป์ทางภูมิศาสตร์ของโครโมโซม Y และ MtDNA
นี่คือรายการของการมอบหมายกลุ่มแฮ็ปโลกรุ๊ปทางภูมิศาสตร์ของโครโมโซม Y และ MtDNA ที่เสนอโดย Bekada และคณะ 2556 [ 14 ]
โครโมโซม Y
ตามข้อมูลจาก SNPS กลุ่มแฮปโลกรุ๊ปที่มีอายุของการสูญพันธุ์ครั้งแรกมักจะอยู่ที่ประมาณ 45-50 พันปีก่อน กลุ่มแฮปโลกรุ๊ปของการสูญพันธุ์ครั้งที่สองดูเหมือนจะแยกตัวออกมาเมื่อ 32-35 พันปีก่อน ตามข้อมูลของMal'taเหตุการณ์การสูญพันธุ์ครั้งแรกสุดดูเหมือนจะเป็นเหตุการณ์โทบา ซึ่งกลุ่มแฮปโลกรุ๊ป CDEF* ดูเหมือนจะแยกตัวออกเป็น C, DE และ F โดย C และ F แทบไม่มีอะไรเหมือนกันเลย ในขณะที่ D และ E มีความเหมือนกันมาก เหตุการณ์การสูญพันธุ์ครั้งที่ 1 ตามการประมาณการในปัจจุบันเกิดขึ้นหลังจากโทบา แม้ว่าดีเอ็นเอโบราณที่เก่ากว่าอาจผลักดันเหตุการณ์การสูญพันธุ์ครั้งแรกสุดให้เกิดขึ้นก่อนโทบา และผลักดันเหตุการณ์การสูญพันธุ์ครั้งแรกกลับไปที่โทบา กลุ่มแฮปโลกรุ๊ปที่มีบันทึกเกี่ยวกับเหตุการณ์การสูญพันธุ์นั้นมีที่มาที่น่าสงสัย เนื่องจากเหตุการณ์การสูญพันธุ์นำไปสู่ภาวะคอขวดอย่างรุนแรง ดังนั้นบันทึกทั้งหมดของกลุ่มเหล่านี้จึงเป็นเพียงการคาดเดา โปรดทราบว่าการนับ SNP ของดีเอ็นเอโบราณนั้นมีความแปรปรวนสูง ซึ่งหมายความว่าแม้ว่ากลุ่มเหล่านี้จะแยกตัวออกมาในช่วงเวลาเดียวกัน แต่ไม่มีใครรู้ว่าเกิดขึ้นเมื่อใด[ 15 ] [ 16 ]
| ต้นทาง | กลุ่มแฮปโลไทป์ | เครื่องหมาย |
|---|---|---|
| ยุโรป (เหตุการณ์การสูญพันธุ์ครั้งที่สอง?) | ฉัน | M170, M253, P259, M227, M507 |
| ยุโรป | ไอ1บี | P215, M438, P37.2, M359, P41.2 |
| ยุโรป | ไอ1บี2 | เอ็ม26 |
| ยุโรป | ไอ1ซี | M223, M284, P78, P95 |
| ยุโรป | เจ2เอ1 | เอ็ม47 |
| ยุโรป | เจ2เอ2 | เอ็ม67, เอ็ม166 |
| ยุโรป | เจ2เอ2เอ | เอ็ม92 |
| ยุโรป | เจ2บี | เอ็ม12, เอ็ม102, เอ็ม280, เอ็ม241 |
| ยุโรป | อาร์1บี1บี1เอ | เอ็ม412, พี310 |
| ยุโรป | อาร์1บี1บี1เอ1 | แอล11 |
| ยุโรป | อาร์1บี1บี1เอ1เอ | ยู106 |
| ยุโรป | อาร์1บี1บี1เอ1บี | U198, P312, S116 |
| ยุโรป | อาร์1บี1บี1เอ1บี1 | ยู152 |
| ยุโรป | อาร์1บี1บี1เอ1บี2 | เอ็ม529 |
| ยุโรป | R1b1b1a1b3,4 | เอ็ม65, เอ็ม153 |
| ยุโรป | อาร์1บี1บี1เอ1บี5 | SRY2627 |
| เอเชียใต้ หรือ เมลานีเซีย | C1 (เดิมชื่อ CxC3) | Z1426 |
| เอเชียเหนือ | ซี2 (เดิมชื่อ ซี3) | เอ็ม217+ |
| อินโดนีเซียหรือเอเชียใต้ | เอฟ | เอ็ม89, เอ็ม282 |
| ยุโรป (คอเคซัส) | จี | M201, M285, P15, P16, M406 |
| เอเชียใต้ | ชม | M69, M52, M82, M197, M370 |
| ยุโรปหรือตะวันออกกลาง | เจ1 | M304, M267, P58, M365, M368, M369 |
| ยุโรปหรือตะวันออกกลาง | เจ2 | M172, M410, M158, M319, DYS445=6, M339, M340 |
| ทางตะวันตกของพม่าในยูเรเซีย (เหตุการณ์การสูญพันธุ์ครั้งแรก?) [ 17 ] | ||
| อินโดนีเซีย (เหตุการณ์การสูญพันธุ์ครั้งแรก?) [ 17 ] | เค2 (น็อปส์) | เอ็ม526 |
| เอเชียใต้ | แอล | M11, M20, M27, M76, M317, M274, M349, M357 |
| เอเชียตะวันออก เอเชียตะวันออกเฉียงใต้ | เอ็น | M231, M214, LLY22g, Tat, M178 |
| เอเชียตะวันออก, เอเชียตะวันออกเฉียงใต้, เอเชียใต้ | โอ | เอ็ม175, เอ็ม119 |
| อินโดนีเซีย ฟิลิปปินส์ | พี (xQR) | 92R7, M207, M173, M45 |
| เอเชียใต้ ไซบีเรีย | R และ Q (QR) แยก[ 17 ] | MEH2, M242, P36.2, M25, M346 |
| ตะวันออกกลาง ยุโรป ไซบีเรีย เอเชียใต้ | ร1า1 | M420, M17, M198, M204, M458 |
| อนาโตเลีย ยุโรปตะวันออกเฉียงใต้ ? | อาร์1บี | เอ็ม173, เอ็ม343, พี25, เอ็ม73 |
| ยุโรป | อาร์1บี1บี | เอ็ม269 |
| ยุโรป | อาร์1บี1บี1 | แอล23 |
| ปากีสถาน อินเดีย | อาร์2 | เอ็ม479, เอ็ม124 |
| ตะวันออกกลาง | ที | เอ็ม70 |
| แอฟริกาเหนือ | อี1บี1บี1 | เอ็ม35 |
| แอฟริกาเหนือ | อี1บี1บี1เอ | เอ็ม78 |
| เอเชียตะวันตก | อี1บี1บี1เอ2 | V13 |
| แอฟริกาเหนือ | อี1บี1บี1เอ1 | วี12 |
| แอฟริกาเหนือ | อี1บี1บี1เอ1บี | วี32 |
| แอฟริกาเหนือ | อี1บี1บี1เอ3 | วี22 |
| แอฟริกาเหนือ | อี1บี1บี1เอ4 | วี65 |
| แอฟริกาเหนือ | อี1บี1บี1บี | เอ็ม81 |
| แอฟริกาเหนือ | อี1บี1บี1ซี | เอ็ม123, เอ็ม34 |
| แอฟริกาตะวันตก, แอฟริกาเหนือ | เอ | เอ็ม91, เอ็ม13 |
| แอฟริกาตะวันออก | บี | M60, M181, SRY10831.1, M150, M109, M112 |
| เอเชีย แอฟริกา | ดีอี | M1, ใช่, M174, M40, M96, M75, M98 |
| เอเชียตะวันออก, เนปาล | ดี | เอ็ม174 |
| แอฟริกาตะวันตก (เหตุการณ์การสูญพันธุ์ครั้งแรก?) | อี1เอ | เอ็ม33 |
| แอฟริกาตะวันออก (เหตุการณ์การสูญพันธุ์ครั้งแรกคือการแยกตัวระหว่าง E1b1 และ E1a เหตุการณ์การสูญพันธุ์ครั้งที่สองคือการแยกตัวระหว่าง E1b1b และ E1b1a) | อี1บี1 | พี2, เอ็ม2, ยู175, เอ็ม191 |
| ตะวันออกกลาง | เจ1 | พี58 |
ดีเอ็นเอไมโทคอนเดรีย
| ต้นทาง | กลุ่มแฮปโลไทป์ |
|---|---|
| ยุโรป | เอช1 |
| ยุโรป | เอช11เอ |
| ยุโรป | ฮ่า |
| ยุโรป | เอช1บี |
| ยุโรป | เอช2เอ |
| ยุโรป | เอช3 |
| ยุโรป | เอช5เอ |
| ยุโรป | เอช6เอ |
| ยุโรป | เอช7 |
| ยุโรป | HV0/HV0a/V |
| ยุโรป | ไอ4 |
| ยุโรป | เจ1ซี7 |
| ยุโรป | เจ2บี1 |
| ยุโรป | ที2บี* |
| ยุโรป | ที2บี4 |
| ยุโรป | ที2อี |
| ยุโรป | ยู4ซี1 |
| ยุโรป | ยู5* |
| ยุโรป | ยู5เอ |
| ยุโรป | ยู5เอ1บี1 |
| ยุโรป | ยู5บี* |
| ยุโรป | ยู5บี1บี* |
| ยุโรป | ยู5บี1ซี |
| ยุโรป | ยู5บี3 |
| ยุโรป | X2c'e |
| ตะวันออกกลาง | ฉัน |
| ตะวันออกกลาง | เอ |
| ตะวันออกกลาง | บี |
| ตะวันออกกลาง | ซี/ซี |
| ตะวันออกกลาง | ดี/จี/เอ็ม9/อี |
| อินเดีย | เอฟ |
| ตะวันออกกลาง | ชม* |
| ตะวันออกกลาง | เอช13เอ1 |
| ตะวันออกกลาง | เอช14เอ |
| ตะวันออกกลาง | เอช20 |
| ตะวันออกกลาง | เอช2เอ1 |
| ตะวันออกกลาง | เอช4 |
| ตะวันออกกลาง | เอช6บี |
| ตะวันออกกลาง | เอช8 |
| ตะวันออกกลาง | เอชวี1 |
| ตะวันออกกลาง | ไอ1 |
| ตะวันออกกลาง | เจ / เจ1ซี / เจ2 |
| ตะวันออกกลาง | เจ1อาบีอี |
| ตะวันออกกลาง | เจ1บี1เอ1 |
| ตะวันออกกลาง | เจ1บี2เอ |
| ตะวันออกกลาง | เจ1ดี / เจ2บี |
| ตะวันออกกลาง | เจ1ดี1 |
| ตะวันออกกลาง | เจ2เอ |
| ตะวันออกกลาง | เจ2เอ2เอ1 |
| ตะวันออกกลาง | เค* |
| ตะวันออกกลาง | เค1อา* |
| ตะวันออกกลาง | เค1บี1* |
| ตะวันออกกลาง | เอ็น1อา* |
| ตะวันออกกลาง | เอ็น1บี |
| ตะวันออกกลาง | เอ็น1ซี |
| ตะวันออกกลาง | เอ็น2 |
| ตะวันออกกลาง | เอ็น9 |
| ตะวันออกกลาง | อาร์* |
| ตะวันออกกลาง | ร0a |
| ตะวันออกกลาง | ที |
| ตะวันออกกลาง | ที1* |
| เอเชียตะวันตก | ที1อา |
| ตะวันออกกลาง | ที2 |
| ตะวันออกกลาง | ที2ซี |
| ตะวันออกกลาง | ที2ไอ |
| ตะวันออกกลาง | ยู1* |
| ตะวันออกกลาง | U2* |
| ตะวันออกกลาง | ยู2อี |
| ยูเรเซีย | ยู3* |
| ตะวันออกกลาง | ยู4 |
| ตะวันออกกลาง | ยู4เอ* |
| ตะวันออกกลาง | ยู7 |
| ตะวันออกกลาง | ยู8* |
| ตะวันออกกลาง | ยู9เอ |
| ตะวันออกกลาง | X |
| ตะวันออกกลาง | เอ็กซ์1เอ |
| ตะวันออกกลาง | เอ็กซ์2บี1 |
| แอฟริกาเหนือ | L3e5 |
| แอฟริกาเหนือ | เอ็ม1 |
| แอฟริกาเหนือ | เอ็ม1เอ1 |
| แอฟริกาเหนือ | ยู6เอ |
| แอฟริกาเหนือ | U6a1'2'3 |
| แอฟริกาเหนือ | ยู6บีซีดี |
| แอฟริกาตะวันออก | ล0* |
| แอฟริกาตะวันออก | ล0a1 |
| แอฟริกาตะวันออก | แอล0เอ1บี |
| แอฟริกาตะวันออก | แอล0เอ2* |
| แอฟริกาตะวันออก | L3c/L4/M |
| แอฟริกาตะวันออก | L3d1a1 |
| แอฟริกาตะวันออก | แอล3ดี1ดี |
| แอฟริกาตะวันออก | L3e1* |
| แอฟริกาตะวันออก | แอล3เอฟ* |
| แอฟริกาตะวันออก | L3h1b* |
| แอฟริกาตะวันออก | L3i* |
| แอฟริกาตะวันออก | แอล3เอ็กซ์* |
| แอฟริกาตะวันออก | L4a'b* |
| แอฟริกาตะวันออก | แอล5* |
| แอฟริกาตะวันออก | แอล6 |
| แอฟริกาตะวันออก | N* / M* / L3* |
| แอฟริกาตะวันตก | แอล1บี* |
| แอฟริกาตะวันตก | แอล1บี3 |
| แอฟริกาตะวันตก | แอล1ซี* |
| แอฟริกาตะวันตก | แอล1ซี2 |
| แอฟริกาตะวันตก | แอล2* |
| แอฟริกาตะวันตก | แอล2เอ |
| แอฟริกาตะวันตก | แอล2เอ1* |
| แอฟริกาตะวันตก | L2a1a2'3'4 |
| แอฟริกาตะวันตก | แอล2เอ1บี |
| แอฟริกาตะวันตก | แอล2เอ1บีเอฟ |
| แอฟริกาตะวันตก | แอล2เอ1ซี1'2 |
| แอฟริกาตะวันตก | L2a1(16189) |
| แอฟริกาตะวันตก | แอล2เอ2 |
| แอฟริกาตะวันตก | แอล2บี* |
| แอฟริกาตะวันตก | แอล2ซี1'2 |
| แอฟริกาตะวันตก | แอล2ดี |
| แอฟริกาตะวันตก | แอล2อี |
| แอฟริกาตะวันตก | แอล3บี |
| แอฟริกาตะวันตก | แอล3บี1เอ3 |
| แอฟริกาตะวันตก | L3b(16124!) |
| แอฟริกาตะวันตก | แอล3บี2เอ |
| แอฟริกาตะวันตก | แอล3ดี* |
| แอฟริกาตะวันตก | L3e2'3'4 |
| แอฟริกาตะวันตก | L3f1b* |
ดูเพิ่มเติม
- โครงการ HapMap ระหว่างประเทศ
- วิวัฒนาการระดับโมเลกุล
- พันธุศาสตร์เชิงโมเลกุล
- พันธุศาสตร์วิวัฒนาการของมนุษย์
- เชื้อชาติ (การจำแนกประเภท) / เชื้อชาติ (การจัดกลุ่มมนุษย์)
- ลำดับวงศ์ตระกูลทางพันธุกรรม
- การตรวจดีเอ็นเอเพื่อสืบสายตระกูล
- รายชื่อหัวข้อทางพันธุศาสตร์เชิงลำดับวงศ์ตระกูล
- รายชื่อแฮปโลกรุ๊ปของบุคคลสำคัญ
ลิงก์ภายนอก
ทั่วไป
- โครงการจีโนกราฟิก
กลุ่มแฮพลอกรุ๊ป DNA ทั้งหมด
โครโมโซม Y
- แผนที่แฮปโลกรุ๊ปทั่วโลก (PDF)
กลุ่มแฮพลอของดีเอ็นเอโครโมโซม Y
- กลุ่มโครโมโซม Y
- แผนผังแฮปโลกรุ๊ป Y ของ ISOGG
- Y-tree ของ PhyloTree:แผนภูมิวิวัฒนาการอ้างอิงแบบง่ายสำหรับโครโมโซม Y ของมนุษย์
- ตัวทำนายแฮปโลกรุ๊ป
- กลุ่ม Y Chromosome Consortium (2002), ระบบการตั้งชื่อสำหรับแผนภูมิของกลุ่มแฮปโลกรุ๊ปไบนารี Y-โครโมโซมของมนุษย์ , Genome Research, Vol. 12 (2), 339–48, กุมภาพันธ์ 2002 ( แผนภูมิ แบบลำดับชั้นโดยละเอียด มีการแปลงจากแผนการตั้งชื่อก่อนหน้านี้)
- Semino et al. (2000), มรดกทางพันธุกรรมของโฮโมเซเปียนส์เซเปียนส์ในยุคหินเก่าในชาวยุโรปปัจจุบัน , Science, Vol 290 (บทความที่แนะนำกลุ่มแฮปโลกรุ๊ป "Eu")
- แผนที่ชาติพันธุ์วิทยาและพันธุศาสตร์ Y-DNA และการรวบรวมข้อมูลแบบโอเพนซอร์ส
กลุ่มแฮพลอไทป์ของดีเอ็นเอไมโตคอนเดรีย
- PhyloTree – แผนภูมิวิวัฒนาการของความแปรผันของดีเอ็นเอไมโทคอนเดรียของมนุษย์ทั่วโลก
- PhyloD3 – แผนภูมิวิวัฒนาการแบบ D3.js โดยอิงจาก PhyloTree
- MitoTool – เว็บเซิร์ฟเวอร์สำหรับการวิเคราะห์และค้นหาความแปรผันของลำดับดีเอ็นเอไมโทคอนเดรียของมนุษย์เก็บถาวรเมื่อวันที่ 19 มิถุนายน 2016 ที่Wayback Machine
- HaploGrep – การจำแนกกลุ่มแฮปโลไทป์ของดีเอ็นเอไมโทคอนเดรียโดยอัตโนมัติโดยใช้ PhyloTree ( เก็บถาวรเมื่อ 12 มิถุนายน 2016 ที่Wayback Machine)
- HaploFind – ระบบค้นหาแฮปโลกรุ๊ปอัตโนมัติที่รวดเร็วสำหรับดีเอ็นเอไมโทคอนเดรียของมนุษย์เก็บถาวรเมื่อ 11 มิถุนายน 2016 ที่Wayback Machine
- โครงร่างแฮปโลกรุ๊ป mtDNA แบบกราฟิก
- การสร้างภูมิทัศน์ mtDNA ของแอฟริกา
- กลุ่มย่อยทั้งสี่ของกลุ่มแฮปโลกรุ๊ป L2 ของ mtDNA มีวิวัฒนาการในอัตราที่แตกต่างกันหรือไม่?
{kind=link}
ซอฟต์แวร์
- โปรแกรมค้นหาแฮปโลกรุ๊ป Y-DNA
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ กลุ่มแฮปโลไทป์
แฮพลอไทป์คือกลุ่มของอัลลีลในสิ่งมีชีวิตที่ได้รับการถ่ายทอดมาด้วยกันจากพ่อแม่เพียงคนเดียว และแฮพลอกรุ๊ป ( haploidมาจากภาษากรีก : ἁπλοῦς , haploûs , "หนึ่งเท่า, ง่าย" และภาษาอังกฤษ:.
การก่อตัวของแฮพลอกรุ๊ป
ไมโทคอนเดรีย เป็น ออร์แกเนลล์ ขนาดเล็ก ที่อยู่ในไซ โทพลาสซึม ของ เซลล์ยูคาริโอติก เช่น เซลล์ของมนุษย์ หน้าที่หลักของมันคือการให้พลังงานแก่เซลล์ เชื่อกันว่าไมโทคอนเดรียเป็นลูกหลานที่ลดขนาดลงของ แบคทีเรียที่ อยู่ ร่วมกันแบบ พึ่งพาอาศัยกัน...
กลุ่มแฮปโลไทป์ของดีเอ็นเอโครโมโซม Y ของมนุษย์
กลุ่มแฮพลอของดีเอ็นเอ โครโมโซม Y ของมนุษย์(Y-DNA) ได้รับการตั้งชื่อตั้งแต่ A ถึง T และยังแบ่งย่อยเพิ่มเติมโดยใช้ตัวเลขและตัวอักษรพิมพ์เล็ก การกำหนดกลุ่มแฮพลอของโครโมโซม Y ได้รับการกำหนดโดย Y Chromosome Consortium [ 5 ]
กลุ่มแฮปโลไทป์ของดีเอ็นเอไมโทคอนเดรียของมนุษย์
กลุ่มแฮ ป โลไทป์ ของ mtDNA ในมนุษย์จะถูกกำหนดด้วยตัวอักษรดังนี้: A , B , C , CZ , D , E , F , G , H , HV , I , J , pre -JT , JT , K , L0 , L1 , L2 , L3 , L4 , L5 , L6 , M , N , O , P , Q , R , R0 , S , T , U , V , W , X , Y และ Z