อ่าน 19 นาที
นิวคลีโอโซม
นิวคลีโอโซมเป็นหน่วยโครงสร้างพื้นฐานของ การบรรจุ DNAในยูคาริโอตโครงสร้างของนิวคลีโอโซมประกอบด้วยส่วนของ DNA ที่พันรอบโปรตีนฮิสโตน แปดตัว และมีลักษณะคล้ายเส้นด้ายที่พันรอบแกนหมุน...
นิวคลีโอโซม

นิวคลีโอโซมเป็นหน่วยโครงสร้างพื้นฐานของ การบรรจุ DNAในยูคาริโอตโครงสร้างของนิวคลีโอโซมประกอบด้วยส่วนของ DNA ที่พันรอบโปรตีนฮิสโตน แปดตัว [ 1 ]และมีลักษณะคล้ายเส้นด้ายที่พันรอบแกนหมุน นิวคลีโอโซมเป็นหน่วยย่อยพื้นฐานของโครมาติน นิวคลีโอโซมแต่ละตัวประกอบด้วย DNA ที่พันรอบโปรตีนแปดตัวที่เรียกว่าฮิสโตน ซึ่งรู้จักกันในชื่อฮิสโตนออกตาเมอร์ ฮิสโตนออกตาเมอร์แต่ละตัวประกอบด้วยโปรตีนฮิ ส โตน H2A , H2B , H3และH4อย่างละสองสำเนา
DNA ต้องถูกอัดแน่นเป็นนิวคลีโอโซมเพื่อให้พอดีกับนิวเคลียสของเซลล์[ 2 ]นอกจากการห่อหุ้มด้วยนิวคลี โอโซมแล้ว โครมาตินยังถูกอัดแน่นยิ่งขึ้นด้วยการพับเป็นโครงสร้างที่ซับซ้อนมากขึ้นเรื่อยๆ จนในที่สุดก็ก่อตัวเป็นโครโมโซม เซลล์มนุษย์แต่ละเซลล์มีนิวคลีโอโซมประมาณ 30 ล้านตัว[ 3 ]
เชื่อกันว่านิวคลีโอโซมจะบรรจุ ข้อมูล ทางพันธุกรรมแบบ เอพิเจเนติก ในรูปแบบของการดัดแปลงโควาเลน ต์ ของฮิสโตน แกนกลาง ตำแหน่งของนิวคลีโอโซมในจีโนมไม่ได้เกิดขึ้นแบบสุ่ม และเป็นสิ่งสำคัญที่จะต้องทราบว่านิวคลีโอโซมแต่ละตัวตั้งอยู่ที่ใด เพราะตำแหน่งนี้จะกำหนดการเข้าถึง DNA ของโปรตีนควบคุม[ 4 ]
นิวคลีโอโซมถูกสังเกตเห็นครั้งแรกในฐานะอนุภาคในกล้องจุลทรรศน์อิเล็กตรอนโดย Don และ Ada Olins ในปี 1974 [ 5 ]และการมีอยู่และโครงสร้างของมัน (เป็นฮิสโตนออกตาเมอร์ที่ล้อมรอบด้วยดีเอ็นเอประมาณ 200 คู่เบส) ได้รับการเสนอโดยRoger Kornberg [ 6 ] [ 7 ] บทบาทของนิวคลีโอโซมในฐานะตัวควบคุมการถอดรหัสได้รับการพิสูจน์โดย Lorch et al. ในหลอดทดลอง[ 8 ]ในปี 1987 และโดย Han และ Grunstein [ 9 ]และ Clark-Adams et al. [ 10 ]ในสิ่งมีชีวิตในปี 1988
อนุภาคแกนกลางของนิวคลีโอโซมประกอบด้วยดีเอ็นเอ ประมาณ 146 คู่เบส (bp) [ 11 ] ที่พัน รอบฮิสโตนออก ตาเมอร์แบบ ซ้าย 1.67 รอบซึ่งประกอบด้วยฮิสโตนแกนกลางH2A , H2B , H3และH4 อย่างละ 2 ชุด [ 12 ] อนุภาคแกนกลางเชื่อมต่อกันด้วยส่วนของดีเอ็นเอเชื่อมโยง ซึ่งอาจมีความยาวได้ถึงประมาณ 80 คู่เบส ในทางเทคนิค นิวคลีโอโซมถูก นิยามว่าเป็นอนุภาคแกนกลางบวกกับบริเวณเชื่อมโยงเหล่านี้หนึ่งส่วน อย่างไรก็ตาม คำนี้มักมีความหมายเหมือนกับอนุภาคแกนกลาง[ 13 ]ปัจจุบันมีแผนที่แสดงตำแหน่งนิวคลีโอโซมทั่วทั้งจีโนมสำหรับสิ่งมีชีวิตต้นแบบและเซลล์มนุษย์หลายชนิด[ 14 ]
ฮิสโตนเชื่อมโยง เช่นH1และไอโซฟอร์มของมัน มีส่วนเกี่ยวข้องกับการอัดแน่นของโครมาติน และอยู่ที่ฐานของนิวคลีโอโซมใกล้กับทางเข้าและทางออกของ DNA โดยจับกับบริเวณเชื่อมโยงของ DNA [ 15 ]นิวคลีโอโซมที่ไม่ควบแน่นโดยไม่มีฮิสโตนเชื่อมโยงจะมีลักษณะคล้าย "ลูกปัดบนสาย DNA" เมื่อมองผ่านกล้องจุลทรรศน์อิเล็กตรอน[ 16 ]
ตรงกันข้ามกับเซลล์ยูคาริโอตส่วนใหญ่ เซลล์สเปิร์มที่เจริญเต็มที่มักใช้โปรตามีนในการบรรจุดีเอ็นเอจีโนม ซึ่งน่าจะเพื่อให้ได้อัตราส่วนการบรรจุที่สูงขึ้น[ 17 ]นอกจากนี้ยังพบสารเทียบเท่าฮิสโตนและโครงสร้างโครมาตินที่เรียบง่ายในอาร์เคีย [ 18 ] ซึ่งบ่งชี้ว่ายูคาริโอตไม่ใช่สิ่งมีชีวิตเพียงชนิดเดียวที่ใช้นิวคลีโอโซม
โครงสร้าง
โครงสร้างของอนุภาคแกนกลาง

ภาพรวม
การศึกษาโครงสร้างบุกเบิกในช่วงทศวรรษ 1980 โดยกลุ่มของ Aaron Klug ได้ให้หลักฐานแรกว่าโปรตีนฮิสโตนแปดตัวพันรอบ DNA เองในเกลียวซูเปอร์เฮลิกซ์แบบซ้ายประมาณ 1.7 รอบ[ 19 ]ในปี 1997 โครงสร้างผลึกนิ วคลีโอโซมที่มีความละเอียดใกล้เคียง อะตอมเป็นครั้งแรกได้รับการแก้ไขโดยกลุ่ม Richmond ที่ETH Zurichซึ่งแสดงรายละเอียดที่สำคัญที่สุดของอนุภาคDNA พาลินโดรมอัลฟาแซ ทเทลไลต์ของมนุษย์ ซึ่งมีความสำคัญต่อการบรรลุโครงสร้างผลึกนิวคลีโอโซมในปี 1997 ได้รับการพัฒนาโดยกลุ่ม Bunick ที่ห้องปฏิบัติการแห่งชาติ Oak Ridgeในรัฐเทนเนสซี[ 20 ] [ 21 ] [ 22 ] [ 23 ] [ 24 ]จนถึงปัจจุบัน โครงสร้างของอนุภาคแกนกลางนิวคลีโอโซมมากกว่า 20 ชนิดได้รับการแก้ไขแล้ว[ 25 ]รวมถึงอนุภาคที่มีฮิสโตนชนิดต่างๆ และฮิสโตนจากสายพันธุ์ต่างๆ โครงสร้างของอนุภาคแกนกลางนิวคลีโอโซมได้รับการอนุรักษ์ไว้อย่างน่าทึ่ง และแม้แต่การเปลี่ยนแปลงมากกว่า 100 หน่วยตกค้างระหว่างฮิสโตนของกบและยีสต์ก็ส่งผลให้แผนที่ความหนาแน่นของอิเล็กตรอนมีค่าเบี่ยงเบนรากกำลังสองเฉลี่ยโดย รวม เพียง 1.6 Å เท่านั้น[ 26 ]
อนุภาคแกนกลางนิวคลีโอโซม (NCP)
อนุภาคแกนกลางนิวคลีโอโซม (แสดงในรูป) ประกอบด้วยดีเอ็นเอ ประมาณ 146 คู่เบส[ 11 ]พันรอบฮิสโตนออกตาเมอร์ แบบ มือซ้าย 1.67 รอบ โดยแต่ละคู่ประกอบด้วยฮิสโตนแกนกลางH2A , H2B , H3และH4 อย่างละ 2 ชุด นิวคลีโอโซมที่อยู่ติดกันจะเชื่อมต่อกันด้วยดีเอ็นเออิสระที่เรียกว่าดีเอ็นเอเชื่อมโยง (ซึ่งมีความยาวแตกต่างกันไปตั้งแต่ 10 - 80 คู่เบส ขึ้นอยู่กับชนิดและประเภทของเนื้อเยื่อ[ 18 ] ) โครงสร้างทั้งหมดสร้างเป็นทรงกระบอกที่มีเส้นผ่านศูนย์กลาง 11 นาโนเมตร และความสูง 5.5 นาโนเมตร



อนุภาคแกนกลางของนิวคลีโอโซมจะถูกสังเกตเมื่อโครมาตินในระยะอินเตอร์เฟสได้รับการบำบัดเพื่อให้โครมาตินคลี่ออกบางส่วน ภาพที่ได้จากกล้องจุลทรรศน์อิเล็กตรอนคือ "ลูกปัดบนเส้นเชือก" เส้นเชือกนั้นคือ DNA ในขณะที่ลูกปัดแต่ละเม็ดในนิวคลีโอโซมคืออนุภาคแกนกลาง อนุภาคแกนกลางของนิวคลีโอโซมประกอบด้วย DNA และโปรตีนฮิสโตน[ 29 ]
การย่อยโครมาติน ด้วย DNAseบางส่วนเผยให้เห็นโครงสร้างนิวคลีโอโซม เนื่องจากส่วนของ DNA ในอนุภาคแกนกลางของนิวคลีโอโซมเข้าถึงได้ยากกว่าส่วนเชื่อมต่อ DNA จึงถูกย่อยเป็นชิ้นส่วนที่มีความยาวเท่ากับผลคูณของระยะห่างระหว่างนิวคลีโอโซม (180, 360, 540 คู่เบส เป็นต้น) ดังนั้นจึงมองเห็นรูปแบบลักษณะเฉพาะที่คล้ายกับบันได ในระหว่างการแยก DNA ด้วยเจลอิเล็กโทรโฟเรซิ ส [ 27 ]การย่อยดังกล่าวสามารถเกิดขึ้นได้ภายใต้สภาวะธรรมชาติในระหว่างอะพอพโทซิส ("การฆ่าตัวตายของเซลล์" หรือการตายของเซลล์ตามโปรแกรม) เนื่องจาก การ ทำลายตัวเองของ DNAเป็นบทบาทหลักของกระบวนการนี้[ 30 ]
ปฏิสัมพันธ์ของโปรตีนภายในนิวคลีโอโซม
โปรตีนฮิสโตนหลักมีโครงสร้างลักษณะ เฉพาะ ที่เรียกว่า "ฮิสโตนโฟลด์" ซึ่งประกอบด้วยอัลฟาเฮลิกซ์ 3 อัน (α1-3) ที่คั่นด้วยลูป 2 อัน (L1-2) ในสารละลาย ฮิสโตนจะสร้างเฮเทอโรไดเมอร์ H2A-H2B และเฮเทอโรเตตระเมอร์ H3-H4 ฮิสโตนจะสร้างไดเมอร์รอบอัลฟาเฮลิกซ์ α2 ที่ยาวในทิศทางตรงข้ามขนานกัน และในกรณีของ H3 และ H4 ไดเมอร์ดังกล่าว 2 ตัวจะสร้างมัดเฮลิกซ์ 4 อันที่เสถียรโดยปฏิกิริยา H3-H3' ที่กว้างขวาง ไดเมอร์ H2A/H2B จะจับกับเตตระเมอร์ H3/H4 เนื่องจากการโต้ตอบระหว่าง H4 และ H2B ซึ่งรวมถึงการสร้างคลัสเตอร์ไฮโดรโฟบิก[ 12 ] ฮิสโตนออกตาเมอร์เกิดจากเตตระเมอร์ H3/H4 ตรงกลางที่ประกบอยู่ระหว่างไดเมอร์ H2A/H2B สองตัว เนื่องจากฮิสโตนหลักทั้งสี่ตัวมีประจุพื้นฐานสูงมาก ฮิสโตนออกตาเมอร์จึงมีเสถียรภาพเฉพาะในสภาวะที่มีดีเอ็นเอหรือความเข้มข้นของเกลือสูงมากเท่านั้น
ปฏิสัมพันธ์ระหว่างฮิสโตนและดีเอ็นเอ
นิวคลีโอโซมประกอบด้วยปฏิสัมพันธ์ระหว่างโปรตีนและ DNA โดยตรงมากกว่า 120 รายการ และปฏิสัมพันธ์ที่เชื่อมโยงกันด้วยน้ำอีกหลายร้อยรายการ[ 31 ]ปฏิสัมพันธ์ระหว่างโปรตีนและ DNA โดยตรงไม่ได้กระจายอย่างสม่ำเสมอทั่วพื้นผิวของออกตาเมอร์ แต่ตั้งอยู่ที่ตำแหน่งเฉพาะเจาะจง เนื่องจากการก่อตัวของตำแหน่งการจับ DNA สองประเภทภายในออกตาเมอร์ ได้แก่ ตำแหน่ง α1α1 ซึ่งใช้เกลียว α1 จากฮิสโตนสองตัวที่อยู่ติดกัน และตำแหน่ง L1L2 ที่เกิดจากลูป L1 และ L2 พันธะเกลือและพันธะไฮโดรเจนระหว่างกลุ่มเบสและไฮดรอกซิลของโซ่ข้างและอะไมด์ของโซ่หลักกับฟอสเฟตของโครงสร้างหลักของ DNA ก่อให้เกิดปฏิสัมพันธ์ส่วนใหญ่กับ DNA สิ่งนี้มีความสำคัญ เนื่องจากนิวคลีโอโซมมีการกระจายตัวอย่างแพร่หลายไปตามจีโนม ทำให้จำเป็นต้องเป็นปัจจัยการจับ DNA ที่ไม่จำเพาะต่อลำดับ แม้ว่านิวคลีโอโซมจะมีแนวโน้มที่จะชอบลำดับ DNA บางลำดับมากกว่าลำดับอื่น[ 32 ]แต่ก็สามารถจับกับลำดับใดๆ ก็ได้ ซึ่งเชื่อกันว่าเป็นผลมาจากความยืดหยุ่นในการสร้างปฏิสัมพันธ์ที่อาศัยน้ำเป็นตัวกลาง นอกจากนี้ ยังมีปฏิสัมพันธ์แบบไม่มีขั้วเกิดขึ้นระหว่างโซ่ข้างของโปรตีนและกลุ่มดีออกซีไรโบส และโซ่ข้างของอาร์จินีนจะแทรกเข้าไปในร่องเล็กของ DNA ที่ตำแหน่งทั้ง 14 ตำแหน่งที่หันเข้าหาพื้นผิวของออกตาเมอร์ การกระจายและความแข็งแรงของตำแหน่งการจับ DNA รอบพื้นผิวของออกตาเมอร์ทำให้ DNA ภายในแกนกลางของนิวคลีโอโซมบิดเบี้ยว DNA จะโค้งงอไม่สม่ำเสมอและยังมีข้อบกพร่องในการบิด การบิดของ DNA รูปแบบ B อิสระในสารละลายคือ 10.5 bp ต่อรอบ อย่างไรก็ตาม การบิดโดยรวมของ DNA ในนิวคลีโอโซมมีเพียง 10.2 bp ต่อรอบ โดยมีค่าแตกต่างกันไปตั้งแต่ 9.4 ถึง 10.9 bp ต่อรอบ
โดเมนหางของฮิสโตน
ส่วนต่อขยายหางของฮิสโตนมีมวลมากถึง 30% ของฮิสโตนทั้งหมด แต่ไม่สามารถมองเห็นได้ในโครงสร้างผลึกของนิวคลีโอโซมเนื่องจากมีความยืดหยุ่นสูง และเชื่อกันว่าส่วนใหญ่ไม่มีโครงสร้าง[ 33 ]หาง N-terminal ของฮิสโตน H3 และ H2B ผ่านช่องที่เกิดจากร่องเล็กของสาย DNA สองสาย โดยยื่นออกมาจาก DNA ทุกๆ 20 bp ในทางกลับกัน หาง N-terminalของฮิสโตน H4 มีบริเวณของกรดอะมิโนเบสสูง (16–25) ซึ่งในโครงสร้างผลึกจะเกิดปฏิกิริยากับบริเวณพื้นผิวที่เป็นกรดสูงของไดเมอร์ H2A-H2B ของนิวคลีโอโซมอื่น ซึ่งอาจเกี่ยวข้องกับโครงสร้างลำดับสูงของนิวคลีโอโซม ปฏิกิริยานี้เชื่อว่าเกิดขึ้นภายใต้สภาวะทางสรีรวิทยาเช่นกัน และชี้ให้เห็นว่าการอะเซทิเลชันของหาง H4 ทำให้โครงสร้างลำดับสูงของโครมาตินบิดเบี้ยว
โครงสร้างลำดับสูงกว่า

การจัดระเบียบของ DNA ที่เกิดขึ้นโดยนิวคลีโอโซมไม่สามารถอธิบายการบรรจุ DNA ที่สังเกตได้ในนิวเคลียสของเซลล์ได้อย่างสมบูรณ์จำเป็นต้อง มีการอัดแน่นของ โครมาติน เข้าไปในนิวเคลียสของเซลล์มากขึ้น แต่ยังไม่เป็นที่เข้าใจอย่างถ่องแท้ ความเข้าใจในปัจจุบัน [ 25 ]คือ นิวคลีโอโซมที่ซ้ำกันโดยมี DNA "ตัวเชื่อม" คั่นกลางจะก่อตัวเป็นเส้นใยขนาด 10 นาโนเมตรซึ่งอธิบายว่าเป็น "ลูกปัดบนเส้นเชือก" และมีอัตราส่วนการบรรจุประมาณห้าถึงสิบ[ 18 ]สายโซ่ของนิวคลีโอโซมสามารถจัดเรียงเป็นเส้นใยขนาด 30 นาโนเมตรซึ่งเป็นโครงสร้างที่อัดแน่นโดยมีอัตราส่วนการบรรจุประมาณ 50 [ 18 ]และการก่อตัวนั้นขึ้นอยู่กับการมีอยู่ของ ฮิ ส โตน H1
โครงสร้างผลึกของเตตระนิวคลีโอโซมได้รับการนำเสนอและนำมาใช้สร้างโครงสร้างที่เสนอของเส้นใย 30 นาโนเมตรเป็นเกลียวสองวง[ 34 ] ยังคงมีการโต้แย้งกันอยู่บ้างเกี่ยวกับแบบจำลองนี้ เนื่องจากไม่สอดคล้องกับข้อมูลกล้องจุลทรรศน์อิเล็กตรอน ล่าสุด [ 35 ] นอกเหนือจากนี้ โครงสร้างของโครมาตินยังไม่เป็นที่เข้าใจอย่างถ่องแท้ แต่โดยทั่วไปแล้วมีการเสนอว่าเส้นใย 30 นาโนเมตรจะเรียงตัวเป็นวงรอบตามโครงสร้างโปรตีนส่วนกลางเพื่อสร้าง ยูโครมาติน ที่ทำงานด้านการถอดรหัสการอัดแน่นเพิ่มเติมจะนำไปสู่เฮเทอโรโครมาตินที่ ไม่ทำงานด้านการถอดรหัส
พลวัต
แม้ว่านิวคลีโอโซมจะเป็นคอมเพล็กซ์โปรตีน-ดีเอ็นเอที่เสถียรมาก แต่ก็ไม่ได้คงที่และได้รับการแสดงให้เห็นว่ามีการเปลี่ยนแปลงโครงสร้างหลายอย่าง รวมถึงการเลื่อนของนิวคลีโอโซมและการเปิดเผยตำแหน่งดีเอ็นเอ ขึ้นอยู่กับบริบท นิวคลีโอโซมสามารถยับยั้งหรืออำนวยความสะดวกในการจับตัวของปัจจัยการถอดรหัสได้ ตำแหน่งของนิวคลีโอโซมถูกควบคุมโดยปัจจัยหลักสามประการ: ประการแรก ความสัมพันธ์ในการจับตัวโดยธรรมชาติของฮิสโตนออกตาเมอร์ขึ้นอยู่กับลำดับดีเอ็นเอ ประการที่สอง นิวคลีโอโซมสามารถถูกแทนที่หรือดึงดูดได้โดยการจับตัวแบบแข่งขันหรือแบบร่วมมือของปัจจัยโปรตีนอื่นๆ ประการที่สาม นิวคลีโอโซมอาจถูกเคลื่อนย้ายอย่างแข็งขันโดยคอมเพล็กซ์การปรับโครงสร้างที่ขึ้นอยู่กับ ATP [ 36 ]
การเลื่อนของนิวคลีโอโซม
เมื่อบ่มด้วยความร้อน นิวคลีโอโซมที่สร้างขึ้นใหม่บนลำดับตำแหน่ง DNA 5S สามารถจัดตำแหน่งตัวเองใหม่ได้โดยการเลื่อนไปยังลำดับที่อยู่ติดกัน[ 37 ]การจัดตำแหน่งใหม่นี้ไม่จำเป็นต้องทำลายฮิสโตนออกตาเมอร์ แต่สอดคล้องกับความสามารถของนิวคลีโอโซมในการ "เลื่อน" ไปตาม DNA ในแบบซิสตำแหน่ง การจับ ของ CTCFทำหน้าที่เป็นจุดยึดตำแหน่งนิวคลีโอโซม ดังนั้น เมื่อใช้ในการจัดเรียงสัญญาณจีโนมต่างๆ นิวคลีโอโซมที่อยู่รอบข้างหลายตัวจึงสามารถระบุได้ง่าย[ 38 ]แม้ว่านิวคลีโอโซมจะเคลื่อนที่ได้โดยธรรมชาติ แต่ยูคาริโอตได้พัฒนาเอนไซม์ปรับโครงสร้างโครมาตินที่ขึ้นอยู่กับ ATP จำนวนมากเพื่อเปลี่ยนแปลงโครงสร้างโครมาติน ซึ่งหลายเอนไซม์ทำเช่นนั้นผ่านการเลื่อนของนิวคลีโอโซม การเลื่อนของนิวคลีโอโซมเป็นหนึ่งในกลไกที่เป็นไปได้สำหรับการแสดงออกของยีนเฉพาะเนื้อเยื่อในวงกว้าง ตำแหน่งเริ่มต้นการถอดรหัสของยีนที่แสดงออกในเนื้อเยื่อเฉพาะนั้นจะมีนิวคลีโอโซมลดลง ในขณะที่ยีนชุดเดียวกันในเนื้อเยื่ออื่นที่ไม่ได้แสดงออกนั้นจะมีนิวคลีโอโซมจับอยู่[ 39 ]
การเปิดเผยตำแหน่ง DNA
ดีเอ็นเอในนิวคลีโอโซมอยู่ในสภาวะสมดุลระหว่างสถานะพันและคลายตัว ดีเอ็นเอภายในนิวคลีโอโซมจะพันกันอย่างสมบูรณ์เป็นเวลาเพียง 250 มิลลิวินาทีก่อนที่จะคลายตัวเป็นเวลา 10-50 มิลลิวินาที แล้วจึงพันกันใหม่อย่างรวดเร็ว โดยวัดด้วย FRET แบบเวลาจำเพาะ[ 40 ]ซึ่งหมายความว่าดีเอ็นเอไม่จำเป็นต้องแยกตัวออกจากนิวคลีโอโซมอย่างแข็งขัน แต่มีช่วงเวลาสำคัญที่สามารถเข้าถึงได้อย่างสมบูรณ์ การนำลำดับการจับดีเอ็นเอเข้าไปในนิวคลีโอโซมจะเพิ่มการเข้าถึงบริเวณที่อยู่ติดกันของดีเอ็นเอเมื่อถูกจับ[ 41 ]
แนวโน้มที่ DNA ภายในนิวคลีโอโซมจะ "หายใจ" นั้นมีผลกระทบเชิงฟังก์ชันที่สำคัญต่อโปรตีนที่จับกับ DNA ทั้งหมดที่ทำงานในสภาพแวดล้อมของโครมาติน[ 40 ]โดยเฉพาะอย่างยิ่ง การหายใจแบบไดนามิกของนิวคลีโอโซมมีบทบาทสำคัญในการจำกัดความก้าวหน้าของRNA polymerase IIในระหว่างการยืดตัวของการถอดรหัส[ 42 ]
บริเวณที่ปราศจากนิวคลีโอโซม
โปรโมเตอร์ของยีนที่ทำงานอยู่จะมีบริเวณที่ปราศจากนิวคลีโอโซม (NFR) ซึ่งช่วยให้โปรตีนต่างๆ เช่น ปัจจัยการถอดรหัส สามารถเข้าถึง DNA ของโปรโมเตอร์ได้ โดยทั่วไปบริเวณที่ปราศจากนิวคลีโอโซมจะมีความยาวประมาณ 200 นิวคลีโอไทด์ในS. cerevisiae [ 43 ]นิวคลีโอโซมที่อยู่ในตำแหน่งที่เหมาะสมจะสร้างขอบเขตของ NFR นิวคลีโอโซมเหล่านี้เรียกว่า +1-นิวคลีโอโซมและ −1-นิวคลีโอโซม และตั้งอยู่ที่ระยะทางมาตรฐานทางด้านล่างและด้านบนตามลำดับจากตำแหน่งเริ่มต้นการถอดรหัส[ 44 ] +1-นิวคลีโอโซมและนิวคลีโอโซมด้านล่างหลายตัวมักจะรวมฮิสโตน H2A.Z เข้าไปด้วย[ 44 ]
การปรับเปลี่ยนโครงสร้างนิวคลีโอโซม
จีโนมของสิ่งมีชีวิตยูคาริโอตจะรวมตัวกันเป็นโครมาตินอย่างแพร่หลาย อย่างไรก็ตาม เซลล์ต้องควบคุมตำแหน่งเฉพาะเจาะจงทั้งในเชิงพื้นที่และเวลาอย่างอิสระจากโครมาตินโดยรวม เพื่อให้ได้ระดับการควบคุมที่สูงซึ่งจำเป็นต่อการประสานกระบวนการภายในนิวเคลียส เช่น การจำลองดีเอ็นเอ การซ่อมแซม และการถอดรหัส เซลล์จึงได้พัฒนากลไกต่างๆ เพื่อปรับเปลี่ยนโครงสร้างและหน้าที่ของโครมาตินในระดับเฉพาะที่และเฉพาะเจาะจง ซึ่งอาจเกี่ยวข้องกับการดัดแปลงฮิสโตนด้วยพันธะโควาเลนต์ การรวมฮิสโตนชนิดต่างๆ และการปรับโครงสร้างแบบไม่ใช้พันธะโควาเลนต์โดยเอนไซม์ปรับโครงสร้างที่ขึ้นอยู่กับ ATP
การดัดแปลงฮิสโตนหลังการสังเคราะห์โปรตีน

นับตั้งแต่มีการค้นพบการดัดแปลงฮิสโตนในช่วงกลางทศวรรษ 1960 ก็มีการคาดการณ์ว่าการดัดแปลงฮิสโตนจะส่งผลต่อการถอดรหัส[ 45 ]ข้อเท็จจริงที่ว่าการดัดแปลงหลังการแปลส่วนใหญ่ที่พบในช่วงแรกนั้นกระจุกตัวอยู่ในส่วนต่อขยายหางที่ยื่นออกมาจากแกนกลางของนิวคลีโอโซม นำไปสู่ทฤษฎีหลักสองทฤษฎีเกี่ยวกับกลไกของการดัดแปลงฮิสโตน ทฤษฎีแรกเสนอว่าการดัดแปลงเหล่านี้อาจส่งผลต่อปฏิกิริยาไฟฟ้าสถิตระหว่างหางฮิสโตนและ DNA เพื่อ "คลาย" โครงสร้างโครมาติน ต่อมามีการเสนอว่าการรวมกันของการดัดแปลงเหล่านี้อาจสร้างเอพิโทปการจับเพื่อดึงดูดโปรตีนอื่นๆ[ 46 ]เมื่อเร็วๆ นี้ เนื่องจากมีการค้นพบการดัดแปลงเพิ่มเติมในบริเวณที่มีโครงสร้างของฮิสโตน จึงมีการเสนอว่าการดัดแปลงเหล่านี้อาจส่งผลต่อปฏิกิริยาระหว่างฮิสโตนกับ DNA [ 47 ]และฮิสโตนกับฮิสโตน[ 48 ]ภายในแกนกลางของนิวคลีโอโซม การดัดแปลง (เช่น การอะเซทิเลชันหรือการฟอสโฟรีเลชัน) ที่ลดประจุของแกนฮิสโตนทรงกลมคาดว่าจะ "คลาย" การเชื่อมโยงระหว่างแกนกับ DNA ความแรงของผลกระทบขึ้นอยู่กับตำแหน่งของการดัดแปลงภายในแกน[ 49 ] การดัดแปลงบางอย่างแสดงให้เห็นว่ามีความสัมพันธ์กับการปิดการทำงานของยีนในขณะที่บางอย่างดูเหมือนจะมีความสัมพันธ์กับการเปิดใช้งานยีน การดัดแปลงทั่วไป ได้แก่การอะเซทิเลชัน การเมทิลเลชันหรือการยูบิควิติเนชันของไลซีนการเมทิลเลชันของอาร์จินีนและการฟอสโฟรีเลชันของซีรีน ข้อมูลที่ จัดเก็บในลักษณะนี้ถือว่าเป็นข้อมูลทางพันธุกรรมเนื่องจากไม่ได้ถูกเข้ารหัสใน DNA แต่ยังคงถ่ายทอดไปยังเซลล์ลูก การรักษาสถานะที่ถูกกดหรือเปิดใช้งานของยีนมักมีความจำเป็นสำหรับการแบ่งเซลล์ [ 18 ]
ฮิสโตนชนิดต่างๆ
แม้ว่าฮิสโตนจะได้รับการอนุรักษ์ไว้อย่างน่าทึ่งตลอดวิวัฒนาการ แต่ก็มีการระบุรูปแบบที่แตกต่างกันหลายรูปแบบ การกระจายตัวของฟังก์ชันฮิสโตนนี้จำกัดอยู่ที่ H2A และ H3 โดยที่ H2B และ H4 ส่วนใหญ่ไม่เปลี่ยนแปลง H2A สามารถถูกแทนที่ด้วยH2AZ (ซึ่งนำไปสู่ความเสถียรของนิวคลีโอโซมที่ลดลง) หรือH2AX (ซึ่งเกี่ยวข้องกับการซ่อมแซม DNA และ การแยกความแตกต่าง ของเซลล์ T ) ในขณะที่โครโมโซม X ที่ไม่ทำงานในสัตว์เลี้ยงลูกด้วยนมจะอุดมไปด้วย macroH2A H3 สามารถถูกแทนที่ด้วย H3.3 (ซึ่งสัมพันธ์กับยีนที่ทำงานและองค์ประกอบควบคุม) และในเซนโทรเมียร์ H3 จะถูกแทนที่ด้วยCENPA [ 18 ]
นอกจากนี้ไวรัสยังสามารถสร้างอนุภาคนิวคลีโอโซมที่ประกอบด้วยฮิสโตนแบบแคนอนิกหลายรูปแบบ[ 50 ]ซึ่งบางครั้งจะหลอมรวมกันเป็นคู่ในสายโพลีเปปไทด์เดียวกัน[ 51 ]จึงทำให้เกิด โครงสร้างฮิส โตน คู่ขึ้น
การปรับโครงสร้างนิวคลีโอโซมที่ขึ้นอยู่กับ ATP
ปฏิกิริยาที่แตกต่างกันหลายอย่างเกี่ยวข้องกับคำว่าการปรับโครงสร้างโครมาตินที่ขึ้นอยู่กับ ATPเอนไซม์ปรับโครงสร้างแสดงให้เห็นว่าสามารถเลื่อนนิวคลีโอโซมไปตาม DNA [ 52 ]ทำลายการสัมผัสระหว่างฮิสโตนกับ DNA จนถึงขั้นทำให้ไดเมอร์ H2A/H2B ไม่เสถียร[ 53 ] [ 54 ]และสร้างการบิดตัวแบบซูเปอร์เฮลิกซ์เชิงลบใน DNA และโครมาติน[ 55 ]เมื่อเร็วๆ นี้ พบว่าเอนไซม์ปรับโครงสร้าง Swr1 สามารถนำฮิสโตน H2A.Z ที่แตกต่างกันเข้าไปในนิวคลีโอโซมได้[ 56 ]ในปัจจุบัน ยังไม่ชัดเจนว่าปฏิกิริยาเหล่านี้ทั้งหมดเป็นปฏิกิริยาที่แตกต่างกันหรือเป็นเพียงผลลัพธ์ทางเลือกของกลไกทั่วไป สิ่งที่เหมือนกันระหว่างปฏิกิริยาทั้งหมด และเป็นลักษณะเด่นของการปรับโครงสร้างโครมาตินที่ขึ้นอยู่กับ ATP ก็คือ ปฏิกิริยาทั้งหมดส่งผลให้การเข้าถึง DNA เปลี่ยนแปลงไป
การศึกษาที่พิจารณาการกระตุ้นยีนในร่างกาย[ 57 ]และที่น่าประหลาดใจยิ่งกว่านั้นคือการปรับโครงสร้างในหลอดทดลอง[ 58 ]ได้เปิดเผยว่าเหตุการณ์การปรับโครงสร้างโครมาตินและการจับตัวของปัจจัยการถอดรหัสมีลักษณะเป็นวัฏจักรและเป็นคาบ แม้ว่าผลที่ตามมาของสิ่งนี้ต่อกลไกปฏิกิริยาของการปรับโครงสร้างโครมาตินจะยังไม่เป็นที่ทราบแน่ชัด แต่ลักษณะไดนามิกของระบบอาจทำให้สามารถตอบสนองต่อสิ่งเร้าภายนอกได้เร็วขึ้น การศึกษาล่าสุดระบุว่าตำแหน่งของนิวคลีโอโซมเปลี่ยนแปลงอย่างมีนัยสำคัญในระหว่างการพัฒนาเซลล์ต้นกำเนิดตัวอ่อนของหนู และการเปลี่ยนแปลงเหล่านี้เกี่ยวข้องกับการจับตัวของปัจจัยการถอดรหัสในการพัฒนา[ 59 ]
การปรับเปลี่ยนโครงสร้างนิวคลีโอโซมแบบไดนามิกทั่วทั้งจีโนมของยีสต์
การศึกษาในปี 2550 ได้จัดทำรายการตำแหน่งนิวคลีโอโซมในยีสต์และแสดงให้เห็นว่านิวคลีโอโซมมีจำนวนน้อยลงใน บริเวณ โปรโมเตอร์และจุดเริ่มต้นของการจำลองแบบ [ 60 ] [ 61 ] [ 62 ] ประมาณ 80% ของจีโนมยีสต์ดูเหมือนจะถูกปกคลุมด้วยนิวคลีโอโซม[ 63 ]และรูปแบบของตำแหน่งนิวคลีโอโซมมีความสัมพันธ์อย่างชัดเจนกับบริเวณ DNA ที่ควบคุมการถอดรหัสบริเวณที่ถูกถอดรหัส และบริเวณที่เริ่มต้นการจำลองแบบ DNA [ 64 ]ล่าสุด การศึกษาใหม่ได้ตรวจสอบการเปลี่ยนแปลงแบบไดนามิกในการจัดตำแหน่งนิวคลีโอโซมใหม่ระหว่างเหตุการณ์การถอดรหัสใหม่ทั่วโลกเพื่อชี้แจงผลกระทบต่อการเคลื่อนที่ของนิวคลีโอโซมระหว่างการเปลี่ยนแปลงการถอดรหัสทั่วทั้งจีโนมในยีสต์ ( Saccharomyces cerevisiae ) [ 65 ]ผลลัพธ์ชี้ให้เห็นว่านิวคลีโอโซมที่อยู่เฉพาะที่บริเวณโปรโมเตอร์จะถูกเคลื่อนย้ายเพื่อตอบสนองต่อความเครียด (เช่นความร้อนช็อก ) นอกจากนี้ การกำจัดนิวคลีโอโซมมักจะสอดคล้องกับการกระตุ้นการถอดรหัส และการแทนที่นิวคลีโอโซมมักจะสอดคล้องกับการยับยั้งการถอดรหัส ซึ่งสันนิษฐานว่าเป็นเพราะ ตำแหน่งการจับ ของปัจจัยการถอดรหัสสามารถเข้าถึงได้มากขึ้นหรือน้อยลงตามลำดับ โดยทั่วไปแล้ว จะมีการจัดตำแหน่งนิวคลีโอโซมใหม่เพียงหนึ่งหรือสองตัวที่โปรโมเตอร์เพื่อทำให้เกิดการเปลี่ยนแปลงการถอดรหัสเหล่านี้ อย่างไรก็ตาม แม้ในบริเวณโครโมโซมที่ไม่เกี่ยวข้องกับการเปลี่ยนแปลงการถอดรหัส ก็ยังพบการจัดตำแหน่งนิวคลีโอโซมใหม่ ซึ่งบ่งชี้ว่าการปกคลุมและการเปิดเผย DNA ที่เกี่ยวข้องกับการถอดรหัสไม่จำเป็นต้องก่อให้เกิดเหตุการณ์การถอดรหัสเสมอไป หลังจากการถอดรหัส บริเวณ rDNA จะต้องได้รับการปกป้องจากความเสียหายใดๆ ซึ่งบ่งชี้ว่าโปรตีน HMGB มีบทบาทสำคัญในการปกป้องบริเวณที่ปราศจากนิวคลีโอโซม[ 66 ] [ 67 ]
ข้อบกพร่องการบิดตัวของ DNA
ข้อบกพร่องการบิดของ DNA เกิดขึ้นเมื่อมีการถ่ายโอนคู่เบสหนึ่งหรือสองคู่จากส่วนของ DNA หนึ่งไปยังส่วนถัดไป ส่งผลให้การบิดของ DNA เปลี่ยนแปลงไป ซึ่งไม่เพียงแต่จะเปลี่ยนการบิดของ DNA เท่านั้น แต่ยังเปลี่ยนความยาวอีกด้วย[ 68 ]ข้อบกพร่องการบิดนี้จะเคลื่อนที่ไปรอบๆ นิวคลีโอโซมผ่านการถ่ายโอนคู่เบส ซึ่งหมายความว่าการบิดของ DNA สามารถทำให้เกิดการเลื่อนของนิวคลีโอโซมได้[ 69 ]โครงสร้างผลึกของนิวคลีโอโซมแสดงให้เห็นว่าตำแหน่งซูเปอร์เฮลิกซ์ 2 และ 5 บนนิวคลีโอโซมมักพบว่าเป็นตำแหน่งที่เกิดข้อบกพร่องการบิดของ DNA เนื่องจากเป็นตำแหน่งการจับของรีโมเดลเลอร์ทั่วไป[ 70 ]มีรีโมเดลเลอร์โครมาตินหลายชนิด แต่ทั้งหมดมีมอเตอร์ ATPase ร่วมกัน ซึ่งอำนวยความสะดวกในการเลื่อนของโครมาตินบน DNA ผ่านการจับและการไฮโดรไลซิสของ ATP [ 71 ] ATPase มีสถานะเปิดและปิด เมื่อมอเตอร์ ATPase เปลี่ยนจากสถานะเปิดเป็นปิด รูปทรงของ DNA duplex จะเปลี่ยนและแสดงการเอียงของคู่เบส[ 70 ]การเริ่มต้นของข้อบกพร่องการบิดผ่านมอเตอร์ ATPase ทำให้เกิดแรงตึงสะสมรอบบริเวณตัวปรับโครงสร้าง แรงตึงจะถูกปลดปล่อยเมื่อการเลื่อนของ DNA เสร็จสมบูรณ์ทั่วทั้งนิวคลีโอโซมผ่านการแพร่กระจายของข้อบกพร่องการบิดสองจุด (จุดหนึ่งบนแต่ละสาย) ในทิศทางตรงกันข้าม[ 71 ]
การประกอบนิวคลีโอโซมในหลอดทดลอง

นิวคลีโอโซมสามารถประกอบขึ้นในหลอดทดลองได้โดยใช้ฮิสโตนบริสุทธิ์จากธรรมชาติหรือฮิสโตนรีคอมบิแนนท์[ 72 ] [ 73 ]เทคนิคมาตรฐานอย่างหนึ่งในการโหลด DNA รอบฮิสโตนเกี่ยวข้องกับการใช้การฟอก ด้วยเกลือ ปฏิกิริยาที่ประกอบด้วยฮิสโตนออกตาเมอร์และแม่แบบ DNA เปลือยสามารถบ่มร่วมกันได้ที่ความเข้มข้นของเกลือ 2 M เมื่อค่อยๆ ลดความเข้มข้นของเกลือลง DNA จะปรับสมดุลไปยังตำแหน่งที่มันพันรอบฮิสโตนออกตาเมอร์ ทำให้เกิดนิวคลีโอโซมขึ้น ภายใต้สภาวะที่เหมาะสม กระบวนการสร้างใหม่นี้ช่วยให้สามารถกำหนดตำแหน่งความสัมพันธ์ของนิวคลีโอโซมของลำดับที่กำหนดได้จากการทดลอง[ 74 ]
อนุภาคแกนกลางนิวคลีโอโซมที่เชื่อมโยงด้วยพันธะไดซัลไฟด์
ความก้าวหน้าล่าสุดในการผลิตอนุภาคแกนนิวคลีโอโซมที่มีเสถียรภาพเพิ่มขึ้นเกี่ยวข้องกับการเชื่อมโยงไดซัลไฟด์ เฉพาะตำแหน่ง [ 75 ]สามารถนำการเชื่อมโยงสองแบบที่แตกต่างกันเข้าไปในอนุภาคแกนนิวคลีโอโซมได้ การเชื่อมโยงแบบแรกเชื่อมโยงสำเนาสองชุดของH2Aผ่านซิสเทอีน (N38C) ที่นำเข้ามา ส่งผลให้เกิดฮิสโตนออกตาเมอร์ซึ่งมีเสถียรภาพต่อการสูญเสียไดเมอร์ H2A/H2B ระหว่างการสร้างนิวคลีโอโซมขึ้นใหม่ การเชื่อมโยงแบบที่สองสามารถนำเข้ามาระหว่างหางฮิสโตนปลาย N ของ H3 และปลาย DNA ของนิวคลีโอโซมผ่านนิวคลีโอไทด์ที่แปลงสภาพได้ที่ถูกรวมเข้าไป[ 76 ]การเชื่อมโยง DNA-ฮิสโตนออกตาเมอร์ทำให้เสถียรภาพของอนุภาคแกนนิวคลีโอโซมต่อการแยกตัวของ DNA ที่ความเข้มข้นของอนุภาคต่ำมากและที่ความเข้มข้นของเกลือสูง
การประกอบนิวคลีโอโซมในร่างกาย

นิวคลีโอโซมเป็นหน่วยบรรจุพื้นฐานของดีเอ็นเอในจีโนม สร้างขึ้นจากโปรตีนฮิสโตนโดยมีดีเอ็นเอขดตัวอยู่รอบๆ นิวคลีโอโซมทำหน้าที่เป็นโครงสร้างค้ำจุนสำหรับการสร้างโครงสร้างโครมาตินระดับสูงขึ้น รวมถึงเป็นชั้นควบคุมการแสดงออกของยีนด้วย นิวคลีโอโซมจะถูกประกอบขึ้นอย่างรวดเร็วบนดีเอ็นเอที่สังเคราะห์ขึ้นใหม่หลังจุดแยกการจำลองแบบ (replication fork)
เอช3 และ เอช4
ฮิสโตนH3และH4จากนิวคลีโอโซมเก่าที่ถูกแยกส่วนจะถูกเก็บไว้ในบริเวณใกล้เคียงและกระจายแบบสุ่มบน DNA ที่สังเคราะห์ขึ้นใหม่[ 77 ]พวกมันถูกประกอบขึ้นโดยคอมเพล็กซ์ปัจจัยการประกอบโครมาติน 1 (CAF-1) ซึ่งประกอบด้วยสามหน่วยย่อย (p150, p60 และ p48) [ 78 ] H3 และ H4 ที่สังเคราะห์ขึ้นใหม่จะถูกประกอบขึ้นโดยปัจจัยการประกอบการจับคู่การจำลองแบบ (RCAF) RCAF ประกอบด้วยหน่วยย่อย Asf1 ซึ่งจับกับโปรตีน H3 และ H4 ที่สังเคราะห์ขึ้นใหม่[ 79 ] โปรตีน H3 และ H4 เก่าจะยังคงรักษาการดัดแปลงทางเคมีไว้ ซึ่งมีส่วนช่วยในการส่งต่อลายเซ็นเอพิเจเนติกส์ โปรตีน H3 และ H4 ที่สังเคราะห์ขึ้นใหม่จะค่อยๆ ถูกอะเซทิเลตที่กรดอะมิโนไลซีนต่างๆ ซึ่งเป็นส่วนหนึ่งของกระบวนการเจริญเติบโตของโครมาติน[ 80 ]เชื่อกันว่าโปรตีน H3 และ H4 เก่าในนิวคลีโอโซมใหม่จะดึงดูดเอนไซม์ปรับเปลี่ยนฮิสโตนที่ทำเครื่องหมายฮิสโตนใหม่ ซึ่งมีส่วนช่วยในการสร้างความทรงจำทางเอพิเจเนติกส์
เอช2เอ และ เอช2บี
ตรงกันข้ามกับ H3 และ H4 เก่า โปรตีนฮิสโตน H2AและH2B เก่า จะถูกปล่อยออกมาและย่อยสลาย ดังนั้นโปรตีน H2A และ H2B ที่ประกอบขึ้นใหม่จึงถูกรวมเข้ากับนิวคลีโอโซมใหม่[ 81 ] H2A และ H2B จะถูกประกอบเป็นไดเมอร์ จากนั้นจะถูกโหลดลงบนนิวคลีโอโซมโดยโปรตีนประกอบนิวคลีโอโซม-1 (NAP-1) ซึ่งช่วยในการเลื่อนของนิวคลีโอโซมด้วย[ 82 ]นิวคลีโอโซมยังถูกเว้นระยะห่างโดยคอมเพล็กซ์การปรับโครงสร้างนิวคลีโอโซมที่ขึ้นอยู่กับ ATP ซึ่งประกอบด้วยเอนไซม์ เช่น Isw1 Ino80 และ Chd1 และต่อมาประกอบเป็นโครงสร้างลำดับสูงขึ้น[ 83 ] [ 84 ]
แกลเลอรี่


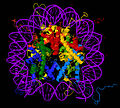
โครงสร้างผลึกของอนุภาคแกนกลางนิวคลีโอโซม ( PDB : 1EQZ [28 ] ) - มุมมองต่างๆ ที่แสดงรายละเอียดของการพับและการจัดระเบียบของฮิสโตน ฮิสโตนH2A , H2B , H3 , H4และDNAถูกระบายสี
ดูเพิ่มเติม
ลิงก์ภายนอก
- MBInfo - นิวคลีโอโซมคืออะไร
- นิวคลีโอโซม บนเว็บไซต์ Richmond Lab
- นิวคลีโอโซมของโปรตีโอพีเดีย
- นิวคลีโอโซมที่ PDB
- การปรับเปลี่ยนโครงสร้างแบบไดนามิกของนิวคลีโอโซมแต่ละตัวทั่วทั้งจีโนมของสิ่งมีชีวิตยูคาริโอตเพื่อตอบสนองต่อการรบกวนการถอดรหัส
- ข้อมูลและเครื่องมือระบุตำแหน่งนิวคลีโอโซมออนไลน์ (รายการพร้อมคำอธิบาย อัปเดตอย่างต่อเนื่อง)
- โครงสร้างโปรตีนฮิสโตน
- HistoneDB 2.0 - ฐานข้อมูลฮิสโตนและรูปแบบต่างๆ ที่NCBI
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ นิวคลีโอโซม
นิวคลีโอโซมเป็นหน่วยโครงสร้างพื้นฐานของ การบรรจุ DNAในยูคาริโอตโครงสร้างของนิวคลีโอโซมประกอบด้วยส่วนของ DNA ที่พันรอบโปรตีนฮิสโตน แปดตัว และมีลักษณะคล้ายเส้นด้ายที่พันรอบแกนหมุน...
โครงสร้างของอนุภาคแกนกลาง
การศึกษาโครงสร้างบุกเบิกในช่วงทศวรรษ 1980 โดยกลุ่มของ Aaron Klug ได้ให้หลักฐานแรกว่าโปรตีนฮิสโตนแปดตัวพันรอบ DNA เองในเกลียวซูเปอร์เฮลิกซ์แบบซ้ายประมาณ 1.
โครงสร้างลำดับสูงกว่า
การจัดระเบียบของ DNA ที่เกิดขึ้นโดยนิวคลีโอโซมไม่สามารถอธิบายการบรรจุ DNA ที่สังเกตได้ในนิวเคลียสของเซลล์ได้อย่างสมบูรณ์จำเป็นต้อง มีการอัดแน่นของ โครมาติน เข้าไปในนิวเคลียสของเซลล์มากขึ้น แต่ยังไม่เป็นที่เข้าใจอย่างถ่องแท้ ความเข้าใจในปัจจุบัน [ 25 ] คือ...
พลวัต
แม้ว่านิวคลีโอโซมจะเป็นคอมเพล็กซ์โปรตีน-ดีเอ็นเอที่เสถียรมาก แต่ก็ไม่ได้คงที่และได้รับการแสดงให้เห็นว่ามีการเปลี่ยนแปลงโครงสร้างหลายอย่าง รวมถึงการเลื่อนของนิวคลีโอโซมและการเปิดเผยตำแหน่งดีเอ็นเอ ขึ้นอยู่กับบริบท...