อ่าน 51 นาที
เอพิเจเนติกส์
เอพิเจเนติกส์ คือการศึกษาการเปลี่ยนแปลงใน การแสดงออกของยีน ที่เกิดขึ้นโดยไม่เปลี่ยนแปลงลำดับ ดีเอ็นเอ [ 1 ] คำนำ หน้าภาษากรีก epi- (ἐπι- "เหนือกว่า, ภายนอก, รอบๆ") ใน...
เอพิเจเนติกส์

เอพิเจเนติกส์คือการศึกษาการเปลี่ยนแปลงในการแสดงออกของยีนที่เกิดขึ้นโดยไม่เปลี่ยนแปลงลำดับดีเอ็นเอ[ 1 ] คำนำ หน้าภาษากรีกepi- (ἐπι- "เหนือกว่า, ภายนอก, รอบๆ") ในเอพิเจเนติกส์หมายถึงคุณลักษณะที่ "อยู่เหนือ" หรือ "เพิ่มเติมจาก" กลไกการถ่ายทอดทางพันธุกรรมแบบดั้งเดิมที่อิงตามลำดับดีเอ็นเอ[ 2 ]เอพิเจเนติกส์มักเกี่ยวข้องกับการเปลี่ยนแปลงที่คงอยู่ตลอดการแบ่งเซลล์ และส่งผลต่อการควบคุมการแสดงออกของยีน[ 3 ]ผลกระทบดังกล่าวต่อลักษณะทางเซลล์และทางสรีรวิทยาอาจเป็นผลมาจากปัจจัยด้านสิ่งแวดล้อม หรือเป็นส่วนหนึ่งของการพัฒนาตามปกติ
คำนี้ยังหมายถึงกลไกเบื้องหลังการเปลี่ยนแปลงเหล่านี้ด้วย นั่นคือ การเปลี่ยนแปลงที่เกี่ยวข้องกับการทำงานของจีโนมที่ไม่เกี่ยวข้องกับการกลายพันธุ์ใน ลำดับนิวคลีโอไทด์ตัวอย่างของกลไกที่ทำให้เกิดการเปลี่ยนแปลงดังกล่าว ได้แก่การเมทิลเลชั่นของ DNAและการดัดแปลงฮิสโตนซึ่งแต่ละอย่างจะเปลี่ยนแปลงวิธีการแสดงออกของยีนโดยไม่เปลี่ยนแปลงลำดับDNA พื้นฐาน [ 4 ]นอกจากนี้ ยังพบว่าลำดับ RNA ที่ไม่เข้ารหัสมีบทบาทสำคัญในการควบคุมการแสดงออกของยีน[ 5 ]การแสดงออกของยีนสามารถควบคุมได้ผ่านการทำงานของโปรตีนยับยั้งที่ยึดติดกับ บริเวณ เงียบของ DNA การเปลี่ยนแปลงทางเอพิเจเนติกส์เหล่านี้อาจคงอยู่ตลอดการแบ่งเซลล์ตลอดช่วงชีวิตของเซลล์ และอาจคงอยู่ได้หลายชั่วอายุคน แม้ว่าจะไม่เกี่ยวข้องกับการเปลี่ยนแปลงในลำดับ DNA พื้นฐานของสิ่งมีชีวิตก็ตาม[ 6 ]แต่ปัจจัยที่ไม่ใช่พันธุกรรมทำให้ยีนของสิ่งมีชีวิตมีพฤติกรรม (หรือ "แสดงออก") ที่แตกต่างกัน[ 7 ]
ตัวอย่างหนึ่งของการเปลี่ยนแปลงเอพิเจเนติกใน ชีววิทยา ของยูคาริโอตคือกระบวนการแบ่งเซลล์ในระหว่างการสร้างรูปร่างเซลล์ต้นกำเนิดที่มีศักยภาพ ทั้งหมดจะกลายเป็นเซลล์ ที่มีศักยภาพหลายอย่างในตัวอ่อนซึ่งต่อมาจะกลายเป็นเซลล์ที่แบ่งตัวอย่างสมบูรณ์ กล่าวอีกนัยหนึ่งคือ เมื่อเซลล์ไข่ ที่ได้รับการปฏิสนธิเพียงเซลล์เดียว – ไซโกต – แบ่งตัวต่อไปเซลล์ลูกที่เกิดขึ้นจะพัฒนาไปเป็นเซลล์ประเภทต่างๆ ในสิ่งมีชีวิต รวมถึงเซลล์ประสาทเซลล์กล้ามเนื้อ เนื้อเยื่อบุผิวเยื่อบุผนังหลอดเลือดฯลฯโดยการกระตุ้นยีนบางตัวในขณะที่ยับยั้งการแสดงออกของยีนอื่นๆ[ 8 ]
คำจำกัดความ
คำว่าepigenesisมีความหมายทั่วไปว่า "การเจริญเติบโตพิเศษ" ซึ่งใช้ในภาษาอังกฤษมาตั้งแต่ศตวรรษที่ 17 [ 9 ]ในสิ่งพิมพ์ทางวิทยาศาสตร์ คำว่าepigeneticsเริ่มปรากฏในทศวรรษที่ 1930 (ดูรูปทางด้านขวา) อย่างไรก็ตาม ความหมายร่วมสมัยของคำนี้เพิ่งปรากฏขึ้นในทศวรรษที่ 1990 [ 10 ]

นิยามของแนวคิดลักษณะทางพันธุกรรมแบบเอพิเจเนติกส์ว่าเป็น "ฟีโนไทป์ที่ถ่ายทอดทางพันธุกรรมได้อย่างเสถียรซึ่งเป็นผลมาจากการเปลี่ยนแปลงในโครโมโซมโดยไม่มีการเปลี่ยนแปลงในลำดับดีเอ็นเอ" ได้รับการกำหนดขึ้นใน การประชุม Cold Spring Harborในปี 2551 [ 11 ]แม้ว่านิยามทางเลือกที่รวมถึงลักษณะที่ไม่สามารถถ่ายทอดทางพันธุกรรมได้จะยังคงถูกนำมาใช้กันอย่างแพร่หลาย[ 12 ]
คลอง Waddington ในทศวรรษ 1940
สมมติฐานเกี่ยวกับการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่มีผลต่อการแสดงออกของโครโมโซมนั้นถูกเสนอโดยนักชีววิทยาชาวรัสเซียNikolai Koltsov [ 13 ] จากความหมายทั่วไปและคำคุณศัพท์ที่เกี่ยวข้องคือเอพิเจเนติกส์นักเอ็มบริโอวิทยาชาวอังกฤษCH Waddingtonได้บัญญัติศัพท์คำว่าเอพิเจเนติกส์ในปี 1942 โดยหมายถึงเอพิเจ เนซิส ในลักษณะเดียวกับ 'ฟีโนเจเนติกส์' ( Phänogenetik ) ของValentin Haecker [ 14 ]เอพิเจเนซิสในบริบทของชีววิทยาในยุคนั้นหมายถึงการแยกตัวของเซลล์จาก สถานะ โทติโพเทนต์เริ่ม ต้น ในระหว่างการพัฒนาของตัวอ่อน[ 15 ]
เมื่อแวดดิงตันบัญญัติศัพท์นี้ขึ้นมา ธรรมชาติทางกายภาพของยีนและบทบาทของยีนในการถ่ายทอดทางพันธุกรรมยังไม่เป็นที่รู้จัก เขาจึงใช้คำนี้เป็นแบบจำลองเชิงแนวคิดว่าองค์ประกอบทางพันธุกรรมอาจมีปฏิสัมพันธ์กับสิ่งแวดล้อมอย่างไรเพื่อสร้างฟีโนไทป์เขาใช้คำว่า " ภูมิทัศน์เอพิเจเนติกส์ " เป็นอุปมาอุปไมยสำหรับการพัฒนาทางชีววิทยาแวดดิงตันเชื่อว่าชะตากรรมของเซลล์ถูกกำหนดขึ้นในระหว่างการพัฒนาในกระบวนการที่เขาเรียกว่าการกำหนดทิศทางเหมือนกับลูกแก้วที่กลิ้งลงไปยังจุดที่ต่ำที่สุดในบริเวณนั้น [ 16 ] แวดดิงตันเสนอให้จินตนาการถึงความไม่สามารถย้อนกลับได้ที่เพิ่มขึ้นของการจำแนกประเภทของเซลล์เป็นสันเขาที่ยกตัวขึ้นระหว่างหุบเขาที่ลูกแก้ว (เปรียบเสมือนเซลล์) กำลังเคลื่อนที่[ 17 ]
เมื่อไม่นานมานี้ แนวคิดของ Waddington เกี่ยวกับภูมิทัศน์เอพิเจเนติกได้รับการกำหนดรูปแบบอย่างเข้มงวดในบริบทของแนวทาง สถานะ พลวัตของระบบในการศึกษาชะตากรรมของเซลล์[ 18 ] [ 19 ]การกำหนดชะตากรรมของเซลล์คาดว่าจะแสดงพลวัตบางอย่าง เช่น การบรรจบกันของตัวดึงดูด (ตัวดึงดูดอาจเป็นจุดสมดุล วงจรจำกัด หรือตัวดึงดูดแปลก ๆ ) หรือการแกว่ง[ 19 ]
ร่วมสมัย
ในปี พ.ศ. 2533 Robin Hollidayได้นิยาม epigenetics ว่า "การศึกษาเกี่ยวกับกลไกการควบคุมกิจกรรมของยีนตามเวลาและพื้นที่ในระหว่างการพัฒนาของสิ่งมีชีวิตที่ซับซ้อน" [ 20 ]
การใช้คำนี้ในชีววิทยาในช่วงหลังๆ เป็นไปตามคำจำกัดความที่เข้มงวดมากขึ้น ตามที่Arthur Riggsและเพื่อนร่วมงานได้นิยามไว้ว่าคือ "การศึกษา การเปลี่ยนแปลงทางพันธุกรรมของยีนที่ถ่ายทอดได้ โดยไมโทซิสและ/หรือไมโอซิสซึ่งไม่สามารถอธิบายได้ด้วยการเปลี่ยนแปลงในลำดับดีเอ็นเอ" [ 21 ]
อย่างไรก็ตาม คำนี้ยังถูกนำมาใช้เพื่ออธิบายกระบวนการที่ยังไม่ได้รับการพิสูจน์ว่าสามารถถ่ายทอดทางพันธุกรรมได้ เช่น การดัดแปลงฮิสโตนบางรูปแบบ ดังนั้นจึงมีความพยายามที่จะกำหนดนิยามใหม่ของ "เอพิเจเนติกส์" ในแง่มุมที่กว้างขึ้นเพื่อหลีกเลี่ยงข้อจำกัดของการต้องอาศัยการ ถ่ายทอดทางพันธุกรรม ตัวอย่างเช่นAdrian Birdได้นิยามเอพิเจเนติกส์ว่า "การปรับโครงสร้างของบริเวณโครโมโซมเพื่อบันทึก ส่งสัญญาณ หรือทำให้สถานะการทำงานที่เปลี่ยนแปลงไปคงอยู่" [ 6 ]นิยามนี้จะรวมถึงการดัดแปลงชั่วคราวที่เกี่ยวข้องกับการซ่อมแซม DNAหรือ ระยะ ของวงจรเซลล์ตลอดจนการเปลี่ยนแปลงที่คงที่ซึ่งคงอยู่ข้ามรุ่นเซลล์หลายรุ่น แต่ไม่รวมถึงสิ่งอื่นๆ เช่น การสร้างแบบจำลองของสถาปัตยกรรมเยื่อหุ้มเซลล์และพรีออนเว้นแต่ว่าสิ่งเหล่านั้นจะส่งผลกระทบต่อการทำงานของโครโมโซม อย่างไรก็ตาม การกำหนดนิยามใหม่ดังกล่าวไม่ได้เป็นที่ยอมรับโดยทั่วไปและยังคงเป็นหัวข้อถกเถียงกันอยู่[ 22 ] โครงการ "Roadmap Epigenomics Project" ของ NIHซึ่งดำเนินการตั้งแต่ปี 2008 ถึง 2017 ใช้คำจำกัดความดังต่อไปนี้: "สำหรับวัตถุประสงค์ของโปรแกรมนี้ เอพิเจเนติกส์หมายถึงทั้งการเปลี่ยนแปลงที่ถ่ายทอดได้ในกิจกรรมและการแสดงออก ของยีน (ในลูกหลานของเซลล์หรือของแต่ละบุคคล) และการเปลี่ยนแปลงที่เสถียรในระยะยาวในศักยภาพการถอดรหัสของเซลล์ที่ไม่จำเป็นต้องถ่ายทอดได้" [ 23 ]ในปี 2008 ได้มีการกำหนดคำจำกัดความที่เป็นเอกฉันท์ของลักษณะเอพิเจเนติกส์ ซึ่งก็คือ "ฟีโนไทป์ที่ถ่ายทอดได้อย่างเสถียรซึ่งเป็นผลมาจากการเปลี่ยนแปลงในโครโมโซมโดยไม่มีการเปลี่ยนแปลงในลำดับดีเอ็นเอ" ในการประชุม ที่ โคลด์สปริงฮาร์เบอร์[ 11 ]
ความคล้ายคลึงกันของคำกับ "พันธุศาสตร์" ทำให้เกิดการใช้งานคู่ขนานมากมาย " เอพิเจโนม " เป็นคำคู่ขนานกับคำว่า " จีโนม " ซึ่งหมายถึงสถานะเอพิเจเนติกโดยรวมของเซลล์ และเอพิเจโนมิกส์หมายถึงการวิเคราะห์การเปลี่ยนแปลงเอพิเจเนติกทั่วโลกทั่วทั้งจีโนม[ 12 ]วลี " รหัสพันธุกรรม " ก็ได้รับการดัดแปลงเช่นกัน โดย " รหัสเอพิเจเนติก " ถูกนำมาใช้เพื่ออธิบายชุดของคุณลักษณะเอพิเจเนติกที่สร้างฟีโนไทป์ที่แตกต่างกันในเซลล์ต่างๆ จากลำดับดีเอ็นเอพื้นฐานเดียวกัน หากนำไปใช้ในระดับสูงสุด "รหัสเอพิเจเนติก" อาจแสดงถึงสถานะโดยรวมของเซลล์ โดยตำแหน่งของแต่ละโมเลกุลจะถูกบันทึกไว้ในแผนที่เอพิเจโนมิกซึ่งเป็นการแสดงภาพแผนผังของการแสดงออกของยีน การเมทิลเลชั่นของดีเอ็นเอ และสถานะการดัดแปลงฮิสโตนของบริเวณจีโนมเฉพาะ โดยทั่วไปแล้ว คำนี้มักใช้ในการอ้างอิงถึงความพยายามอย่างเป็นระบบในการวัดข้อมูลเอพิเจเนติกที่เฉพาะเจาะจงและเกี่ยวข้อง เช่นรหัสฮิสโตนหรือรูปแบบ การเมทิลเลชั่นของดีเอ็นเอ
กลไก
การดัดแปลง โควาเลนต์ของ DNA (เช่น การเติมหมู่เมทิลและหมู่ไฮดรอกซีเมทิลของไซโตซีน) หรือของโปรตีนฮิสโตน (เช่น การเติมหมู่อะเซทิลของไลซีน การเติมหมู่เมทิลของไลซีนและอาร์จินีน การเติมหมู่ฟอสโฟริเลชันของเซรินและทรีโอนีน และการเติมหมู่ยูบิควิตินและหมู่ซูโมอิเลชันของไลซีน) มีบทบาทสำคัญในการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกหลายประเภท ดังนั้น คำว่า "เอพิเจเนติกส์" จึงถูกใช้เป็นคำพ้องความหมายสำหรับกระบวนการเหล่านี้ในบางครั้ง อย่างไรก็ตาม สิ่งนี้อาจทำให้เข้าใจผิดได้ การปรับโครงสร้างโครมาตินไม่ได้ถูกถ่ายทอดทางพันธุกรรมเสมอไป และการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์ไม่ได้เกี่ยวข้องกับการปรับโครงสร้างโครมาตินเสมอไป[ 24 ]ในปี 2019 มีการดัดแปลงไลซีนเพิ่มเติมปรากฏในเอกสารทางวิทยาศาสตร์ที่เชื่อมโยงการดัดแปลงเอพิเจเนติกส์กับการเผาผลาญของเซลล์ นั่นคือ การเติมหมู่แลคติล[ 25 ]

เนื่องจากฟีโนไทป์ของเซลล์หรือบุคคลได้รับผลกระทบจากยีนที่ถูกถอดรหัสสถานะการถอดรหัส ที่ถ่ายทอดทางพันธุกรรมจึงสามารถก่อให้เกิดผลกระทบทางเอพิเจเนติกส์ได้ การควบคุม การแสดงออกของยีนมีหลายระดับวิธีหนึ่งในการควบคุมยีนคือการปรับโครงสร้างของโครมาติน โครมาตินเป็นสารประกอบของดีเอ็นเอและ โปรตีน ฮิสโตนที่มันจับอยู่ด้วย หากวิธีการที่ดีเอ็นเอพันรอบฮิสโตนเปลี่ยนไป การแสดงออกของยีนก็สามารถเปลี่ยนแปลงได้เช่นกัน การปรับโครงสร้างโครมาตินเกิดขึ้นผ่านกลไกหลักสองอย่าง:
- วิธีแรกคือการดัดแปลงหลังการสังเคราะห์โปรตีนของกรดอะมิโนที่เป็นส่วนประกอบของโปรตีนฮิสโตน โปรตีนฮิสโตนประกอบด้วยสายโซ่ยาวของกรดอะมิโน หากกรดอะมิโนในสายโซ่เปลี่ยนไป รูปร่างของฮิสโตนอาจเปลี่ยนแปลงไปได้ ในระหว่างการจำลองดีเอ็นเอ ดีเอ็นเอจะไม่คลายตัวออกทั้งหมด ดังนั้นจึงเป็นไปได้ที่ฮิสโตนที่ถูกดัดแปลงแล้วอาจถูกนำไปยังสำเนาดีเอ็นเอใหม่แต่ละชุด เมื่อไปถึงที่นั่น ฮิสโตนเหล่านี้อาจทำหน้าที่เป็นแม่แบบ เริ่มต้นการสร้างฮิสโตนใหม่ที่อยู่รอบๆ ในรูปแบบใหม่ โดยการเปลี่ยนแปลงรูปร่างของฮิสโตนรอบๆ ฮิสโตนที่ถูกดัดแปลงเหล่านี้จะช่วยให้มั่นใจได้ว่าโปรแกรมการถอดรหัสเฉพาะสายพันธุ์จะยังคงอยู่หลังจากการแบ่งเซลล์
- วิธีที่สองคือการเพิ่มหมู่เมทิลลงใน DNA ส่วนใหญ่ที่ตำแหน่ง CpGเพื่อเปลี่ยนไซโตซีนเป็น5-เมทิลไซโตซีน 5-เมทิลไซโตซีนทำหน้าที่คล้ายกับไซโตซีนทั่วไป โดยจับคู่กับกัวนีนใน DNA สองสาย อย่างไรก็ตาม เมื่อมีไซโตซีนที่มีหมู่เมทิลอยู่ในตำแหน่ง CpG ในบริเวณโปรโมเตอร์และเอนแฮนเซอร์ของยีน ยีนมักจะถูกยับยั้ง[ 26 ] [ 27 ]เมื่อมีไซโตซีนที่มีหมู่เมทิลอยู่ในตำแหน่ง CpG ในส่วนของยีน (ในบริเวณการเข้ารหัสยกเว้นตำแหน่งเริ่มต้นการถอดรหัส) การแสดงออกของยีนมักจะเพิ่มขึ้น การถอดรหัสของยีนมักขึ้นอยู่กับปัจจัยการถอดรหัส ที่จับกับ ลำดับการจดจำ (10 เบสหรือน้อยกว่า) ที่เอนแฮนเซอร์ซึ่งมีปฏิสัมพันธ์กับบริเวณโปรโมเตอร์ของยีนนั้น ( การแสดงออกของยีน#เอนแฮนเซอร์ ปัจจัยการถอดรหัส คอมเพล็กซ์ตัวกลาง และลูป DNA ในการถอดรหัสของสัตว์เลี้ยงลูกด้วยนม ) [ 28 ]ประมาณ 22% ของปัจจัยการถอดรหัสถูกยับยั้งไม่ให้จับเมื่อลำดับการจดจำมีไซโตซีนที่ถูกเมทิลเลต นอกจากนี้ การมีไซโตซีนที่ถูกเมทิลเลตในบริเวณโปรโมเตอร์สามารถดึงดูด โปรตีนโดเมน ที่จับกับเมทิล-CpG (MBD) ได้ MBD ทั้งหมดจะโต้ตอบกับ การปรับโครงสร้าง นิวคลีโอโซมและ คอมเพล็กซ์ ฮิสโตนดีอะเซทิเลสซึ่งนำไปสู่การปิดการทำงานของยีน นอกจากนี้ การดัดแปลงโควาเลนต์อีกอย่างหนึ่งที่เกี่ยวข้องกับไซโตซีนที่ถูกเมทิลเลตคือการดีเมทิลเลชันโดยเอนไซม์ TETการดีเมทิลเลชันดังกล่าวเกิดขึ้นหลายร้อยครั้ง ตัวอย่างเช่น ในระหว่าง เหตุการณ์ การเรียนรู้และการสร้างความทรงจำในเซลล์ประสาท[ 29 ] [ 30 ]
มักมีความสัมพันธ์แบบต่างตอบแทนระหว่างการเมทิลเลชันของ DNA และการเมทิลเลชันของไลซีนในฮิสโตน[ 31 ]ตัวอย่างเช่นโปรตีนโดเมนจับเมทิล MBD1ซึ่งถูกดึงดูดและเชื่อมโยงกับไซโตซีนที่ถูกเมทิลเลชันในตำแหน่ง CpG ของ DNA ยังสามารถเชื่อมโยงกับ กิจกรรม เมทิลทรานสเฟอเรส H3K9 เพื่อเมทิลเลชันฮิสโตน 3 ที่ไลซีน 9 ในทางกลับกัน การเมทิลเลชันเพื่อการบำรุงรักษา DNA โดยDNMT1ดูเหมือนจะอาศัยการรับรู้การเมทิลเลชันของฮิสโตนบนนิวคลีโอโซมที่มีอยู่ในตำแหน่ง DNA เพื่อดำเนินการเมทิลเลชันของไซโตซีนบน DNA ที่สังเคราะห์ขึ้นใหม่[ 31 ]นอกจากนี้ยังมีการสื่อสารข้ามกันระหว่างการเมทิลเลชันของ DNA ที่ดำเนินการโดยDNMT3AและDNMT3Bและการเมทิลเลชันของฮิสโตน ดังนั้นจึงมีความสัมพันธ์ระหว่างการกระจายตัวของการเมทิลเลชันของ DNA และการเมทิลเลชันของฮิสโตนทั่วทั้งจีโนม[ 32 ]
กลไกการถ่ายทอดทางพันธุกรรมของสถานะฮิสโตนยังไม่เป็นที่เข้าใจดีนัก อย่างไรก็ตาม เป็นที่ทราบกันดีเกี่ยวกับกลไกการถ่ายทอดทางพันธุกรรมของสถานะเมทิลเลชั่นของ DNA ในระหว่างการแบ่งเซลล์และการเปลี่ยนแปลงรูปร่าง การถ่ายทอดทางพันธุกรรมของสถานะเมทิลเลชั่นขึ้นอยู่กับเอนไซม์บางชนิด (เช่นDNMT1 ) ที่มีความสัมพันธ์กับ 5-เมทิลไซโตซีนมากกว่าไซโตซีน หากเอนไซม์นี้ไปถึงส่วนของ DNA ที่ "มีเมทิลเลชั่นครึ่งหนึ่ง" (ซึ่ง 5-เมทิลไซโตซีนอยู่ในสาย DNA เพียงสายเดียวจากสองสาย) เอนไซม์จะทำการเมทิลเลชั่นอีกครึ่งหนึ่ง อย่างไรก็ตาม ปัจจุบันเป็นที่ทราบกันแล้วว่า DNMT1 มีปฏิสัมพันธ์ทางกายภาพกับโปรตีนUHRF1 UHRF1 เพิ่งได้รับการยอมรับว่าเป็นสิ่งจำเป็นสำหรับการบำรุงรักษาเมทิลเลชั่นของ DNA ที่เกิดจาก DNMT1 UHRF1 เป็นโปรตีนที่จดจำ DNA ที่มีเมทิลเลชั่นครึ่งหนึ่งโดยเฉพาะ ดังนั้นจึงนำ DNMT1 ไปยังสารตั้งต้นเพื่อรักษาเมทิลเลชั่นของ DNA [ 32 ]
แม้ว่าการดัดแปลงฮิสโตนจะเกิดขึ้นตลอดทั้งลำดับ แต่ปลาย N-terminus ที่ไม่มีโครงสร้างของฮิสโตน (เรียกว่าหางฮิสโตน) จะได้รับการดัดแปลงอย่างมากเป็นพิเศษ การดัดแปลงเหล่านี้ได้แก่การอะเซทิเล ชัน การเมทิลเลชันการยูบิควิทิเลชัน การฟอสโฟ รี เลชัน การซูโมอิเลชัน การไรโบซิเลชัน และการซิทรูลลิเนชัน การอะเซทิเลชันเป็นการดัดแปลงที่ได้รับการศึกษามากที่สุด ตัวอย่างเช่น การอะเซทิเลชันของไลซีน K14 และ K9 ของหางฮิสโตน H3 โดยเอนไซม์ฮิสโตนอะเซทิลทรานสเฟอเรส (HATs) โดยทั่วไปเกี่ยวข้องกับความสามารถในการถอดรหัส[ 34 ] (ดูรูป)
แนวคิดหนึ่งคือ แนวโน้มของการอะเซทิเลชันที่เกี่ยวข้องกับการถอดรหัส "ที่ทำงานอยู่" นั้นมีลักษณะทางชีวฟิสิกส์ เนื่องจากโดยปกติแล้วไลซีนจะมีไนโตรเจนที่มีประจุบวกอยู่ที่ปลาย จึงสามารถจับกับฟอสเฟตที่มีประจุลบของโครงสร้างหลักของ DNA ได้ เหตุการณ์อะเซทิเลชันจะเปลี่ยนกลุ่มอะมีนที่มีประจุบวกบนโซ่ข้างให้กลายเป็นพันธะอะไมด์ที่เป็นกลาง ซึ่งจะกำจัดประจุบวกออกไป ทำให้ DNA หลุดออกจากฮิสโตน เมื่อเกิดเหตุการณ์นี้ขึ้น คอมเพล็กซ์เช่นSWI/SNFและปัจจัยการถอดรหัสอื่นๆ สามารถจับกับ DNA และทำให้เกิดการถอดรหัสได้ นี่คือแบบจำลอง "ซิส" ของการทำงานทางเอพิเจเนติกส์ กล่าวอีกนัยหนึ่ง การเปลี่ยนแปลงที่หางของฮิสโตนมีผลโดยตรงต่อ DNA เอง[ 35 ]
อีกแบบจำลองหนึ่งของหน้าที่ทางเอพิเจเนติกส์คือแบบจำลอง "ทรานส์" ในแบบจำลองนี้ การเปลี่ยนแปลงที่หางของฮิสโตนจะส่งผลต่อดีเอ็นเอทางอ้อม ตัวอย่างเช่น การอะเซทิเลชันของไลซีนอาจสร้างตำแหน่งการจับสำหรับเอนไซม์ที่ปรับเปลี่ยนโครมาติน (หรือกลไกการถอดรหัสเช่นกัน) ตัวปรับเปลี่ยนโครมาตินนี้สามารถทำให้เกิดการเปลี่ยนแปลงสถานะของโครมาตินได้ อันที่จริง โบรโมโดเมน ซึ่งเป็นโดเมนโปรตีนที่จับกับอะเซทิลไลซีนโดยเฉพาะ พบได้ในเอนไซม์หลายชนิดที่ช่วยกระตุ้นการถอดรหัส รวมถึงคอมเพล็กซ์ SWI/SNF เป็นไปได้ว่าการอะเซทิเลชันอาจทำหน้าที่ในลักษณะนี้และในลักษณะก่อนหน้านี้เพื่อช่วยในการกระตุ้นการถอดรหัส
แนวคิดที่ว่าการดัดแปลงทำหน้าที่เป็นโมดูลเชื่อมต่อสำหรับปัจจัยที่เกี่ยวข้องนั้นได้รับการยืนยันโดยการเมทิลเลชันของฮิสโตนเช่นกัน การเมทิลเลชันของไลซีน 9 ในฮิสโตน H3 เกี่ยวข้องกับโครมาตินที่เงียบงันในการถอดรหัสอย่างถาวร ( เฮเทอโรโครมาติน แบบถาวร ) มานานแล้ว (ดูรูปด้านล่าง) มีการค้นพบว่าโครโมโดเมน (โดเมนที่จับกับเมทิลไลซีนโดยเฉพาะ) ในโปรตีนHP1 ที่ยับยั้งการถอดรหัส จะดึงดูด HP1 ไปยังบริเวณที่มีการเมทิลเลชันของ K9 ตัวอย่างหนึ่งที่ดูเหมือนจะหักล้างแบบจำลองทางชีวฟิสิกส์นี้สำหรับการเมทิลเลชันคือ การไตรเมทิลเลชันของฮิสโตน H3 ที่ไลซีน 4 มีความสัมพันธ์อย่างมากกับ (และจำเป็นสำหรับ) การกระตุ้นการถอดรหัสอย่างสมบูรณ์ (ดูรูปด้านบน) ในกรณีนี้ การไตรเมทิลเลชันจะทำให้เกิดประจุบวกคงที่ที่ส่วนหาง
มีการแสดงให้เห็นว่าฮิสโตนไลซีนเมทิลทรานสเฟอเรส (KMT) มีหน้าที่รับผิดชอบต่อกิจกรรมเมทิลเลชันนี้ในรูปแบบของฮิสโตน H3 และ H4 เอนไซม์นี้ใช้ไซต์ที่ออกฤทธิ์เร่งปฏิกิริยาที่เรียกว่าโดเมน SET (Suppressor of variegation, Enhancer of Zeste, Trithorax) โดเมน SET เป็นลำดับกรดอะมิโน 130 ตัวที่เกี่ยวข้องกับการปรับเปลี่ยนกิจกรรมของยีน โดเมนนี้ได้รับการพิสูจน์แล้วว่าสามารถจับกับหางของฮิสโตนและทำให้เกิดการเมทิลเลชันของฮิสโตน[ 36 ]
การดัดแปลงฮิสโตนที่แตกต่างกันมีแนวโน้มที่จะทำงานในลักษณะที่แตกต่างกัน การอะเซทิเลชันที่ตำแหน่งหนึ่งมีแนวโน้มที่จะทำงานแตกต่างจากการอะเซทิเลชันที่ตำแหน่งอื่น นอกจากนี้ การดัดแปลงหลายอย่างอาจเกิดขึ้นพร้อมกัน และการดัดแปลงเหล่านี้อาจทำงานร่วมกันเพื่อเปลี่ยนพฤติกรรมของนิวคลีโอโซมแนวคิดที่ว่าการดัดแปลงแบบไดนามิกหลายอย่างควบคุมการถอดรหัสยีนในลักษณะที่เป็นระบบและทำซ้ำได้เรียกว่ารหัสฮิสโตนแม้ว่าแนวคิดที่ว่าสถานะของฮิสโตนสามารถอ่านได้แบบเชิงเส้นในฐานะตัวนำข้อมูลดิจิทัลนั้นถูกหักล้างไปแล้วเป็นส่วนใหญ่ หนึ่งในระบบที่เข้าใจได้ดีที่สุดที่ประสานการปิดกั้นการทำงานของโครมาตินคือการปิดกั้นการทำงานของยีน HML และ HMR ในยีสต์โดยใช้ โปรตีน SIR
การเมทิลเลชั่นของดีเอ็นเอ
การเมทิลเลชันของ DNAมักเกิดขึ้นในลำดับที่ซ้ำกัน และช่วยยับยั้งการแสดงออกและการเคลื่อนที่ของ ' องค์ประกอบที่เคลื่อนย้ายได้ ': [ 37 ]เนื่องจาก5-เมทิลไซโตซีนสามารถดีอะมิเนตเป็นไทมิดีน ได้เอง (แทนที่ไนโตรเจนด้วยออกซิเจน) ตำแหน่ง CpG จึงมักกลายพันธุ์และหายากในจีโนม ยกเว้นที่เกาะ CpGซึ่งโดยทั่วไปจะยังคงไม่ถูกเมทิลเลชัน การเปลี่ยนแปลงทางเอพิเจเนติกส์ประเภทนี้จึงมีศักยภาพที่จะนำไปสู่ความถี่ที่เพิ่มขึ้นของการกลายพันธุ์ทางพันธุกรรมถาวร รูปแบบการเมทิลเลชันของ DNA เป็นที่ทราบกันดีว่าถูกสร้างขึ้นและแก้ไขเพื่อตอบสนองต่อปัจจัยด้านสิ่งแวดล้อมโดยการทำงานร่วมกันที่ซับซ้อนของเอนไซม์เมทิลทรานสเฟอเรสของ DNA อย่างน้อยสามชนิดที่เป็นอิสระต่อกัน ได้แก่ DNMT1, DNMT3A และ DNMT3B การสูญเสียเอนไซม์ใดเอนไซม์หนึ่งในสามชนิดนี้จะทำให้หนูตายได้[ 38 ] DNMT1 เป็นเมทิลทรานสเฟอเรสที่พบมากที่สุดในเซลล์ร่างกาย[ 39 ]อยู่ในบริเวณจุดจำลองแบบ[ 40 ]มีความชอบต่อ DNA ที่มีเมทิลเพียงครึ่งเดียว 10–40 เท่า และมีปฏิสัมพันธ์กับแอนติเจนนิวเคลียร์ของเซลล์ที่กำลังแบ่งตัว (PCNA) [ 41 ]
DNMT1 ถ่ายโอนรูปแบบการเติมหมู่เมทิลไปยังสายดีเอ็นเอที่สังเคราะห์ขึ้นใหม่หลังจากการจำลองดีเอ็นเอ โดยการปรับเปลี่ยนดีเอ็นเอที่มีการเติมหมู่เมทิล เพียงครึ่งเดียว ดังนั้นจึงมักถูกเรียกว่าเมทิลทรานสเฟอเรสแบบ 'บำรุงรักษา' [ 42 ] DNMT1 มีความสำคัญต่อการพัฒนาตัวอ่อน การประทับตรา และการปิดใช้งานโครโมโซม X อย่างเหมาะสม[ 38 ] [ 43 ]เพื่อเน้นความแตกต่างของกลไกการถ่ายทอดทางพันธุกรรมระดับโมเลกุลนี้จากกลไกการจับคู่เบสแบบวัตสัน-คริกแบบดั้งเดิม จึงมีการนำคำว่า 'การสร้างแม่แบบเอพิเจเนติกส์' มาใช้[ 44 ]นอกจากนี้ นอกเหนือจากการบำรุงรักษาและการส่งผ่านสถานะดีเอ็นเอที่มีการเติมหมู่เมทิลแล้ว หลักการเดียวกันนี้ยังสามารถใช้ในการบำรุงรักษาและการส่งผ่านการดัดแปลงฮิสโตนและแม้แต่สถานะทางพันธุกรรมของไซโตพลาสซึม ( โครงสร้าง ) ได้อีกด้วย [ 45 ]
การเมทิลเลชันของอาร์เอ็นเอ
การเติมหมู่เมทิล N6-methyladenosine (m6A) ลงใน RNA ซึ่งเป็นการดัดแปลง RNA ที่พบมากที่สุดในยูคาริโอต ได้รับการยอมรับเมื่อเร็ว ๆ นี้ว่าเป็นกลไกการควบคุมยีนที่สำคัญ[ 46 ]
ในปี 2011 ได้มีการพิสูจน์แล้วว่าการเมทิลเลชันของmRNAมีบทบาทสำคัญในการรักษาสมดุลพลังงาน ของมนุษย์ ยีน FTOที่เกี่ยวข้องกับโรคอ้วนแสดงให้เห็นว่าสามารถดีเมทิลเลชันN6-เมทิลอะดีโนซีนใน RNA ได้ [ 47 ] [ 48 ]
การดัดแปลงฮิสโตน
ฮิสโตน H3 และ H4 ยังสามารถถูกจัดการได้ผ่านการกำจัดหมู่เมทิลโดยใช้ฮิสโตนไลซีนดีเมทิเลส (KDM) เอนไซม์ที่เพิ่งค้นพบนี้มีไซต์ที่ออกฤทธิ์ทางเร่งปฏิกิริยาที่เรียกว่าโดเมน Jumonji (JmjC) การกำจัดหมู่เมทิลเกิดขึ้นเมื่อ JmjC ใช้โคแฟคเตอร์หลายตัวเพื่อไฮดรอกซิเลตหมู่เมทิล ทำให้หมู่เมทิลถูกกำจัดออกไป JmjC สามารถกำจัดหมู่เมทิลออกจากสารตั้งต้นที่มีหมู่เมทิลหนึ่ง สอง และสามหมู่ได้[ 49 ]
บริเวณโครโมโซมสามารถปรับใช้สถานะทางเลือกที่เสถียรและถ่ายทอดได้ ส่งผลให้เกิดการแสดงออกของยีนแบบสองสถานะโดยไม่ต้องเปลี่ยนแปลงลำดับดีเอ็นเอ การควบคุมเอพิเจเนติกมักเกี่ยวข้องกับการดัดแปลงโควาเลนต์ ทางเลือก ของฮิสโตน[ 50 ]ความเสถียรและการถ่ายทอดของสถานะของบริเวณโครโมโซมขนาดใหญ่คาดว่าจะเกี่ยวข้องกับปฏิกิริยาป้อนกลับเชิงบวก โดยที่นิวคลีโอโซม ที่ถูกดัดแปลง จะดึงดูดเอนไซม์ที่ดัดแปลงนิวคลีโอโซมที่อยู่ใกล้เคียงในลักษณะเดียวกัน[ 51 ]แบบจำลองสุ่มแบบง่ายสำหรับเอพิเจเนติกประเภทนี้พบได้ที่นี่[ 52 ] [ 53 ]
มีการเสนอแนะว่าการควบคุมการถอดรหัสตามโครมาตินอาจได้รับอิทธิพลจากผลของ RNA ขนาดเล็กRNA ที่รบกวนขนาดเล็กสามารถปรับเปลี่ยนการแสดงออกของยีนผ่านการดัดแปลงเอพิเจเนติกของโปรโมเตอร์ เป้าหมาย ได้[ 54 ]
ทรานสคริปต์ RNA
บางครั้ง ยีนเมื่อถูกกระตุ้นแล้วจะถอดรหัสผลิตภัณฑ์ที่ช่วยรักษาการทำงานของยีนนั้นโดยตรงหรือโดยอ้อม ตัวอย่างเช่นHnf4และMyoDช่วยเพิ่มการถอดรหัสของยีนเฉพาะตับและยีนเฉพาะกล้ามเนื้อตามลำดับ รวมถึงยีนของพวกมันเอง ผ่าน กิจกรรมของ ปัจจัย การถอดรหัส ของโปรตีนที่พวกมันเข้ารหัส การส่งสัญญาณ RNA รวมถึงการดึงดูดที่แตกต่างกันของลำดับชั้นของคอมเพล็กซ์ปรับเปลี่ยนโครมาตินทั่วไปและ DNA เมทิลทรานสเฟอเรสไปยังตำแหน่งเฉพาะโดย RNA ในระหว่างการแยกแยะและการพัฒนา[ 55 ]การเปลี่ยนแปลงทางเอพิเจเนติกส์อื่นๆ เกิดขึ้นจากการสร้างรูปแบบการต่อเชื่อมที่แตกต่างกันของRNAหรือโดยการสร้าง RNA สองสาย ( RNAi ) ลูกหลานของเซลล์ที่ยีนถูกเปิดใช้งานจะสืบทอดกิจกรรมนี้ แม้ว่าสิ่งกระตุ้นดั้งเดิมสำหรับการเปิดใช้งานยีนจะไม่มีอยู่อีกต่อไปแล้ว ยีนเหล่านี้มักจะถูกเปิดหรือปิดโดยการส่งสัญญาณแม้ว่าในบางระบบที่ซิงไซเทียหรือช่องว่างเชื่อมต่อมีความสำคัญ RNA อาจแพร่กระจายไปยังเซลล์หรือนิวเคลียสอื่นๆ โดยตรงโดยการแพร่แม่จะส่ง RNA และโปรตีนจำนวนมากไปยังไซโกต ในระหว่าง การสร้างไข่หรือผ่านเซลล์พยาบาลส่งผลให้เกิด ฟีโนไทป์ที่ มีผลจากแม่มีการถ่ายทอด RNA จากอสุจิจากพ่อในปริมาณที่น้อยกว่า แต่มีหลักฐานล่าสุดว่าข้อมูลเอพิเจเนติกนี้สามารถนำไปสู่การเปลี่ยนแปลงที่มองเห็นได้ในลูกหลานหลายรุ่น[ 56 ]
ไมโครอาร์เอ็นเอ
ไมโครอาร์เอ็นเอ (miRNAs) เป็นสมาชิกของอาร์เอ็นเอที่ไม่เข้ารหัสซึ่งมีขนาดตั้งแต่ 17 ถึง 25 นิวคลีโอไทด์ miRNAs ควบคุมการทำงานทางชีวภาพที่หลากหลายในพืชและสัตว์[ 57 ]จนถึงปัจจุบันในปี 2013 มีการค้นพบ miRNAs ประมาณ 2,000 ชนิดในมนุษย์ และสามารถค้นหาได้ทางออนไลน์ในฐานข้อมูล miRNA [ 58 ] miRNA แต่ละตัวที่แสดงออกในเซลล์อาจกำหนดเป้าหมาย mRNA ประมาณ 100 ถึง 200 ตัวที่มันลดระดับลง[ 59 ]การลดระดับ mRNA ส่วนใหญ่เกิดขึ้นจากการทำให้ mRNA เป้าหมายสลายตัว ในขณะที่การลดระดับบางส่วนเกิดขึ้นในระดับการแปลเป็นโปรตีน[ 60 ]
ดูเหมือนว่าประมาณ 60% ของยีนที่เข้ารหัสโปรตีนของมนุษย์จะถูกควบคุมโดย miRNA [ 61 ] miRNA จำนวนมากถูกควบคุมโดยเอพิเจเนติกส์ ประมาณ 50% ของยีน miRNA เกี่ยวข้องกับเกาะ CpG [ 57 ]ซึ่งอาจถูกยับยั้งโดยการเมทิลเลชันทางเอพิเจเนติกส์ การถอดรหัสจากเกาะ CpG ที่ถูกเมทิลเลชันจะถูกยับยั้งอย่างรุนแรงและถ่ายทอดทางพันธุกรรมได้[ 62 ] miRNA อื่นๆ ถูกควบคุมโดยเอพิเจเนติกส์โดยการดัดแปลงฮิสโตนหรือโดยการเมทิลเลชันของ DNA และการดัดแปลงฮิสโตนร่วมกัน[ 57 ]
sRNA
sRNAเป็นชิ้นส่วน RNA ขนาดเล็ก (50–250 นิวคลีโอไทด์) ที่มีโครงสร้างสูงและไม่มีรหัส ซึ่งพบในแบคทีเรีย พวกมันควบคุมการแสดงออกของยีน รวมถึง ยีน ก่อโรคในเชื้อก่อโรค และถูกมองว่าเป็นเป้าหมายใหม่ในการต่อสู้กับแบคทีเรียดื้อยา[ 63 ]พวกมันมีบทบาทสำคัญในกระบวนการทางชีววิทยาหลายอย่าง โดยจับกับ mRNA และโปรตีนเป้าหมายในโปรคาริโอต การวิเคราะห์ทางวิวัฒนาการของพวกมัน เช่น ผ่านปฏิสัมพันธ์ระหว่าง sRNA กับเป้าหมาย mRNA หรือคุณสมบัติการจับกับ โปรตีน ถูกนำมาใช้เพื่อสร้างฐานข้อมูลที่ครอบคลุม[ 64 ]แผนที่ sRNA- ยีน ที่สร้างขึ้นจากเป้าหมายในจีโนมของจุลินทรีย์ก็ถูกสร้างขึ้นเช่นกัน[ 65 ]
อาร์เอ็นเอที่ไม่เข้ารหัสแบบยาว
การตรวจสอบจำนวนมากได้แสดงให้เห็นถึงการมีส่วนร่วมที่สำคัญของ RNA ที่ไม่เข้ารหัสแบบยาว (lncRNAs) ในการควบคุมการแสดงออกของยีนและการดัดแปลงโครโมโซม ซึ่งส่งผลให้มีการควบคุมอย่างมีนัยสำคัญต่อการแบ่งเซลล์ RNA ที่ไม่เข้ารหัสแบบยาวเหล่านี้ยังมีส่วนช่วยในการประทับตราทางพันธุกรรมและการปิดใช้งานโครโมโซม X [ 66 ]ในสัตว์ไม่มีกระดูกสันหลัง เช่น แมลงสังคมอย่างผึ้ง RNA ที่ไม่เข้ารหัสแบบยาวถูกตรวจพบว่าเป็นกลไกทางพันธุกรรมที่เป็นไปได้ผ่านยีนเฉพาะอัลลีลที่อยู่เบื้องหลังความก้าวร้าวผ่านการผสมข้ามแบบต่างตอบแทน[ 67 ]
พรีออน
พรีออนเป็นโปรตีนรูปแบบติดเชื้อโดยทั่วไป โปรตีนจะพับตัวเป็นหน่วยที่แยกจากกันซึ่งทำหน้าที่เฉพาะในเซลล์ แต่โปรตีนบางชนิดก็สามารถสร้างสถานะโครงสร้างที่ติดเชื้อได้ ซึ่งเรียกว่าพรีออน แม้ว่ามักจะถูกมองในบริบทของโรคติดเชื้อ แต่ พรีออนก็ถูกนิยามอย่างหลวมๆ ด้วยความสามารถในการเปลี่ยนโปรตีนในสถานะดั้งเดิมอื่นๆ ให้เป็นสถานะโครงสร้างที่ติดเชื้อได้ ในความหมายหลังนี้ พวกมันสามารถถูกมองว่าเป็นเอพิเจเนติกส์ที่สามารถชักนำให้เกิดการเปลี่ยนแปลงฟีโนไทป์โดยไม่ต้องดัดแปลงจีโนม[ 68 ]
บางคนถือว่า พรีออนของเชื้อราเป็นเอพิเจเนติก เนื่องจากฟีโนไทป์การติดเชื้อที่เกิดจากพรีออนสามารถถ่ายทอดทางพันธุกรรมได้โดยไม่ต้องมีการปรับเปลี่ยนจีโนมPSI+และ URE3 ซึ่งค้นพบในยีสต์ในปี 1965 และ 1971 เป็นพรีออนสองชนิดที่ได้รับการศึกษามากที่สุด[ 69 ] [ 70 ]พรีออนสามารถมีผลต่อฟีโนไทป์ได้โดยการกักเก็บโปรตีนไว้ในกลุ่มก้อน ทำให้กิจกรรมของโปรตีนนั้นลดลง ในเซลล์ PSI+ การสูญเสียโปรตีน Sup35 (ซึ่งเกี่ยวข้องกับการยุติการแปล) ทำให้ไรโบโซมมีอัตราการอ่านผ่านโคดอน หยุดสูงขึ้น ซึ่งเป็นผลที่ทำให้เกิดการยับยั้งการกลายพันธุ์แบบไร้ความหมายในยีนอื่นๆ[ 71 ]ความสามารถของ Sup35 ในการสร้างพรีออนอาจเป็นลักษณะที่ได้รับการอนุรักษ์ไว้ อาจให้ข้อได้เปรียบในการปรับตัวโดยการให้เซลล์สามารถเปลี่ยนเป็นสถานะ PSI+และแสดงลักษณะทางพันธุกรรมที่สงบนิ่งซึ่งปกติจะสิ้นสุดลงด้วยการกลายพันธุ์ของรหัสหยุด[ 72 ] [ 73 ] [ 74 ] [ 75 ]
เอพิเจเนติ กส์ที่อิงตามพรีออนยังพบได้ในSaccharomyces cerevisiae อีกด้วย [ 76 ]
พื้นฐานระดับโมเลกุล
การเปลี่ยนแปลงทางเอพิเจเนติกส์จะปรับเปลี่ยนการทำงานของยีนบางตัว แต่ไม่ใช่ลำดับรหัสพันธุกรรมของ DNA [ 77 ]โครงสร้างจุลภาค (ไม่ใช่รหัส) ของ DNA เองหรือ โปรตีน โครมาติน ที่เกี่ยวข้อง อาจถูกปรับเปลี่ยน ทำให้เกิดการทำงานหรือการยับยั้ง กลไกนี้ช่วยให้เซลล์ที่แตกต่างกันในสิ่งมีชีวิตหลายเซลล์สามารถแสดงออกเฉพาะยีนที่จำเป็นสำหรับกิจกรรมของตนเองเท่านั้น การเปลี่ยนแปลงทางเอพิเจเนติกส์จะคงอยู่เมื่อเซลล์แบ่งตัว การเปลี่ยนแปลงทางเอพิเจเนติกส์ส่วนใหญ่เกิดขึ้นภายในช่วงชีวิตของสิ่งมีชีวิตแต่ละตัวเท่านั้น อย่างไรก็ตาม การเปลี่ยนแปลงทางเอพิเจเนติกส์เหล่านี้สามารถส่งต่อไปยังลูกหลานของสิ่งมีชีวิตนั้นได้ผ่านกระบวนการที่เรียกว่าการถ่ายทอดทางเอพิเจเนติกส์ข้ามรุ่นยิ่งไปกว่านั้น หากการปิดใช้งานยีนเกิดขึ้นในเซลล์อสุจิหรือเซลล์ไข่ที่ส่งผลให้เกิดการปฏิสนธิ การปรับเปลี่ยนทางเอพิเจเนติกส์นี้อาจถูกถ่ายทอดไปยังรุ่นต่อไปได้เช่นกัน[ 78 ]
กระบวนการเอพิเจเนติกส์เฉพาะ ได้แก่พารามิวเทชันการบุ๊กมาร์ก การประทับตราการปิดกั้นยีนการปิดใช้งานโครโมโซม X ผลกระทบจากตำแหน่งการ ตั้งโปรแกรม ใหม่ ของการเมทิลเล ชันDNA ทรานส์เวคชันผลกระทบจากมารดาความก้าวหน้าของการเกิดมะเร็งผลกระทบหลายประการของสารก่อความพิการแต่กำเนิด การควบคุมการดัดแปลงฮิสโตนและเฮเทอโรโครมาตินและข้อจำกัดทางเทคนิคที่ส่งผลต่อพาร์ทีโนเจเนซิสและการโคลนนิ่ง[ 79 ] [ 80 ] [ 81 ]
ความเสียหายของดีเอ็นเอ
ความเสียหายของ DNA ยังสามารถทำให้เกิดการเปลี่ยนแปลงทางเอพิเจเนติกส์ได้[ 82 ] [ 83 ] [ 84 ]ความเสียหายของ DNA เกิดขึ้นบ่อยมาก โดยเฉลี่ยประมาณ 60,000 ครั้งต่อวันต่อเซลล์ในร่างกายมนุษย์ (ดูความเสียหายของ DNA (ที่เกิดขึ้นตามธรรมชาติ) ) ความเสียหายเหล่านี้ส่วนใหญ่ได้รับการซ่อมแซม อย่างไรก็ตาม การเปลี่ยนแปลงทางเอพิเจเนติกส์ยังคงสามารถคงอยู่ได้ในบริเวณที่มีการซ่อมแซม DNA โดยเฉพาะอย่างยิ่ง การแตกของสายคู่ใน DNA สามารถเริ่มต้นการปิดการทำงานของยีนทางเอพิเจเนติกส์ที่ไม่ได้กำหนดไว้ล่วงหน้าได้ ทั้งโดยการทำให้เกิดเมทิลเลชั่นของ DNA และโดยการส่งเสริมการดัดแปลงฮิสโตนประเภทการปิดการทำงาน (การปรับโครงสร้างโครมาติน - ดูส่วนถัดไป) [ 85 ]นอกจากนี้ เอนไซม์Parp1 (poly(ADP)-ribose polymerase)และผลิตภัณฑ์ของมันคือ poly(ADP)-ribose (PAR) จะสะสมอยู่ที่บริเวณที่มีความเสียหายของ DNA ซึ่งเป็นส่วนหนึ่งของกระบวนการซ่อมแซม[ 86 ]การสะสมนี้จะนำไปสู่การสรรหาและการกระตุ้นโปรตีนปรับโครงสร้างโครมาติน ALC1 ซึ่งสามารถทำให้เกิดการปรับโครงสร้างนิวคลีโอโซม ได้ [ 87 ]พบว่าการปรับโครงสร้างนิวคลีโอโซมทำให้เกิดการปิดการทำงานของยีนซ่อมแซม DNA MLH1 ในระดับเอพิเจเนติกส์[ 21 ] [ 88 ]สารเคมีที่ทำลาย DNA เช่นเบนซีนไฮโดรควินอน สไตรีนคาร์บอนเตตระคลอไรด์และไตรคลอโรเอทิลีนทำให้เกิดการลดเมทิลเลชั่นของ DNA อย่างมาก บางส่วนเกิดจากการกระตุ้นวิถีทางความเครียดออกซิเดชั่น[ 89 ]
เป็นที่ทราบกันดีว่าอาหารสามารถเปลี่ยนแปลงเอพิเจเนติกส์ของหนูที่กินอาหารต่างกันได้[ 90 ]ส่วนประกอบอาหารบางชนิดเพิ่มระดับเอนไซม์ซ่อมแซม DNA เช่นMGMTและMLH1 [ 91 ]และp53 [ 92 ]ส่วนประกอบอาหารอื่นๆ สามารถลดความเสียหายของ DNA ได้ เช่น ไอโซฟลาโวนจากถั่วเหลือง ในการศึกษาหนึ่งพบว่า ตัวบ่งชี้ความเครียดออกซิเดชัน เช่น นิวคลีโอไทด์ที่เปลี่ยนแปลงไปซึ่งอาจเกิดจากความเสียหายของ DNA ลดลงเมื่อรับประทานอาหารเสริมถั่วเหลืองเป็นเวลา 3 สัปดาห์[ 93 ] นอกจากนี้ยังพบว่าความเสียหายของ DNA จากออกซิเดชันลดลง 2 ชั่วโมงหลังจากรับประทานสารสกัดจากกากบลูเบอร์รี่ ( Vaccinium myrtillius L.) ที่อุดมไปด้วยแอนโทไซยานิน [ 94 ]
การซ่อมแซมดีเอ็นเอ
ความเสียหายต่อดีเอ็นเอเป็นเรื่องที่พบได้บ่อยและมีการซ่อมแซมอยู่ตลอดเวลา การเปลี่ยนแปลงทางเอพิเจเนติกส์อาจเกิดขึ้นพร้อมกับการซ่อมแซมดีเอ็นเอจากความเสียหายจากออกซิเดชันหรือการแตกหักของสายดีเอ็นเอสองสาย ในเซลล์มนุษย์ ความเสียหายต่อดีเอ็นเอจากออกซิเดชันเกิดขึ้นประมาณ 10,000 ครั้งต่อวัน และการแตกหักของสายดีเอ็นเอสองสายเกิดขึ้นประมาณ 10 ถึง 50 ครั้งต่อรอบการแบ่งเซลล์ในเซลล์ร่างกายที่กำลังแบ่งตัว (ดูความเสียหายต่อดีเอ็นเอ (ที่เกิดขึ้นตามธรรมชาติ) ) ข้อได้เปรียบเชิงคัดเลือกของการซ่อมแซมดีเอ็นเอคือการช่วยให้เซลล์อยู่รอดได้เมื่อเผชิญกับความเสียหายต่อดีเอ็นเอ ส่วนข้อได้เปรียบเชิงคัดเลือกของการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่เกิดขึ้นพร้อมกับการซ่อมแซมดีเอ็นเอนั้นยังไม่ชัดเจน
การซ่อมแซมความเสียหายของดีเอ็นเอจากปฏิกิริยาออกซิเดชันสามารถเปลี่ยนแปลงเครื่องหมายทางพันธุกรรมได้
ในสภาวะคงที่ (โดยมีการเกิดความเสียหายภายในและได้รับการซ่อมแซม) จะมีกัวนีนที่เสียหายจากการออกซิเดชันประมาณ 2,400 ตัวที่ก่อตัวเป็น8-oxo-2'-deoxyguanosine (8-OHdG) ในดีเอ็นเอของเซลล์สัตว์เลี้ยงลูกด้วยนมโดยเฉลี่ย[ 95 ] 8-OHdG คิดเป็นประมาณ 5% ของความเสียหายจากการออกซิเดชันที่พบได้ทั่วไปในดีเอ็นเอ[ 96 ]กัวนีนที่ถูกออกซิไดซ์ไม่ได้เกิดขึ้นแบบสุ่มในบรรดากัวนีนทั้งหมดในดีเอ็นเอ มีความชอบลำดับสำหรับกัวนีนที่ ไซต์ CpG ที่ถูกเมทิลเลต (ไซโตซีนตามด้วยกัวนีนตามทิศทาง 5' → 3'และไซโตซีนถูกเมทิลเลต (5-mCpG)) [ 97 ]ไซต์ 5-mCpG มีศักยภาพในการแตกตัวเป็นไอออนต่ำที่สุดสำหรับการออกซิเดชันของกัวนีน

กัวนีนที่ถูกออกซิไดซ์มีศักยภาพในการจับคู่ผิดพลาดและก่อให้เกิดการกลายพันธุ์[ 99 ] ออก โซกัวนีนไกลโคซิเลส (OGG1) เป็นเอนไซม์หลักที่รับผิดชอบในการตัดกัวนีนที่ถูกออกซิไดซ์ออกในระหว่างการซ่อมแซม DNA OGG1 ค้นหาและจับกับ 8-OHdG ภายในไม่กี่วินาที[ 100 ]อย่างไรก็ตาม OGG1 ไม่ได้ตัด 8-OHdG ออกทันที ในเซลล์ HeLa การกำจัด 8-OHdG ครึ่งหนึ่งของค่าสูงสุดเกิดขึ้นใน 30 นาที[ 101 ]และในหนูที่ได้รับรังสี 8-OHdG ที่เกิดขึ้นในตับของหนูจะถูกกำจัดออกไปโดยมีครึ่งชีวิต 11 นาที[ 96 ]
เมื่อ OGG1 อยู่ที่กัวนีนที่ถูกออกซิไดซ์ภายในตำแหน่ง CpG ที่ถูกเมทิลเลชัน มันจะดึงดูดTET1มายังรอยโรค 8-OHdG (ดูรูป) ซึ่งทำให้ TET1 สามารถกำจัดหมู่เมทิลออกจากไซโตซีนที่ถูกเมทิลเลชันที่อยู่ติดกันได้ การกำจัดหมู่เมทิลออกจากไซโตซีนเป็นการเปลี่ยนแปลงทางเอพิเจเนติกส์
ตัวอย่างเช่น เมื่อเซลล์เยื่อบุผิวเต้านมของมนุษย์ได้รับการรักษาด้วย H2O2 เป็นเวลาหกชั่วโมง 8-OHdG จะเพิ่มขึ้นประมาณ 3.5 เท่าใน DNA และทำให้เกิดการลดเมทิลเลชั่นของ 5-เมทิลไซโตซีนในจีโนมประมาณ 80% [ 98 ] การลดเมทิลเลชั่นของ CpG ในโปรโมเตอร์ของยีนโดย กิจกรรม ของเอนไซม์ TETจะเพิ่มการถอดรหัสของยีนเป็น mRNA [ 102 ]ในเซลล์ที่ได้รับการรักษาด้วยH2O2 ยีนเฉพาะหนึ่งตัวได้รับการตรวจสอบคือBACE1 [ 98 ] ระดับเมทิลเลชั่นของ CpG island ของBACE1 ลดลง (การเปลี่ยนแปลงทางเอ พิเจเนติก) และทำให้การแสดงออกของmRNA ของ BACE1 เพิ่มขึ้นประมาณ 6.5 เท่า
ในขณะที่การบ่มด้วย H2O2 เป็นเวลาหกชั่วโมงทำให้เกิดการลดเมทิลเลชั่นของไซต์ 5-mCpG อย่างมาก แต่การบ่มด้วย H2O2 ในระยะเวลาที่สั้นกว่าดูเหมือนจะส่งเสริมการเปลี่ยนแปลงทางเอพิเจเนติกส์อื่นๆ การรักษาเซลล์ด้วย H2O2 เป็น เวลา 30 นาทีทำให้โปรตีนซ่อมแซมความผิดพลาดแบบเฮเทอโรไดเม อร์ MSH2-MSH6 ดึงดูด DNA เมทิลทรานสเฟอเรส 1 ( DNMT1 )ไปยังไซต์ที่มีความเสียหายของ DNA จากออกซิเดชั่นบางชนิด[ 103 ]ซึ่งอาจทำให้เกิดการเพิ่มเมทิลเลชั่นของไซโตซีน (การเปลี่ยนแปลงทางเอพิเจเนติกส์) ในตำแหน่งเหล่านี้
Jiang et al. [ 104 ]ได้ทำการรักษาเซลล์ HEK 293ด้วยสารที่ก่อให้เกิดความเสียหายต่อ DNA จากออกซิเดชัน ( โพแทสเซียมโบรเมต (KBrO 3 ) หรือโพแทสเซียมโครเมต (K 2 CrO 4 )) การซ่อมแซมแบบตัดฐาน (BER) ของความเสียหายจากออกซิเดชันเกิดขึ้นโดยเอนไซม์ซ่อมแซม DNA โพลีเมอเรสเบตาที่ไปอยู่ที่กัวนีนที่ถูกออกซิไดซ์ โพลีเมอเรสเบตาเป็นโพลีเมอเรสหลักของมนุษย์ในการซ่อมแซม BER แบบสั้นๆ ของความเสียหายต่อ DNA จากออกซิเดชัน Jiang et al. [ 104 ]ยังพบว่าโพลีเมอเรสเบตาได้ดึงดูด โปรตีน DNA เมทิลทรานสเฟอเรส DNMT3b ไปยังตำแหน่งการซ่อมแซม BER จากนั้นพวกเขาได้ประเมินรูปแบบการเมทิลเลชันในระดับนิวคลีโอไทด์เดี่ยวในบริเวณเล็กๆ ของ DNA ซึ่งรวมถึง บริเวณ โปรโมเตอร์และบริเวณการถอดรหัสช่วงต้นของ ยีน BRCA1ความเสียหายต่อ DNA จากออกซิเดชันจากโบรเมตได้ปรับเปลี่ยนรูปแบบการเมทิลเลชันของ DNA (ทำให้เกิดการเปลี่ยนแปลงทางเอพิเจเนติกส์) ที่ไซต์ CpG ภายในบริเวณของ DNA ที่ศึกษา ในเซลล์ที่ไม่ได้รับการรักษา CpG ที่ตำแหน่ง −189, −134, −29, −19, +16 และ +19 ของยีน BRCA1 มีไซโตซีนที่ถูกเมทิลเลชัน (โดยการนับเลขเริ่มจาก ตำแหน่งเริ่มต้นการถอดรหัส mRNAและเลขลบแสดงถึงนิวคลีโอไทด์ในบริเวณโปรโมเตอร์ต้นน้ำ) การเกิดออกซิเดชันที่เกิดจากการรักษาด้วยโบรเมตส่งผลให้ไซโตซีนที่ถูกเมทิลเลชันที่ตำแหน่ง −189, −134, +16 และ +19 หายไป และยังนำไปสู่การสร้างเมทิลเลชันใหม่ที่ CpG ที่ตำแหน่ง −80, −55, −21 และ +8 หลังจากที่การซ่อมแซม DNA เกิดขึ้น
การซ่อมแซมโดยการรวมตัวของยีนที่เหมือนกันจะเปลี่ยนแปลงเครื่องหมายทางเอพิเจเนติกส์
บทความหลายฉบับรายงานการคัดเลือกDNA เมทิลทรานสเฟอเรส 1 (DNMT1)ไปยังตำแหน่งที่มีการแตกของสาย DNA สองสาย[ 105 ] [ 106 ] [ 107 ]ในระหว่างการซ่อมแซมแบบโฮโมโลจัสรีคอมบิเนชัน (HR)ของการแตกของสาย DNA สองสาย การมีส่วนร่วมของ DNMT1 ทำให้สาย DNA ที่ได้รับการซ่อมแซมทั้งสองสายมีระดับของไซโตซีนที่ถูกเมทิลเลชันแตกต่างกัน สายหนึ่งจะถูกเมทิลเลชันบ่อยครั้งที่ตำแหน่ง CpG ประมาณ 21 ตำแหน่งที่อยู่ถัดจากจุดแตกของสาย DNA สองสายที่ได้รับการซ่อมแซม สาย DNA อีกสายหนึ่งจะสูญเสียการเมทิลเลชันที่ตำแหน่ง CpG ประมาณ 6 ตำแหน่งที่เคยถูกเมทิลเลชันมาก่อนที่อยู่ถัดจากจุดแตกของสาย DNA สองสาย รวมถึงสูญเสียการเมทิลเลชันที่ตำแหน่ง CpG ประมาณ 5 ตำแหน่งที่เคยถูกเมทิลเลชันมาก่อนที่อยู่ก่อนหน้าจุดแตกของสาย DNA สองสาย เมื่อโครโมโซมถูกจำลองแบบ จะทำให้เกิดโครโมโซมลูกสาวหนึ่งตัวที่ถูกเมทิลเลชันอย่างมากที่อยู่ถัดจากจุดแตกเดิม และอีกตัวหนึ่งที่ไม่ถูกเมทิลเลชันในบริเวณทั้งด้านบนและด้านล่างของจุดแตกเดิม ในส่วนของยีนที่ถูกทำลายโดยการแตกของสายคู่ เซลล์ลูกหลานครึ่งหนึ่งจะแสดงยีนนั้นในระดับสูง และเซลล์ลูกหลานอีกครึ่งหนึ่งจะแสดงยีนนั้นถูกยับยั้ง เมื่อโคลนของเซลล์เหล่านี้ถูกเลี้ยงไว้เป็นเวลาสามปี รูปแบบเมทิลเลชั่นใหม่จะคงอยู่ตลอดช่วงเวลานั้น[ 108 ]
ในหนูที่มีการแทรกการรวมตัวใหม่แบบโฮโมโลยีที่ควบคุมโดย CRISPR ในจีโนมของพวกมัน พบว่ามีการเติมเมทิลเลชั่นของไซต์ CpG จำนวนมากเพิ่มขึ้นภายในการแทรกที่เกี่ยวข้องกับการแตกของสายคู่[ 109 ]
การเชื่อมต่อปลายที่ไม่เหมือนกันอาจทำให้เกิดการเปลี่ยนแปลงของเครื่องหมายทางพันธุกรรมบางอย่าง
การเชื่อมต่อปลายที่ไม่เหมือนกัน (NHEJ) ของการแตกของสายคู่ DNA ในเซลล์สามารถทำให้เกิดเซลล์ลูกหลานบางส่วนที่มีการแสดงออกของยีนที่มีการแตกของสายคู่เริ่มต้นลดลง และเซลล์ลูกหลานบางส่วนมีการแสดงออกของยีนนั้นสูงขึ้นเนื่องจากการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่เกี่ยวข้องกับการซ่อมแซม NHEJ ความถี่ของการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่ทำให้เกิดการยับยั้งยีนหลังจากการซ่อมแซม NHEJ ของการแตกของสายคู่ DNA ในยีนนั้นอาจอยู่ที่ประมาณ 0.9% [ 106 ]
การสืบทอดโครงสร้าง
ในซีลิเอตเช่นTetrahymenaและParameciumเซลล์ที่มีพันธุกรรมเหมือนกันจะแสดงความแตกต่างที่ถ่ายทอดได้ในรูปแบบของแถวซีเลียบนพื้นผิวเซลล์ รูปแบบที่เปลี่ยนแปลงไปจากการทดลองสามารถส่งต่อไปยังเซลล์ลูกได้ ดูเหมือนว่าโครงสร้างที่มีอยู่จะทำหน้าที่เป็นแม่แบบสำหรับโครงสร้างใหม่ กลไกของการถ่ายทอดทางพันธุกรรมดังกล่าวยังไม่ชัดเจน แต่มีเหตุผลให้สันนิษฐานได้ว่าสิ่งมีชีวิตหลายเซลล์ก็ใช้โครงสร้างเซลล์ที่มีอยู่เพื่อประกอบเป็นโครงสร้างใหม่เช่นกัน[ 110 ] [ 111 ] [ 112 ]
การจัดตำแหน่งนิวคลีโอโซม
จีโนมยูคาริโอตมีนิวคลีโอโซม จำนวนมาก ตำแหน่งของนิวคลีโอโซมไม่ได้เกิดขึ้นโดยบังเอิญ และเป็นตัวกำหนดการเข้าถึง DNA ของโปรตีนควบคุม โปรโมเตอร์ที่ทำงานในเนื้อเยื่อต่าง ๆ แสดงให้เห็นว่ามีลักษณะตำแหน่งของนิวคลีโอโซมที่แตกต่างกัน[ 113 ]ซึ่งเป็นตัวกำหนดความแตกต่างในการแสดงออกของยีนและการจำแนกเซลล์ มีการแสดงให้เห็นว่าอย่างน้อยนิวคลีโอโซมบางส่วนยังคงอยู่ในเซลล์สเปิร์ม (ซึ่งฮิสโตนส่วนใหญ่แต่ไม่ใช่ทั้งหมดถูกแทนที่ด้วยโปรตามีน ) ดังนั้นตำแหน่งของนิวคลีโอโซมจึงสามารถถ่ายทอดทางพันธุกรรมได้ในระดับหนึ่ง การศึกษาล่าสุดได้ค้นพบความเชื่อมโยงระหว่างตำแหน่งของนิวคลีโอโซมกับปัจจัยทางพันธุกรรมอื่น ๆ เช่น การเมทิลเลชันของ DNA และการไฮดรอกซีเมทิลเลชัน[ 114 ]
ฮิสโตนชนิดต่างๆ
ฮิสโตนชนิดต่างๆจะถูกรวมเข้าในบริเวณเฉพาะของจีโนมโดยไม่สุ่ม ลักษณะทางชีวเคมีที่แตกต่างกันของฮิสโตนเหล่านี้สามารถส่งผลต่อการทำงานของจีโนมผ่านบทบาทของฮิสโตนในการควบคุมยีน[ 115 ]และการรักษาโครงสร้างของโครโมโซม[ 116 ]
โครงสร้างจีโนม
โครงสร้างสามมิติของจีโนม (จีโนม 3 มิติ) มีความซับซ้อน มีพลวัต และมีความสำคัญต่อการควบคุมการทำงานของจีโนมและกระบวนการนิวเคลียร์ เช่น การจำลองดีเอ็นเอ การถอดรหัส และการซ่อมแซมความเสียหายของดีเอ็นเอ[ 117 ]
หน้าที่และผลที่ตามมา
ในสมอง
หน่วยความจำ
การสร้างและการบำรุงรักษาความทรงจำเกิดจากการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่ทำให้เกิดการเปลี่ยนแปลงแบบไดนามิกที่จำเป็นในการถอดรหัสยีนซึ่งสร้างและต่ออายุความทรงจำในเซลล์ประสาท[ 30 ]
เหตุการณ์หนึ่งสามารถก่อให้เกิดปฏิกิริยาลูกโซ่ที่ส่งผลให้เกิดการเปลี่ยนแปลงเมทิลเลชันของยีนจำนวนมากในเซลล์ประสาท ซึ่งทำให้เกิดการแสดงเหตุการณ์หรือความทรงจำ[ 30 ]
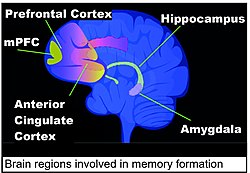
บริเวณสมองที่สำคัญในการสร้างความทรงจำ ได้แก่ ฮิปโปแคมปัส คอร์เทกซ์พรีฟรอนทัลส่วนกลาง (mPFC) คอร์เทกซ์ซิงกูเลตส่วนหน้า และอะมิกดาลา ดังแสดงในแผนภาพสมองมนุษย์ในส่วนนี้[ 118 ]
เมื่อความทรงจำที่แข็งแกร่งถูกสร้างขึ้น เช่นในหนูที่ได้รับการปรับสภาพความกลัวตามบริบท (CFC) หนึ่งในเหตุการณ์แรกๆ ที่เกิดขึ้นคือ การแตกของสายดีเอ็นเอคู่มากกว่า 100 จุดเกิดขึ้นโดยโทโปไอโซเมอเรส IIBในเซลล์ประสาทของฮิปโปแคมปัสและคอร์เทกซ์พรีฟรอนทัลส่วนกลาง (mPFC) [ 119 ]การแตกของสายคู่เหล่านี้อยู่ที่ตำแหน่งเฉพาะที่อนุญาตให้มีการกระตุ้นการถอดรหัสของยีนเริ่มต้นทันที (IEGs) ซึ่งมีความสำคัญในการสร้างความทรงจำ ทำให้เกิดการแสดงออกในmRNAโดยมีการถอดรหัส mRNA สูงสุดที่เจ็ดถึงสิบนาทีหลังจาก CFC [ 119 ] [ 120 ]
ยีน IEG ที่สำคัญสอง ยีนในการสร้างความทรงจำคือEGR1 [ 121 ]และตัวแปรโปรโมเตอร์ทางเลือกของDNMT3AคือDNMT3A2 [ 122 ]โปรตีน EGR1 จับกับ DNA ที่โมทีฟการจับ 5′-GCGTGGGCG-3′ หรือ 5′-GCGGGGGCGG-3' และมีตำแหน่งในจีโนมประมาณ 12,000 ตำแหน่งที่โปรตีน EGR1 สามารถจับได้[ 123 ]โปรตีน EGR1 จับกับ DNA ใน บริเวณ โปรโมเตอร์และเอนแฮนเซอร์ ของยีน EGR1 ดึงดูดเอนไซม์ดีเมทิลเลตTET1มายังบริเวณที่เกี่ยวข้อง และนำ TET1 ไปยังตำแหน่งประมาณ 600 ตำแหน่งบนจีโนม ซึ่ง TET1 สามารถดีเมทิลเลตและกระตุ้นยีนที่เกี่ยวข้องได้[ 123 ]

เอนไซม์ DNA เมทิลทรานสเฟอเรส DNMT3A1, DNMT3A2 และ DNMT3B สามารถเติมหมู่เมทิลให้กับไซโตซีน (ดูภาพในส่วนนี้) ที่ไซต์ CpG ในหรือใกล้โปรโมเตอร์ของยีนได้ ดังที่แสดงโดย Manzo et al. [ 124 ]เอนไซม์ DNA เมทิลทรานสเฟอเรสทั้งสามชนิดนี้แตกต่างกันในตำแหน่งการจับกับจีโนมและกิจกรรมการเติมหมู่เมทิลให้กับ DNA ที่ไซต์ควบคุมที่แตกต่างกัน Manzo et al. ระบุตำแหน่งจีโนม 3,970 แห่งที่อุดมไปด้วย DNMT3A1 โดยเฉพาะ 3,838 แห่งสำหรับ DNMT3A2 และ 3,432 แห่งสำหรับ DNMT3B เมื่อ DNMT3A2 ถูกเหนี่ยวนำขึ้นใหม่เป็น IEG (เมื่อเซลล์ประสาทถูกกระตุ้น) จะเกิดการเติมหมู่เมทิลให้กับไซโตซีนใหม่จำนวนมาก ซึ่งคาดว่าอยู่ในบริเวณเป้าหมายของ DNMT3A2 Oliviera et al. [ 122 ]พบว่าระดับ IEG ของ Dnmt3a2 ที่เหนี่ยวนำโดยกิจกรรมของเซลล์ประสาทในฮิปโปแคมปัสเป็นตัวกำหนดความสามารถในการสร้างความทรงจำระยะยาว
หนูสร้างความทรงจำแบบเชื่อมโยงระยะยาวหลังจากได้รับการปรับสภาพความกลัวตามบริบท[ 125 ] Duke และคณะ[ 29 ]พบว่า 24 ชั่วโมงหลังจาก CFC ในหนู ในเซลล์ประสาทฮิปโปแคมปัส ยีน 2,097 ยีน (9.17% ของยีนในจีโนมของหนู) มีการเปลี่ยนแปลงเมทิลเลชั่น เมื่อมีไซโตซีนที่มีเมทิลเลชั่นใหม่ปรากฏในไซต์ CpG ในบริเวณโปรโมเตอร์ของยีน ยีนมักจะถูกยับยั้ง และเมื่อมีไซโตซีนที่ไม่มีเมทิลเลชั่นใหม่ ยีนอาจถูกกระตุ้น[ 126 ]หลังจาก CFC มี 1,048 ยีนที่มีการแสดงออกของ mRNA ลดลง และ 564 ยีนที่มีการแสดงออกของ mRNA เพิ่มขึ้น ในทำนองเดียวกัน เมื่อหนูได้รับการ CFC หนึ่งชั่วโมงต่อมาในบริเวณฮิปโปแคมปัสของสมองหนู มี 675 ยีนที่ไม่มีเมทิลเลชั่น และ 613 ยีนที่มีเมทิลเลชั่นมากเกินไป[ 127 ]อย่างไรก็ตาม ความทรงจำไม่ได้คงอยู่ในฮิปโปแคมปัส แต่หลังจากสี่หรือห้าสัปดาห์ ความทรงจำจะถูกเก็บไว้ในคอร์เทกซ์ซิงกูเลตด้านหน้า[ 128 ]ในการศึกษาเกี่ยวกับหนูหลังจาก CFC Halder และคณะ[ 127 ]แสดงให้เห็นว่าสี่สัปดาห์หลังจาก CFC มีอย่างน้อย 1,000 ยีนที่มีการเมทิลเลชั่นแตกต่างกัน และมากกว่า 1,000 ยีนที่มีการแสดงออกแตกต่างกันในคอร์เทกซ์ซิงกูเลตด้านหน้า ในขณะเดียวกัน การเมทิลเลชั่นที่เปลี่ยนแปลงไปในฮิปโปแคมปัสก็กลับคืนสู่สภาพเดิม
การเปลี่ยนแปลงทางเอพิเจเนติกของเมทิลเลชันหลังจากการสร้างความทรงจำใหม่จะสร้างกลุ่ม mRNA ในนิวเคลียสที่แตกต่างกัน ดังที่ Bernstein ได้ทบทวนไว้[ 30 ]ส่วนผสมใหม่ของmRNA ในนิวเคลียสที่กำหนดโดยเอพิเจเนติก มักจะถูกบรรจุลงในเม็ดประสาทหรือmessenger RNPซึ่งประกอบด้วย mRNA หน่วยย่อยไรโบโซมขนาดเล็กและขนาดใหญ่ปัจจัยเริ่มต้นการแปล และโปรตีนที่จับกับ RNA ซึ่งควบคุมการทำงานของ mRNA เม็ดประสาทเหล่านี้ถูกขนส่งจากนิวเคลียสของเซลล์ประสาทและถูกกำหนดทิศทางตามบริเวณที่ไม่ได้รับการแปล 3′ ของ mRNA ในเม็ดประสาท ("รหัสไปรษณีย์") ไปยังเดนไดรต์ ของเซลล์ประสาท โดยประมาณ 2,500 mRNA อาจอยู่ในเดนไดรต์ของเซลล์ประสาทพีระมิดในฮิปโปแคมปัส และอาจมีทรานสคริปต์ประมาณ 450 รายการอยู่ในปลายประสาทก่อนซิแนปส์ที่กระตุ้น (หนามเดนไดรต์) การเปลี่ยนแปลงของลำดับสารถอดรหัส (ขึ้นอยู่กับการเปลี่ยนแปลงทางอีพีเจเนติกในนิวเคลียสของเซลล์ประสาท) มีความไวในการตอบสนองต่อสัญญาณที่แตกต่างกัน ซึ่งเป็นพื้นฐานของความยืดหยุ่นของไซแนปส์ที่เปลี่ยนแปลงไป ความยืดหยุ่นของไซแนปส์ที่เปลี่ยนแปลงไปนี้มักถูกพิจารณาว่าเป็นรากฐานทางเคมีประสาทของการเรียนรู้และความทรงจำ
ความชรา
เอพิเจเนติกส์มีบทบาทสำคัญในการเสื่อมสภาพของสมอง และ ภาวะความเสื่อมถอยทางสติปัญญาที่เกี่ยวข้องกับอายุ ซึ่งมีความเกี่ยวข้องกับการยืดอายุขัย[ 129 ] [ 130 ] [ 131 ] [ 132 ] [ 133 ]
อื่นๆ และทั่วไป
ในวัยผู้ใหญ่ การเปลี่ยนแปลงในเอพิเจโนมมีความสำคัญต่อการทำงานของระบบการรับรู้ระดับสูงต่างๆ การทำงานผิดปกติของกลไกเอพิเจเนติกส์เกี่ยวข้องกับความผิดปกติและโรคทางระบบประสาทเสื่อม การดัดแปลงเอพิเจเนติกส์ในเซลล์ประสาทมีความเปลี่ยนแปลงได้และย้อนกลับได้[ 134 ]การควบคุมเอพิเจเนติกส์ส่งผลต่อการทำงานของเซลล์ประสาท ส่งผลต่อการเรียนรู้ ความจำ และกระบวนการรับรู้ ด้านอื่นๆ [ 135 ]
เหตุการณ์ในช่วงต้น รวมถึงในช่วงการพัฒนาของตัวอ่อนสามารถส่งผลต่อการพัฒนา การรับรู้ และผลลัพธ์ด้านสุขภาพผ่านกลไกทางพันธุกรรม [ 136 ]
กลไกเอพิเจเนติกส์ได้รับการเสนอให้เป็น "กลไกโมเลกุลที่เป็นไปได้สำหรับผลกระทบของฮอร์โมน ภายในร่างกาย ต่อการจัดระเบียบวงจรสมองที่กำลังพัฒนา" [ 137 ]
สารอาหารอาจมีปฏิสัมพันธ์กับเอพิเจโนมเพื่อ "ปกป้องหรือส่งเสริมกระบวนการรับรู้ตลอดช่วงชีวิต" [ 138 ] [ 139 ]
การทบทวนชี้ให้เห็นว่าผลกระทบทางชีววิทยาประสาทของการออกกำลังกายผ่านเอพิเจเนติกส์ดูเหมือนจะเป็น "หัวใจสำคัญในการสร้าง 'ความทรงจำเอพิเจเนติกส์' เพื่อส่งผลต่อการทำงานของสมองและพฤติกรรมในระยะยาว" และอาจถ่ายทอดทางพันธุกรรมได้ด้วย[ 140 ]
ด้วยไซแนปส์ แอกโซ-ซิเลียรี จะมีการสื่อสารระหว่างแอกซอนเซโรโทเนอร์จิก และซิเลียหลักคล้ายเสาอากาศของเซลล์ประสาทพีระมิดCA1 ซึ่งเปลี่ยนแปลงสถานะ เอพิเจเนติกของเซลล์ประสาทในนิวเคลียสผ่านการส่งสัญญาณที่แตกต่างจากที่เยื่อหุ้มพลาสมา (และในระยะยาว) [ 141 ] [ 142 ]
เอพิเจเนติกส์ยังมีบทบาทสำคัญในการวิวัฒนาการของสมองในมนุษย์ด้วย[ 143 ]
การพัฒนา
เอพิเจเนติกส์เชิงพัฒนาการสามารถแบ่งออกเป็นเอพิเจเนติกส์แบบกำหนดไว้ล่วงหน้าและแบบความน่าจะเป็น เอพิเจเนติกส์แบบกำหนดไว้ล่วงหน้าคือการเคลื่อนไหวแบบทิศทางเดียวจากพัฒนาการเชิงโครงสร้างใน DNA ไปสู่การเจริญเติบโตเชิงหน้าที่ของโปรตีน คำว่า "กำหนดไว้ล่วงหน้า" ในที่นี้หมายความว่าพัฒนาการนั้นถูกกำหนดไว้และสามารถคาดการณ์ได้ ในทางกลับกัน เอพิเจเนติกส์แบบความน่าจะเป็นคือพัฒนาการเชิงโครงสร้างและหน้าที่แบบสองทิศทางโดยอาศัยประสบการณ์และการพัฒนาแบบหล่อหลอมจากภายนอก[ 144 ]
การถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกในเซลล์ร่างกาย โดยเฉพาะอย่างยิ่งผ่านการดัดแปลงโคเวเลนต์ของ DNA และฮิสโตน และ การจัดตำแหน่งนิว คลีโอโซมใหม่ มีความสำคัญอย่างยิ่งต่อการพัฒนาของสิ่งมีชีวิตยูคาริโอตหลายเซลล์[ 114 ]ลำดับจีโนมคงที่ (ยกเว้นบางกรณีที่สำคัญ) แต่เซลล์จะแตกต่างไปเป็นเซลล์หลายประเภท ซึ่งทำหน้าที่ต่างกัน และตอบสนองต่อสิ่งแวดล้อมและการส่งสัญญาณระหว่างเซลล์แตกต่างกัน ดังนั้น เมื่อแต่ละบุคคลพัฒนาขึ้นมอร์โฟเจนจะกระตุ้นหรือยับยั้งยีนในลักษณะที่ถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติก ทำให้เซลล์มีความทรงจำ ในสัตว์เลี้ยงลูกด้วยนม เซลล์ส่วนใหญ่จะแตกต่างไปเป็นเซลล์ชนิดสุดท้าย โดยมีเพียงเซลล์ต้นกำเนิด เท่านั้น ที่ยังคงความสามารถในการแตกต่างไปเป็นเซลล์หลายประเภท ("totipotency" และ "multipotency") ในสัตว์เลี้ยงลูกด้วยนมเซลล์ต้นกำเนิดบางส่วนยังคงผลิตเซลล์ที่แตกต่างใหม่ต่อไปตลอดชีวิต เช่น ในกระบวนการสร้างเซลล์ประสาทแต่สัตว์เลี้ยงลูกด้วยนมไม่สามารถตอบสนองต่อการสูญเสียเนื้อเยื่อบางส่วนได้ ตัวอย่างเช่น ความไม่สามารถสร้างแขนขาขึ้นใหม่ได้ ซึ่งสัตว์ชนิดอื่นบางชนิดสามารถทำได้ การดัดแปลงเอพิเจเนติกส์ควบคุมการเปลี่ยนผ่านจากเซลล์ต้นกำเนิดประสาทไปเป็นเซลล์ต้นกำเนิดเกลีย (ตัวอย่างเช่น การแยกตัวเป็นโอลิโกเดนโดรไซต์ถูกควบคุมโดยการกำจัดอะเซทิลและการเติมเมทิลของฮิสโตน) [ 145 ]ต่างจากสัตว์ เซลล์พืชไม่แยกตัวอย่างสมบูรณ์ แต่ยังคงมีศักยภาพในการสร้างพืชใหม่ได้ แม้ว่าพืชจะใช้กลไกเอพิเจเนติกส์หลายอย่างเช่นเดียวกับสัตว์ เช่นการปรับโครงสร้างโครมาตินแต่ก็มีสมมติฐานว่าเซลล์พืชบางชนิดไม่ใช้หรือต้องการ "ความทรงจำของเซลล์" ในการรีเซ็ตรูปแบบการแสดงออกของยีนโดยใช้ข้อมูลตำแหน่งจากสิ่งแวดล้อมและเซลล์รอบข้างเพื่อกำหนดชะตากรรมของพวกมัน[ 146 ]
การเปลี่ยนแปลงทางเอพิเจเนติกส์สามารถเกิดขึ้นได้เพื่อตอบสนองต่อการสัมผัสกับสิ่งแวดล้อม – ตัวอย่างเช่น การเสริมอาหารของมารดาด้วยเจนิสเตอิน (250 มก./กก.) ทำให้เกิดการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่ส่งผลต่อการแสดงออกของ ยีน อะกูติซึ่งส่งผลต่อสีขน น้ำหนัก และแนวโน้มที่จะเป็นมะเร็ง[ 147 ] [ 148 ] [ 149 ]งานวิจัยที่กำลังดำเนินอยู่มุ่งเน้นไปที่การสำรวจผลกระทบของสารก่อความพิการแต่กำเนิด อื่นๆ ที่รู้จัก เช่นภาวะผิดปกติของตัวอ่อนจากเบาหวานต่อ ลาย เซ็นเมทิลเลชัน[ 150 ]
ผลการศึกษาหนึ่งที่ก่อให้เกิดข้อถกเถียงชี้ให้เห็นว่าประสบการณ์ที่กระทบกระเทือนจิตใจอาจก่อให้เกิดสัญญาณเอพิเจเนติกส์ที่สามารถส่งต่อไปยังรุ่นต่อๆ ไปได้ หนูถูกฝึกโดยใช้ไฟฟ้าช็อตที่เท้าให้กลัวกลิ่นดอกซากุระ นักวิจัยรายงานว่าลูกหลานของหนูมีความรังเกียจกลิ่นนี้มากขึ้น[ 151 ] [ 152 ]พวกเขาเสนอว่าการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่เพิ่มการแสดงออกของยีน แทนที่จะเป็นในดีเอ็นเอเอง ในยีน M71 ซึ่งควบคุมการทำงานของตัวรับกลิ่นในจมูกที่ตอบสนองต่อกลิ่นดอกซากุระโดยเฉพาะ มีการเปลี่ยนแปลงทางกายภาพที่สัมพันธ์กับการทำงานของระบบรับกลิ่นในสมองของหนูที่ได้รับการฝึกฝนและลูกหลานของพวกมัน มีการรายงานคำวิจารณ์หลายประการ รวมถึงพลังทางสถิติที่ต่ำของการศึกษาเป็นหลักฐานของความผิดปกติบางอย่าง เช่น อคติในการรายงานผล[ 153 ]เนื่องจากข้อจำกัดของขนาดตัวอย่าง จึงมีความเป็นไปได้ที่ผลกระทบจะไม่ได้รับการแสดงให้เห็นอย่างมีนัยสำคัญทางสถิติแม้ว่าจะมีอยู่จริงก็ตาม คำวิจารณ์ชี้ให้เห็นว่าความน่าจะเป็นที่การทดลองทั้งหมดที่รายงานจะแสดงผลลัพธ์เชิงบวกหากปฏิบัติตามโปรโตคอลที่เหมือนกัน โดยสมมติว่าผลกระทบที่อ้างนั้นมีอยู่จริง มีเพียง 0.4% เท่านั้น ผู้เขียนยังไม่ได้ระบุว่าหนูตัวใดเป็นพี่น้องกัน และถือว่าหนูทุกตัวเป็นอิสระทางสถิติ[ 154 ]นักวิจัยดั้งเดิมชี้ให้เห็นผลลัพธ์เชิงลบในภาคผนวกของเอกสารที่คำวิจารณ์ละเว้นในการคำนวณ และดำเนินการติดตามว่าหนูตัวใดเป็นพี่น้องกันในอนาคต[ 155 ]
ข้ามรุ่น
กลไกเอพิเจเนติกส์เป็นส่วนที่จำเป็นของต้นกำเนิดวิวัฒนาการของ การ จำแนกเซลล์[ 156 ]แม้ว่าโดยทั่วไปแล้วเอพิเจเนติกส์ในสิ่งมีชีวิตหลายเซลล์จะถูกมองว่าเป็นกลไกที่เกี่ยวข้องกับการจำแนก โดยมีรูปแบบเอพิเจเนติกส์ "รีเซ็ต" เมื่อสิ่งมีชีวิตสืบพันธุ์ แต่ก็มีการสังเกตการถ่ายทอดทางเอพิเจเนติกส์ข้ามรุ่น (เช่น ปรากฏการณ์พารามิวเทชันที่พบในข้าวโพด ) แม้ว่าลักษณะเอพิเจเนติกส์ข้ามรุ่นส่วนใหญ่จะค่อยๆ หายไปในหลายชั่วอายุคน แต่ก็ยังมีความเป็นไปได้ที่เอพิเจเนติกส์ข้ามรุ่นอาจเป็นอีกแง่มุมหนึ่งของวิวัฒนาการและการปรับตัว ดังที่กล่าวไว้ข้างต้น บางคนนิยามเอพิเจเนติกส์ว่าสามารถถ่ายทอดได้
เชื้อพันธุ์ที่ถูกแยกหรือกำแพงไวส์แมนน์เป็นลักษณะเฉพาะของสัตว์ และการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์พบได้ทั่วไปในพืชและจุลินทรีย์อีวา จาบลอนกา , มาริออน เจ. แลมบ์และเอเตียน ดันชิน ได้โต้แย้งว่าผลกระทบเหล่านี้อาจต้องการการปรับปรุงกรอบแนวคิดมาตรฐานของการสังเคราะห์สมัยใหม่และเรียกร้องให้มีการสังเคราะห์วิวัฒนาการแบบขยาย [ 157 ] [ 158 ] [ 159 ] นักชีววิทยาวิวัฒนาการคนอื่นๆ เช่นจอห์น เมย์นาร์ด สมิธได้รวมการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์เข้าไว้ในแบบจำลองพันธุศาสตร์ประชากร[ 160 ]หรือมีความสงสัยอย่างเปิดเผยเกี่ยวกับการสังเคราะห์วิวัฒนาการแบบขยาย ( ไมเคิล ลินช์ ) [ 161 ]โทมัส ดิกกินส์ และกาซี ราห์มาน ระบุว่ากลไกเอพิเจเนติกส์ เช่น การเมทิลเลชั่นของดีเอ็นเอและการดัดแปลงฮิสโตนนั้นได้รับการถ่ายทอดทางพันธุกรรมภายใต้การควบคุมของการคัดเลือกโดยธรรมชาติดังนั้นจึงสอดคล้องกับ"การสังเคราะห์สมัยใหม่" ก่อน หน้า นี้ [ 162 ]
การถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์แตกต่างจากการถ่ายทอดทางพันธุกรรมแบบดั้งเดิมในสองประเด็นสำคัญ ซึ่งส่งผลสำคัญต่อวิวัฒนาการ ได้แก่:
- อัตราการเกิด epimutation อาจเร็วกว่าอัตราการกลายพันธุ์มาก[ 163 ]
- การกลายพันธุ์ของเอพิเจเนซิสสามารถย้อนกลับได้ง่ายกว่า[ 164 ]
ในพืช การกลายพันธุ์ของเมทิลเลชั่นของ DNA ที่ถ่ายทอดทางพันธุกรรมมีโอกาสเกิดขึ้นมากกว่าการกลายพันธุ์ของ DNA ถึง 100,000 เท่า[ 165 ]องค์ประกอบที่ถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์ เช่น ระบบ PSI+สามารถทำหน้าที่เป็น "ตัวหยุด" ซึ่งดีพอสำหรับการปรับตัวในระยะสั้นที่ช่วยให้สายพันธุ์อยู่รอดได้นานพอสำหรับการกลายพันธุ์และ/หรือการรวมตัวใหม่เพื่อผสมผสานการเปลี่ยนแปลงฟีโนไทป์ที่ปรับตัวเข้า กับพันธุกรรม [ 166 ]การมีอยู่ของความเป็นไปได้นี้เพิ่มความสามารถในการวิวัฒนาการของสายพันธุ์
มีรายงานกรณี การถ่ายทอดลักษณะทางพันธุกรรมข้ามรุ่นมากกว่า 100 กรณีในสิ่งมีชีวิตหลากหลายชนิด รวมถึงโปรคาริโอต พืช และสัตว์[ 167 ]ตัวอย่างเช่นผีเสื้อเสื้อคลุมไว้ทุกข์จะเปลี่ยนสีผ่านการเปลี่ยนแปลงของฮอร์โมนเพื่อตอบสนองต่อการทดลองที่อุณหภูมิแตกต่างกัน[ 168 ]
เชื้อราเส้นใยNeurospora crassaเป็นระบบแบบจำลองที่โดดเด่นสำหรับการทำความเข้าใจการควบคุมและการทำงานของการเมทิลเลชันของไซโตซีน ในสิ่งมีชีวิตนี้ การเมทิลเลชันของ DNA เกี่ยวข้องกับซากของระบบป้องกันจีโนมที่เรียกว่า RIP (repeat-induced point mutation) และปิดการแสดงออกของยีนโดยการยับยั้งการยืดตัวของการถอดรหัส[ 169 ]
พ รีออน PSI ของยีสต์ถูกสร้างขึ้นโดยการเปลี่ยนแปลงโครงสร้างของปัจจัยยุติการแปล ซึ่งจะถูกส่งต่อไปยังเซลล์ลูก สิ่งนี้สามารถให้ข้อได้เปรียบในการอยู่รอดภายใต้สภาวะที่ไม่เอื้ออำนวย ซึ่งเป็นตัวอย่างของการควบคุมเอพิเจเนติกส์ที่ช่วยให้สิ่งมีชีวิตเซลล์เดียวสามารถตอบสนองต่อความเครียดจากสิ่งแวดล้อมได้อย่างรวดเร็ว พรีออนสามารถมองได้ว่าเป็นตัวแทนเอพิเจเนติกส์ที่สามารถชักนำให้เกิดการเปลี่ยนแปลงฟีโนไทป์โดยไม่ต้องแก้ไขจีโนม[ 170 ]
การตรวจจับเครื่องหมายเอพิเจเนติกส์โดยตรงในจุลินทรีย์เป็นไปได้ด้วยการจัดลำดับแบบเรียลไทม์ของโมเลกุลเดี่ยวซึ่งความไวของพอลิเมอเรสทำให้สามารถวัดเมทิลเลชันและการดัดแปลงอื่นๆ ในขณะที่โมเลกุล DNA กำลังถูกจัดลำดับ[ 171 ]โครงการหลายโครงการได้แสดงให้เห็นถึงความสามารถในการรวบรวมข้อมูลเอพิเจเนติกส์ทั่วทั้งจีโนมในแบคทีเรีย[ 172 ] [ 173 ] [ 174 ] [ 175 ]
เอพิเจเนติกส์ในแบคทีเรีย

แม้ว่าเอพิเจเนติกส์จะมีความสำคัญพื้นฐานในยูคาริโอตโดยเฉพาะอย่างยิ่ง ใน เมตาโซแอนแต่ในแบคทีเรียกลับมีบทบาทที่แตกต่างออกไป[ 176 ]ที่สำคัญที่สุดคือ ยูคาริโอตใช้กลไกเอพิเจเนติกส์เป็นหลักในการควบคุมการแสดงออกของยีน ซึ่งแบคทีเรียแทบจะไม่ทำเช่นนั้น อย่างไรก็ตาม แบคทีเรียใช้การเมทิลเลชันของดีเอ็นเอหลังการจำลองแบบอย่างแพร่หลายเพื่อควบคุมเอพิเจเนติกส์ของปฏิสัมพันธ์ระหว่างดีเอ็นเอและโปรตีน แบคทีเรียยังใช้ การเมทิลเลชัน ของอะดีนีนใน ดีเอ็นเอ (แทนที่จะเป็นการ เมทิลเลชัน ของไซโตซีน ในดีเอ็นเอ ) เป็นสัญญาณเอพิเจเนติกส์ การเมทิลเลชันของอะดีนีนในดีเอ็นเอมีความสำคัญต่อความรุนแรงของแบคทีเรียในสิ่งมีชีวิต เช่นEscherichia coli , Salmonella , Vibrio , Yersinia , HaemophilusและBrucellaในAlphaproteobacteriaการเมทิลเลชันของอะดีนีนจะควบคุมวงจรเซลล์และเชื่อมโยงการถอดรหัสยีนกับการจำลองแบบดีเอ็นเอ ในGammaproteobacteriaการเมทิลเลชันของอะดีนีนให้สัญญาณสำหรับการจำลองดีเอ็นเอ การแยกโครโมโซม การซ่อมแซมความผิดพลาด การบรรจุแบคทีริโอเฟจ กิจกรรมทรานสโพเซส และการควบคุมการแสดงออกของยีน[ 170 ] [ 177 ]มีสวิตช์ทางพันธุกรรมที่ควบคุมStreptococcus pneumoniae (นิวโมค็อกคัส) ซึ่งช่วยให้แบคทีเรียสามารถเปลี่ยนลักษณะเฉพาะของมันแบบสุ่มไปเป็นสถานะทางเลือกหกสถานะที่อาจปูทางไปสู่การพัฒนาวัคซีนที่ดีขึ้น แต่ละรูปแบบถูกสร้างขึ้นแบบสุ่มโดยระบบเมทิลเลชันแบบแปรผันเฟส ความสามารถของนิวโมค็อกคัสในการก่อให้เกิดการติดเชื้อร้ายแรงนั้นแตกต่างกันในแต่ละสถานะทั้งหกนี้ ระบบที่คล้ายกันนี้มีอยู่ในสกุลแบคทีเรียอื่นๆ[ 178 ]ในBacillotaเช่นClostridioides difficileการเมทิลเลชันของอะดีนีนควบคุม การสร้างสปอร์ การ สร้าง ไบโอฟิล์มและการปรับตัวเข้ากับโฮสต์[ 179 ]
ยา
เอพิเจเนติกส์มีศักยภาพในการประยุกต์ใช้ทางการแพทย์ที่หลากหลายและมากมาย[ 180 ]
ฝาแฝด
การเปรียบเทียบโดยตรงของฝาแฝดเหมือนกันถือเป็นแบบจำลองที่เหมาะสมที่สุดสำหรับการตรวจสอบเอพิเจเนติกส์สิ่งแวดล้อมในกรณีของมนุษย์ที่มีการสัมผัสกับสิ่งแวดล้อมที่แตกต่างกัน ฝาแฝดโมโนไซโกติก (ฝาแฝดเหมือนกัน) จะแยกแยะเอพิเจเนติกส์ไม่ได้ในช่วงปีแรก ๆ ในขณะที่ฝาแฝดที่โตขึ้นจะมีความแตกต่างอย่างเห็นได้ชัดในปริมาณโดยรวมและการกระจายตัวของจีโนมของดีเอ็นเอ 5-เมทิลไซโตซีนและการอะเซทิเลชันของฮิสโตน[ 10 ]คู่ฝาแฝดที่ใช้ชีวิตร่วมกันน้อยกว่าและ/หรือมีความแตกต่างกันมากขึ้นในประวัติทางการแพทย์จะเป็นคู่ที่แสดงความแตกต่างมากที่สุดในระดับของ ดีเอ็นเอ 5-เมทิลไซโตซีนและการอะเซทิเลชันของฮิสโตน H3 และ H4 [ 181 ]
แฝดไดไซโกติก (แฝดต่างไข่) และแฝดโมโนไซโกติก (แฝดเหมือน) แสดงหลักฐานของอิทธิพลเอพิเจเนติกในมนุษย์[ 181 ] [ 182 ] [ 183 ]ความแตกต่างของลำดับดีเอ็นเอที่พบได้มากในการศึกษาแบบบุคคลเดียวไม่ได้รบกวนการวิเคราะห์ ความแตกต่างของสิ่งแวดล้อมสามารถสร้างผลกระทบเอพิเจเนติกในระยะยาว และแฝดโมโนไซโกติกชนิดย่อยที่แตกต่างกันในด้านพัฒนาการอาจแตกต่างกันในแง่ของความไวต่อการไม่สอดคล้องกันจากมุมมองเอพิเจเนติก[ 184 ]
การ ศึกษา แบบความเร็วสูงซึ่งหมายถึงเทคโนโลยีที่ตรวจสอบเครื่องหมายทางพันธุกรรมจำนวนมาก มุ่งเน้นไปที่ความแตกต่างทางเอพิเจเนติกส์ระหว่างฝาแฝดโมโนไซโกติก เพื่อเปรียบเทียบการเปลี่ยนแปลงทั่วโลกและเฉพาะตำแหน่งในเมทิลเลชั่นของดีเอ็นเอและการดัดแปลงฮิสโตนในตัวอย่างคู่ฝาแฝดโมโนไซโกติก 40 คู่[ 181 ]ในกรณีนี้ ศึกษาเฉพาะคู่ฝาแฝดที่มีสุขภาพดีเท่านั้น แต่มีช่วงอายุที่กว้าง ตั้งแต่ 3 ถึง 74 ปี ข้อสรุปสำคัญประการหนึ่งจากการศึกษานี้คือ มีการสะสมความแตกต่างทางเอพิเจเนติกส์ที่ขึ้นอยู่กับอายุระหว่างพี่น้องสองคนของคู่ฝาแฝด การสะสมนี้ชี้ให้เห็นถึงการมีอยู่ของ "การเปลี่ยนแปลง" ทางเอพิเจเนติกส์ การเปลี่ยนแปลงทางเอพิเจเนติกส์เป็นคำที่ใช้เรียกการดัดแปลงทางเอพิเจเนติกส์ที่เกิดขึ้นโดยตรงตามอายุ ในขณะที่อายุเป็นปัจจัยเสี่ยงที่ทราบกันดีสำหรับหลายโรค พบว่าเมทิลเลชั่นที่เกี่ยวข้องกับอายุเกิดขึ้นแตกต่างกันที่ตำแหน่งเฉพาะตามจีโนม เมื่อเวลาผ่านไป สิ่งนี้อาจส่งผลให้เกิดความแตกต่างที่วัดได้ระหว่างอายุทางชีวภาพและอายุตามปฏิทิน พบว่าการเปลี่ยนแปลงทางเอพิเจเนติกส์สะท้อนถึงวิถีชีวิตและอาจทำหน้าที่เป็นตัวบ่งชี้ทางชีวภาพ เชิงฟังก์ชัน ของโรคก่อนที่จะถึงเกณฑ์ ทางคลินิก [ 185 ]
การศึกษาล่าสุดที่วิเคราะห์แฝดโมโนไซโกติก 114 คู่ และแฝดไดไซโกติก 80 คู่ เพื่อตรวจสอบสถานะการเมทิลเลชั่นของดีเอ็นเอในบริเวณจีโนมที่ไม่ซ้ำกันประมาณ 6,000 บริเวณ สรุปได้ว่าความคล้ายคลึงทางเอพิเจเนติกส์ในช่วงเวลาที่บลาสโตซิสต์แบ่งตัวอาจมีส่วนทำให้เกิดความคล้ายคลึงทางฟีโนไทป์ในแฝดโมโนไซโกติกด้วยกัน ซึ่งสนับสนุนแนวคิดที่ว่าสภาพแวดล้อมจุลภาคในระยะเริ่มต้นของการพัฒนาตัวอ่อนมีความสำคัญอย่างยิ่งต่อการสร้างเครื่องหมายเอพิเจเนติกส์[ 182 ]โรคทางพันธุกรรมแต่กำเนิดเป็นที่เข้าใจกันดี และเป็นที่ชัดเจนว่าเอพิเจเนติกส์สามารถมีบทบาทได้ เช่น ในกรณีของกลุ่มอาการแองเจิลแมนและกลุ่มอาการพราเดอร์-วิลลี โรคเหล่านี้เป็นโรคทางพันธุกรรมปกติที่เกิดจากการลบยีนหรือการปิดใช้งานยีน แต่พบได้บ่อยผิดปกติเนื่องจากบุคคลส่วนใหญ่เป็นเฮมิไซโกสเนื่องจากการประทับตราทางจีโนมดังนั้นการกำจัดยีนเพียงยีนเดียวก็เพียงพอที่จะทำให้เกิดโรคได้ ในขณะที่กรณีส่วนใหญ่จะต้องมีการกำจัดยีนทั้งสองสำเนา[ 186 ]
การประทับตราทางพันธุกรรม
ความผิดปกติในมนุษย์บางอย่างเกี่ยวข้องกับการประทับตราทางพันธุกรรม ซึ่งเป็นปรากฏการณ์ในสัตว์เลี้ยงลูกด้วยนมที่พ่อและแม่ถ่ายทอดรูปแบบเอพิเจเนติกส์ที่แตกต่างกันสำหรับตำแหน่งทางพันธุกรรมเฉพาะในเซลล์สืบพันธุ์ ของพวก เขา[ 187 ]กรณีที่รู้จักกันดีที่สุดของการประทับตราในความผิดปกติในมนุษย์คือกลุ่มอาการแองเจิลแมนและกลุ่มอาการพราเดอร์-วิลลีซึ่งทั้งสองกลุ่มอาการนี้สามารถเกิดขึ้นได้จากการกลายพันธุ์ทางพันธุกรรมเดียวกัน คือการลบส่วนหนึ่งของโครโมโซม 15qและกลุ่มอาการเฉพาะที่จะพัฒนาขึ้นนั้นขึ้นอยู่กับว่าการกลายพันธุ์นั้นได้รับการถ่ายทอดมาจากแม่หรือพ่อของเด็ก[ 188 ]
ในการศึกษาที่ Överkalixหลานชายทางฝั่งพ่อ (แต่ไม่ใช่ทางฝั่งแม่) [ 189 ]ของชายชาวสวีเดนที่เผชิญกับภาวะอดอยากในช่วงก่อนวัยรุ่นในศตวรรษที่ 19 มีโอกาสเสียชีวิตจากโรคหัวใจและหลอดเลือดน้อยลง หากอาหารอุดมสมบูรณ์ อัตรา การเสียชีวิต จากโรคเบาหวานในหลานชายจะเพิ่มขึ้น ซึ่งบ่งชี้ว่านี่เป็นการถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์ข้ามรุ่น[ 190 ]ในทางกลับกัน พบผลตรงกันข้ามในเพศหญิง – หลานสาวทางฝั่งพ่อ (แต่ไม่ใช่ทางฝั่งแม่) ของผู้หญิงที่ประสบกับภาวะอดอยากขณะอยู่ในครรภ์ (และดังนั้นในขณะที่ไข่ของพวกเธอกำลังก่อตัว) มีอายุขัยเฉลี่ยสั้นกว่า[ 191 ]
ตัวอย่างของยาที่เปลี่ยนแปลงการแสดงออกของยีนจากเหตุการณ์ทางเอพิเจเนติกส์
การใช้ ยา ปฏิชีวนะ กลุ่มเบตา-แลคแทม สามารถเปลี่ยนแปลงการทำงานของตัวรับกลูตาเมตและการทำงานของไซโคลสปอรินต่อปัจจัยการถอดรหัสหลายตัว นอกจากนี้ลิเธียมยังสามารถส่งผลกระทบต่อออโตฟาจีของโปรตีนที่ผิดปกติ และ ยา โอปิออยด์จากการใช้เรื้อรังสามารถเพิ่มการแสดงออกของยีนที่เกี่ยวข้องกับลักษณะการเสพติดได้[ 192 ]
โภชนาการของพ่อแม่ การสัมผัสกับ ความเครียด หรือ สารเคมีที่รบกวนต่อมไร้ท่อในครรภ์[ 193 ]ผลกระทบจากฝ่ายชายที่มีต่อแม่ เช่น การดึงดูดคุณภาพของคู่ครองที่แตกต่างกัน อายุของแม่และพ่อ และเพศของลูก อาจส่งผลต่อการแสดงออกของเอพิมิวเทชันในเซลล์สืบพันธุ์ในลูกหลาน และระดับของการถ่ายทอดทางพันธุกรรมระหว่างรุ่นให้คงที่ตลอดช่วงชีวิต[ 194 ]อย่างไรก็ตาม ยังไม่ชัดเจนว่าผลกระทบทางเอพิเจเนติกส์สามารถส่งต่อข้ามรุ่นได้หรือไม่และในระดับใด โดยเฉพาะในมนุษย์[ 195 ] [ 196 ]
การเสพติด
การเสพติด เป็นความผิดปกติของ ระบบรางวัลของสมองซึ่งเกิดขึ้นผ่าน กลไก การถอดรหัสและการเปลี่ยนแปลงทางพันธุกรรมของระบบประสาท และเกิดขึ้นเมื่อเวลาผ่านไปจากการได้รับสารกระตุ้นการเสพติดในระดับสูงเรื้อรัง (เช่น มอร์ฟีน โคเคน การมีเพศสัมพันธ์ การพนัน) [ 197 ] [ 198 ] [ 199 ] มีการสังเกต การถ่ายทอด ลักษณะทางพันธุกรรมของการเสพติดข้ามรุ่นในงานวิจัยทางคลินิกก่อนหน้านี้[ 200 ] [ 201 ]อย่างไรก็ตาม ยังไม่มีหลักฐานที่แน่ชัดสนับสนุนความคงอยู่ของผลกระทบทางพันธุกรรมข้ามรุ่นในมนุษย์ ตัวอย่างเช่น ผลกระทบทางพันธุกรรมของการได้รับควันบุหรี่ในครรภ์ที่พบในเหลนที่ไม่ได้รับควันบุหรี่[ 195 ]
เอพิเจเนติกส์และมะเร็ง
การศึกษาชี้ให้เห็นว่าเอพิเจเนติกส์ยังสามารถมีบทบาทในโรคมะเร็ง โดยเฉพาะความผิดปกติทางเอพิเจเนติกส์และการเริ่มต้นและการลุกลามของโรคมะเร็ง[ 202 ] ซึ่งนำไปสู่การพัฒนาการรักษาด้วยเอพิเจเนติกส์ รวมถึงยาที่ได้รับการอนุมัติจาก FDA เมื่อเร็วๆ นี้ เช่น สารยับยั้ง DNA เมทิลทรานสเฟอเรส (DNMT) สารยับยั้งฮิสโตนดีอะเซทิเลส (HDACi) และสารยับยั้งฮิสโตนเมทิลทรานสเฟอเรส (HMTi) ซึ่งปัจจุบันรวมอยู่ในการรักษาโรคมะเร็งที่มีอยู่แล้ว[ 203 ] โรคมะเร็งถือเป็นทั้งโรคทางพันธุกรรมและโรคทางเอพิเจเนติกส์[ 204 ]ซึ่งรวมถึงองค์ประกอบต่างๆ เช่น การเมทิลเลชั่นของ DNA การดัดแปลงฮิสโตน และการแสดงออกของไมโครอาร์เอ็นเอ ซึ่งอาจนำไปสู่การเจริญเติบโตของเซลล์มะเร็ง[ 204 ]
วิจัย
ข้อมูลทางพันธุกรรมสองรูปแบบ ได้แก่ พันธุกรรมและเอพิเจเนติกส์ เรียกรวมกันว่าการถ่ายทอดแบบคู่ สมาชิกในกลุ่ม APOBEC/AID ของไซโตซีนดีอะมิเนสอาจมีอิทธิพลต่อการถ่ายทอดทางพันธุกรรมและเอพิเจเนติกส์พร้อมกันโดยใช้กลไกโมเลกุลที่คล้ายคลึงกัน และอาจเป็นจุดเชื่อมต่อระหว่างกระบวนการที่แยกส่วนกันตามแนวคิดเหล่านี้[ 205 ]
ยาปฏิชีวนะ กลุ่มฟลูออโรควิโนโลนทำให้เกิดการเปลี่ยนแปลงทางเอพิเจเนติกในเซลล์ของสัตว์เลี้ยงลูกด้วยนม ผ่าน การจับกับ เหล็ก ซึ่งนำไปสู่ผลกระทบทางเอพิเจเนติกผ่านการยับยั้ง ไดออกซิเจเนสที่ขึ้นอยู่กับอัลฟา-คีโตกลูตาเรตซึ่งต้องการเหล็กเป็นโคแฟคเตอร์[ 206 ]
มีการใช้สารทางเภสัชวิทยาหลายชนิดในการผลิตเซลล์ต้นกำเนิดแบบเหนี่ยวนำให้เกิดความสามารถในการเปลี่ยนแปลงเป็นเซลล์ชนิดต่างๆ (iPSC) หรือรักษาสภาพฟีโนไทป์ของเซลล์ต้นกำเนิดจากตัวอ่อน (ESC) ผ่านวิธีการทางเอพิเจเนติกส์ เซลล์ต้นกำเนิดจากผู้ใหญ่ เช่น เซลล์ต้นกำเนิดจากไขกระดูก ยังแสดงให้เห็นถึงศักยภาพในการเปลี่ยนแปลงไปเป็นเซลล์หัวใจที่มีความสามารถในการทำงาน เมื่อได้รับการรักษาด้วยสารยับยั้งเอนไซม์เมทิลทรานสเฟอเรสของฮิสโตน G9a BIX01294 [ 207 ] [ 208 ]
ความยืดหยุ่นของเซลล์ ซึ่งเป็นการปรับตัวของเซลล์ต่อสิ่งเร้าโดยไม่ต้องเปลี่ยนแปลงรหัสพันธุกรรม จำเป็นต้องมีการเปลี่ยนแปลงทางเอพิเจเนติกส์ การเปลี่ยนแปลงเหล่านี้ได้รับการสังเกตในความยืดหยุ่นของเซลล์ในเซลล์มะเร็งระหว่างการเปลี่ยนผ่านจากเยื่อบุผิวเป็นเนื้อเยื่อเกี่ยวพัน[ 209 ]และในเซลล์ภูมิคุ้มกัน เช่น แมโครฟาจ[ 210 ]ที่น่าสนใจคือ การเปลี่ยนแปลงทางเมตาบอลิซึมเป็นพื้นฐานของการปรับตัวเหล่านี้ เนื่องจากเมตาบอไลต์ต่างๆ มีบทบาทสำคัญในเคมีของเครื่องหมายเอพิเจเนติกส์ ตัวอย่างเช่น อัลฟา-คีโตกลูตาเรต ซึ่งจำเป็นสำหรับการกำจัดหมู่เมทิลออกจากฮิสโตน และอะเซทิล-โคเอนไซม์เอ ซึ่งจำเป็นสำหรับการเติมหมู่อะเซทิลให้กับฮิสโตน
การแก้ไขเอพิเจโนม
การควบคุมการแสดงออกของยีนแบบเอพิเจเนติกที่สามารถเปลี่ยนแปลงหรือใช้ในการแก้ไขเอพิเจโนมได้แก่การดัดแปลง mRNA/lncRNA การดัดแปลง เมทิลเลชั่นของ DNAและการดัดแปลงฮิสโตน[ 211 ] [ 212 ] [ 213 ]
ตำแหน่ง CpG, SNP และลักษณะทางชีวภาพ
การเมทิลเลชันเป็นกลไกการควบคุมทางพันธุกรรมที่มีลักษณะเฉพาะอย่างกว้างขวางซึ่งสามารถกำหนดลักษณะทางชีวภาพได้ อย่างไรก็ตาม หลักฐานเชิงทดลองที่แข็งแกร่งเชื่อมโยงรูปแบบการเมทิลเลชันใน SNP เข้ากับคุณลักษณะเพิ่มเติมที่สำคัญสำหรับหลักการทางเอพิเจเนติกส์แบบคลาสสิกของการกระตุ้น/ยับยั้ง ข้อมูลปฏิสัมพันธ์ระดับโมเลกุล ซึ่งได้รับการสนับสนุนโดยการวิเคราะห์โคโลคัลไลเซชัน ระบุเส้นทางการควบคุมนิวเคลียร์หลายเส้นทางที่เชื่อมโยงความแปรผันของลำดับกับความผิดปกติในการเมทิลเลชันของ DNA และความแปรผันระดับโมเลกุลและฟีโนไทป์[ 214 ]
ตำแหน่งUBASH3B
UBASH3Bเข้ารหัสโปรตีนที่มีกิจกรรมไทโรซีนฟอสฟาเตส ซึ่งก่อนหน้านี้เชื่อมโยงกับเนื้องอกขั้นสูง[ 215 ] SNP rs7115089 ถูกระบุว่ามีอิทธิพลต่อการเมทิลเลชั่นของ DNA และการแสดงออกของตำแหน่งนี้ รวมถึงดัชนีมวลกาย (BMI) [ 214 ]ในความเป็นจริง SNP rs7115089 มีความสัมพันธ์อย่างมากกับ BMI [ 216 ]และกับตัวแปรทางพันธุกรรมที่เชื่อมโยงกับลักษณะทางหัวใจและหลอดเลือดและเมตาบอลิซึมอื่นๆ ใน GWAS [ 217 ] [ 218 ] [ 219 ]การศึกษาใหม่ๆ ชี้ให้เห็นว่าUBASH3Bเป็นตัวกลางที่มีศักยภาพของภาวะอ้วนและโรคหัวใจและหลอดเลือด[ 214 ]นอกจากนี้ แบบจำลองสัตว์แสดงให้เห็นว่า การแสดงออกของ UBASH3Bเป็นตัวบ่งชี้ของการจำกัดแคลอรี่ที่อาจขับเคลื่อนความไวต่อโรคอ้วนตามโปรแกรม และมีความสัมพันธ์กับการวัดภาวะอ้วนอื่นๆ ในเลือดส่วนปลายของมนุษย์[ 220 ]
ตำแหน่งNFKBIE
SNP rs730775 ตั้งอยู่ในอินตรอนแรกของNFKBIEและเป็นcis eQTL สำหรับNFKBIEในเลือดทั้งหมด[ 214 ]ตัวยับยั้งนิวเคลียร์แฟคเตอร์ (NF)-κB ε (NFKBIE) ยับยั้งกิจกรรมของ NF-κB1 โดยตรงและมีการแสดงออกร่วมกับ NF-κB1 อย่างมีนัยสำคัญ นอกจากนี้ยังเกี่ยวข้องกับโรคข้ออักเสบรูมาตอยด์[ 221 ]การวิเคราะห์โคโลคัลไลเซชันสนับสนุนว่าตัวแปรสำหรับไซต์ CpG ส่วนใหญ่ใน SNP rs730775 ทำให้เกิดความแปรผันทางพันธุกรรมที่ โลคั ส NFKBIEซึ่งอาจเชื่อมโยงกับโรคข้ออักเสบรูมาตอยด์ผ่าน การควบคุม แบบทรานส์แอคติ้งของการเมทิลเลชันของ DNA โดย NF-κB [ 214 ]
ตำแหน่งFADS1
เอนไซม์ Fatty acid desaturase 1 (FADS1) เป็นเอนไซม์สำคัญในการเผาผลาญกรดไขมัน[ 222 ]นอกจากนี้ rs174548 ใน ยีน FADS1 ยัง แสดงความสัมพันธ์ที่เพิ่มขึ้นกับการเมทิลเลชั่นของ DNA ในผู้ที่มีเซลล์ CD8+ T จำนวนมาก[ 214 ] SNP rs174548 มีความสัมพันธ์อย่างมากกับความเข้มข้นของกรดอะราคิโดนิกและเมตาบอไลต์อื่นๆ ในการเผาผลาญกรดไขมัน[ 223 ] [ 224 ]จำนวนอีโอซิโนฟิลในเลือด[ 225 ]และโรคอักเสบ เช่น โรคหอบหืด[ 226 ]ผลการวิเคราะห์ปฏิสัมพันธ์บ่งชี้ถึงความสัมพันธ์ระหว่าง rs174548 กับโรคหอบหืด ซึ่งให้ข้อมูลเชิงลึกใหม่เกี่ยวกับการเผาผลาญกรดไขมันในเซลล์ CD8+ T ที่มีฟีโนไทป์ภูมิคุ้มกัน[ 214 ]
วิทยาศาสตร์เทียม
เนื่องจากเอพิเจเนติกส์ยังอยู่ในช่วงเริ่มต้นของการพัฒนาในฐานะวิทยาศาสตร์และถูกห้อมล้อมด้วยความตื่นเต้นเร้าใจในสื่อสาธารณะเดวิด กอร์สกีและนักพันธุศาสตร์อดัม รัทเธอร์ฟอร์ดจึงแนะนำให้ระมัดระวังการแพร่กระจายของข้อสรุปที่ผิดพลาดและวิทยาศาสตร์เทียมโดย ผู้เขียน ยุคใหม่ที่เสนอแนะอย่างไม่มีมูลความจริงว่ายีนและสุขภาพของบุคคลสามารถถูกควบคุมโดยการควบคุมจิตใจการใช้คำศัพท์ทางวิทยาศาสตร์ในทางที่ผิดโดยผู้เขียนที่หลอกลวงได้ก่อให้เกิดข้อมูลที่ผิดพลาดในหมู่ประชาชนทั่วไป[ 2 ] [ 227 ]
ดูเพิ่มเติม
- ผลกระทบของบอลด์วิน
- เอพิเจเนติกส์เชิงพฤติกรรม
- ผลกระทบทางชีวภาพของรังสีต่ออีพิเจโนม
- เอพิเจเนติกส์เชิงคำนวณ
- บทบาทของการเปลี่ยนแปลงทางอีพีเจเนติกส์ต่อวิวัฒนาการ
- ฐานข้อมูล DAnCER (2010)
- เอพิเจเนซิส (ชีววิทยา)
- เอพิเจเนติกส์ในนิติวิทยาศาสตร์
- เอพิเจเนติกส์ของโรคภูมิคุ้มกันบกพร่อง
- การกำหนดลักษณะเอพิฟีโนไทป์
- การบำบัดด้วยเอพิเจเนติกส์
- เอพิเจเนติกส์ของโรคความเสื่อมของระบบประสาท
- เอพิเจเนติกส์ของอาการปวดเรื้อรัง
- พันธุศาสตร์
- ลามาร์คิสม์
- โภชนาการเอพิเจโนมิกส์
- การแปรผันของผลกระทบจากตำแหน่ง
- ลัทธิก่อนการก่อตัว
- เอพิไทป์ทางกายภาพ
- อาร์เรย์พันธุกรรมสังเคราะห์
- เอพิเจเนติกส์การนอนหลับ
- หน่วยความจำการถอดรหัส
- การถ่ายทอดทางพันธุกรรมแบบเอพิเจเนติกส์ข้ามรุ่น
อ่านเพิ่มเติม
- Haque FN, Gottesman II, Wong AH (พฤษภาคม 2552). "ไม่เหมือนกันอย่างแท้จริง: ความแตกต่างทางเอพิเจเนติกส์ในฝาแฝดโมโนไซโกติกและนัยสำคัญสำหรับการศึกษาฝาแฝดในจิตเวชศาสตร์" American Journal of Medical Genetics. Part C, Seminars in Medical Genetics . 151C (2): 136– 41. doi : 10.1002/ajmg.c.30206 . PMID 19378334 . S2CID 205327825 .
- "พันธุศาสตร์เหนือพันธุกรรมคืออะไร?"ศูนย์ควบคุมและป้องกันโรค 15 สิงหาคม 2565 สืบค้นเมื่อ 11 กันยายน 2566
ลิงก์ภายนอก
- "พันธุศาสตร์เหนือยีนและการถ่ายทอดทางพันธุกรรม" learn.genetics.utah.edu สืบค้นเมื่อ 17 เมษายน 2562
- โครงการเอพิเจโนมของมนุษย์ (HEP)
- เครือข่ายความเป็นเลิศด้านเอพิเจโนม (NoE)
- กลุ่มวิจัยด้านเอพิเจเนติกส์ สิ่งแวดล้อม และสุขภาพของแคนาดา (CEEHRC)
- เครือข่ายความเป็นเลิศด้านเอพิเจโนม (NoE) – เว็บไซต์สาธารณะระดับนานาชาติเก็บถาวรเมื่อวันที่ 28 พฤษภาคม 2010 ที่Wayback Machine
- "ดีเอ็นเอไม่ใช่โชคชะตา" – บทความหน้าปกนิตยสารDiscover
- "ผีในยีนของคุณ" , Horizon (2005), BBC
- บทความเกี่ยวกับเอพิเจเนติกส์ที่ Hopkins Medicine
- สู่แผนที่โลกของการเปลี่ยนแปลงทางพันธุกรรมระดับเอพิเจเนติกส์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เอพิเจเนติกส์
เอพิเจเนติกส์ คือการศึกษาการเปลี่ยนแปลงใน การแสดงออกของยีน ที่เกิดขึ้นโดยไม่เปลี่ยนแปลงลำดับ ดีเอ็นเอ [ 1 ] คำนำ หน้าภาษากรีก epi- (ἐπι- "เหนือกว่า, ภายนอก, รอบๆ") ใน...
คำจำกัดความ
คำว่า epigenesis มีความหมายทั่วไปว่า "การเจริญเติบโตพิเศษ" ซึ่งใช้ในภาษาอังกฤษมาตั้งแต่ศตวรรษที่ 17 [ 9 ] ในสิ่งพิมพ์ทางวิทยาศาสตร์ คำว่า epigenetics เริ่มปรากฏในทศวรรษที่ 1930 (ดูรูปทางด้านขวา) อย่างไรก็ตาม ความหมายร่วมสมัยของคำนี้เพิ่งปรากฏขึ้นในทศวรรษที่...
คลอง Waddington ในทศวรรษ 1940
สมมติฐานเกี่ยวกับการเปลี่ยนแปลงทางเอพิเจเนติกส์ที่มีผลต่อการแสดงออกของ โครโมโซม นั้นถูกเสนอโดยนักชีววิทยาชาวรัสเซีย Nikolai Koltsov [ 13 ] จาก ความหมายทั่วไปและคำคุณศัพท์ที่เกี่ยวข้องคือ เอพิเจเนติกส์ นักเอ็มบริโอวิทยาชาวอังกฤษ CH Waddington...
ร่วมสมัย
ในปี พ.ศ. 2533 Robin Holliday ได้นิยาม epigenetics ว่า "การศึกษาเกี่ยวกับกลไกการควบคุมกิจกรรมของยีนตามเวลาและพื้นที่ในระหว่างการพัฒนาของสิ่งมีชีวิตที่ซับซ้อน" [ 20 ]