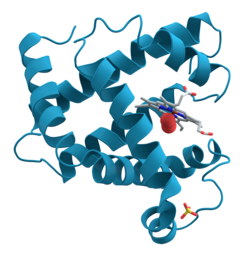
โปรตีนเป็นชีวโมเลกุลและโมเลกุล ขนาดใหญ่ ที่ประกอบด้วยกรดอะมิโน ที่เรียงตัวกันเป็นสายยาวหนึ่งสายหรือมากกว่า โปรตีนมีหน้าที่หลากหลายในสิ่งมีชีวิต รวมถึงการเร่งปฏิกิริยาเมแทบอลิซึม การจำลองดีเอ็นเอการตอบสนองต่อสิ่งกระตุ้นการสร้างโครงสร้างให้กับเซลล์และสิ่งมีชีวิตและการลำเลียงโมเลกุลจากที่หนึ่งไปยังอีกที่หนึ่ง โปรตีนแต่ละชนิดมีความแตกต่างกันโดยหลักๆ แล้วอยู่ที่ลำดับกรดอะมิโน ซึ่งถูกกำหนดโดยลำดับนิวคลีโอไทด์ของยีนและโดยปกติแล้วโปรตีนจะพับตัวเป็นโครงสร้างสามมิติ เฉพาะ ที่กำหนดกิจกรรมของโปรตีน
สายโซ่เชิงเส้นของกรดอะมิโนที่ตกค้างเรียกว่าพอลิเพปไทด์โปรตีนประกอบด้วยพอลิเพปไทด์ยาวอย่างน้อยหนึ่งพอลิเพปไทด์ พอลิเพปไทด์สั้นที่มีกรดอะมิโนน้อยกว่า 20–30 ตัว มักไม่ถือว่าเป็นโปรตีนและมักเรียกว่าเพ ปไทด์ กรดอะมิโนที่ตกค้างแต่ละตัวจะยึดติดกันด้วยพันธะเพปไทด์และกรดอะมิโนที่อยู่ติดกันลำดับของกรดอะมิโนที่ตกค้างในโปรตีนถูกกำหนดโดยลำดับของยีน ซึ่งถูกเข้ารหัสไว้ในรหัสพันธุกรรมโดยทั่วไป รหัสพันธุกรรมระบุกรดอะมิโนมาตรฐาน 20 ชนิด แต่ในสิ่งมีชีวิตบางชนิด รหัสพันธุกรรมอาจรวมถึงซีลีโนซิสเทอีนและในอาร์เคียบาง ชนิด อาจรวมถึง ไพร์โรไลซีนไม่นานหลังจากหรือแม้กระทั่งในระหว่างการสังเคราะห์ กรดอะมิโนที่ตกค้างในโปรตีนมักถูกดัดแปลงทางเคมีโดยการดัดแปลงหลังการแปลรหัสซึ่งจะเปลี่ยนแปลงคุณสมบัติทางกายภาพและทางเคมี การพับตัว ความเสถียร กิจกรรม และท้ายที่สุดคือหน้าที่ของโปรตีน โปรตีนบางชนิดมีหมู่ที่ไม่ใช่เพปไทด์เกาะอยู่ ซึ่งอาจเรียกว่าหมู่เทียมหรือโคแฟกเตอร์ โปรตีนสามารถทำงานร่วมกันเพื่อบรรลุหน้าที่เฉพาะอย่างหนึ่ง และมักจะรวมตัวกันเพื่อสร้างคอมเพล็กซ์โปรตีนที่ มีเสถียรภาพ
เมื่อโปรตีนถูกสร้างขึ้นแล้ว โปรตีนจะมีอายุอยู่เพียงช่วงระยะเวลาหนึ่งเท่านั้น จากนั้นจะถูกย่อยสลายและนำกลับมาใช้ใหม่โดยกลไกของเซลล์ผ่านกระบวนการหมุนเวียนโปรตีนอายุขัยของโปรตีนวัดจากครึ่งชีวิตและครอบคลุมช่วงกว้าง โปรตีนสามารถคงอยู่ได้นานหลายนาทีหรือหลายปี โดยมีอายุขัยเฉลี่ย 1-2 วันในเซลล์ของสัตว์เลี้ยงลูกด้วยนม โปรตีนที่ผิดปกติหรือมีการพับตัวผิดปกติจะเสื่อมสลายเร็วขึ้น เนื่องจากถูกกำหนดเป้าหมายเพื่อทำลาย หรือเนื่องจากไม่เสถียร
เช่นเดียวกับโมเลกุลขนาดใหญ่ทางชีววิทยาอื่นๆ เช่นโพลีแซ็กคาไรด์และกรดนิวคลีอิกโปรตีนเป็นส่วนประกอบสำคัญของสิ่งมีชีวิตและมีส่วนร่วมในแทบทุกกระบวนการภายในเซลล์โปรตีนหลายชนิดเป็นเอนไซม์ที่เร่งปฏิกิริยาทางชีวเคมีและมีความสำคัญต่อกระบวนการ เมแทบอลิ ซึม โปรตีนบางชนิดมีหน้าที่เชิงโครงสร้างหรือเชิงกล เช่นแอคตินและไมโอซินในกล้ามเนื้อ และโปรตีนโครงร่างของไซโตสเกเลตัน ที่รักษารูปร่างของเซลล์ โปรตีนชนิดอื่นๆ มีความสำคัญต่อการส่งสัญญาณของเซลล์ การตอบสนองทางภูมิคุ้มกันการยึดเกาะของเซลล์และวัฏจักรของเซลล์ในสัตว์ โปรตีนเป็นสิ่งจำเป็นในอาหารเพื่อให้กรดอะมิโนจำเป็นที่ไม่สามารถสังเคราะห์ได้การย่อยจะสลายโปรตีนเพื่อนำไปใช้ในกระบวนการเมแทบอลิซึม
ประวัติศาสตร์และนิรุกติศาสตร์
การค้นพบและการศึกษาในระยะเริ่มต้น
โปรตีนได้รับการศึกษาและรับรองมาตั้งแต่คริสต์ศตวรรษที่ 18 โดยAntoine Fourcroyและคนอื่นๆซึ่งมักเรียกโปรตีนเหล่านี้รวมกันว่า " อัลบูมิน " หรือ "สารอัลบูมินัส" ( Eiweisskörperในภาษาเยอรมัน) ตัวอย่างเช่นกลูเตน ถูกแยกออกจากข้าวสาลีเป็นครั้งแรกในงานวิจัยที่ตีพิมพ์เมื่อประมาณปี ค.ศ. 1747 และต่อมาพบว่ามีอยู่ในพืชหลายชนิด ในปี ค.ศ. 1789 Antoine Fourcroy ได้ระบุโปรตีนจากสัตว์สามชนิดที่แตกต่างกัน ได้แก่อัลบูมินไฟบรินและเจลาติน [ โปรตีนจากพืช (พืช) ที่ได้รับการศึกษาในช่วงปลายคริสต์ศตวรรษที่ 18 และต้นคริสต์ศตวรรษที่ 19 ได้แก่กลูเตนอั ล บูมินจากพืช กลี อะดิน และ เลกู มิน
โปรตีนได้รับการอธิบายครั้งแรกโดย Gerardus Johannes Mulderนักเคมีชาวดัตช์ และตั้งชื่อโดย Jöns Jacob Berzeliusนักเคมีชาวสวีเดนในปี ค.ศ. 1838 Mulder ได้ทำการวิเคราะห์องค์ประกอบ ของโปรตีนทั่วไปและพบว่าโปรตีนเกือบทั้งหมดมี สูตรเชิงประจักษ์เดียวกันคือ C 400 H 620 N 100 O 120 P 1 S 1 เขาสรุปอย่างผิดพลาดว่าโปรตีนอาจประกอบด้วยโมเลกุลชนิดเดียว (ขนาดใหญ่มาก) คำว่า "โปรตีน" เพื่ออธิบายโมเลกุลเหล่านี้ถูกเสนอโดย Berzelius ผู้ร่วมงานของ Mulder; โปรตีนมาจากคำภาษากรีกπρώτειος ( proteios ) ซึ่งแปลว่า "ปฐมภูมิ" "นำหน้า" หรือ "ยืนอยู่ข้างหน้า" [ + -inมัลเดอร์ได้ระบุผลิตภัณฑ์จากการย่อยสลายโปรตีน เช่น กรดอะมิโน ลิวซีนซึ่งเขาพบว่ามีน้ำหนักโมเลกุล (เกือบถูกต้อง)131 ดา .
นักวิทยาศาสตร์ด้านโภชนาการยุคแรกๆ เช่นคาร์ล ฟอน วอยต์ ชาวเยอรมัน เชื่อว่าโปรตีนเป็นสารอาหารที่สำคัญที่สุดในการรักษาโครงสร้างของร่างกาย เพราะโดยทั่วไปเชื่อกันว่า "เนื้อสร้างเนื้อ" ประมาณปี ค.ศ. 1862 คาร์ล ไฮน์ริช ริตเฮา เซน ได้แยกกรดอะมิโนกลูตามิก [ โทมัส เบอร์ ออสบอร์นได้รวบรวมบทวิจารณ์โดยละเอียดเกี่ยวกับโปรตีนจากพืชที่สถานีทดลองเกษตรคอนเนตทิคัต ออสบอร์น ร่วมกับลาฟาแยตต์ เมนเดล ได้ค้นพบ กรดอะมิโนที่จำเป็นทางโภชนาการหลายชนิดในการทดลองให้อาหารหนูทดลองในห้องปฏิบัติการอาหารที่ขาดกรดอะมิโนจำเป็นจะทำให้หนูเจริญเติบโตช้าลง ซึ่งสอดคล้องกับ กฎของลีบิกเกี่ยวกับค่า ต่ำสุดกรดอะมิโนจำเป็นชนิดสุดท้ายที่ถูกค้นพบ คือทรีโอนีนถูกระบุโดยวิลเลียม คัมมิง โรส [
ความยากลำบากในการทำให้โปรตีนบริสุทธิ์เป็นอุปสรรคต่อการทำงานของนักชีวเคมีโปรตีนในยุคแรกๆ โปรตีนสามารถหาได้จากเลือด ไข่ขาว และเคราติน ในปริมาณมาก แต่โปรตีนแต่ละชนิดยังไม่สามารถหาได้ ในช่วงทศวรรษ 1950 บริษัท Armour Hot Dog ได้ ทำให้เอนไซม์ ไรโบนิวคลีเอส เอของตับอ่อนวัวบริสุทธิ์ 1 กิโลกรัมและเปิดให้นักวิทยาศาสตร์เข้าถึงได้ฟรี การกระทำนี้ช่วยให้ไรโบนิวคลีเอส เอ กลายเป็นเป้าหมายหลักสำหรับการศึกษาทางชีวเคมีในทศวรรษต่อมา
โพลีเปปไทด์

ความเข้าใจเกี่ยวกับโปรตีนในฐานะโพลีเปปไทด์หรือสายของกรดอะมิโนเกิดขึ้นจากงานของFranz HofmeisterและHermann Emil Fischerในปี 1902 บทบาทสำคัญของโปรตีนในฐานะเอนไซม์ในสิ่งมีชีวิตที่เร่งปฏิกิริยายังไม่เป็นที่เข้าใจอย่างสมบูรณ์จนกระทั่งปี 1926 เมื่อJames B. Sumnerแสดงให้เห็นว่าเอนไซม์ยูรีเอสเป็นโปรตีน
Linus Paulingได้รับการยกย่องว่าเป็นผู้ทำนายโครงสร้างรอง ของโปรตีนปกติได้สำเร็จ โดยอาศัยพันธะไฮโดรเจนซึ่งเป็นแนวคิดที่เสนอครั้งแรกโดยWilliam Astburyในปี 1933 งานยุคหลังของWalter Kauzmannเกี่ยวกับการเปลี่ยนแปลงสภาพ [ ซึ่งอิงจากการศึกษาครั้งก่อนของKaj Linderstrøm-Lang บางส่วน [ มีส่วนสนับสนุนความเข้าใจเกี่ยวกับการพับตัวของโปรตีนและโครงสร้างที่เกิดจากปฏิสัมพันธ์ที่ไม่ชอบน้ำ [
โปรตีนตัวแรกที่มีลำดับกรดอะมิโนโซ่คืออินซูลินโดยเฟรเดอริก แซงเกอร์ในปี 1949 แซงเกอร์ได้กำหนดลำดับกรดอะมิโนของอินซูลินได้อย่างถูกต้อง จึงแสดงให้เห็นอย่างชัดเจนว่าโปรตีนประกอบด้วยพอลิเมอร์เชิงเส้นของกรดอะมิโน ไม่ใช่โซ่กิ่งคอลลอยด์หรือไซโคล [ เขาได้รับรางวัลโนเบลจากความสำเร็จนี้ในปี 1958 การศึกษาของคริสเตียน อันฟินเซน เกี่ยวกับกระบวนการ พับออกซิเดชันของไรโบนิวคลีเอสเอ ซึ่งเขาได้รับรางวัลโนเบลในปี 1972 ทำให้สมมติฐานทางอุณหพลศาสตร์ของการพับโปรตีนแข็งแกร่งขึ้น ซึ่งตามทฤษฎีนี้ รูปแบบการพับของโปรตีนแสดงถึงพลังงานอิสระขั้นต่ำ
โครงสร้าง

ด้วยการพัฒนาของผลึกศาสตร์รังสีเอกซ์ทำให้สามารถระบุโครงสร้างโปรตีนและลำดับเบสได้โครงสร้างโปรตีนแรกๆที่ถูกค้นพบคือฮีโมโกลบินโดยMax PerutzและไมโอโกลบินโดยJohn Kendrewในปี 1958 การใช้คอมพิวเตอร์และพลังการประมวลผลที่เพิ่มขึ้นได้สนับสนุนการจัดลำดับโปรตีนที่ซับซ้อน ในปี 1999 Roger Kornbergได้จัดลำดับโครงสร้างที่ซับซ้อนอย่างยิ่งของRNA polymeraseโดยใช้รังสีเอกซ์ความเข้มสูงจากซินโครตรอน [
นับแต่นั้นมา ได้มีการพัฒนากล้องจุลทรรศน์อิเล็กตรอนแบบแช่แข็ง (cryo-EM) ของโครงสร้างโมเลกุล ขนาดใหญ่ ใช้ลำแสงอิเล็กตรอนแทนรังสีเอกซ์ วิธีนี้สร้างความเสียหายต่อตัวอย่างน้อยกว่า ทำให้นักวิทยาศาสตร์ได้รับข้อมูลมากขึ้นและวิเคราะห์โครงสร้างขนาดใหญ่ได้การทำนายโครงสร้างโปรตีนด้วยคอมพิวเตอร์ของโดเมนโครงสร้าง โปรตีนขนาดเล็ก ช่วยให้นักวิจัยสามารถเข้าถึงความละเอียดของโครงสร้างโปรตีนในระดับอะตอมได้ ณ เดือนเมษายน พ.ศ. 2567 ธนาคารข้อมูลโปรตีนมีโครงสร้างโปรตีนจากรังสีเอกซ์ 181,018 โครงสร้าง, EM 19,809 โครงสร้าง และNMR 12,697 โครงสร้าง
การจำแนกประเภท
โปรตีนถูกจำแนกประเภทโดยลำดับและโครงสร้างเป็นหลัก แม้ว่าโดยทั่วไปจะใช้การจำแนกประเภทอื่นๆ ก็ตาม โดยเฉพาะอย่างยิ่งสำหรับเอนไซม์ ระบบเลข EC จะให้รูปแบบการจำแนกประเภทเชิงหน้าที่ในทำนองเดียวกันออนโทโลยีของยีนจำแนกทั้งยีนและโปรตีนตามหน้าที่ทางชีวภาพและชีวเคมี และตามตำแหน่งภายในเซลล์
ความคล้ายคลึงกันของลำดับเบสถูกนำมาใช้เพื่อจำแนกโปรตีนทั้งในแง่ของความคล้ายคลึงกันทางวิวัฒนาการและหน้าที่ ซึ่งอาจใช้ทั้งโปรตีนทั้งหมดหรือโดเมนโปรตีนโดยเฉพาะอย่างยิ่งในโปรตีนหลายโดเมนโดเมนโปรตีนช่วยให้สามารถจำแนกโปรตีนได้โดยการรวมกันของลำดับ โครงสร้าง และหน้าที่ และสามารถนำมารวมกันได้หลายวิธี ในการศึกษาเบื้องต้นเกี่ยวกับโปรตีน 170,000 ชนิด พบว่าประมาณสองในสามถูกกำหนดให้มีโดเมนอย่างน้อยหนึ่งโดเมน โดยโปรตีนที่มีขนาดใหญ่กว่าจะมีโดเมนมากกว่า (เช่น โปรตีนที่มีกรดอะมิโน มากกว่า 600 ตัว จะมีโดเมนมากกว่า 5 โดเมนโดยเฉลี่ย)
ชีวเคมี


โปรตีนส่วนใหญ่ประกอบด้วยพอลิเมอร์ เชิงเส้นที่สร้างขึ้นจากกรดอะมิโน L -αมากถึง 20 ตัว กรดอะมิ โนโปรตีนเจนิกทั้งหมดมีโครงสร้างร่วมกัน โดยคาร์บอน αจะจับกับหมู่อะมิโน หมู่ คาร์บอกซิลและโซ่ข้าง ที่แปรผันได้ มี เพียงโพรลีน เท่านั้น ที่แตกต่างจากโครงสร้างพื้นฐานนี้ เนื่องจากโซ่ข้างเป็นวัฏจักร จับกับหมู่อะมิโน ซึ่งเป็นข้อจำกัดความยืดหยุ่นของโซ่โปรตีนโซ่ข้างของกรดอะมิโนมาตรฐานมีโครงสร้างและคุณสมบัติทางเคมีที่หลากหลาย และผลรวมของกรดอะมิโนทั้งหมดเป็นตัวกำหนดโครงสร้างสามมิติและปฏิกิริยาเคมี
กรดอะมิโนในสายพอลิเพปไทด์เชื่อมต่อกันด้วยพันธะเปปไทด์ระหว่างหมู่อะมิโนและหมู่คาร์บอกซิล กรดอะมิโนแต่ละตัวในสายเรียกว่าสารตกค้างและอะตอมของคาร์บอน ไนโตรเจน และออกซิเจนที่เชื่อมต่อกันเรียกว่าสายหลักหรือ แกนหลัก ของโปรตีนพันธะเปปไทด์มี รูปแบบ เรโซแนนซ์ สอง รูปแบบที่ทำให้ แกนหลักของโปรตีนมีลักษณะ พันธะคู่คาร์บอนแอลฟาจะอยู่ในระนาบ เดียว กับไนโตรเจนและหมู่คาร์บอนิล (C=O) มุมไดฮีดรัลอีกสองมุมในพันธะเปปไทด์กำหนดรูปร่างเฉพาะที่ของแกนหลักของโปรตีน ผลที่ตามมาอย่างหนึ่งของลักษณะพันธะคู่ NC(O) คือโปรตีนมีความแข็งเล็กน้อยสายพอลิเพปไทด์สิ้นสุดด้วยหมู่อะมิโนอิสระที่เรียกว่าปลาย Nหรือปลายอะมิโนและหมู่คาร์บอกซิลอิสระที่เรียกว่าปลาย Cหรือปลายคาร์บอกซีตามธรรมเนียม ลำดับเปปไทด์จะถูกเขียนจากปลาย N ถึงปลาย C ซึ่งสัมพันธ์กับลำดับที่โปรตีนถูกสังเคราะห์โดยไรโบโซม [
คำว่าโปรตีนโพลีเปปไทด์และเปปไทด์ค่อนข้างคลุมเครือและอาจมีความหมายทับซ้อนกัน โดย ทั่วไปแล้ว โปรตีนมักใช้เรียกโมเลกุลชีวภาพที่สมบูรณ์ในโครงสร้าง ที่เสถียร ในขณะที่เปปไทด์มักถูกสงวนไว้สำหรับโอลิโกเมอร์กรดอะมิโนสั้นๆ ซึ่งมักไม่มีโครงสร้างสามมิติที่เสถียร แต่ขอบเขตระหว่างทั้งสองยังไม่ชัดเจนนัก และมักอยู่ที่ประมาณ 20–30 เรซิดิว
โปรตีนสามารถโต้ตอบกับโมเลกุลและไอออนหลายประเภท รวมถึงกับโปรตีนชนิดอื่นกับไขมันกับคาร์โบไฮเดรตและกับดีเอ็นเอ [
ความอุดมสมบูรณ์ในเซลล์
เซลล์ แบคทีเรียทั่วไปเช่นE. coliและStaphylococcus aureusคาดว่ามีโปรตีนประมาณ 2 ล้านตัว แบคทีเรียขนาดเล็กกว่า เช่นMycoplasmaหรือSpirocheteมีโมเลกุลน้อยกว่า ประมาณ 50,000 ถึง 1 ล้านตัว ในทางตรงกันข้าม เซลล์ ยูคาริโอตมีขนาดใหญ่กว่า จึงมีโปรตีนมากกว่ามาก ตัวอย่างเช่น เซลล์ ยีสต์มีโปรตีนประมาณ 50 ล้านตัว และ เซลล์ มนุษย์ประมาณ 1 ถึง 3 พันล้านตัวความเข้มข้นของสำเนาโปรตีนแต่ละชุดมีตั้งแต่ไม่กี่โมเลกุลต่อเซลล์ไปจนถึง 20 ล้านตัวยีนที่เข้ารหัสโปรตีนไม่ได้แสดงออกทั้งหมดในเซลล์ส่วนใหญ่ และจำนวนยีนเหล่านี้ขึ้นอยู่กับชนิดของเซลล์และสิ่งกระตุ้นภายนอก ตัวอย่างเช่น ในโปรตีนประมาณ 20,000 ตัวที่เข้ารหัสโดยจีโนมมนุษย์ มีเพียง 6,000 ตัวเท่านั้นที่ตรวจพบในเซลล์lymphoblastoid โปรตีนที่มีมากที่สุดในธรรมชาติเชื่อกันว่าคือRuBisCOซึ่งเป็นเอนไซม์ที่เร่งปฏิกิริยาการรวมตัวของคาร์บอนไดออกไซด์เข้ากับสารอินทรีย์ในกระบวนการสังเคราะห์แสงพืชอาจมีเอนไซม์นี้มากถึง 1% โดยน้ำหนัก
สังเคราะห์
การสังเคราะห์ทางชีวภาพ
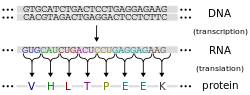
โปรตีนประกอบขึ้นจากกรดอะมิโนโดยใช้ข้อมูลที่เข้ารหัสในยีน โปรตีนแต่ละชนิดมีลำดับกรดอะมิโนเฉพาะของตัวเอง ซึ่งกำหนดโดย ลำดับ นิวคลีโอไทด์ของยีนที่เข้ารหัสโปรตีนนี้รหัสพันธุกรรมคือชุดของชุดนิวคลีโอไทด์สามชุดที่เรียกว่าโคดอนและแต่ละชุดที่มีสามนิวคลีโอไทด์จะกำหนดกรดอะมิโน ตัวอย่างเช่น AUG ( adenine – uracil – guanine ) เป็นรหัสของเมไทโอนีนเนื่องจากDNAมีนิวคลีโอไทด์สี่ตัว จำนวนโคดอนที่เป็นไปได้ทั้งหมดคือ 64 ตัว ดังนั้นจึงมีความซ้ำซ้อนในรหัสพันธุกรรม โดยกรดอะมิโนบางชนิดถูกกำหนดโดยโคดอนมากกว่าหนึ่งตัว ยีนที่เข้ารหัสใน DNA จะ ถูกถอดรหัสเป็น pre- messenger RNA (mRNA) ก่อนโดยโปรตีน เช่นRNA polymerase สิ่งมีชีวิตส่วนใหญ่จะประมวลผล pre-mRNA (ทรานสคริปต์หลัก ) โดยใช้รูปแบบต่างๆ ของการดัดแปลงหลังการถอดรหัสเพื่อสร้าง mRNA ที่โตเต็มที่ ซึ่งจะถูกใช้เป็นแม่แบบสำหรับการสังเคราะห์โปรตีนโดยไรโบโซมในโปรคาริโอต mRNA อาจถูกนำมาใช้ทันทีที่สร้างขึ้น หรือถูกผูกมัดโดยไรโบโซมหลังจากเคลื่อนออกจากนิวคลีโอตในทางตรงกันข้ามยูคาริโอตจะสร้าง mRNA ในนิวเคลียสของเซลล์แล้วจึงเคลื่อนย้ายผ่านเยื่อหุ้มนิวเคลียสเข้าสู่ไซโทพลาสซึมซึ่งเป็นที่ที่การสังเคราะห์โปรตีนจะเกิดขึ้น อัตราการสังเคราะห์โปรตีนในโปรคาริโอตสูงกว่ายูคาริโอต และสามารถมีกรดอะมิโนได้ถึง 20 ตัวต่อวินาที
กระบวนการสังเคราะห์โปรตีนจากแม่แบบ mRNA เรียกว่าการแปลรหัส mRNA จะถูกโหลดลงบนไรโบโซมและอ่านนิวคลีโอไทด์ครั้งละสามตัวโดยการจับคู่โคดอนแต่ละตัวกับ แอ นติโคดอน ที่จับคู่กับ เบส ซึ่งอยู่บน โมเลกุล RNA ถ่ายโอนซึ่งบรรจุกรดอะมิโนที่สอดคล้องกับโคดอนที่มันจดจำ เอนไซม์อะมิโนแอซิล ทีอาร์เอ็นเอ ซินเทส จะ "เติม" กรดอะมิโนที่ถูกต้องให้กับโมเลกุล tRNA พอลิเพปไทด์ที่กำลังเติบโตมักเรียกว่าnascent chainโปรตีนจะถูกสังเคราะห์ทางชีวภาพจากปลาย Nไปยังปลาย Cเสมอ
ขนาดของโปรตีนที่สังเคราะห์ขึ้นสามารถวัดได้จากจำนวนกรดอะมิโนที่ประกอบขึ้น และมวลโมเลกุล รวม ซึ่งปกติจะรายงานด้วยหน่วยดาลตัน (Da) หรือหน่วยอนุพันธ์กิโลดาลตัน (kDa) ขนาดเฉลี่ยของโปรตีนจะเพิ่มขึ้นจากอาร์เคียไปเป็นแบคทีเรียไปเป็นยูคาริโอต (283, 311, 438 เรซิดิว และ 31, 34, 49 kDa ตามลำดับ) เนื่องจากจำนวนโดเมนโปรตีนที่ประกอบเป็นโปรตีนในสิ่งมีชีวิตชั้นสูง มีจำนวนมากกว่า ตัวอย่างเช่น โปรตีน ยีสต์มีความยาวโดยเฉลี่ย 466 กรดอะมิโน และมีมวล 53 kDa โปรตีนที่ใหญ่ที่สุดที่รู้จักคือไททินซึ่งเป็นส่วนประกอบของซาร์โคเมียร์ของกล้ามเนื้อ มีมวลโมเลกุลเกือบ3000 kDaและความยาวรวมเกือบกรดอะมิโน27,000 ตัว[
การสังเคราะห์ทางเคมี

โปรตีนสั้นสามารถสังเคราะห์ทางเคมีได้โดย วิธี การสังเคราะห์เปปไทด์ หลายแบบ ซึ่งอาศัย เทคนิค การสังเคราะห์สารอินทรีย์เช่นการผูกทางเคมีเพื่อผลิตเปปไทด์ที่ให้ผลผลิตสูงการสังเคราะห์ทางเคมีช่วยให้สามารถนำกรดอะมิโนที่ไม่ใช่ธรรมชาติเข้าสู่สายพอลิเพปไทด์ได้ เช่น การยึด โพ รบฟลูออเรสเซนต์เข้ากับสายข้างของกรดอะมิโนวิธีการเหล่านี้มีประโยชน์ในห้องปฏิบัติการชีวเคมีและชีววิทยาของเซลล์แม้ว่าโดยทั่วไปจะไม่เหมาะสำหรับการใช้งานเชิงพาณิชย์ การสังเคราะห์ทางเคมีไม่มีประสิทธิภาพสำหรับพอลิเพปไทด์ที่มีกรดอะมิโนมากกว่า 300 ตัว และโปรตีนที่สังเคราะห์อาจไม่สามารถมีโครงสร้างตติยภูมิ ดั้งเดิมได้ วิธีการสังเคราะห์ทางเคมีส่วนใหญ่ดำเนินจากปลาย C ไปยังปลาย N ซึ่งตรงข้ามกับปฏิกิริยาทางชีวภาพ
โครงสร้าง


โปรตีนส่วนใหญ่พับตัวเป็นโครงสร้างสามมิติที่มีลักษณะเฉพาะ รูปร่างที่โปรตีนพับตัวตามธรรมชาติเรียกว่าโครงสร้างดั้งเดิมแม้ว่าโปรตีนหลายชนิดสามารถพับตัวได้โดยไม่ต้องอาศัยความช่วยเหลือ เพียงแค่ผ่านคุณสมบัติทางเคมีของกรดอะมิโน โปรตีนบางชนิดก็ต้องการความช่วยเหลือจากchaperone โมเลกุล เพื่อพับตัวเป็นโครงสร้างดั้งเดิมนักชีวเคมีมักอ้างถึงโครงสร้างโปรตีนสี่ประการที่แตกต่างกัน:
- โครงสร้างหลัก :ลำดับกรดอะมิโนโปรตีนเป็นโพลีเอไมด์
- โครงสร้างทุติยภูมิ : โครงสร้างเฉพาะที่ที่ซ้ำกันอย่างสม่ำเสมอและเสถียรด้วยพันธะไฮโดรเจนตัวอย่างที่พบบ่อยที่สุด ได้แก่ α-helix , β-sheetและ turnเนื่องจากโครงสร้างทุติยภูมิเป็นโครงสร้างเฉพาะที่ จึงสามารถมีบริเวณที่มีโครงสร้างทุติยภูมิที่แตกต่างกันหลายส่วนอยู่ในโมเลกุลโปรตีนเดียวกันได้
- โครงสร้างตติยภูมิ : รูปร่างโดยรวมของโมเลกุลโปรตีนเดี่ยว ความสัมพันธ์เชิงพื้นที่ของโครงสร้างทุติยภูมิต่อกัน โครงสร้างตติยภูมิโดยทั่วไปจะคงตัวโดยปฏิกิริยานอกท้องถิ่น ซึ่งส่วนใหญ่มักเกิดจากการก่อตัวของแกนที่ไม่ชอบน้ำแต่ยังรวมถึงสะพานเกลือพันธะไฮโดรเจน พันธะไดซัลไฟด์และแม้แต่การดัดแปลงหลังการแปลรหัส คำว่า "โครงสร้างตติยภูมิ" มักใช้เป็นคำพ้องความหมายกับคำว่า "พับ " โครงสร้างตติยภูมิเป็นส่วนที่ควบคุมการทำงานพื้นฐานของโปรตีน
- โครงสร้างควอเทอร์นารี : โครงสร้างที่ก่อตัวจากโมเลกุลโปรตีนหลายโมเลกุล (โซ่โพลีเปปไทด์) ซึ่งในบริบทนี้ มักเรียกว่าซับยูนิตโปรตีนซึ่งทำหน้าที่เป็นคอมเพล็กซ์โปรตีน เดี่ยว
- โครงสร้างควินารี : ลักษณะของพื้นผิวโปรตีนที่ทำหน้าที่จัดระเบียบภายในเซลล์ที่หนาแน่น โครงสร้างควินารีขึ้นอยู่กับปฏิสัมพันธ์ของโมเลกุลขนาดใหญ่ที่เกิดขึ้นภายในเซลล์ที่มีชีวิต ซึ่งเกิดขึ้นชั่วคราวแต่มีความสำคัญ
โปรตีนไม่ใช่โมเลกุลที่แข็งกระด้างอย่างสมบูรณ์ นอกจากระดับโครงสร้างเหล่านี้แล้ว โปรตีนอาจเปลี่ยนแปลงไปมาระหว่างโครงสร้างที่เกี่ยวข้องกันหลายโครงสร้างในขณะที่มันทำหน้าที่ของมัน ในบริบทของการจัดเรียงตัวใหม่เหล่านี้ โครงสร้างตติยภูมิหรือควอเทอร์นารีเหล่านี้มักเรียกว่า " คอนฟอร์เมชัน " และการเปลี่ยนผ่านระหว่างโครงสร้างเหล่านี้เรียกว่าการเปลี่ยนแปลงคอนฟอร์เมชันการเปลี่ยนแปลงดังกล่าวมักเกิดจากการจับกันของ โมเลกุล สารตั้งต้น กับ บริเวณแอคทีฟของเอนไซม์หรือบริเวณทางกายภาพของโปรตีนที่มีส่วนร่วมในการเร่งปฏิกิริยาทางเคมี ในสารละลาย โครงสร้างโปรตีนจะเปลี่ยนแปลงไปเนื่องจากการสั่นสะเทือนทางความร้อนและการชนกับโมเลกุลอื่นๆ

โปรตีนสามารถแบ่งอย่างไม่เป็นทางการออกเป็นสามกลุ่มหลัก ซึ่งสัมพันธ์กับโครงสร้างตติยภูมิทั่วไป ได้แก่โปรตีนทรงกลมโปรตีนเส้นใยและโปรตีนเยื่อหุ้มเซลล์โปรตีนทรงกลมเกือบทั้งหมดละลายน้ำได้และหลายชนิดเป็นเอนไซม์ โปรตีนเส้นใยมักมีโครงสร้าง เช่นคอลลาเจนซึ่งเป็นองค์ประกอบหลักของเนื้อเยื่อเกี่ยวพัน หรือเคราตินซึ่งเป็นองค์ประกอบโปรตีนของเส้นผมและเล็บ โปรตีนเยื่อหุ้มเซลล์มักทำหน้าที่เป็นตัวรับหรือเป็นช่องทางให้โมเลกุลที่มีขั้วหรือมีประจุผ่าน เยื่อ หุ้มเซลล์
กรณีพิเศษของพันธะไฮโดรเจนภายในโมเลกุลภายในโปรตีนซึ่งป้องกันการโจมตีของน้ำได้ไม่ดีนัก จึงส่งเสริมการคายน้ำ ของตัวเอง เรียกว่าดีไฮดรอน [
โดเมนโปรตีน
โปรตีนหลายชนิดประกอบด้วยโดเมนโปรตีน หลายโดเมน กล่าว คือ ส่วนของโปรตีนที่พับเป็นหน่วยโครงสร้างที่แตกต่างกันโดเมนมักมีหน้าที่เฉพาะ เช่นกิจกรรมของเอนไซม์ (เช่น ไคเนส ) หรือทำหน้าที่เป็นโมดูลการจับ

รูปแบบลำดับ
ลำดับกรดอะมิโนสั้น ๆ ภายในโปรตีนมักทำหน้าที่เป็นจุดจดจำโปรตีนอื่น ๆตัวอย่างเช่นโดเมน SH3มักจะจับกับโมทิฟ PxxP สั้น ๆ (เช่นโพรลีน 2 ตัว [P] คั่นด้วย กรดอะมิโนที่ไม่ระบุสองตัว[x] แม้ว่ากรดอะมิโนที่อยู่รอบ ๆ อาจกำหนดความจำเพาะในการจับที่แน่นอน) โมทิฟเหล่านี้จำนวนมากถูกรวบรวมไว้ใน ฐานข้อมูล Eukaryotic Linear Motif (ELM)
ฟังก์ชันของเซลล์
โปรตีนเป็นสารออกฤทธิ์หลักภายในเซลล์ กล่าวกันว่าทำหน้าที่ตามที่กำหนดไว้ในข้อมูลที่เข้ารหัสในยีนยกเว้นRNA บางชนิด โมเลกุลชีวภาพอื่นๆ ส่วนใหญ่มักเป็นองค์ประกอบที่ค่อนข้างเฉื่อยซึ่งโปรตีนทำหน้าที่ โปรตีนมีน้ำหนักแห้งครึ่งหนึ่งของ เซลล์ Escherichia coliในขณะที่โมเลกุลขนาดใหญ่อื่นๆ เช่น DNA และ RNA มีเพียง 3% และ 20% ตามลำดับกลุ่มโปรตีนที่แสดงออกในเซลล์หรือชนิดของเซลล์เฉพาะเรียกว่าโปรตีโอม [

ลักษณะสำคัญของโปรตีนที่ทำหน้าที่หลากหลาย คือ ความสามารถในการจับโมเลกุลอื่นๆ ได้อย่างแน่นหนาและเฉพาะเจาะจง บริเวณของโปรตีนที่ทำหน้าที่จับโมเลกุลอื่นเรียกว่าบริเวณจับและมักเป็นแอ่งหรือ "ช่อง" บนพื้นผิวโมเลกุล ความสามารถในการจับนี้เกิดจากโครงสร้างตติยภูมิของโปรตีน ซึ่งกำหนดช่องของบริเวณจับ และจากคุณสมบัติทางเคมีของโซ่ข้างเคียงของกรดอะมิโนโดยรอบ การจับของโปรตีนสามารถแน่นหนาและจำเพาะเจาะจงอย่างยิ่ง ตัวอย่างเช่น โปรตีนที่ยับยั้งไรโบนิวคลี เอส จะจับกับแองจิโอเจนิน ของมนุษย์โดยมีค่า คงที่การแยก ตัวแบบซับเฟมโตโมลาร์ (< 10 โมลาร์) แต่จะไม่จับกับโฮโมล็อกออน โคเนสของสัตว์ ครึ่งบกครึ่งน้ำ เลย (> 1 โมลาร์) การเปลี่ยนแปลงทางเคมีเล็กน้อยมาก เช่น การเพิ่มหมู่เมทิลเดี่ยวเข้าไปในคู่จับ บางครั้งอาจเพียงพอที่จะกำจัดการจับได้เกือบหมด ตัวอย่างเช่นอะมิโนแอซิล tRNA ซินเทสที่จำเพาะต่อกรดอะมิโนวาลีนจะแยกแยะกับกลุ่มข้างเคียงที่คล้ายกันมากของกรดอะมิโนไอโซลิวซีน [
โปรตีนสามารถจับกับโปรตีนชนิดอื่นได้เช่นเดียวกับ ซับสเตรต โมเลกุลขนาดเล็กเมื่อโปรตีนจับกับสำเนาอื่นของโมเลกุลเดียวกันโดยเฉพาะ โปรตีนเหล่านั้นสามารถโอลิโก เมอไรเซชัน เพื่อสร้างเส้นใย ซึ่งกระบวนการนี้มักเกิดขึ้นในโปรตีนโครงสร้างที่ประกอบด้วยโมโนเมอร์ทรงกลมที่รวมตัวกันเองเพื่อสร้างเส้นใยแข็ง ปฏิกิริยาระหว่าง โปรตีนกับโปรตีนควบคุมการทำงานของเอนไซม์ ควบคุมการดำเนินไปตลอดวัฏจักรเซลล์และทำให้เกิดการรวมตัวของคอมเพล็กซ์โปรตีน ขนาดใหญ่ ซึ่งทำปฏิกิริยาที่เกี่ยวข้องอย่างใกล้ชิดมากมายและมีหน้าที่ทางชีววิทยาร่วมกัน โปรตีนสามารถจับกับหรือรวมเข้ากับเยื่อหุ้มเซลล์ได้ ความสามารถของคู่จับในการเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างในโปรตีนทำให้สามารถสร้างเครือข่ายการส่งสัญญาณ ที่ซับซ้อนมหาศาลได้ เนื่องจากปฏิกิริยาระหว่างโปรตีนสามารถย้อนกลับได้และขึ้นอยู่กับความพร้อมของกลุ่มโปรตีนคู่จับที่แตกต่างกันอย่างมากในการสร้างกลุ่มโปรตีนที่สามารถทำหน้าที่แยกกัน การศึกษาปฏิกิริยาระหว่างโปรตีนเฉพาะจึงเป็นกุญแจสำคัญในการทำความเข้าใจแง่มุมสำคัญของการทำงานของเซลล์ และท้ายที่สุดคือคุณสมบัติที่จำแนกเซลล์แต่ละชนิด
เอนไซม์
บทบาทที่รู้จักกันดีที่สุดของโปรตีนในเซลล์คือเอนไซม์ซึ่ง ทำหน้าที่ เร่งปฏิกิริยาเคมี เอนไซม์มักจะมีความจำเพาะสูงและเร่งปฏิกิริยาเคมีเพียงหนึ่งหรือสองปฏิกิริยาเท่านั้น เอนไซม์ทำปฏิกิริยาส่วนใหญ่ที่เกี่ยวข้องกับกระบวนการเมแทบอลิซึมรวมถึงควบคุมดีเอ็นเอในกระบวนการต่างๆ เช่นการจำลองดีเอ็นเอการซ่อมแซมดีเอ็นเอและการถอดรหัสเอนไซม์บางชนิดทำปฏิกิริยากับโปรตีนชนิดอื่นเพื่อเพิ่มหรือกำจัดหมู่เคมีในกระบวนการที่เรียกว่าการดัดแปลงหลังการแปล เป็นที่ทราบกันว่ามีปฏิกิริยาประมาณ 4,000 ปฏิกิริยาที่ถูกเร่งปฏิกิริยาโดยเอนไซม์อัตราเร่งที่เกิดจากปฏิกิริยาเร่งปฏิกิริยาด้วยเอนไซม์มักจะสูงมาก โดยเพิ่มขึ้นถึง 10-17 เมื่อเทียบกับปฏิกิริยาที่ไม่ได้เร่งปฏิกิริยาในกรณีของออโรเตตดีคาร์บอกซิเลส (78 ล้านปีที่ไม่มีเอนไซม์ และ 18 มิลลิวินาทีสำหรับเอนไซม์)
โมเลกุลที่เอนไซม์จับและกระทำเรียกว่าสารตั้งต้นแม้ว่าเอนไซม์อาจประกอบด้วยกรดอะมิโนหลายร้อยชนิด แต่โดยปกติแล้วจะมีสารตกค้างเพียงเล็กน้อยเท่านั้นที่สัมผัสกับสารตั้งต้น และสารตกค้างที่น้อยกว่านั้น คือโดยเฉลี่ยแล้วมีสารตกค้างสามถึงสี่ชนิด ซึ่งเกี่ยวข้องโดยตรงกับกระบวนการเร่งปฏิกิริยาบริเวณของเอนไซม์ที่จับสารตั้งต้นและบรรจุสารตกค้างที่เป็นตัวเร่งปฏิกิริยา เรียกว่าบริเวณแอคทีฟไซต์ [
โปรตีน Dirigentเป็นสมาชิกของกลุ่มโปรตีนที่กำหนดสเตอริโอเคมีของสารประกอบที่สังเคราะห์โดยเอนไซม์อื่น
การส่งสัญญาณเซลล์และการจับกับลิแกนด์

โปรตีนหลายชนิดมีส่วนเกี่ยวข้องในกระบวนการ ส่งสัญญาณ และการถ่ายทอดสัญญาณ ของ เซลล์โปรตีนบางชนิด เช่นอินซูลินเป็นโปรตีนนอกเซลล์ที่ส่งสัญญาณจากเซลล์ที่สังเคราะห์ไปยังเซลล์อื่นๆ ในเนื้อเยื่อ ที่อยู่ห่างไกล โปรตีน บางชนิดเป็นโปรตีนเยื่อหุ้มเซลล์ที่ทำหน้าที่เป็นตัวรับซึ่งหน้าที่หลักคือการจับกับโมเลกุลส่งสัญญาณและกระตุ้นการตอบสนองทางชีวเคมีภายในเซลล์ ตัวรับหลายชนิดมีจุดจับที่โผล่ออกมาบนพื้นผิวเซลล์และมีโดเมนเอฟเฟกเตอร์ภายในเซลล์ ซึ่งอาจมีกิจกรรมทางเอนไซม์หรืออาจเกิดการเปลี่ยนแปลงโครงสร้างที่ตรวจพบโดยโปรตีนอื่นๆ ภายในเซลล์
แอนติบอดีเป็นส่วนประกอบโปรตีนของระบบภูมิคุ้มกันแบบปรับตัวซึ่งหน้าที่หลักคือการจับแอนติเจนหรือสารแปลกปลอมในร่างกาย และกำหนดเป้าหมายเพื่อทำลาย แอนติบอดีสามารถหลั่งออกสู่สิ่งแวดล้อมภายนอกเซลล์ หรือยึดเกาะในเยื่อหุ้มเซลล์บี เฉพาะทาง ที่เรียกว่าพลาสมาเซลล์ในขณะที่เอนไซม์มีความสัมพันธ์ในการจับกับสารตั้งต้นที่จำกัดเนื่องจากความจำเป็นในการดำเนินปฏิกิริยา แต่แอนติบอดีไม่มีข้อจำกัดดังกล่าว ความสัมพันธ์ในการจับของแอนติบอดีกับสารตั้งต้นนั้นสูงมากเป็นพิเศษ
โปรตีนขนส่งลิแกนด์หลายชนิดจับกับไบโอโมเลกุลขนาดเล็กบาง ชนิด และขนส่งไปยังตำแหน่งอื่นๆ ในร่างกายของสิ่งมีชีวิตหลายเซลล์ โปรตีนเหล่านี้ต้องมีความสัมพันธ์ในการจับสูงเมื่อมีลิแกนด์อยู่ในความเข้มข้นสูง และปล่อยลิแกนด์ออกมาเมื่อมีลิแกนด์อยู่ในความเข้มข้นต่ำในเนื้อเยื่อเป้าหมาย ตัวอย่างทั่วไปของโปรตีนที่จับกับลิแกนด์คือฮีโมโกลบินซึ่งขนส่งออกซิเจนจากปอดไปยังอวัยวะและเนื้อเยื่ออื่นๆ ในสัตว์มีกระดูกสันหลัง ทุกชนิด และมีโฮโมล็อกใกล้เคียงกันในทุกอาณาจักรชีวภาพ เลกตินเป็นโปรตีนที่จับกับน้ำตาลซึ่งมีความจำเพาะสูงสำหรับหมู่น้ำตาลของมัน โดยทั่วไปแล้ว เลกตินมีบทบาทใน ปรากฏการณ์ การจดจำ ทางชีวภาพ ที่เกี่ยวข้องกับเซลล์และโปรตีน ตัวรับและฮอร์โมนเป็นโปรตีนที่จับกับความจำเพาะสูง
โปรตีนทรานส์เมมเบรนสามารถทำหน้าที่เป็นโปรตีนขนส่งลิแกนด์ที่เปลี่ยนการซึมผ่านของเยื่อหุ้มเซลล์ไปยังโมเลกุลและไอออนขนาดเล็ก เยื่อหุ้มเซลล์มี แกนกลางที่ไม่ ชอบน้ำซึ่งโมเลกุลที่มีขั้ว หรือประจุไม่สามารถ แพร่ ผ่าน ได้ โปรตีนเยื่อหุ้มเซลล์มีช่องภายในที่ช่วยให้โมเลกุลเหล่านี้เข้าและออกจากเซลล์ได้ โปรตีน ช่องไอออน หลายชนิด ถูกออกแบบเฉพาะเพื่อคัดเลือกเฉพาะไอออนเฉพาะ ตัวอย่างเช่น ช่อง โพแทสเซียมและโซเดียมมักจะเลือกเฉพาะไอออนใดไอออนหนึ่งจากสองไอออน
โปรตีนโครงสร้าง

โปรตีนโครงสร้างให้ความแข็งแกร่งและความยืดหยุ่นแก่ส่วนประกอบทางชีวภาพที่มีลักษณะเป็นของเหลว โปรตีนโครงสร้างส่วนใหญ่เป็นโปรตีนเส้นใยตัวอย่างเช่นคอลลาเจนและอีลาสตินเป็นองค์ประกอบสำคัญของเนื้อเยื่อเกี่ยวพันเช่นกระดูกอ่อนและเคราตินพบได้ในโครงสร้างแข็งหรือเส้นใย เช่นผมเล็บขนกีบและกระดองสัตว์บางชนิด[ 42 โปรตีนทรงกลมบางชนิด สามารถทำหน้าที่เชิงโครงสร้าง ได้เช่นแอคตินและทูบูลินเป็นโมโนเมอร์ที่มีลักษณะเป็นทรงกลมและละลายน้ำได้ แต่เกิด การพอลิเมอไรเซ ชันจนกลายเป็นเส้นใยยาวแข็งที่ประกอบกันเป็นโครงร่างเซลล์ซึ่งช่วยให้เซลล์คงรูปร่างและขนาดไว้ได้
โปรตีนอื่นๆ ที่ทำหน้าที่โครงสร้าง ได้แก่โปรตีนมอเตอร์เช่นไมโอซินไคเนซินและไดนีอินซึ่งสามารถสร้างแรงเชิงกลได้ โปรตีนเหล่านี้มีความสำคัญอย่างยิ่งต่อการเคลื่อนที่ ของเซลล์ ของสิ่งมีชีวิตเซลล์เดียว และอสุจิของสิ่งมีชีวิตหลายเซลล์หลายชนิดที่สืบพันธุ์แบบอาศัยเพศโปรตีนเหล่านี้สร้างแรงที่เกิดจากการหดตัวของกล้ามเนื้อและมีบทบาทสำคัญในการขนส่งภายในเซลล์
วิธีการศึกษา
วิธีการที่ใช้กันทั่วไปในการศึกษาโครงสร้างและหน้าที่ของโปรตีน ได้แก่อิมมูโนฮิสโตเคมี , การกลายพันธุ์แบบกำหนดตำแหน่ง , ผลึกศาสตร์รังสีเอกซ์ , เรโซแนนซ์แม่เหล็กนิวเคลียร์และแมสสเปกโตรมิเตอร์ กิจกรรมและโครงสร้างของโปรตีนสามารถตรวจสอบได้ ทั้งในหลอดทดลอง , ในสิ่งมีชีวิตและในซิลิโกการ ศึกษา ใน หลอดทดลอง ของโปรตีนบริสุทธิ์ในสภาพแวดล้อมควบคุมมีประโยชน์สำหรับการเรียนรู้ว่าโปรตีนทำงานอย่างไรตัวอย่างเช่น การศึกษา จลนพลศาสตร์ของเอนไซม์จะสำรวจกลไกทางเคมีของกิจกรรมการเร่งปฏิกิริยาของเอนไซม์และความสัมพันธ์สัมพัทธ์กับโมเลกุลสารตั้งต้นต่างๆ ] ในทางตรงกันข้าม การทดลอง ในสิ่งมีชีวิตสามารถให้ข้อมูลเกี่ยวกับบทบาททางสรีรวิทยาของโปรตีนในบริบทของเซลล์หรือแม้แต่สิ่งมีชีวิต ทั้งหมด และมักจะให้ข้อมูลเพิ่มเติมเกี่ยวกับพฤติกรรมของโปรตีนในบริบทที่แตกต่างกัน ในซิลิโกใช้วิธีการคำนวณเพื่อศึกษาโปรตีน
การฟอกโปรตีน
โปรตีนอาจถูกทำให้บริสุทธิ์จากส่วนประกอบของเซลล์อื่น ๆ โดยใช้เทคนิคต่าง ๆ เช่นการแยกด้วยแรงเหวี่ยงสูง การตกตะกอนอิ เล็กโตร โฟรีซิสและโครมาโทกราฟี [ การถือกำเนิดของวิศวกรรมพันธุกรรมทำให้มีวิธีการต่าง ๆ มากมายในการอำนวยความสะดวกในการทำให้บริสุทธิ์
ในการ วิเคราะห์ ในหลอดทดลองโปรตีนจะต้องถูกทำให้บริสุทธิ์ออกจากส่วนประกอบอื่นๆ ของเซลล์ กระบวนการนี้มักเริ่มต้นด้วยการสลายเซลล์ซึ่งเยื่อหุ้มเซลล์จะถูกทำลายและสารภายในจะถูกปล่อยลงในสารละลายที่เรียกว่าไลเสทหยาบส่วนผสมที่ได้สามารถถูกทำให้บริสุทธิ์ได้โดยใช้การปั่นเหวี่ยงด้วยพลังสูงซึ่งจะแยกส่วนประกอบต่างๆ ของเซลล์ออกเป็นเศษส่วนที่มีโปรตีนที่ละลายน้ำได้ ไขมันและโปรตีนในเยื่อหุ้มเซลล์ ออร์แกเนลล์ของเซลล์และกรดนิวคลีอิกการตกตะกอนด้วยวิธีที่เรียกว่าการตกเกลือ (salting out) สามารถทำให้โปรตีนจากไลเสทนี้เข้มข้นขึ้นได้ จากนั้นจึงใช้ โครมาโทกราฟีหลายประเภทเพื่อแยกโปรตีนที่ต้องการโดยพิจารณาจากคุณสมบัติต่างๆ เช่น น้ำหนักโมเลกุล ประจุสุทธิ และค่าสัมประสิทธิ์การจับยึดสามารถตรวจสอบระดับการทำให้บริสุทธิ์ได้โดยใช้เจลอิเล็กโทรโฟเรซิส หลายประเภท หาก ทราบ น้ำหนักโมเลกุลและ จุดไอโซอิเล็กทริกของโปรตีนที่ต้องการ ใช้ สเปกโทรสโกปีหากโปรตีนมีคุณสมบัติทางสเปกโทรสโกปีที่สามารถแยกแยะได้ หรือโดยใช้เอนไซม์หากโปรตีนมีกิจกรรมทางเอนไซม์ นอกจากนี้ โปรตีนยังสามารถแยกตามประจุได้โดยใช้อิเล็กโทรโฟกัส [
สำหรับโปรตีนธรรมชาติ อาจจำเป็นต้องมีขั้นตอนการทำให้บริสุทธิ์หลายขั้นตอนเพื่อให้ได้โปรตีนที่มีความบริสุทธิ์เพียงพอสำหรับการประยุกต์ใช้ในห้องปฏิบัติการ เพื่อลดความซับซ้อนของกระบวนการนี้ มักใช้ พันธุวิศวกรรมเพื่อเพิ่มคุณสมบัติทางเคมีให้กับโปรตีน ซึ่งทำให้โปรตีนบริสุทธิ์ได้ง่ายขึ้นโดยไม่ส่งผลกระทบต่อโครงสร้างหรือกิจกรรมของโปรตีน ในกรณีนี้ จะมี "แท็ก" ซึ่งประกอบด้วยลำดับกรดอะมิโนเฉพาะ ซึ่งมักจะเป็นชุดของ กรดอะ มิโนฮิสทิดีน (หรือ " His-tag ") ติดอยู่ที่ปลายด้านหนึ่งของโปรตีน ดังนั้น เมื่อไลเสทถูกส่งผ่านคอลัมน์โครมาโทกราฟีที่มีนิกเกิล กรดอะมิโนฮิสทิดีนจะเชื่อมนิกเกิลและยึดติดกับคอลัมน์ ในขณะที่ส่วนประกอบที่ไม่มีแท็กของไลเสทจะผ่านได้อย่างไม่ติดขัด มีการพัฒนาแท็กจำนวนหนึ่งเพื่อช่วยนักวิจัยในการทำให้โปรตีนเฉพาะบริสุทธิ์จากส่วนผสมที่ซับซ้อน
การระบุตำแหน่งเซลล์

การศึกษาโปรตีนในร่างกายมักเกี่ยวข้องกับการสังเคราะห์และการแปลตำแหน่งของโปรตีนภายในเซลล์ แม้ว่าโปรตีนภายในเซลล์หลายชนิดจะถูกสังเคราะห์ในไซโทพลาสซึมและโปรตีนที่ยึดติดหรือหลั่งออกมาในเอนโดพลาสมิกเรติคูลัม แต่รายละเอียดเฉพาะเกี่ยวกับวิธีการที่โปรตีนถูกกำหนดเป้าหมายไปยังออร์แกเนลล์หรือโครงสร้างเซลล์ที่เฉพาะเจาะจงนั้นมักไม่ชัดเจน เทคนิคที่มีประโยชน์สำหรับการประเมินการแปลตำแหน่งของเซลล์คือการใช้พันธุวิศวกรรมเพื่อแสดงโปรตีนฟิวชันหรือไคเมรา ในเซลล์ ซึ่งประกอบด้วยโปรตีนธรรมชาติที่น่าสนใจซึ่งเชื่อมโยงกับ " รีพอร์เตอร์ " เช่นโปรตีนเรืองแสงสีเขียว (GFP) จากนั้นสามารถมองเห็นตำแหน่งของโปรตีนที่หลอมรวมภายในเซลล์ได้อย่างชัดเจนและมีประสิทธิภาพโดยใช้กล้องจุลทรรศน์[
วิธีการอื่นๆ ในการอธิบายตำแหน่งของโปรตีนในเซลล์ จำเป็นต้องใช้เครื่องหมายช่องที่ทราบแล้วสำหรับบริเวณต่างๆ เช่น ER, กอลจิ, ไลโซโซมหรือแวคิวโอล, ไมโทคอนเดรีย, คลอโรพลาสต์, เยื่อหุ้มพลาสมา ฯลฯ การใช้เครื่องหมายเหล่านี้ในรูปแบบที่มีการติดแท็กเรืองแสง หรือแอนติบอดีต่อเครื่องหมายที่ทราบแล้ว ทำให้การระบุตำแหน่งของโปรตีนที่ต้องการทำได้ง่ายขึ้นมาก ตัวอย่างเช่นการเรืองแสงทางอ้อม (indirect immunofluorescence)จะช่วยให้สามารถระบุตำแหน่งที่อยู่ร่วมกันของฟลูออเรสเซนซ์และแสดงตำแหน่งได้ สีย้อมเรืองแสงถูกนำมาใช้เพื่อติดฉลากช่องเซลล์เพื่อวัตถุประสงค์ที่คล้ายคลึงกัน
นอกจากนี้ยังมีความเป็นไปได้อื่นๆ อีกด้วย ตัวอย่างเช่นอิมมูโนฮิสโตเคมีมักใช้แอนติบอดีต่อโปรตีนที่น่าสนใจอย่างน้อยหนึ่งชนิด ซึ่งถูกจับคู่กับเอนไซม์ที่ให้สัญญาณเรืองแสงหรือสัญญาณโครโมเจนิก ซึ่งสามารถนำมาเปรียบเทียบระหว่างตัวอย่างต่างๆ ได้ ทำให้ได้ข้อมูลการระบุตำแหน่งอีกเทคนิคหนึ่งที่สามารถใช้ได้คือการแบ่งส่วนร่วมในความชันของซูโครส (หรือวัสดุอื่นๆ) โดยใช้ การปั่นเหวี่ยงแบบไอ โซพิกนิกแม้ว่าเทคนิคนี้จะไม่สามารถพิสูจน์การอยู่ร่วมกันของส่วนที่มีความหนาแน่นที่ทราบและโปรตีนที่น่าสนใจได้ แต่ก็แสดงให้เห็นถึงความเป็นไปได้ที่เพิ่มขึ้น
สุดท้าย วิธีการมาตรฐานทองคำในการระบุตำแหน่งเซลล์คือกล้องจุลทรรศน์อิมมูโนอิเล็กตรอนเทคนิคนี้ใช้แอนติบอดีต่อโปรตีนที่ต้องการ ร่วมกับเทคนิคกล้องจุลทรรศน์อิเล็กตรอนแบบคลาสสิก ตัวอย่างจะถูกเตรียมไว้สำหรับการตรวจด้วยกล้องจุลทรรศน์อิเล็กตรอนแบบปกติ จากนั้นจึงใช้แอนติบอดีต่อโปรตีนที่ต้องการ ซึ่งถูกจับคู่กับวัสดุที่มีความหนาแน่นสูงเป็นพิเศษ ซึ่งโดยปกติแล้วจะเป็นทองคำ วิธีนี้ช่วยให้สามารถระบุตำแหน่งทั้งรายละเอียดทางจุลภาคโครงสร้างและโปรตีนที่ต้องการได้
ด้วยการประยุกต์ใช้ทางพันธุวิศวกรรมอีกวิธีหนึ่งที่เรียกว่า การกลายพันธุ์แบบกำหนดตำแหน่ง ( site-directed mutagenesis ) นักวิจัยสามารถเปลี่ยนแปลงลำดับโปรตีน ส่งผลให้โครงสร้าง ตำแหน่งของโปรตีน และความไวต่อการควบคุมเปลี่ยนแปลงไป เทคนิคนี้ยังช่วยให้สามารถรวมกรดอะมิโนที่ไม่เป็นธรรมชาติเข้ากับโปรตีนได้ โดยใช้ tRNA ที่ผ่านการดัดแปลงและอาจช่วยให้สามารถออกแบบโปรตีนใหม่ที่มีคุณสมบัติใหม่ๆ ได้ อย่างมีเหตุผล
โปรตีโอมิกส์
ส่วนประกอบทั้งหมดของโปรตีนที่ปรากฏ ณ เวลาใดเวลาหนึ่งในเซลล์หรือชนิดของเซลล์ เรียกว่าโปรตีโอมและการศึกษาชุดข้อมูลขนาดใหญ่ดังกล่าวได้กำหนดขอบเขตของโปรตีโอมิกส์ซึ่งตั้งชื่อโดยเปรียบเทียบกับขอบเขตที่เกี่ยวข้องของจีโนมิกส์เทคนิคการทดลองที่สำคัญในโปรตีโอมิกส์ประกอบด้วย อิเล็กโทรโฟรีซิ สแบบสองมิติซึ่งช่วยให้สามารถแยกโปรตีนได้หลายชนิดแมสสเปกโตรมิเตอร์ [ ซึ่งช่วยให้สามารถระบุโปรตีนและจัดลำดับเปปไทด์ได้อย่างรวดเร็วและมีประสิทธิภาพสูง (ส่วนใหญ่หลังจากการย่อยในเจล ) โปรตีนไมโครอาร์เรย์ซึ่งช่วยให้สามารถตรวจจับระดับสัมพัทธ์ของโปรตีนต่างๆ ที่มีอยู่ในเซลล์ และการคัดกรองแบบไฮบริดสองแบบซึ่งช่วยให้สามารถสำรวจปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีน ได้อย่างเป็นระบบ ส่วนประกอบทั้งหมดของปฏิสัมพันธ์ที่เป็นไปได้ทางชีวภาพดังกล่าว เรียกว่าอินเทอร์แอค โท มความพยายามอย่างเป็นระบบในการกำหนดโครงสร้างของโปรตีนที่แสดงถึงการพับที่เป็นไปได้ทั้งหมด เรียกว่าจีโนมิกส์เชิงโครงสร้าง
การกำหนดโครงสร้าง
การค้นพบโครงสร้างตติยภูมิของโปรตีน หรือโครงสร้างจตุภาคของสารประกอบเชิงซ้อน สามารถให้เบาะแสสำคัญเกี่ยวกับการทำงานของโปรตีนและผลกระทบที่อาจเกิดขึ้นได้ เช่น ในการออกแบบยาเนื่องจากโปรตีนมีขนาดเล็กเกินกว่าจะมองเห็นได้ด้วยกล้องจุลทรรศน์แบบใช้แสงจึงจำเป็นต้องใช้วิธีการอื่นเพื่อระบุโครงสร้างของโปรตีน วิธีการทดลองทั่วไป ได้แก่ผลึกศาสตร์รังสีเอกซ์และสเปกโทรสโกปี NMRซึ่งทั้งสองวิธีนี้สามารถให้ข้อมูลโครงสร้างที่ ความละเอียดระดับ อะตอมอย่างไรก็ตาม การทดลอง NMR สามารถให้ข้อมูลที่สามารถประมาณระยะทางย่อยระหว่างคู่อะตอม และหาโครงสร้างขั้นสุดท้ายที่เป็นไปได้ของโปรตีนโดยการแก้ปัญหาเรขาคณิตระยะทางอินเตอร์เฟอโรมิเตอร์แบบโพลาไรเซชันคู่เป็นวิธีการวิเคราะห์เชิงปริมาณสำหรับการวัดโครงสร้างโดยรวมของโปรตีนและการเปลี่ยนแปลงโครงสร้างอัน เนื่อง มาจากปฏิกิริยาหรือสิ่งกระตุ้นอื่นๆ ได โค รอิซึมแบบวงกลม เป็นอีกเทคนิคหนึ่งในห้องปฏิบัติการสำหรับการกำหนดองค์ประกอบภายในของโปรตีนแบบแผ่นเบตา/เกลียวอัลฟากล้องจุลทรรศน์อิเล็กตรอนแบบแช่แข็งใช้เพื่อสร้างข้อมูลโครงสร้างที่มีความละเอียดต่ำเกี่ยวกับกลุ่มโปรตีนขนาดใหญ่มาก รวมถึงไวรัสที่ประกอบเข้าด้วยกัน[ 41 การวิเคราะห์ผลึกอิเล็กตรอนแบบแปรผันสามารถผลิตข้อมูลความละเอียดสูงได้ในบางกรณี โดยเฉพาะอย่างยิ่งสำหรับผลึกสองมิติของโปรตีนเยื่อหุ้มเซลล์โครงสร้างที่แก้ไขแล้วมักจะถูกเก็บไว้ในProtein Data Bank (PDB) ซึ่งเป็นแหล่งข้อมูลที่เข้าถึงได้ฟรีซึ่งสามารถรับข้อมูลโครงสร้างเกี่ยวกับโปรตีนหลายพันชนิดในรูปแบบพิกัดคาร์ทีเซียนสำหรับแต่ละอะตอมในโปรตีน
มีลำดับยีนที่รู้จักมากกว่าโครงสร้างโปรตีนเสียอีก ยิ่งไปกว่านั้น ชุดของโครงสร้างที่แก้ไขแล้วยังเอนเอียงไปทางโปรตีนที่สามารถอยู่ภายใต้เงื่อนไขที่จำเป็นในการวิเคราะห์ผลึกศาสตร์ด้วยรังสีเอกซ์ซึ่งเป็นหนึ่งในวิธีการกำหนดโครงสร้างหลัก โดยเฉพาะอย่างยิ่ง โปรตีนทรงกลมนั้นค่อนข้างตกผลึก ได้ง่าย เมื่อเตรียมสำหรับการวิเคราะห์ผลึกศาสตร์ด้วยรังสีเอกซ์ ในทางตรงกันข้าม โปรตีนเมมเบรนและโปรตีนเชิงซ้อนขนาดใหญ่นั้นตกผลึกได้ยากและมีสัดส่วนน้อยใน PDB จีโนมิกส์เชิงโครงสร้างได้พยายามแก้ไขข้อบกพร่องเหล่านี้โดยการแก้โครงสร้างตัวแทนของคลาสพับหลักอย่างเป็นระบบ วิธี การทำนายโครงสร้างโปรตีนพยายามหาวิธีการสร้างโครงสร้างที่น่าเชื่อถือสำหรับโปรตีนที่ยังไม่สามารถระบุโครงสร้างได้จากการทดลอง
การทำนายโครงสร้าง

การทำนายโครงสร้างโปรตีนเป็นงานเสริมของสาขาจีโนมิกส์เชิงโครงสร้างโดยพัฒนาแบบจำลองทางคณิตศาสตร์ ที่มีประสิทธิภาพ ของโปรตีนเพื่อทำนายการก่อตัวของโมเลกุลในทางทฤษฎีด้วยการคำนวณ แทนที่จะตรวจจับโครงสร้างด้วยการสังเกตในห้องปฏิบัติการการทำนายโครงสร้างที่ประสบความสำเร็จมากที่สุด เรียกว่า การสร้างแบบ จำลองโฮ โมโลยี (homology modeling ) ซึ่งอาศัยการมีโครงสร้าง "แม่แบบ" ที่มีความคล้ายคลึงกันของลำดับกับโปรตีนที่กำลังสร้างแบบจำลอง เป้าหมายของจีโนมิกส์เชิงโครงสร้างคือการให้การแสดงที่เพียงพอในโครงสร้างที่แก้ไขแล้วเพื่อสร้างแบบจำลองของโครงสร้างส่วนใหญ่ที่เหลืออยู่แม้ว่าการสร้างแบบจำลองที่แม่นยำจะยังคงเป็นความท้าทายเมื่อมีเพียงโครงสร้างแม่แบบที่สัมพันธ์กันในระยะไกลเท่านั้น แต่ก็มีการเสนอว่าการจัดเรียงลำดับเป็นคอขวดในกระบวนการนี้ เนื่องจากสามารถสร้างแบบจำลองที่มีความแม่นยำได้มากหากทราบการจัดเรียงลำดับที่ "สมบูรณ์แบบ" วิธีการทำนายโครงสร้างหลายวิธีได้ถูกนำมาใช้เป็นข้อมูลสำหรับสาขาวิศวกรรมโปรตีน ที่กำลังเกิดขึ้น ใหม่ ซึ่งได้มีการออกแบบโครงสร้างพับโปรตีนแบบใหม่ไว้แล้วโปรตีนหลายชนิด (ในยูคาริโอต ~33%) ประกอบด้วยส่วนขนาดใหญ่ที่ไม่มีโครงสร้างแต่ทำหน้าที่ทางชีวภาพ และสามารถจัดเป็นโปรตีนที่มีความผิดปกติภายในได้การทำนายและวิเคราะห์ความผิดปกติของโปรตีนเป็นส่วนสำคัญของการจำแนกลักษณะโครงสร้างโปรตีน
การจำลองกระบวนการไดนามิกในซิลิโก
ปัญหาการคำนวณที่ซับซ้อนกว่าคือการทำนายปฏิสัมพันธ์ระหว่างโมเลกุล เช่นการเชื่อมต่อโมเลกุล [ พับตัวของโปรตีนปฏิสัมพันธ์ระหว่างโปรตีนและโปรตีนและปฏิกิริยาเคมี แบบจำลองทางคณิตศาสตร์เพื่อจำลองกระบวนการพลวัตเหล่านี้เกี่ยวข้องกับกลศาสตร์โมเลกุลโดยเฉพาะอย่างยิ่งพลวัตโมเลกุลในเรื่องนี้ การจำลองแบบ in silicoค้นพบการพับตัวของโดเมนโปรตีน ขนาดเล็กแบบ α-helical เช่นส่วนหัวของวิลลินโปรตีนเสริมของ HIV และวิธีการแบบไฮบริดที่ผสมผสานพลวัตโมเลกุลมาตรฐานเข้ากับ คณิตศาสตร์ กลศาสตร์ควอนตัมได้สำรวจสถานะทางอิเล็กทรอนิกส์ของโรดอปซิน [
นอกเหนือจากพลศาสตร์โมเลกุลแบบคลาสสิกแล้ว วิธี พลศาสตร์ควอนตัมยังช่วยให้สามารถจำลองโปรตีนได้อย่างละเอียดในระดับอะตอม พร้อมคำอธิบายผลกระทบทางกลควอนตัมที่แม่นยำ ตัวอย่างเช่น วิธี Hartree แบบหลายชั้นหลายรูปแบบที่ขึ้นกับเวลาและวิธี สมการ การเคลื่อนที่แบบลำดับชั้นซึ่งถูกนำไปประยุกต์ใช้กับคริปโทโครมของพืชและสารเชิงซ้อนสำหรับการเก็บเกี่ยวแสงของแบคทีเรียตามลำดับ ทั้งการจำลองเชิงกลแบบควอนตัมและแบบคลาสสิกของระบบในระดับชีวภาพล้วนต้องการการประมวลผลอย่างมาก ดังนั้น โครงการริเริ่ม การประมวลผลแบบกระจายเช่นโครงการFolding@home จึงช่วยอำนวยความสะดวกในการสร้าง แบบจำลองโมเลกุลโดยใช้ประโยชน์จากความก้าวหน้าใน การประมวลผลแบบขนาน ของ GPUและเทคนิคMonte Carlo
การวิเคราะห์ทางเคมี
ปริมาณไนโตรเจนรวมของสารอินทรีย์ส่วนใหญ่เกิดจากหมู่อะมิโนในโปรตีนไนโตรเจนรวมแบบเจลดาห์ล (TKN) เป็นหน่วยวัดไนโตรเจนที่ใช้กันอย่างแพร่หลายในการวิเคราะห์น้ำ (ของเสีย) ดิน อาหาร อาหารสัตว์ และสารอินทรีย์โดยทั่วไป ดังชื่อที่บ่งบอกวิธีเจลดาห์ลถูกนำมาใช้ มีวิธีการที่ละเอียดอ่อนกว่านี้
การย่อยอาหาร

เมื่อไม่มีตัวเร่งปฏิกิริยา โปรตีนจะไฮโดรไลซ์ได้ ช้า การสลายโปรตีนให้เป็นเปปไทด์และกรดอะมิโนขนาดเล็ก ( โปรตีโอไลซิส ) เป็นขั้นตอนหนึ่งในการย่อยอาหารซึ่งผลิตภัณฑ์ที่ได้จากการสลายเหล่านี้จะถูกดูดซึมในลำไส้เล็กการไฮโดรไลซิสของโปรตีนอาศัยเอนไซม์ที่เรียกว่าโปรตีเอสหรือเปปไทเดส โปรตีเอสซึ่งเป็นโปรตีนเองก็มีหลายประเภทตามพันธะเปปไทด์ที่มันตัดออก รวมถึงแนวโน้มที่จะตัดพันธะเปปไทด์ที่ปลายของโปรตีน (เอ็กโซเปปไทเดส) เทียบกับพันธะเปปไทด์ที่ด้านในของโปรตีน (เอนโดเปปไทเดส) เปปซินเป็นเอนโดเปปไทเดสในกระเพาะอาหาร หลังจากกระเพาะอาหาร ตับอ่อนจะหลั่งโปรตีเอสชนิดอื่นเพื่อให้การไฮโดรไลซิสเสร็จสมบูรณ์ ซึ่งรวมถึงทริปซินและไคโมทริปซิน [
การไฮโดรไลซิสโปรตีนถูกนำมาใช้ในเชิงพาณิชย์เพื่อผลิตกรดอะมิโนจากแหล่งโปรตีนจำนวนมาก เช่น เลือดป่น ขน และเคราติน วัสดุเหล่านี้จะถูกนำไปผ่านกระบวนการไฮโดรไลซิสด้วยกรดไฮโดรคลอริก ร้อน ซึ่งมีผลต่อการไฮโดรไลซิสของพันธะเปปไทด์
คุณสมบัติเชิงกล
คุณสมบัติเชิงกลของโปรตีนมีความหลากหลายอย่างมากและมักเป็นศูนย์กลางของหน้าที่ทางชีวภาพของโปรตีน เช่น ในกรณีของโปรตีนอย่างเคราตินและคอลลาเจน [ ยกตัวอย่างเช่น ความสามารถของเนื้อเยื่อกล้ามเนื้อในการขยายตัวและหดตัวอย่างต่อเนื่องนั้นเชื่อมโยงโดยตรงกับคุณสมบัติความยืดหยุ่นของโครงสร้างโปรตีนพื้นฐานนอกเหนือจากโปรตีนเส้นใยแล้ว พลวัตเชิงโครงสร้างของเอนไซม์ และโครงสร้างของเยื่อหุ้มเซลล์ชีวภาพรวมถึงหน้าที่ทางชีวภาพอื่นๆ ยังถูกควบคุมโดยคุณสมบัติเชิงกลของโปรตีน นอกเหนือจากบริบททางชีวภาพ คุณสมบัติเชิงกลเฉพาะตัวของโปรตีนหลายชนิด รวมถึงความยั่งยืนเมื่อเทียบกับพอลิเมอร์สังเคราะห์ทำให้โปรตีนเหล่านี้เป็นเป้าหมายที่พึงประสงค์สำหรับการออกแบบวัสดุรุ่นต่อไป
โมดูลั สของยังEคำนวณจากความเค้นตามแนวแกนσเหนือความเครียดที่เกิดขึ้นεเป็นการวัดความแข็ง สัมพัทธ์ ของวัสดุ ในบริบทของโปรตีน ความแข็งนี้มักสัมพันธ์โดยตรงกับหน้าที่ทางชีวภาพ ตัวอย่างเช่นคอลลาเจนซึ่งพบในเนื้อเยื่อเกี่ยวพันกระดูกและกระดูกอ่อนและเคราตินซึ่งพบในเล็บ กรงเล็บ และผม พบว่ามีความแข็งที่สูงกว่าอีลาสติน หลายเท่า [ ซึ่งเชื่อกันว่าให้ความยืดหยุ่นแก่โครงสร้างต่างๆ เช่นหลอดเลือดเนื้อเยื่อปอดและเนื้อเยื่อกระเพาะปัสสาวะเป็นต้นเมื่อเปรียบเทียบกับสิ่งนี้โปรตีนทรงกลมเช่นBovine Serum Albuminซึ่งลอยได้ค่อนข้างอิสระในไซโทซอลและมักทำหน้าที่เป็นเอนไซม์ (และจึงมีการเปลี่ยนแปลงโครงสร้างบ่อยครั้ง) จะมีโมดูลัสของยังต่ำกว่ามาก
โมดูลัสของยังของโปรตีนเดี่ยวสามารถพบได้ผ่าน การจำลอง พลวัตเชิงโมเลกุลการใช้สนามแรงแบบอะตอม เช่นCHARMMหรือGROMOSหรือสนามแรงแบบหยาบ เช่น Martini โมเลกุลโปรตีนเดี่ยวสามารถถูกยืดออกได้ด้วยแรงแกนเดียว ในขณะที่การยืดออกที่เกิดขึ้นจะถูกบันทึกเพื่อคำนวณความเครียดในการทดลอง วิธีการต่างๆ เช่นกล้องจุลทรรศน์แรงอะตอมสามารถใช้เพื่อให้ได้ข้อมูลที่คล้ายคลึงกันพลวัตภายในของโปรตีนเกี่ยวข้องกับความยืดหยุ่นและการเปลี่ยนรูปพลาสติกที่ละเอียดอ่อนซึ่งเกิดจาก แรง หนืดหยุ่นซึ่งสามารถตรวจสอบได้ด้วยเทคนิค นา โนรีโอโลยีการประมาณค่าเหล่านี้ให้ค่าคงที่ของสปริงทั่วไปที่ประมาณk ≈ 100 pN/nm เทียบเท่ากับโมดูลัสของยังที่E ≈ 100 MPa และค่าสัมประสิทธิ์แรงเสียดทานทั่วไปที่γ ≈ 0.1 pN·s/nm ซึ่งสอดคล้องกับความหนืดที่η ≈ 0.01 pN·s/nm = 10 cP (นั่นคือ มีความหนืดมากกว่าน้ำ 10
ในระดับมหภาค โมดูลัสของยังของโครงข่ายโปรตีนที่เชื่อมขวางสามารถหาได้จากการทดสอบเชิงกล แบบดั้งเดิม สามารถดูค่าที่สังเกตได้จากการทดลองของโปรตีนบางชนิดได้ด้านล่าง
| โปรตีน | คลาสโปรตีน | โมดูลัสของยัง |
|---|---|---|
| เคราติน (เชื่อมขวาง) | เส้นใย | เกรดเฉลี่ย 1.5–10 |
| อีลาสติน (เชื่อมขวาง) | เส้นใย | 1 เมกะปาสคาล |
| ไฟบริน (เชื่อมขวาง) | เส้นใย | 1–10 เมกะปาสคาล |
| คอลลาเจน (เชื่อมขวาง) | เส้นใย | เกรดเฉลี่ย 5–7.5 |
| เรซิลิน (เชื่อมขวาง) | เส้นใย | 1–2 เมกะปาสคาล |
| อัลบูมินในซีรั่มวัว (เชื่อมโยง) | ทรงกลม | 2.5–15 กิโลปาสคาล |
| โปรตีนเยื่อหุ้มชั้นนอกของเบตาบาร์เรล | เยื่อหุ้มเซลล์ | เกรดเฉลี่ย 20–45 |
ดูเพิ่มเติม
- การสูญเสียโปรตีน – เทคนิคในการวิจัยวัสดุมีชีวิต
- โปรตีนที่จับกับดีเอ็นเอ – โปรตีนที่จับกับดีเอ็นเอ
- ดัชนีบทความที่เกี่ยวข้องกับโปรตีน
- อินเทน
- รายชื่อโปรตีน
- โมเลกุลขนาด ใหญ่ – โมเลกุลขนาดใหญ่มาก เช่น โปรตีน
- วิวัฒนาการของโปรตีน – การศึกษาการเปลี่ยนแปลงใน DNA และ RNA เมื่อเวลาผ่านไป
- พื้นที่ลำดับโปรตีน – การแสดงลำดับพันธุกรรมที่เป็นไปได้
- โปรตีนซูเปอร์แฟมิลี – การจัดกลุ่มของโปรตีน
- ความเป็นพิษของโปรตีน – การสะสมของเสียจากการเผาผลาญเนื่องจากการทำงานของไตผิดปกติ
- โปรทีโอพาธี – โรคที่เกิดจากโครงสร้างโปรตีนที่ผิดปกติหน้าที่แสดงคำอธิบายสั้น ๆ ของเป้าหมายการเปลี่ยนเส้นทาง
- Proteopedia – สารานุกรม 3 มิติของโปรตีนและโมเลกุลอื่นๆ
อ่านเพิ่มเติม
- หนังสือเรียน
- Branden C, Tooze J (1999). บทนำสู่โครงสร้างโปรตีน . นิวยอร์ก: Garland Pub. ISBN 978-0-8153-2305-1-
- Murray RF, Harper HW, Granner DK, Mayes PA, Rodwell VW (2006). Harper's Illustrated Biochemistry . นิวยอร์ก: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9-
- Van Holde KE, Mathews CK (1996). ชีวเคมี. เมนโลพาร์ก, แคลิฟอร์เนีย: Benjamin/Cummings Pub. Co., Inc. ISBN 978-0-8053-3931-4-
- ประวัติศาสตร์
- Tanford C, Reynolds JA (2001). Nature's Robots: A History of Proteins . Oxford New York: Oxford University Press, USA. ISBN 978-0-19-850466-5-
ลิงค์ภายนอก
ฐานข้อมูลและโครงการ
- ฐานข้อมูลโปรตีน NCBI Entrez
- ฐานข้อมูลโครงสร้างโปรตีนของ NCBI
- ฐานข้อมูลอ้างอิงโปรตีนของมนุษย์
- ฮิวแมนโปรตีนพีเดีย
- Folding@Home (มหาวิทยาลัยสแตนฟอร์ด) เก็บถาวรเมื่อ 2012-09-08 ที่เวย์แบ็กแมชชีน
- ธนาคารข้อมูลโปรตีนในยุโรป (ดู PDBeQuips บทความสั้นๆ และบทช่วยสอนเกี่ยวกับโครงสร้าง PDB ที่น่าสนใจ)
- ความร่วมมือด้านการวิจัยสำหรับชีวสารสนเทศเชิงโครงสร้าง (ดูเพิ่มเติมที่ Molecule of the Month Archived 2020-07-24 ที่Wayback Machineซึ่งนำเสนอรายละเอียดสั้นๆ เกี่ยวกับโปรตีนที่เลือกจาก PDB)
- Proteopedia – ชีวิตในรูปแบบ 3 มิติ: โมเดล 3 มิติที่หมุนและซูมได้พร้อมคำอธิบายประกอบจาก wiki สำหรับโครงสร้างโมเลกุลโปรตีนที่รู้จักทุกชนิด
- UniProt แหล่งโปรตีนสากล
บทช่วยสอนและเว็บไซต์เพื่อการศึกษา
- "การแนะนำโปรตีน" จากHOPES (โครงการเผยแพร่ความรู้ด้านโรคฮันติงตันเพื่อการศึกษาที่มหาวิทยาลัยสแตนฟอร์ด)
- โปรตีน: การเกิดชีวภาพจนถึงการย่อยสลาย – ห้องสมุดเสมือนของชีวเคมีและชีววิทยาเซลล์







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
