อ่าน 14 นาที
การดัดแปลงหลังการแปล
การดัดแปลงหลังการแปล ( PTMs ) คือ กระบวนการ โควาเลนต์ที่เปลี่ยนแปลงโปรตีนหลังจากการสังเคราะห์และการปล่อยออกจากไรโบโซม PTMs...
การดัดแปลงหลังการแปล

การดัดแปลงหลังการแปล ( PTMs ) คือ กระบวนการ โควาเลนต์ที่เปลี่ยนแปลงโปรตีนหลังจากการสังเคราะห์และการปล่อยออกจากไรโบโซม PTMs เป็นเหตุการณ์การแก้ไขที่ย้อนกลับได้ซึ่งใช้และดำเนินการในกระบวนการโดยรวมของการควบคุมหลังการแปล – การควบคุมระดับของโปรตีนที่ออกฤทธิ์ เหตุการณ์ที่ไม่สามารถย้อนกลับได้คือโปรตีโอไลซิส(การย่อยสลายโปรตีน) PTMs ช่วยให้หน้าที่ของโปรตีนมีความหลากหลายและขยายออกไปนอกเหนือจากข้อกำหนดของการถอดรหัส [ 1 ]ณ ปี 2023 มี PTM ที่รู้จักมากกว่า 650 ชนิด[ 2 ] PTMs ยังเป็นกระบวนการของโปรคาริโอต อีกด้วย [ 3 ]
การดัดแปลงหลังการสังเคราะห์ โปรตีน (PTMs) อาจเกี่ยวข้องกับเอนไซม์หรือเกิดขึ้นเองโดยธรรมชาติ โปรตีนถูกสร้างขึ้นโดยไรโบโซม ซึ่งแปลmRNAเป็นสายโพลีเปปไทด์ จากนั้นสายโพลีเปป ไทด์อาจเปลี่ยนแปลงไปเป็นโปรตีนที่สมบูรณ์ แล้วจึงปล่อยออกจากไรโบโซม การดัดแปลงหลังการสังเคราะห์โปรตีนเป็นองค์ประกอบสำคัญในการ ส่งสัญญาณภายในเซลล์เช่น เมื่อโปรฮอร์โมนถูกเปลี่ยนเป็นฮอร์โมน
การดัดแปลงหลังการแปลสามารถเกิดขึ้นได้ที่โซ่ข้างของกรดอะมิโน หรือที่ ปลายC-หรือN-ของโปรตีน[ 4 ]พวกมันสามารถขยายชุดทางเคมีของกรดอะมิโน 22 ชนิดได้ โดยการเปลี่ยนหมู่ฟังก์ชัน ที่มีอยู่ หรือเพิ่มหมู่ฟังก์ชันใหม่ เช่น ฟอสเฟตการฟอสฟอริเลชันมีประสิทธิภาพสูงในการควบคุมกิจกรรมของเอนไซม์และเป็นการเปลี่ยนแปลงที่พบบ่อยที่สุดหลังการแปล[ 5 ] โปรตีน ยูคาริโอตและโปรคาริโอตจำนวนมากยังมี โมเลกุล คาร์โบไฮเดรตติดอยู่ด้วยในกระบวนการที่เรียกว่าไกลโคซิเลชันซึ่งสามารถส่งเสริม การพับตัว ของ โปรตีนและปรับปรุงความเสถียร ตลอดจนทำหน้าที่ควบคุม การติด โมเลกุล ไขมันที่เรียกว่าลิพิเดชันมักจะเกิดขึ้นกับโปรตีนหรือส่วนหนึ่งของโปรตีนที่ติดอยู่กับเยื่อหุ้มเซลล์
รูปแบบอื่นๆ ของการดัดแปลงหลังการแปลประกอบด้วยการตัดพันธะเปปไทด์เช่น การประมวลผลโปรเปปไทด์ให้เป็นรูปแบบที่สมบูรณ์หรือการกำจัด หมู่ เมไท โอนีนที่เป็นตัวเริ่มต้น การสร้างพันธะไดซัลไฟด์จาก หมู่ ซิสเทอีนก็อาจเรียกได้ว่าเป็นการดัดแปลงหลังการแปลเช่นกัน[ 6 ]ตัวอย่างเช่น ฮอร์โมนเปปไทด์อินซูลินถูกตัดสองครั้งหลังจากสร้างพันธะไดซัลไฟด์ และโปรเปปไทด์ถูกกำจัดออกจากตรงกลางของโซ่ โปรตีนที่ได้จะประกอบด้วยโซ่โพลีเปปไทด์สองโซ่ที่เชื่อมต่อกันด้วยพันธะไดซัลไฟด์
การดัดแปลงหลังการแปลบางประเภทเป็นผลมาจากความเครียดออกซิเดชันการคาร์บอนิเลชันเป็นตัวอย่างหนึ่งที่กำหนดเป้าหมายโปรตีนที่ถูกดัดแปลงเพื่อการย่อยสลายและอาจส่งผลให้เกิดการก่อตัวของโปรตีนรวมกลุ่ม[ 7 ] [ 8 ]การดัดแปลงกรดอะมิโนเฉพาะสามารถใช้เป็นตัวบ่งชี้ทางชีวภาพที่บ่งบอกถึงความเสียหายจากออกซิเดชัน[ 9 ] PTMs และไอออนโลหะมีบทบาทสำคัญและมีปฏิสัมพันธ์กันในการควบคุมการทำงานของโปรตีน ส่งผลต่อกระบวนการของเซลล์ เช่น การส่งสัญญาณและการแสดงออกของยีน โดยปฏิสัมพันธ์ที่ผิดปกติเกี่ยวข้องกับโรคต่างๆ เช่น มะเร็งและโรคความเสื่อมของระบบประสาท[ 10 ]
ตำแหน่งที่มักได้รับการดัดแปลงหลังการแปลรหัสคือตำแหน่งที่มีหมู่ฟังก์ชันที่สามารถทำหน้าที่เป็นนิวคลีโอไฟล์ในปฏิกิริยา ได้แก่ หมู่ ไฮดรอกซิลของซีรีนทรีโอนีนและไทโร ซีน รูป แบบ อะมีนของไลซีนอาร์จินีนและฮิ สติดีน แอนไอออนไทโอเลต ของซิส เทอี น คาร์บอกซิเลตของแอสปาร์เทตและกลูตาเมตและปลาย N- และ C-เทอร์มินัส นอกจากนี้ แม้ว่าอะไมด์ของแอสปาราจีนจะเป็นนิวคลีโอไฟล์ที่อ่อนแอ แต่ก็สามารถทำหน้าที่เป็นจุดยึดสำหรับไกลแคนได้ การดัดแปลงที่หายากกว่าอาจเกิดขึ้นที่ เมไทโอนีนที่ถูกออกซิไดซ์และที่หมู่เมทิลีน บางหมู่ ในโซ่ข้าง[ 11 ]
การดัดแปลงโปรตีนหลังการสังเคราะห์สามารถตรวจจับได้ด้วยวิธีการทดลองหลากหลายวิธี รวมถึงแมสสเปกโทรเมตรีอีสเทิร์นบลอตติงและเวสเทิร์นบลอตติง
การดัดแปลงหลังการสังเคราะห์ที่เกี่ยวข้องกับการเพิ่มหมู่ฟังก์ชัน
การเติมโดยเอนไซม์ในร่างกาย
กลุ่มไฮโดรโฟบิกสำหรับการกำหนดตำแหน่งในเยื่อหุ้มเซลล์
- ไมริสโตอิลเลชัน ( อะซิเลชันชนิดหนึ่ง) คือการเติมไมริสเตต ซึ่งเป็น กรดอิ่มตัวC14 เข้าไป
- การเติมหมู่ปาล์มิโตอิล (ซึ่งเป็นกระบวนการอะซิเลชันชนิดหนึ่ง) คือการเติมหมู่ปาล์มิเตต ซึ่ง เป็นกรดอิ่มตัวที่มีคาร์บอน16 อะตอม
- ไอโซพรีนิเลชันหรือพรีนิเลชันคือการเติม หมู่ ไอโซพรีนอยด์ (เช่นฟาร์เนซอลและเจอรานิลเจอรานิออล )
- การไกลโคซิเอชันคือ การสร้าง แองเคอร์ไกลโคซิลฟอสฟาติดิลอิโนซิทอล (GPI)ผ่านพันธะอะไมด์กับส่วนหาง C-เทอร์มินัล
โคแฟคเตอร์เพื่อเพิ่มประสิทธิภาพการทำงานของเอนไซม์
- ลิโปอิเลชัน (อะซิเลชันชนิดหนึ่ง) คือการเติม หมู่ฟังก์ชัน ลิโปเอต (C8 )เข้าไป
- หมู่ ฟลาวิน ( ฟลาวินโมโนนิวคลีโอไทด์ (FMN) หรือฟลาวินอะดีนีนไดนิวคลีโอไทด์ (FAD)) อาจถูกเชื่อมต่อด้วยพันธะโควาเลนต์
- การยึดเกาะของ ฮีมซีผ่าน พันธะ ไทโออีเทอร์กับซิสเทอีน
- ฟอสโฟแพนเททีนิเลชันคือการเติมหมู่ 4'-ฟอสโฟแพนเททีนิลจากโคเอนไซม์เอเช่น ในการสังเคราะห์กรดไขมัน โพลีคีไทด์ เปปไทด์ที่ไม่ใช่ไรโบโซม และลิวซีน
- การก่อตัวของฐานชิฟฟ์เรตินิลิ ดีน
การปรับเปลี่ยนปัจจัยการแปล
- การก่อตัว ของไดฟ์ทาไมด์ (บนฮิสติดีนที่พบในeEF2 )
- การยึดเกาะ เอทานอลอะมีนฟอสโฟกลีเซอรอล (บนกลูตาเมตที่พบในeEF1α ) [ 12 ]
- การสร้าง ไฮพูซีน (บนไลซีนที่คงสภาพของeIF5A (ยูคาริโอต) และaIF5A (อาร์เคีย))
- การเติม เบต้า-ไลซีนบนไลซีนที่อนุรักษ์ไว้ของปัจจัยการยืด P (EFP) ในแบคทีเรียส่วนใหญ่[ 13 ] EFP เป็นโฮโมล็อกของeIF5A (ยูคาริโอต) และaIF5A (อาร์เคีย) (ดูด้านบน)
กลุ่มเคมีขนาดเล็ก
- การเติมหมู่แอซิลเช่น หมู่แอซิลที่ตำแหน่ง O ( เอสเทอร์ ), หมู่แอซิลที่ตำแหน่ง N ( เอไมด์ ), หมู่แอซิลที่ตำแหน่ง S ( ไทโอเอสเทอร์ )
- การอะซิทิเลชันคือการเพิ่ม กลุ่ม อะซิทิลที่ปลาย Nของโปรตีนหรือที่หมู่ไลซีน[ 14 ]กระบวนการตรงกันข้ามเรียกว่าการดีอะซิทิเลชัน
- ฟอร์ไมเลชัน
- การเติม หมู่แอลคิล (Alkylation)คือการเติม หมู่ แอลคิลเช่นหมู่เมทิลหมู่เอทิล
- การเติม หมู่เมทิล (methylation)คือการเติม หมู่ เมทิล เข้าไป โดยปกติจะเกิดขึ้นที่ กรด อะมิโนไลซีนหรืออาร์จินีนส่วนกระบวนการตรงกันข้ามเรียกว่าการดีเมทิลเลชัน (demethylation )
- การเกิดอะมิเดชันที่ปลาย C เกิดจากการแยกตัวแบบออกซิเดชันของหมู่ไกลซีนที่ปลาย C [ 15 ]
- โมโนอะมินิเลชันการเพิ่มโมโนอะมีนให้กับ สารตกค้าง กลูตามีนผ่านทรานส์อะมิเดชัน[ 16 ]
- โดปามินิเลชันคือการเติมโดปามีนเข้ากับ หมู่ กลูตามีนผ่านกระบวนการทรานส์อะมิเดชัน
- ฮิสตามินิเลชันคือการเติมฮิสตามีนเข้ากับ หมู่ กลูตามีนผ่านกระบวนการทรานส์อะมิเดชัน
- เซโรโทนิเลชันคือการเติมเซโรโทนินเข้ากับ หมู่ กลูตามีนผ่านกระบวนการทรานส์อะมิเดชัน
- โมโนอะมินิเลชันการเพิ่มโมโนอะมีนให้กับ สารตกค้าง กลูตามีนผ่านทรานส์อะมิเดชัน[ 16 ]
- การสร้างพันธะ อะไมด์
- การเติม กรดอะมิโน
- การเติม อาร์จินีน (arginylation)เป็นกระบวนการเติมที่อาศัยtRNA เป็นตัวกลาง
- โพลีกลูตาไมเลชันคือการเชื่อมต่อแบบโควาเลนต์ของ สารตกค้าง ของกรดกลูตามิกกับปลาย N ของทูบูลินและโปรตีนอื่นๆ บางชนิด[ 17 ] (ดูทูบูลินโพลีกลูตาไมเลส )
- โพลีไกลซิเลชัน คือการเชื่อมต่อแบบโควาเลนต์ของ กรดอะมิโนไกลซีน 1 ถึงมากกว่า 40 โมเลกุล เข้ากับส่วนปลาย C-terminal ของทูบูลิน
- การเติม กรดอะมิโน
- บิวทิริเลชัน
- แกมมาคาร์บอกซิเลชันขึ้นอยู่กับวิตามินเค[ 18 ]
- การเติม หมู่ไกลโค ซิล (glycosylation) คือการเติมหมู่ ไกลโคซิลเข้าไป ใน กรดอะมิโนอาร์จินีนแอส พารา จีนซิสเทอีน ไฮดรอก ซีไลซีน เซ รีน ทรีโอ นีน ไทโรซีนหรือทริปโตเฟน ทำให้เกิดเป็นไกลโคโปรตีนซึ่งแตกต่างจาก การเติม หมู่ไกลเคชั่น (glycation)ซึ่งถือเป็นการเติมหมู่ของน้ำตาลโดยไม่ใช้เอนไซม์
- พอซิแมนโนซิเลชันคือการเติมไกลแคนอย่างง่าย ซึ่งส่วนใหญ่ประกอบด้วยแมนโนสและเอ็น-อะเซทิลกลูโคซามีน (GlcNAc) เข้ากับกรดอะมิโนแอสปาราจีน
- O -GlcNAcคือการเติม N -acetylglucosamine เข้ากับหมู่เซรินหรือทรีโอนีนในพันธะ β-ไกลโคไซด์
- โพลีไซอะลิเลชัน คือการเติมกรดโพลีไซอะลิก (PSA) เข้ากับโมเลกุลการยึดเกาะของเซลล์ประสาท (NCAM)
- ไฮดรอกซิเลชัน : การเติมอะตอมออกซิเจนเข้าไปที่โซ่ข้างของกรดอะมิโนโพรลีนหรือไลซีน
- การเติมไอโอดีน : การเติมอะตอมของไอโอดีนเข้าไปในวงแหวนอะโรมาติกของหมู่ไทโรซีน (เช่น ในไทโรโกลบูลิน )
- การเติมสารนิวคลีโอไทด์เช่น การเติม ADP-ไรโบซิเลชัน
- เพอร์ซัลฟิเดชันคือการเติมโมเลกุลกำมะถันลงบน หมู่ ไทออลของ กรดอะ มิโนซิสเทอี น
- การก่อตัว ของฟอสเฟตเอสเทอร์ ( O -linked) หรือฟอสโฟรามิเดต ( N -linked)
- การฟอสโฟรีเลชันคือการเติม หมู่ ฟอสเฟตโดยปกติจะเติมที่ซีรีน ทรีโอนีนและไทโรซีน ( แบบ O -linked) หรือฮิสติดีน ( แบบ N -linked)
- การเติม หมู่แอดีนิล (adenylylation)คือการเติม หมู่ แอดีนิลเข้าไปโดยปกติจะเติมที่ไทโรซีน ( O -linked) หรือฮิสติดีนและไลซีน ( N -linked)
- การเติมหมู่ยูริดิล (uridylylation) คือการเติมหมู่ยูริดิล (เช่นยูริดีนโมโนฟอสเฟต (UMP)) เข้าไปที่ไทโรซีนเป็นส่วนใหญ่
- โพรพิโอนิเลชัน
- การก่อตัวของไพโรกลูตาเมต
- เอส -กลูตาไธโอนิเลชัน
- เอส -ไนโตรซิเลชัน
- S-ซัลฟีนิเลชัน ( หรือเรียกอีกอย่างว่าS-ซัลฟีนิเลชัน) การเพิ่มโคเวเลนต์แบบย้อนกลับของอะตอมออกซิเจนหนึ่งอะตอมไปยัง กลุ่ม ไทออลของสารตกค้างซิสเทอีน[ 19 ]
- S-ซัลฟินิเลชัน ซึ่งโดยปกติแล้วเป็นการเติมโคเวเลนต์ที่ไม่สามารถย้อนกลับได้ของอะตอมออกซิเจนสองอะตอมไปยัง กลุ่ม ไทออลของสารตกค้างซิสเทอีน[ 19 ]
- S-ซัลโฟนิเลชัน ซึ่งโดยปกติแล้วเป็นการเติมโคเวเลนต์ที่ไม่สามารถย้อนกลับได้ของอะตอมออกซิเจนสามอะตอมไปยัง กลุ่ม ไทออลของ สารตกค้าง ซิสเทอีนส่งผลให้เกิดสารตกค้างกรดซิสเทอิก[ 19 ]
- การเติมหมู่ซัลเฟตเข้าไปในไทโรซีน(sulfation )
การดัดแปลงที่ไม่เกี่ยวข้องกับเอนไซม์ในร่างกาย
ตัวอย่างของ PTM ที่ไม่ใช่เอนไซม์ ได้แก่ ไกลเคชั่น ไกลโคออกซิเดชั่น ไนโตรซิเลชั่น ออกซิเดชั่น ซัคซิเนชั่น และลิโปออกซิเดชั่น[ 20 ]
- การคาร์บาไมเลชัน คือการเติมกรดไอโซไซยานิกที่ปลาย N ของโปรตีนหรือโซ่ข้างของไลซีน[ 21 ]
- กระบวนการคาร์บอนิเลชันคือการเติมคาร์บอนมอนอกไซด์เข้าไปในสารประกอบอินทรีย์/อนินทรีย์อื่นๆ
- กระบวนการไกลเคชั่นคือการเติมโมเลกุลน้ำตาลเข้าไปในโปรตีนโดยปราศจากการควบคุมของเอนไซม์
- กลูตาริเลชันคือ การเพิ่มกลุ่มกลูตาริลให้กับหมู่ไลซีน[ 22 ]
- มาโลนิเลชันคือการเพิ่มกลุ่มมาโลนิลให้กับสารตกค้างไลซีน[ 23 ]
- เมทิลมาโลนิเลชันคือการเพิ่มกลุ่มเมทิลมาโลนิลให้กับหมู่ไลซีน[ 24 ]
- การสร้าง พันธะไอโซเปปไทด์โดยธรรมชาติระหว่างไลซีนและกรดแอสปาร์ติกหรือแอสปาราจีน ดังที่พบในโปรตีนพื้นผิวหลายชนิดของแบคทีเรียแกรมบวก[ 25 ]
- ซัคซินิเลชันการเพิ่ม กลุ่ม ซัคซินิลให้กับไลซีน[ 26 ]
การเติมสารโดยไม่ใช้เอนไซม์ในหลอดทดลอง
- การไบโอติไนเลชัน : การเชื่อมต่อโมเลกุลไบโอตินเข้ากับโปรตีนด้วยพันธะโควาเลนต์โดยใช้สารเคมีไบโอติไนเลชัน ซึ่งโดยทั่วไปมีวัตถุประสงค์เพื่อติดฉลากโปรตีน
- การคาร์บาไมเลชัน: การเพิ่มกรดไอโซไซยานิกที่ปลาย N ของโปรตีนหรือโซ่ข้างของสารตกค้าง Lys หรือ Cys ซึ่งโดยทั่วไปเกิดจากการสัมผัสกับสารละลายยูเรีย[ 27 ]
- ออกซิเดชัน: การเติมอะตอมออกซิเจนหนึ่งอะตอมหรือมากกว่านั้นลงในหมู่ข้างเคียงที่ไวต่อปฏิกิริยา โดยเฉพาะอย่างยิ่งในหมู่เมไทโอนีน (Met), ทริปโตเฟน (Trp), ฮิสติดีน (His) หรือซิสทีน (Cys) การเกิด พันธะ ไดซัลไฟด์ระหว่างหมู่ซิสทีน
- การเพกิเลชัน : การเชื่อมต่อโพลีเอทิลีนไกลคอล (PEG) ด้วยพันธะโควาเลนต์โดยใช้สารเพกิเลชัน ซึ่งโดยทั่วไปจะเชื่อมต่อกับปลาย N-terminus หรือหมู่ข้างเคียงของกรดอะมิโนไลซีน การเพกิเลชันใช้เพื่อเพิ่มประสิทธิภาพของยาโปรตีน
การเชื่อมต่อกับโปรตีนหรือเปปไทด์อื่นๆ
- กระบวนการยูบิควิติเนชันคือ การเชื่อมต่อ แบบโควาเลนต์กับโปรตีนยูบิควิติน
- SUMOylationคือการเชื่อมต่อแบบโควาเลนต์กับโปรตีน SUMO (ตัวดัดแปลงที่เกี่ยวข้องกับยูบิควิตินขนาดเล็ก) [ 28 ]
- เนดดิเลชันคือการเชื่อมต่อแบบโควาเลนต์กับโปรตีนเน็ด
- ISGylation คือการเชื่อมโยงโควาเลนต์กับ โปรตีน ISG15 (ยีนกระตุ้นอินเตอร์เฟรอน 15) [ 29 ]
- พูพิเลชันคือการเชื่อมต่อแบบโควาเลนต์กับโปรตีนคล้ายยูบิควิตินของโปรคาริโอต
การดัดแปลงทางเคมีของกรดอะมิโน
- ซิทรูลลิเนชันหรือ ดีอิมิเนชัน คือ การแปลงอาร์จินีนเป็นซิทรูลลีน[ 30 ]
- การดีแอมิเดชันคือการเปลี่ยนกลูตามีนเป็นกรดกลูตามิกหรือแอสพาราจีนเป็นกรดแอสปาร์ติก
- การกำจัดการแปลงเป็นแอลคีนโดยการกำจัดเบต้าของฟอสโฟทรีโอนีนและฟอสโฟเซอรีนหรือการกำจัดน้ำของทรีโอนีนและเซอรีน[ 31 ]
การเปลี่ยนแปลงโครงสร้าง
- พันธะไดซัลไฟด์คือพันธะโควาเลนต์ระหว่างกรดอะมิโนซิสเทอีน สอง โมเลกุล
- สะพานไลซีน-ซิสเทอีนการเชื่อมต่อแบบโควาเลนต์ของไลซีน 1 ตัวและซิสเทอีน 1 หรือ 2 ตัวผ่านอะตอมออกซิเจน (สะพาน NOS และ SONOS) [ 32 ]
- การแตกตัวของโปรตีนด้วยเอนไซม์โปรตีเอสคือการแตกตัวของโปรตีนที่พันธะเปปไทด์
- การสร้าง ไอโซแอสปาร์เทตผ่านการสร้างวงแหวนของกรดอะมิโนแอสพาราจีนหรือกรดแอสปาร์ติก
- การเกิดราซีไมเซชัน
- ของซีรีนโดยโปรตีน-ซีรีนอีพิเมอเรส
- ของอะลานีนในเดอร์มอร์ฟิน ซึ่งเป็น เปปไทด์โอปิออยด์จากกบ
- ของเมไทโอนีนในเดลทอร์ฟินซึ่งเป็นเปปไทด์โอปิออยด์ของกบเช่นกัน
- การตัดต่อโปรตีน การกำจัด อินเทอินแบบเร่งปฏิกิริยาด้วยตนเองซึ่งคล้ายคลึงกับการประมวลผล mRNA
สถิติ
PTM ทั่วไปตามความถี่
ในปี 2554 สถิติของการดัดแปลงหลังการแปลแต่ละครั้งที่ตรวจพบจากการทดลองและที่คาดการณ์ไว้ได้ถูกรวบรวมโดยใช้ข้อมูลทั่วทั้งโปรตีโอมจากฐานข้อมูล Swiss-Prot [ 33 ]การดัดแปลงที่พบจากการทดลองที่พบบ่อยที่สุด 10 อันดับแรกมีดังนี้: [ 34 ]
| ความถี่ | การแก้ไข |
|---|---|
| 58383 | การฟอสฟอริเลชัน |
| 6751 | อะเซทิเลชัน |
| 5526 | ไกลโคซิเลชันแบบ N-linked |
| 2844 | อามิเดชั่น |
| 1619 | ไฮดรอกซิเลชัน |
| 1523 | เมทิลเลชัน |
| 1133 | ไกลโคซิเลชันแบบ O-linked |
| 878 | การยูบิควิตินิเลชัน |
| 826 | กรดไพร์โรลิโดนคาร์บอกซิลิก |
| 504 | การซัลเฟต |
การดัดแปลงหลังการสังเคราะห์โปรตีนทั่วไปตามสารตกค้าง
ตัวอย่างการดัดแปลงหลังการสังเคราะห์โปรตีนที่พบได้ทั่วไปในกรดอะมิโนบางชนิดแสดงไว้ด้านล่าง การดัดแปลงเกิดขึ้นที่หมู่ข้างเคียงเว้นแต่จะระบุไว้เป็นอย่างอื่น
| กรดอะมิโน | อักษรย่อ. | การแก้ไข |
|---|---|---|
| อะลานีน | อะลา หรือ เอ | การเติมหมู่แอเซทิลที่ปลาย N (N-terminus) |
| อาร์จินีน | อาร์ก หรือ อาร์ | การดีอิมิเนชันเป็นซิทรูลีน , การเมทิลเลชัน |
| แอสพาราจีน | แอสน หรือ เอ็น | การดีแอมิเดชันเป็นแอสปาร์ติกแอซิด (Asp) หรือไอโซ(แอสปาร์ติกแอซิด) (iso(Asp)), การ ไกลโคซิเลชันแบบ N-linked , การสร้างพันธะไอโซเปปไทด์โดยธรรมชาติ |
| กรดแอสปาร์ติก | แอสป์ หรือ ดี | การไอโซเมอไรเซชันไปเป็นกรดไอโซแอสปาร์ติก การสร้างพันธะไอโซเปปไทด์โดยธรรมชาติ |
| ซิสเทอีน | ซิสทีนหรือซี | การสร้างพันธะ ไดซัลไฟด์ , การออกซิเดชันเป็นกรดซัลฟีนิก, กรดซัลฟินิก หรือกรดซัลโฟนิก, การพาลมิโทอิลเลชัน , การอะเซทิลเลชันที่ปลาย N (N-terminus), การไนโตรซิเลชันที่ปลาย S |
| กลูตามีน | กลูตาเมตหรือคิว | การเกิดวงแหวนเป็นกรดไพโรกลูตามิก (ปลาย N-terminus) การกำจัดหมู่เอมีนเป็นกรดกลูตามิกหรือ การสร้าง พันธะไอโซเปปไทด์กับไลซีนโดยทรานส์กลูตามิเนส |
| กรดกลูตามิก | กาวหรืออี | การเกิดวงแหวนเป็นกรดไพโรกลูตามิก (ปลาย N-terminus) และการเติมหมู่แกมมาคาร์บอกซิล |
| ไกลซีน | ไกล หรือ จี | การเติมหมู่ไมริสโตอิลที่ปลาย N (N-terminus), การเติมหมู่แอซีทิลที่ปลาย N (N-terminus) |
| ฮิสติดีน | ของเขาหรือ H | การฟอสโฟรีเลชัน |
| ไอโซลิวซีน | อิเล หรือ ไอ | |
| ลิวซีน | ลิว หรือ แอล | |
| ไลซีน | ไลส์ หรือ เค | การอะเซทิเลชัน , การยูบิควิทิเลชัน , การซูโมอิเลชัน , การเมทิเลชัน , การไฮดรอกซิเลชันที่นำไปสู่แอลลิซีน , การสร้างพันธะไอโซเปปไทด์โดยธรรมชาติ |
| เมไทโอนีน | เมโทรหรือเอ็ม | การเติมหมู่แอเซทิลที่ปลาย N (N-terminus), การเติมหมู่ยูบิควิตินที่ปลาย N (N-linked ubiquitination), การออกซิเดชันเป็นซัลฟอกไซด์หรือซัลโฟน |
| ฟีนิลอะลานีน | ฟีนิลอะลานีนหรือเอฟ | |
| โปรไลน์ | โปร หรือ พี | ไฮดรอกซิเลชัน |
| เซรีน | เซอร์ หรือ เอส | การฟอสโฟรีเลชัน , การไกลโคซิเลชันแบบ O-linked , การอะเซทิเลชันแบบ N-terminus (N-terminus) |
| ทรีโอนีน | ทรอหรือที | การฟอสโฟรีเลชัน, การไกลโคซิเลชันแบบ O-linked, การอะเซทิเลชันแบบ N-terminus (N-terminus) |
| ทริปโตแฟน | ทริป หรือ ดับเบิลยู | โมโนหรือไดออกซิเดชัน การก่อตัวของคีนูเรนีนทริปโตแฟน ทริปโตฟิลควิโนน |
| ไทโรซีน | ไทร์ หรือ วาย | การซัลเฟต , การฟอสโฟรีเลชัน |
| วาลีน | วาล หรือ วี | การเติมหมู่แอเซทิลที่ปลาย N (N-terminus) |
ฐานข้อมูลและเครื่องมือ
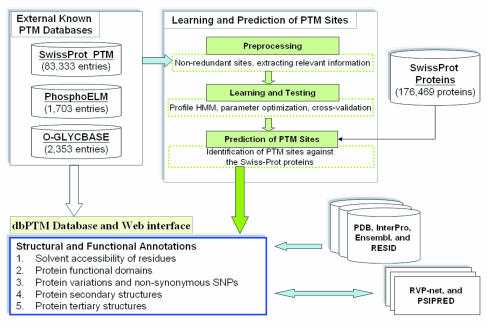
ลำดับโปรตีนประกอบด้วยลวดลายลำดับที่เอนไซม์ดัดแปลงจดจำได้ และสามารถบันทึกหรือทำนายได้ในฐานข้อมูล PTM (Post-Transformation Technology) เนื่องจากมีการค้นพบการดัดแปลงที่แตกต่างกันจำนวนมาก จึงมีความจำเป็นต้องบันทึกข้อมูลประเภทนี้ไว้ในฐานข้อมูล ข้อมูล PTM สามารถรวบรวมได้จากวิธีการทดลองหรือทำนายได้จากข้อมูลคุณภาพสูงที่ได้รับการตรวจสอบและคัดกรองด้วยตนเอง มีการสร้างฐานข้อมูลจำนวนมาก โดยมักเน้นไปที่กลุ่มอนุกรมวิธานบางกลุ่ม (เช่น โปรตีนของมนุษย์) หรือคุณลักษณะอื่นๆ
รายชื่อแหล่งข้อมูล
- PhosphoSitePlus [ 36 ] – ฐานข้อมูลข้อมูลและเครื่องมือที่ครอบคลุมสำหรับการศึกษาการดัดแปลงหลังการแปลโปรตีนของสัตว์เลี้ยงลูกด้วยนม
- ProteomeScout [ 37 ] – ฐานข้อมูลโปรตีนและการดัดแปลงหลังการแปลรหัสแบบทดลอง
- ฐานข้อมูลอ้างอิงโปรตีนของมนุษย์[ 37 ] – ฐานข้อมูลสำหรับการดัดแปลงที่แตกต่างกันและทำความเข้าใจโปรตีนที่แตกต่างกัน คลาส และหน้าที่/กระบวนการที่เกี่ยวข้องกับโปรตีนที่ก่อให้เกิดโรค
- PROSITE [ 38 ] – ฐานข้อมูลของรูปแบบฉันทามติสำหรับ PTM หลายประเภท รวมถึงไซต์
- RESID [ 39 ] – ฐานข้อมูลที่ประกอบด้วยชุดคำอธิบายประกอบและโครงสร้างสำหรับ PTM
- iPTMnet [ 40 ] – ฐานข้อมูลที่รวมข้อมูล PTM จากฐานความรู้หลายแห่งและผลลัพธ์การขุดข้อความ
- dbPTM [ 35 ] – ฐานข้อมูลที่แสดง PTM ต่างๆ และข้อมูลเกี่ยวกับองค์ประกอบทางเคมี/โครงสร้าง และความถี่สำหรับตำแหน่งที่มีการดัดแปลงกรดอะมิโน
- Uniprotมีข้อมูลเกี่ยวกับ PTM แม้ว่าข้อมูลอาจจะไม่ครบถ้วนเท่ากับฐานข้อมูลเฉพาะทางอื่นๆ

ผลของ PTM ต่อการทำงานของโปรตีนและกระบวนการทางสรีรวิทยา[ 41 ] - ฐาน ข้อมูล O -GlcNAc [ 42 ] [ 43 ] - ฐานข้อมูลที่รวบรวมไว้สำหรับการเติมหมู่ O-GlcNAc ให้กับโปรตีนและอ้างอิงถึงรายการโปรตีนมากกว่า 14,000 รายการและไซต์O -GlcNAc มากกว่า 10,000 ไซต์

เครื่องมือ
รายชื่อซอฟต์แวร์สำหรับแสดงภาพโปรตีนและการดัดแปลงหลังการสังเคราะห์ (PTMs)
- PyMOL [ 44 ] – แนะนำชุด PTM ทั่วไปลงในแบบจำลองโปรตีน
- สุดยอด[ 45 ] – เครื่องมือแบบโต้ตอบเพื่อดูบทบาทของโพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยวต่อ PTM
- ไคเมรา[ 46 ] – ฐานข้อมูลเชิงโต้ตอบเพื่อแสดงภาพโมเลกุล
ตัวอย่างกรณีศึกษา
- การแตกตัวและการสร้างพันธะไดซัลไฟด์ระหว่างการผลิตอินซูลิน
- การดัดแปลง โปรตีนฮิสโตน หลังการสังเคราะห์ (PTM) เป็นกลไกควบคุมการถอดรหัส : การควบคุมเอนไซม์ RNA polymerase โดยโครงสร้างโครมาติน
- การดัดแปลงหลังการสังเคราะห์โปรตีน (PTM) ของRNA polymerase IIในการควบคุมการถอดรหัส
- การแตกตัวของโซ่โพลีเปปไทด์มีความสำคัญต่อความจำเพาะของเลคติน[ 47 ]
- อิทธิพลของ Ni(II) ในการอะซิทิเลชันของโปรตีนฮิสโตน H4 [ 10 ]
ดูเพิ่มเติม
ลิงก์ภายนอก
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การดัดแปลงหลังการแปล
การดัดแปลงหลังการแปล ( PTMs ) คือ กระบวนการ โควาเลนต์ที่เปลี่ยนแปลงโปรตีนหลังจากการสังเคราะห์และการปล่อยออกจากไรโบโซม PTMs...
การเติมโดยเอนไซม์ ในร่างกาย
กลุ่มไฮโดรโฟบิกสำหรับการกำหนดตำแหน่งในเยื่อหุ้มเซลล์ ไมริสโตอิลเลชัน ( อะซิเลชัน ชนิดหนึ่ง) คือการเติม ไมริสเตต ซึ่งเป็น กรดอิ่มตัวC14 เข้าไป การเติมหมู่ปาล์มิโตอิล (ซึ่งเป็นกระบวนการอะซิเลชันชนิดหนึ่ง) คือการเติม หมู่ปาล์มิเตต ซึ่ง เป็นกรดอิ่มตัวที่มีคาร์บอน...
การดัดแปลงที่ไม่เกี่ยวข้องกับเอนไซม์ ในร่างกาย
ตัวอย่างของ PTM ที่ไม่ใช่เอนไซม์ ได้แก่ ไกลเคชั่น ไกลโคออกซิเดชั่น ไนโตรซิเลชั่น ออกซิเดชั่น ซัคซิเนชั่น และลิโปออกซิเดชั่น [ 20 ]
การเติมสารโดยไม่ใช้เอนไซม์ ในหลอดทดลอง
การไบโอติไนเลชัน : การเชื่อมต่อโมเลกุลไบโอตินเข้ากับโปรตีนด้วยพันธะโควาเลนต์โดยใช้สารเคมีไบโอติไนเลชัน ซึ่งโดยทั่วไปมีวัตถุประสงค์เพื่อติดฉลากโปรตีน การคาร์บาไมเลชัน: การเพิ่มกรดไอโซไซยานิกที่ปลาย N ของโปรตีนหรือโซ่ข้างของสารตกค้าง Lys หรือ Cys...