อ่าน 30 นาที
โปรตีน
โปรตีน เป็น โมเลกุลชีวภาพ ขนาดใหญ่ และ โมเลกุลขนาดมหึมา ที่ประกอบด้วยสายโซ่ยาวของกรดอะมิโนหนึ่งสายหรือมากกว่า โปรตีน ทำ หน้าที่มากมายในสิ่งมีชีวิต รวมถึง การเร่งปฏิกิริยาทางเม...
โปรตีน
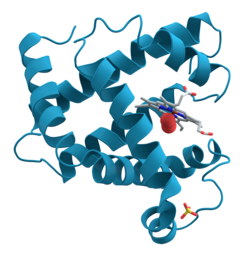
โปรตีนเป็นโมเลกุลชีวภาพ ขนาดใหญ่ และโมเลกุลขนาดมหึมาที่ประกอบด้วยสายโซ่ยาวของกรดอะมิโนหนึ่งสายหรือมากกว่าโปรตีนทำ หน้าที่มากมายในสิ่งมีชีวิต รวมถึงการเร่งปฏิกิริยาทางเม ตาบอลิ ซึมการจำลองดีเอ็นเอการตอบสนองต่อสิ่งเร้าการให้โครงสร้างแก่เซลล์และสิ่งมีชีวิตและการขนส่งโมเลกุลจากที่หนึ่งไปยังอีกที่หนึ่ง โปรตีนแตกต่างกันหลักๆ ในลำดับของกรดอะมิโน ซึ่งถูกกำหนดโดยลำดับนิวคลีโอไทด์ของยีนและโดยปกติจะส่งผลให้โปรตีนพับตัวเป็นโครงสร้างสามมิติ เฉพาะ ที่กำหนดกิจกรรมของมัน
สายโซ่เชิงเส้นของกรดอะมิโนเรียกว่าพอลิเปปไทด์โปรตีนประกอบด้วยพอลิเปปไทด์สายยาวอย่างน้อยหนึ่งสาย พอลิเปปไทด์สายสั้นที่มีกรดอะมิโนน้อยกว่า 20-30 ตัว มักไม่ถือว่าเป็นโปรตีนและเรียกว่าเปปไทด์ กรดอะมิโนแต่ละตัวเชื่อมต่อกันด้วยพันธะเปปไทด์และพันธะระหว่างกรดอะมิโนที่อยู่ติดกันลำดับของกรดอะมิโนในโปรตีนถูกกำหนดโดยลำดับของยีน ซึ่งถูกเข้ารหัสอยู่ในรหัสพันธุกรรมโดยทั่วไป รหัสพันธุกรรมจะระบุกรดอะมิโนมาตรฐาน 20 ชนิด แต่ในสิ่งมีชีวิตบางชนิด รหัสพันธุกรรมอาจรวมถึงซีลีโนซิสเทอีนและในอาร์เคียบาง ชนิด อาจรวมถึงไพโร ไลซีนด้วย ไม่นานหลังจากหรือแม้กระทั่งในระหว่างการสังเคราะห์ กรดอะมิโนในโปรตีนมักถูกดัดแปลงทางเคมีโดยการดัดแปลงหลังการแปลซึ่งจะเปลี่ยนแปลงคุณสมบัติทางกายภาพและเคมี การพับตัว ความเสถียร กิจกรรม และท้ายที่สุดคือหน้าที่ของโปรตีน โปรตีนบางชนิดมีกลุ่มที่ไม่ใช่เปปไทด์ติดอยู่ ซึ่งอาจเรียกว่ากลุ่มโปรสเตติกหรือโคแฟคเตอร์ โปรตีนสามารถทำงานร่วมกันเพื่อบรรลุหน้าที่เฉพาะอย่าง และมักจะรวมตัวกันเพื่อสร้างโปรตีนเชิงซ้อนที่ มีเสถียรภาพ
เมื่อโปรตีนถูกสร้างขึ้นแล้ว โปรตีนจะมีอายุอยู่ได้เพียงช่วงเวลาหนึ่งเท่านั้น จากนั้นก็จะถูกย่อยสลายและนำกลับมาใช้ใหม่โดยกลไกของเซลล์ผ่านกระบวนการหมุนเวียนโปรตีนอายุขัยของโปรตีนวัดได้จากครึ่งชีวิตซึ่งมีช่วงกว้างมาก โปรตีนอาจอยู่ได้นานเพียงไม่กี่นาทีหรือหลายปี โดยมีอายุขัยเฉลี่ย 1-2 วันในเซลล์สัตว์เลี้ยงลูกด้วยนม โปรตีนที่ผิดปกติหรือพับตัวผิดรูปจะถูกย่อยสลายได้เร็วกว่า โดยส่วนใหญ่มักทำโดยโปรตีเอโซมซึ่งเป็นกลุ่มโปรตีนขนาดใหญ่ โปรตีนเหล่านี้จะถูกย่อยสลายเนื่องจากถูกกำหนดเป้าหมาย ( ยูบิควิตินไลเกสสามารถทำเครื่องหมายโปรตีนเพื่อทำลายได้) หรือเนื่องจากไม่เสถียรหรือเสียหาย
เช่นเดียวกับโมเลกุลชีวภาพขนาดใหญ่อื่นๆ เช่นโพลีแซ็กคาไรด์และกรดนิวคลีอิกโปรตีนเป็นส่วนประกอบที่สำคัญของสิ่งมีชีวิตและมีส่วนร่วมในกระบวนการเกือบทุกอย่างภายในเซลล์โปรตีนหลายชนิดเป็นเอนไซม์ที่เร่ง ปฏิกิริยาทาง ชีวเคมีและมีความสำคัญต่อ กระบวนการเผา ผลาญโปรตีนบางชนิดมีหน้าที่เชิงโครงสร้างหรือเชิงกล เช่นแอคตินและไมโอซินในกล้ามเนื้อ และโปรตีนโครงสร้างของโครงร่างเซลล์ที่ช่วยรักษารูปร่างของเซลล์ โปรตีนอื่นๆ มีความสำคัญในการส่งสัญญาณของเซลล์การตอบสนองทางภูมิคุ้มกันการยึดเกาะของเซลล์และวงจรเซลล์ในสัตว์ โปรตีนจำเป็นต้องได้รับจากอาหารเพื่อจัดหาอะมิโนแอซิดที่จำเป็นซึ่งร่างกายไม่สามารถสังเคราะห์ได้เองการย่อยอาหารจะย่อยสลายโปรตีนเพื่อนำไปใช้ในกระบวนการเผาผลาญ
ประวัติศาสตร์และรากศัพท์
การค้นพบและการศึกษาเบื้องต้น
โปรตีนได้รับการศึกษาและรู้จักมาตั้งแต่ศตวรรษที่ 17 โดยAntoine Fourcroyและคนอื่นๆ[ 1 ] [ 2 ]ซึ่งมักเรียกโดยรวมว่า " อัลบูมิน " หรือ "สารอัลบูมิน" ( Eiweisskörperในภาษาเยอรมัน) [ 2 ] ตัวอย่างเช่นกลูเตน ถูกแยกออกจากข้าวสาลีเป็นครั้งแรกในการวิจัยที่ตีพิมพ์ราวปี 1747 และต่อมาพบว่ามีอยู่ในพืชหลายชนิด [ 1 ]ในปี 1789 Antoine Fourcroy ได้จำแนกโปรตีนจากสัตว์ออกเป็น 3 ชนิดที่แตกต่างกัน ได้แก่อัลบูมินไฟบรินและเจลาติน [ 3 ] โปรตีนจากพืชที่ศึกษาในช่วงปลายศตวรรษที่ 17 และต้นศตวรรษที่ 18 ได้แก่กลูเตน อัลบูมิ นจากพืช กลีอะดินและเลกู มิ น[ 1 ]
โปรตีนได้รับการอธิบายครั้งแรกโดยนักเคมีชาวดัตช์Gerardus Johannes Mulderและตั้งชื่อโดยนักเคมีชาวสวีเดนJöns Jacob Berzeliusในปี 1838 [ 4 ] [ 5 ] Mulder ได้ทำการวิเคราะห์ธาตุ ของโปรตีนทั่วไปและพบว่าโปรตีนเกือบทั้งหมดมี สูตรเชิงประจักษ์เดียวกันคือ C 400 H 620 N 100 O 120 P 1 S 1 [ 6 ]เขาได้ข้อสรุปที่ผิดพลาดว่าโปรตีนอาจประกอบด้วยโมเลกุลชนิดเดียว (ขนาดใหญ่มาก) คำว่า "โปรตีน" ที่ใช้อธิบายโมเลกุลเหล่านี้ได้รับการเสนอโดย Berzelius ผู้ร่วมงานของ Mulder คำว่าโปรตีนมาจากคำภาษากรีกπρώτειος ( proteios )ซึ่งหมายถึง "หลัก" [ 7 ] "นำหน้า" หรือ "ยืนอยู่ข้างหน้า" [ 2 ] + -inมัลเดอร์ได้ทำการระบุผลิตภัณฑ์จากการสลายตัวของโปรตีน เช่นกรดอะมิโนลิวซีนซึ่งเขาพบว่ามีน้ำหนักโมเลกุล (เกือบถูกต้อง) เท่ากับ131 ดา . [ 6 ]
นักวิทยาศาสตร์ด้านโภชนาการยุคแรก เช่นคาร์ล ฟอน วอยต์ ชาวเยอรมัน เชื่อว่าโปรตีนเป็นสารอาหารที่สำคัญที่สุดในการรักษาสภาพโครงสร้างของร่างกาย เนื่องจากโดยทั่วไปเชื่อกันว่า "เนื้อสร้างเนื้อ" [ 8 ]ประมาณปี 1862 คา ร์ล ไฮน์ริช ริทเฮา เซน ได้แยกกรดอะมิโนกลูตามิกออก มา [ 9 ]โทมัส เบอร์ ออสบอร์นได้รวบรวมบทวิจารณ์โดยละเอียดเกี่ยวกับโปรตีนจากพืชที่สถานีทดลองทางการเกษตรคอนเนตทิคัต ออสบอร์น ร่วมกับลาฟาแยตต์ เมนเดลได้กำหนดกรดอะมิโนที่จำเป็นต่อโภชนาการ หลายชนิด ในการทดลองให้อาหารกับหนูทดลอง[ 10 ]อาหารที่ขาดกรดอะมิโนที่จำเป็นจะทำให้การเจริญเติบโตของหนูชะงักงัน ซึ่งสอดคล้องกับกฎขั้นต่ำของไลบิก [ 11 ] กรดอะมิโนที่จำเป็นตัวสุดท้ายที่ถูกค้นพบ คือ ทรีโอนีนซึ่งระบุโดยวิลเลียม คัมมิงโรส[ 12 ]
ความยากลำบากในการทำให้โปรตีนบริสุทธิ์เป็นอุปสรรคต่องานของนักชีวเคมีโปรตีนในยุคแรก โปรตีนสามารถหาได้ในปริมาณมากจากเลือด ไข่ขาว และเคราตินแต่โปรตีนแต่ละชนิดหาได้ยาก ในช่วงทศวรรษ 1950 บริษัท Armour Hot Dog ได้ทำให้ไร โบเอนไซม์ Aจากตับอ่อนของวัวบริสุทธิ์ 1 กิโลกรัมและแจกจ่ายให้แก่นักวิทยาศาสตร์โดยไม่คิดค่าใช้จ่าย การกระทำนี้ช่วยให้ไรโบเอนไซม์ A กลายเป็นเป้าหมายหลักในการศึกษาทางชีวเคมีในทศวรรษต่อมา[ 6 ]
โพลีเปปไทด์

ความเข้าใจเกี่ยวกับโปรตีนในฐานะพอลิเปปไทด์หรือสายโซ่ของกรดอะมิโน เกิดขึ้นจากผลงานของFranz HofmeisterและHermann Emil Fischerในปี พ.ศ. 2445 [ 13 ] [ 14 ]บทบาทสำคัญของโปรตีนในฐานะเอนไซม์ในสิ่งมีชีวิตที่เร่งปฏิกิริยายังไม่เป็นที่เข้าใจอย่างถ่องแท้จนกระทั่งปี พ.ศ. 2469 เมื่อJames B. Sumnerแสดงให้เห็นว่าเอนไซม์ยูรีเอสเป็นโปรตีน[ 15 ]
Linus Paulingได้รับการยกย่องว่าสามารถทำนายโครงสร้างทุติยภูมิ ของโปรตีนปกติได้อย่างแม่นยำ โดยอาศัยพันธะไฮโดรเจน ซึ่ง เป็น แนวคิดที่ William Astburyเสนอเป็นครั้งแรกในปี พ.ศ. 2476 [ 16 ]ต่อมางานของWalter Kauzmannเกี่ยวกับการเสียสภาพ [ 17 ] [ 18 ]ซึ่งอิงจากงานวิจัยก่อนหน้านี้ของKaj Linderstrøm-Lang บางส่วน [ 19 ]มีส่วนช่วยให้เข้าใจการพับและโครงสร้างของโปรตีนที่เกิดจากการโต้ตอบแบบไฮโดรโฟบิก[ 20 ]
โปรตีนตัวแรกที่มี การจัดลำดับกรดอะมิโนคืออินซูลินโดยเฟรเดอริก แซงเกอร์ในปี 1949 แซงเกอร์ได้กำหนดลำดับกรดอะมิโนของอินซูลินได้อย่างถูกต้อง จึงแสดงให้เห็นอย่างชัดเจนว่าโปรตีนประกอบด้วยพอลิเมอร์เชิงเส้นของกรดอะมิโน ไม่ใช่โซ่กิ่งคอลลอยด์หรือไซโคล [ 21 ] เขาได้รับรางวัลโนเบลจากความสำเร็จนี้ในปี 1958 [ 22 ] การศึกษาของคริสเตียน แอนฟินเซน เกี่ยวกับกระบวนการ พับตัวแบบออกซิเดชันของไรโบเอนไซม์เอ ซึ่งทำให้เขาได้รับรางวัลโนเบลในปี 1972 ได้ยืนยันสมมติฐานทางเทอร์โมไดนามิกของการพับตัวของโปรตีน ซึ่งระบุว่ารูปแบบที่พับตัวของโปรตีนแสดงถึงพลังงานอิสระต่ำสุด[ 23 ] [ 24 ]
โครงสร้าง

ด้วยการพัฒนาของผลึกศาสตร์รังสีเอกซ์ทำให้สามารถกำหนดโครงสร้างโปรตีนรวมถึงลำดับของโปรตีนได้[ 25 ]โครงสร้างโปรตีนแรกที่ได้รับการแก้ไขคือฮีโมโกลบินโดยMax PerutzและไมโอโกลบินโดยJohn Kendrewในปี 1958 [ 26 ] [ 27 ]การใช้คอมพิวเตอร์และกำลังการคำนวณที่เพิ่มขึ้นได้สนับสนุนการจัดลำดับโปรตีนที่ซับซ้อน ในปี 1999 Roger Kornbergได้จัดลำดับโครงสร้างที่ซับซ้อนมากของRNA polymeraseโดยใช้รังสีเอกซ์ความเข้มสูงจากซินโครตรอน[ 25 ]
ตั้งแต่นั้นมากล้องจุลทรรศน์อิเล็กตรอนแบบแช่แข็ง (cryo-EM) ของโครงสร้างโมเลกุล ขนาดใหญ่ [ 28 ]ได้รับการพัฒนาขึ้น Cryo-EM ใช้ตัวอย่างโปรตีนที่แช่แข็งแทนผลึก และใช้ลำแสงอิเล็กตรอนแทนรังสีเอกซ์ ซึ่งทำให้เกิดความเสียหายต่อตัวอย่างน้อยลง ช่วยให้นักวิทยาศาสตร์ได้รับข้อมูลมากขึ้นและวิเคราะห์โครงสร้างขนาดใหญ่ขึ้นได้[ 25 ]การทำนายโครงสร้างโปรตีนด้วยคอมพิวเตอร์ของโดเมนโครงสร้าง โปรตีนขนาดเล็ก [ 29 ]ช่วยให้นักวิจัยเข้าใกล้ความละเอียดระดับอะตอมของโครงสร้างโปรตีนได้ ณ เดือนเมษายน 2024 ฐานข้อมูลโปรตีนมีโครงสร้างโปรตีนจากรังสีเอกซ์ 181,018 โครงสร้าง จากกล้องจุลทรรศน์อิเล็กตรอน 19,809 โครงสร้าง และ จาก NMR 12,697 โครงสร้าง[ 30 ]
การจำแนกประเภท
โปรตีนส่วนใหญ่จะถูกจำแนกตามลำดับและโครงสร้าง แม้ว่าจะมีการใช้การจำแนกประเภทอื่นๆ ทั่วไปก็ตาม โดยเฉพาะอย่างยิ่งสำหรับเอนไซม์ ระบบหมายเลข EC จะให้แผนการจำแนกประเภทตามหน้าที่[ 31 ]ในทำนองเดียวกันออนโทโลยีของยีนจะจำแนกทั้งยีนและโปรตีนตามหน้าที่ทางชีววิทยาและชีวเคมี และตามตำแหน่งภายในเซลล์[ 32 ]
ความคล้ายคลึงของลำดับถูกนำมาใช้ในการจำแนกโปรตีนทั้งในแง่ของความคล้ายคลึงทางวิวัฒนาการและหน้าที่ ซึ่งอาจใช้โปรตีนทั้งหมดหรือโดเมนของโปรตีนโดยเฉพาะอย่างยิ่งในโปรตีนที่มีหลายโดเมนโดเมนของโปรตีนช่วยให้สามารถจำแนกโปรตีนได้โดยการรวมกันของลำดับ โครงสร้าง และหน้าที่ และสามารถรวมกันได้หลายวิธี ในการศึกษาเบื้องต้นของโปรตีน 170,000 ชนิด พบว่าประมาณสองในสามได้รับการกำหนดอย่างน้อยหนึ่งโดเมน โดยโปรตีนขนาดใหญ่จะมีโดเมนมากกว่า (เช่น โปรตีนที่มีขนาดใหญ่กว่า 600 กรดอะมิโนจะมีโดเมนโดยเฉลี่ยมากกว่า 5 โดเมน) [ 33 ]
ชีวเคมี


โปรตีนส่วนใหญ่ประกอบด้วยพอลิเมอร์ เชิงเส้นที่สร้างขึ้นจากกรดอะมิโน L -αมากถึง 20 โมเลกุลกรดอะมิโนที่สร้างโปรตีนทั้งหมดมีโครงสร้างทั่วไปที่คาร์บอนอัลฟาเชื่อมต่อกับหมู่เอมีน หมู่ คาร์บอกซิลและโซ่ข้าง ที่แปรผันได้ มี เพียงโพรลีน เท่านั้น ที่แตกต่างจากโครงสร้างพื้นฐานนี้ เนื่องจากโซ่ข้างของโพรลีนเป็นวงแหวน เชื่อมต่อกับหมู่เอมีน ซึ่งจำกัดความยืดหยุ่นของโซ่โปรตีน[ 34 ]โซ่ข้างของกรดอะมิโนมาตรฐานมีโครงสร้างทางเคมีและคุณสมบัติที่หลากหลาย และผลรวมของกรดอะมิโนทั้งหมดเป็นตัวกำหนดโครงสร้างสามมิติและปฏิกิริยาทางเคมี[ 35 ]
กรดอะมิโนในสายโพลีเปปไทด์เชื่อมต่อกันด้วยพันธะเปปไทด์ระหว่างหมู่เอมีโนและหมู่คาร์บอกซิล กรดอะมิโนแต่ละตัวในสายเรียกว่าหน่วยย่อยและลำดับของอะตอมคาร์บอน ไนโตรเจน และออกซิเจนที่เชื่อมต่อกันเรียกว่าสายหลักหรือ โครงสร้างหลัก ของโปรตีน[ 36 ] : 19 พันธะเปปไทด์มี รูปแบบ เรโซแนนซ์ สองแบบ ที่ทำให้ โครงสร้างหลักมีลักษณะ พันธะคู่คาร์บอนอัลฟาอยู่ในระนาบเดียวกัน กับไนโตรเจนและหมู่คาร์บอนิล (C=O) โดยประมาณ มุมไดเฮดรัลอีกสอง มุม ในพันธะเปปไทด์กำหนดรูปร่างเฉพาะที่โครงสร้างหลักของโปรตีนมี ผลที่ตามมาประการหนึ่งของลักษณะพันธะคู่ NC(O) คือโปรตีนค่อนข้างแข็ง[ 36 ] : 31 สายโพลีเปปไทด์สิ้นสุดด้วยหมู่เอมีโนอิสระที่เรียกว่าปลาย Nหรือปลายเอมีโนและหมู่คาร์บอกซิลอิสระที่เรียกว่าปลาย Cหรือปลายคาร์บอกซิล[ 37 ]ตามธรรมเนียมแล้ว ลำดับของเปปไทด์จะเขียนจากปลาย N ไปยังปลาย C ซึ่งสอดคล้องกับลำดับที่โปรตีนถูกสังเคราะห์โดยไรโบโซม[ 37 ] [ 38 ]
คำว่าโปรตีนโพลีเปปไทด์และเปปไทด์มีความหมายกำกวมเล็กน้อยและอาจทับซ้อนกันได้ โดยทั่วไปแล้ว โปรตีนจะใช้เรียกโมเลกุลทางชีวภาพที่สมบูรณ์ในโครงสร้าง ที่เสถียร ในขณะที่เปปไทด์โดยทั่วไปจะสงวนไว้สำหรับโอลิโกเมอร์ของกรดอะมิโนสั้นๆ ซึ่งมักไม่มีโครงสร้างสามมิติที่เสถียร แต่ขอบเขตระหว่างทั้งสองนั้นไม่ชัดเจนนักและมักจะอยู่ที่ประมาณ 20–30 หน่วยย่อย[ 39 ]
โปรตีนสามารถ โต้ตอบกับโมเลกุลและไอออนหลายประเภท รวมถึงโปรตีนอื่นๆไขมันคาร์โบไฮเดรตและDNA [ 40 ] [ 41 ] [ 42 ]
ความอุดมสมบูรณ์ในเซลล์
โดยทั่วไป แล้ว เซลล์ แบคทีเรียเช่นE. coliและStaphylococcus aureusจะมีโปรตีนประมาณ 2 ล้านโมเลกุล แบคทีเรียขนาดเล็ก เช่นMycoplasmaหรือspirochetesจะมีโมเลกุลน้อยกว่า ประมาณ 50,000 ถึง 1 ล้านโมเลกุล ในทางตรงกันข้าม เซลล์ ยูคาริโอตมีขนาดใหญ่กว่า จึงมีโปรตีนมากกว่ามาก ตัวอย่างเช่น เซลล์ ยีสต์คาดว่าจะมีโมเลกุลโปรตีนประมาณ 50 ล้านโมเลกุล และ เซลล์ มนุษย์มีประมาณ 1 ถึง 3 พันล้าน โมเลกุล [ 43 ]ความเข้มข้นของสำเนาโปรตีนแต่ละชนิดมีตั้งแต่ไม่กี่โมเลกุลต่อเซลล์ไปจนถึง 20 ล้านโมเลกุล[ 44 ]ยีนที่เข้ารหัสโปรตีนไม่ได้แสดงออกในเซลล์ส่วนใหญ่เสมอไป และจำนวนของยีนขึ้นอยู่กับปัจจัยต่างๆ เช่น ชนิดของเซลล์และสิ่งกระตุ้นภายนอก ตัวอย่างเช่น จากโปรตีนประมาณ 20,000 ชนิดที่เข้ารหัสโดยจีโนมของมนุษย์ มีเพียง 6,000 ชนิดเท่านั้นที่ตรวจพบในเซลล์ลิมโฟบลาสตอยด์[ 45 ]เชื่อกันว่าโปรตีนที่มีมากที่สุดในธรรมชาติคือRuBisCOซึ่งเป็นเอนไซม์ที่เร่งปฏิกิริยาการรวมตัวของคาร์บอนไดออกไซด์เข้ากับสารอินทรีย์ในกระบวนการสังเคราะห์แสงพืชอาจมีเอนไซม์นี้มากถึง 1% โดยน้ำหนัก[ 46 ]
สังเคราะห์
การสังเคราะห์ทางชีวภาพ
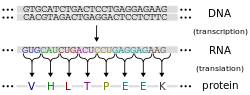
โปรตีนถูกประกอบขึ้นจากกรดอะมิโนโดยใช้ข้อมูลที่เข้ารหัสอยู่ในยีน โปรตีนแต่ละชนิดมีลำดับกรดอะมิโนเฉพาะตัวซึ่งกำหนดโดย ลำดับ นิวคลีโอไทด์ของยีนที่เข้ารหัสโปรตีนนั้นรหัสพันธุกรรมเป็นชุดของนิวคลีโอไทด์สามตัวที่เรียกว่าโคดอนและการรวมกันของนิวคลีโอไทด์สามตัวแต่ละชุดจะกำหนดกรดอะมิโนหนึ่งตัว ตัวอย่างเช่น AUG ( อะดีนีน – ยูราซิล – กัวนีน ) เป็นรหัสสำหรับเมไทโอนีนเนื่องจากDNAประกอบด้วยนิวคลีโอไทด์สี่ตัว จำนวนโคดอนที่เป็นไปได้ทั้งหมดจึงมี 64 ตัว ดังนั้นจึงมีความซ้ำซ้อนในรหัสพันธุกรรมอยู่บ้าง โดยกรดอะมิโนบางตัวถูกกำหนดโดยโคดอนมากกว่าหนึ่งตัว[ 42 ] : 1002–42 ยีนที่เข้ารหัสใน DNA จะ ถูกถอดรหัสเป็นพรีเมสเซนเจอร์อาร์เอ็นเอ (mRNA) ก่อนโดยโปรตีน เช่นอาร์เอ็นเอพอลิเมอเรส สิ่งมีชีวิตส่วนใหญ่จะประมวลผล pre-mRNA (ทรานสคริปต์หลัก ) โดยใช้การดัดแปลงหลังการถอดรหัสใน รูปแบบต่างๆ เพื่อสร้าง mRNA ที่สมบูรณ์ ซึ่งจะถูกใช้เป็นแม่แบบสำหรับการสังเคราะห์โปรตีนโดยไรโบโซมในโปรคาริโอต mRNA อาจถูกนำมาใช้ทันทีที่ผลิต หรือถูกจับโดยไรโบโซมหลังจากเคลื่อนตัวออกจากนิวคลีออยด์ในทางตรงกันข้ามยูคาริโอตสร้าง mRNA ในนิวเคลียสของเซลล์แล้วเคลื่อนย้ายข้ามเยื่อหุ้มนิวเคลียสไปยังไซโตพลาสซึมซึ่งการสังเคราะห์โปรตีนจะเกิดขึ้น อัตราการสังเคราะห์โปรตีนในโปรคาริโอตสูงกว่าในยูคาริโอตและสามารถสูงถึง 20 กรดอะมิโนต่อวินาที[ 47 ]
กระบวนการสังเคราะห์โปรตีนจากแม่แบบ mRNA เรียกว่าการแปล (translation ) mRNA จะถูกโหลดลงบนไรโบโซมและถูกอ่านทีละสามนิวคลีโอไทด์โดยการจับคู่โคดอนแต่ละตัวกับ แอ นติโคดอน ที่ จับคู่เบส ซึ่งอยู่บน โมเลกุล tRNAซึ่งบรรทุกกรดอะมิโนที่สอดคล้องกับโคดอนที่มันรู้จัก เอนไซม์อะมิโนเอซิล tRNA ซินเทสจะ "เติม" กรดอะมิโนที่ถูกต้องให้กับโมเลกุล tRNA โพลีเปปไทด์ที่กำลังเติบโตมักเรียกว่าสายโซ่เกิดใหม่ (nascent chain ) โปรตีนจะถูกสังเคราะห์ทางชีวภาพจากปลาย Nไปยังปลาย Cเสมอ[ 42 ] : 1002–42
ขนาดของโปรตีนที่สังเคราะห์ขึ้นสามารถวัดได้จากจำนวนกรดอะมิโนที่ประกอบอยู่และมวลโมเลกุล ทั้งหมด ซึ่งโดยปกติจะรายงานด้วยหน่วยดาลตัน (Da) หรือหน่วยอนุพันธ์กิโลดาลตัน (kDa) ขนาดเฉลี่ยของโปรตีนเพิ่มขึ้นจากอาร์เคียไปจนถึงแบคทีเรียและยูคาริโอต (283, 311, 438 หน่วย และ 31, 34, 49 kDa ตามลำดับ) เนื่องจากจำนวนโดเมนโปรตีนที่ประกอบเป็นโปรตีนในสิ่งมีชีวิตชั้นสูง มีจำนวนมากขึ้น [ 48 ]ตัวอย่างเช่น โปรตีน ของยีสต์มีความยาวเฉลี่ย 466 กรดอะมิโนและมีมวล 53 kDa [ 39 ]โปรตีนที่ใหญ่ที่สุดที่รู้จักคือไททินซึ่งเป็นส่วนประกอบของซาร์โคเมียร์ของกล้ามเนื้อ โดยมีมวลโมเลกุลเกือบ3000 กิโลดาลตันและความยาวรวมเกือบกรดอะมิโน27,000 ตัว[ 49 ]
การสังเคราะห์ทางเคมี

โปรตีนขนาดสั้นสามารถสังเคราะห์ได้ทางเคมีโดยกลุ่มของ วิธี การสังเคราะห์เปปไทด์ซึ่งอาศัย เทคนิค การสังเคราะห์อินทรีย์เช่นการเชื่อมต่อทางเคมีเพื่อผลิตเปปไทด์ในปริมาณมาก[ 50 ]การสังเคราะห์ทางเคมีช่วยให้สามารถนำกรดอะมิโนที่ไม่เป็นธรรมชาติเข้าสู่สายโซ่โพลีเปปไทด์ได้ เช่น การติด โพ รบเรืองแสงเข้ากับหมู่ข้างของกรดอะมิโน (ซึ่งสามารถทำได้โดยวิศวกรรมพันธุกรรมเมื่อผลิตโปรตีนรีคอมบิแนนท์) [ 51 ] [ 52 ]วิธีการเหล่านี้มีประโยชน์ในชีวเคมี ในห้องปฏิบัติการ และชีววิทยาของเซลล์แม้ว่าโดยทั่วไปแล้วจะไม่เหมาะสำหรับการใช้งานเชิงพาณิชย์ การสังเคราะห์ทางเคมีไม่มีประสิทธิภาพสำหรับโพลีเปปไทด์ที่มีความยาวมากกว่าประมาณ 300 กรดอะมิโน และโปรตีนที่สังเคราะห์ขึ้นอาจไม่สามารถมีโครงสร้างตติยภูมิ แบบดั้งเดิมได้ง่าย วิธีการสังเคราะห์ทางเคมีส่วนใหญ่ดำเนินการจากปลาย C ไปยังปลาย N ซึ่งตรงกันข้ามกับปฏิกิริยาทางชีวภาพ[ 53 ]
โครงสร้าง


โปรตีนส่วนใหญ่จะพับตัวเป็นโครงสร้างสามมิติที่เป็นเอกลักษณ์ รูปร่างที่โปรตีนพับตัวตามธรรมชาติเรียกว่าโครงสร้างดั้งเดิม[ 36 ] : 36 แม้ว่าโปรตีนหลายชนิดสามารถพับตัวได้โดยไม่ต้องอาศัยความช่วยเหลือ เพียงแค่คุณสมบัติทางเคมีของกรดอะมิโน แต่บางชนิดก็ต้องการความช่วยเหลือจากโมเลกุลชาเปอโรนเพื่อพับตัวเป็นโครงสร้าง ดั้งเดิม [ 36 ] : 37 นักชีวเคมีมักกล่าวถึงลักษณะที่แตกต่างกันสี่ประการของโครงสร้างโปรตีน: [ 36 ] : 30–34
- โครงสร้างปฐมภูมิ :ลำดับของกรดอะมิโนโปรตีนเป็นพอลิอะไมด์
- โครงสร้างทุติยภูมิ : โครงสร้างเฉพาะที่ที่ซ้ำกันอย่างสม่ำเสมอและมีความเสถียรโดยพันธะไฮโดรเจนตัวอย่างที่พบได้บ่อยที่สุด ได้แก่อัลฟาเฮลิกซ์ เบต้าชีทและส่วนโค้งเนื่องจากโครงสร้างทุติยภูมิเป็นโครงสร้างเฉพาะที่ จึงสามารถมีหลายบริเวณที่มีโครงสร้างทุติยภูมิที่แตกต่างกันอยู่ในโมเลกุลโปรตีนเดียวกันได้
- โครงสร้างระดับตติยภูมิ : รูปร่างโดยรวมของโมเลกุลโปรตีนเดี่ยว ความสัมพันธ์เชิงพื้นที่ของโครงสร้างระดับทุติยภูมิกับโครงสร้างระดับอื่นๆ โครงสร้างระดับตติยภูมิโดยทั่วไปจะมีความเสถียรโดยปฏิกิริยาที่ไม่ใช่เฉพาะที่ ซึ่งส่วนใหญ่คือการก่อตัวของแกนไฮโดรโฟบิกแต่ก็อาจเกิดขึ้นผ่านพันธะเกลือพันธะไฮโดรเจน พันธะไดซัลไฟด์และแม้แต่การดัดแปลงหลังการสังเคราะห์โปรตีน คำว่า "โครงสร้างระดับตติยภูมิ" มักใช้เป็นคำพ้องความหมายกับคำว่า " การพับ " โครงสร้างระดับตติยภูมิเป็นสิ่งที่ควบคุมการทำงานพื้นฐานของโปรตีน
- โครงสร้างระดับควอเทอร์นารี : โครงสร้างที่เกิดจากโมเลกุลโปรตีนหลายโมเลกุล (สายโซ่โพลีเปปไทด์) ซึ่งในบริบทนี้มักเรียกว่าหน่วยย่อยของโปรตีนทำหน้าที่รวมกันเป็นโปรตีนเชิงซ้อน เดียว
- โครงสร้างควินารี : ลักษณะเฉพาะของพื้นผิวโปรตีนที่จัดระเบียบภายในเซลล์ที่แออัด โครงสร้างควินารีขึ้นอยู่กับปฏิกิริยาระหว่างโมเลกุลขนาดใหญ่ที่เกิดขึ้นชั่วคราวแต่มีความสำคัญอย่างยิ่งภายในเซลล์สิ่งมีชีวิต
โปรตีนไม่ใช่โมเลกุลที่แข็งตัวอย่างสมบูรณ์ นอกจากโครงสร้างในระดับเหล่านี้แล้ว โปรตีนอาจเปลี่ยนไปมาระหว่างโครงสร้างที่เกี่ยวข้องหลายโครงสร้างในขณะที่ทำหน้าที่ ในบริบทของการจัดเรียงหน้าที่ใหม่เหล่านี้ โครงสร้างระดับตติยภูมิหรือจตุรภูมิเหล่านี้มักเรียกว่า " คอนฟอร์เมชัน " และการเปลี่ยนผ่านระหว่างโครงสร้างเหล่านี้เรียกว่าการเปลี่ยนแปลงคอนฟอร์เมชันการเปลี่ยนแปลงดังกล่าวมักเกิดจากการจับตัวของ โมเลกุล ซับสเตรต กับ ไซต์ที่ออกฤทธิ์ของเอนไซม์หรือบริเวณทางกายภาพของโปรตีนที่เข้าร่วมในการเร่งปฏิกิริยาทางเคมี ในสารละลาย โครงสร้างของโปรตีนจะเปลี่ยนแปลงไปเนื่องจากการสั่นสะเทือนทางความร้อนและการชนกับโมเลกุลอื่น[ 42 ] : 368–75

โดยทั่วไปแล้ว โปรตีนสามารถแบ่งออกเป็น 3 ประเภทหลัก ซึ่งสัมพันธ์กับโครงสร้างตติยภูมิทั่วไป ได้แก่โปรตีนทรงกลมโปรตีนเส้นใยและโปรตีนเยื่อหุ้มเซลล์โปรตีนทรงกลมเกือบทั้งหมดละลายน้ำได้และหลายชนิดเป็นเอนไซม์ โปรตีนเส้นใยมักเป็นโครงสร้าง เช่นคอลลาเจนซึ่งเป็นส่วนประกอบหลักของเนื้อเยื่อเกี่ยวพัน หรือเคราตินซึ่งเป็นส่วนประกอบโปรตีนของเส้นผมและเล็บ โปรตีนเยื่อหุ้มเซลล์มักทำหน้าที่เป็นตัวรับหรือเป็นช่องทางให้โมเลกุลที่มีขั้วหรือมีประจุผ่านเยื่อหุ้มเซลล์ได้[ 42 ] : 165–85
พันธะไฮโดรเจนภายในโมเลกุลของโปรตีนในกรณีพิเศษ ซึ่งได้รับการปกป้องจากการโจมตีของน้ำได้ไม่ดี จึงส่งเสริมให้เกิดการขาดน้ำ ของตัวมันเอง เรียกว่าดีไฮดรอน[ 54 ]
โดเมนโปรตีน
โปรตีนหลายชนิดประกอบด้วยโดเมนโปรตีน หลายโดเมน กล่าว คือ ส่วนของโปรตีนที่พับตัวเป็นหน่วยโครงสร้างที่แตกต่างกัน[ 55 ] : 134 โดเมนมักมีหน้าที่เฉพาะ เช่นกิจกรรมของเอนไซม์ (เช่น ไคเนส ) หรือทำหน้าที่เป็นโมดูลการจับ[ 55 ] : 155–156

ลำดับโมทีฟ
ลำดับกรดอะมิโนสั้นๆ ภายในโปรตีนมักทำหน้าที่เป็นไซต์การจดจำสำหรับโปรตีนอื่นๆ[ 57 ]ตัวอย่างเช่นโดเมน SH3มักจะจับกับโมทีฟ PxxP สั้นๆ (เช่นโพรลีน 2 ตัว [P] คั่นด้วย กรดอะมิโนที่ไม่ระบุ 2 ตัว[x] แม้ว่ากรดอะมิโนโดยรอบอาจกำหนดความจำเพาะในการจับที่แน่นอนได้) โมทีฟดังกล่าวจำนวนมากได้รับการรวบรวมไว้ใน ฐานข้อมูล Eukaryotic Linear Motif (ELM) [ 58 ]
หน้าที่ของเซลล์
โปรตีนเป็นตัวการสำคัญภายในเซลล์ กล่าวกันว่าทำหน้าที่ตามข้อมูลที่กำหนดในยีน[ 39 ]ยกเว้นRNA บางประเภท โมเลกุลทางชีวภาพอื่นๆ ส่วนใหญ่เป็นองค์ประกอบเฉื่อยที่โปรตีนทำหน้าที่ โปรตีนประกอบขึ้นเป็นครึ่งหนึ่งของน้ำหนักแห้งของ เซลล์ Escherichia coliในขณะที่โมเลกุลขนาดใหญ่อื่นๆ เช่น DNA และ RNA ประกอบขึ้นเพียง 3% และ 20% ตามลำดับ[ 59 ]ชุดของโปรตีนที่แสดงออกในเซลล์หรือชนิดของเซลล์ใดเซลล์หนึ่งเรียกว่าโปรตีโอม [ 55 ] : 120

คุณลักษณะสำคัญของโปรตีนที่ทำให้โปรตีนทำหน้าที่หลากหลายคือความสามารถในการจับกับโมเลกุลอื่นอย่างจำเพาะเจาะจงและแน่นหนา บริเวณของโปรตีนที่รับผิดชอบในการจับกับโมเลกุลอื่นเรียกว่าตำแหน่งจับ (binding site)ซึ่งมักจะเป็นรอยบุ๋มหรือ "ช่อง" บนพื้นผิวของโมเลกุล ความสามารถในการจับนี้เกิดจากโครงสร้างระดับตติยภูมิของโปรตีน ซึ่งกำหนดช่องตำแหน่งจับ และจากคุณสมบัติทางเคมีของหมู่ข้างเคียงของกรดอะมิโนที่อยู่รอบๆ การจับกันของโปรตีนอาจแน่นหนาและจำเพาะเจาะจงอย่างมาก ตัวอย่างเช่น โปรตีน ยับยั้งไรโบเอนไซม์จับกับแอนจิโอเจนิน ของมนุษย์ ด้วยค่าคงที่การแยกตัว ต่ำกว่าเฟมโตโมลาร์ (< 10 −15 M) แต่ไม่จับกับออน โคเนสซึ่งเป็นโฮโมล็อกของสัตว์ครึ่งบกครึ่งน้ำ เลย (> 1 M) การเปลี่ยนแปลงทางเคมีเพียงเล็กน้อย เช่น การเพิ่มหมู่เมทิลเพียงหมู่เดียวให้กับคู่จับก็อาจเพียงพอที่จะกำจัดการจับได้เกือบทั้งหมด ตัวอย่างเช่นอะมิโนเอซิล tRNA ซินเทสที่จำเพาะต่อกรดอะมิโนวาลีนจะแยกแยะออกจากโซ่ข้างที่คล้ายคลึงกันมากของกรดอะมิโนไอโซลิวซีน[ 60 ]
โปรตีนสามารถจับกับโปรตีนอื่น ๆ รวมถึง สารตั้งต้น โมเลกุลขนาดเล็กได้เมื่อโปรตีนจับกับโมเลกุลเดียวกันอย่างจำเพาะเจาะจง โปรตีนเหล่านั้นสามารถรวมตัวกันเป็นโอลิโกเมอร์เพื่อสร้างเส้นใยได้ กระบวนการนี้มักเกิดขึ้นในโปรตีนโครงสร้างซึ่งประกอบด้วยโมโนเมอร์ทรงกลมที่รวมตัวกันเองเพื่อสร้างเส้นใยที่แข็งแรง ปฏิสัมพันธ์ ระหว่างโปรตีนควบคุมกิจกรรมของเอนไซม์ ควบคุมความก้าวหน้าของวงจรเซลล์และช่วยให้เกิดการประกอบกันของโปรตีนเชิงซ้อน ขนาดใหญ่ ที่ดำเนินการปฏิกิริยาที่เกี่ยวข้องอย่างใกล้ชิดหลายอย่างที่มีหน้าที่ทางชีวภาพร่วมกัน โปรตีนสามารถจับกับหรือรวมเข้ากับเยื่อหุ้มเซลล์ได้ ความสามารถของคู่พันธะในการเหนี่ยวนำให้เกิดการเปลี่ยนแปลงโครงสร้างในโปรตีนช่วยให้สามารถสร้างเครือข่ายการส่งสัญญาณ ที่ซับซ้อนอย่างมากได้ [ 42 ] : 830–49 เนื่องจากปฏิสัมพันธ์ระหว่างโปรตีนสามารถย้อนกลับได้และขึ้นอยู่กับความพร้อมของกลุ่มโปรตีนคู่พันธะที่แตกต่างกันในการสร้างกลุ่มที่สามารถดำเนินการชุดหน้าที่เฉพาะได้ การศึกษาปฏิสัมพันธ์ระหว่างโปรตีนเฉพาะจึงเป็นกุญแจสำคัญในการทำความเข้าใจแง่มุมที่สำคัญของหน้าที่ของเซลล์ และท้ายที่สุดคือคุณสมบัติที่แยกแยะเซลล์ประเภทต่างๆ[ 61 ] [ 62 ]
เอนไซม์
บทบาทที่รู้จักกันดีที่สุดของโปรตีนในเซลล์คือการเป็นเอนไซม์ซึ่ง ทำหน้าที่ เร่งปฏิกิริยาเคมี เอนไซม์มักมีความจำเพาะสูงและเร่งปฏิกิริยาเคมีเพียงหนึ่งหรือสองปฏิกิริยาเท่านั้น เอนไซม์ทำหน้าที่ในปฏิกิริยาส่วนใหญ่ที่เกี่ยวข้องกับการเผาผลาญรวมถึงการจัดการ DNA ในกระบวนการต่างๆ เช่นการจำลอง DNAการซ่อมแซม DNAและการถอดรหัสเอนไซม์บางชนิดทำหน้าที่กับโปรตีนอื่นๆ เพื่อเพิ่มหรือลบกลุ่มเคมีในกระบวนการที่เรียกว่าการดัดแปลงหลังการแปลรหัส มีปฏิกิริยาประมาณ 4,000 ปฏิกิริยาที่ทราบกันว่าถูกเร่งปฏิกิริยาโดยเอนไซม์[ 63 ]การเร่งอัตราที่เกิดจากการเร่งปฏิกิริยาโดยเอนไซม์มักมีขนาดใหญ่มาก—มากถึง 10 17เท่าของอัตราเมื่อเทียบกับปฏิกิริยาที่ไม่มีตัวเร่งปฏิกิริยาในกรณีของออโรเทตดีคาร์บอก ซิเลส (78 ล้านปีหากไม่มีเอนไซม์ 18 มิลลิวินาทีเมื่อมีเอนไซม์) [ 64 ]
โมเลกุลที่ถูกจับและกระทำโดยเอนไซม์เรียกว่าสารตั้งต้นแม้ว่าเอนไซม์จะประกอบด้วยกรดอะมิโนหลายร้อยตัว แต่โดยปกติแล้วจะมีเพียงส่วนน้อยของสารตกค้างเท่านั้นที่สัมผัสกับสารตั้งต้น และมีเพียงส่วนน้อยยิ่งกว่านั้น—โดยเฉลี่ยสามถึงสี่สารตกค้าง—ที่เกี่ยวข้องโดยตรงกับการเร่งปฏิกิริยา[ 65 ]บริเวณของเอนไซม์ที่จับกับสารตั้งต้นและมีสารตกค้างเร่งปฏิกิริยาเรียกว่าไซต์ที่ใช้งาน[ 55 ] : 389
โปรตีนไดริเจนท์เป็นสมาชิกของกลุ่มโปรตีนที่กำหนดสเตอริโอเคมีของสารประกอบที่สังเคราะห์โดยเอนไซม์อื่น[ 66 ]
การส่งสัญญาณของเซลล์และการจับกับลิแกนด์

โปรตีนหลายชนิดมีส่วนเกี่ยวข้องในกระบวนการส่งสัญญาณของเซลล์และการส่งสัญญาณ โปรตีนบางชนิด เช่นอินซูลินเป็นโปรตีนนอกเซลล์ที่ส่งสัญญาณจากเซลล์ที่สังเคราะห์ขึ้นไปยังเซลล์อื่นในเนื้อเยื่อ ที่อยู่ห่างไกล ส่วนโปรตีน อื่นๆ เป็นโปรตีนเยื่อหุ้มเซลล์ที่ทำหน้าที่เป็นตัวรับซึ่งหน้าที่หลักคือการจับกับโมเลกุลส่งสัญญาณและกระตุ้นการตอบสนองทางชีวเคมีในเซลล์ ตัวรับหลายชนิดมีตำแหน่งการจับที่อยู่บนพื้นผิวเซลล์และโดเมนตัวกระตุ้นภายในเซลล์ ซึ่งอาจมีกิจกรรมของเอนไซม์หรืออาจมีการเปลี่ยนแปลงโครงสร้างที่ตรวจจับได้โดยโปรตีนอื่นๆ ภายในเซลล์[ 41 ] : 251–81
แอนติบอดีเป็นส่วนประกอบโปรตีนของระบบภูมิคุ้มกันแบบปรับตัวซึ่งมีหน้าที่หลักในการจับกับแอนติเจนหรือสารแปลกปลอมในร่างกาย และกำหนดเป้าหมายเพื่อทำลาย แอนติบอดีสามารถหลั่งออกมาสู่สิ่งแวดล้อมภายนอกเซลล์ หรือยึดติดอยู่ในเยื่อหุ้มเซลล์ของเซลล์ B พิเศษ ที่เรียกว่าเซลล์พลาสมาในขณะที่เอนไซม์มีข้อจำกัดในความสามารถในการจับกับสารตั้งต้นเนื่องจากความจำเป็นในการดำเนินการปฏิกิริยา แต่แอนติบอดีไม่มีข้อจำกัดดังกล่าว ความสามารถในการจับของแอนติบอดีกับเป้าหมายนั้นสูงมาก[ 42 ] : 275–50
โปรตีนขนส่งลิแกนด์จำนวนมากจะจับกับโมเลกุลชีวภาพขนาดเล็ก ที่เฉพาะเจาะจง และขนส่งไปยังตำแหน่งอื่น ๆ ในร่างกายของสิ่งมีชีวิตหลายเซลล์ โปรตีนเหล่านี้ต้องมีความสัมพันธ์ในการจับสูงเมื่อลิแกนด์มีอยู่ในความเข้มข้นสูง และจะปล่อยลิแกนด์เมื่อมีอยู่ในความเข้มข้นต่ำในเนื้อเยื่อเป้าหมาย ตัวอย่างที่เป็นแบบอย่างของโปรตีนที่จับกับลิแกนด์คือฮีโมโกลบินซึ่งขนส่งออกซิเจนจากปอด ไปยังอวัยวะและเนื้อเยื่ออื่น ๆ ในสัตว์ มีกระดูกสันหลังทั้งหมดและมีโฮโมล็อกที่ใกล้เคียงกันในทุกอาณาจักรชีวภาพ[ 42 ] : 222–29 เลคตินเป็นโปรตีนที่จับกับน้ำตาลซึ่งมีความจำเพาะสูงต่อหมู่ของน้ำตาลเลคตินมักมีบทบาทใน ปรากฏการณ์ การจดจำ ทางชีวภาพ ที่เกี่ยวข้องกับเซลล์และโปรตีน[ 67 ]ตัวรับเป็นโปรตีนที่จับกับสารที่มีความจำเพาะสูง และฮอร์โมน หลายชนิด ทำงานโดยการจับกับตัวรับของพวกมันโดยเฉพาะ[ 68 ]
โปรตีนทรานส์เมมเบรนสามารถทำหน้าที่เป็นโปรตีนขนส่งลิแกนด์ที่เปลี่ยนแปลงการซึมผ่านของเยื่อหุ้มเซลล์ต่อโมเลกุลขนาดเล็กและไอออน เยื่อหุ้มเซลล์เองมี แกน ไฮโดรโฟบิกซึ่งโมเลกุลที่มีขั้ว หรือมีประจุไม่สามารถ แพร่ ผ่าน ได้ โปรตีนเยื่อหุ้มเซลล์มีช่องภายในที่อนุญาตให้โมเลกุลดังกล่าวเข้าและออกจากเซลล์ได้ โปรตีน ช่องไอออน หลายชนิด มีความเชี่ยวชาญในการเลือกเฉพาะไอออนบางชนิดเท่านั้น ตัวอย่างเช่น ช่อง โพแทสเซียมและโซเดียมมักจะเลือกเฉพาะไอออนใดไอออนหนึ่งจากสองไอออน[ 41 ] : 232–34
โปรตีนโครงสร้าง

โปรตีนโครงสร้างทำให้ส่วนประกอบทางชีวภาพที่ปกติแล้วเป็นของเหลวมีความแข็งและคงรูป โปรตีนโครงสร้างส่วนใหญ่เป็นโปรตีนเส้นใยตัวอย่างเช่นคอลลาเจนและอีลาสตินเป็นส่วนประกอบสำคัญของเนื้อเยื่อเกี่ยวพันเช่นกระดูกอ่อนและเคราตินพบในโครงสร้างที่แข็งหรือเป็นเส้นใย เช่นเส้นผมเล็บขนนกกีบและเปลือกหอยบางชนิด[ 42 ] : 178–81 โปรตีนทรง กลมบางชนิดสามารถทำหน้าที่โครงสร้างได้ ตัวอย่างเช่นแอคตินและทูบูลินเป็นโปรตีนทรงกลมและละลายได้ในรูปโมโนเมอร์ แต่จะรวมตัวกันเป็นพอลิเมอร์เพื่อสร้างเส้นใยยาวและแข็งที่ประกอบเป็นโครงกระดูกเซลล์ซึ่งช่วยให้เซลล์รักษารูปร่างและขนาดไว้ได้[ 55 ] : 490
โปรตีนอื่นๆ ที่ทำหน้าที่เชิงโครงสร้าง ได้แก่โปรตีนมอเตอร์เช่นไมโอซินไคเนซินและไดเนอินซึ่งสามารถสร้างแรงเชิงกลได้ โปรตีนเหล่านี้มีความสำคัญต่อการเคลื่อนที่ ของเซลล์ ในสิ่งมีชีวิตเซลล์เดียวและอสุจิของสิ่งมีชีวิตหลายเซลล์จำนวนมากที่สืบพันธุ์แบบอาศัยเพศพวกมันสร้างแรงที่เกิดจากการหดตัวของกล้ามเนื้อ[ 42 ] : 258–64, 272 และมีบทบาทสำคัญในการขนส่งภายในเซลล์[ 55 ] : 481, 490
วิธีการศึกษา
วิธีการที่ใช้กันทั่วไปในการศึกษาโครงสร้างและหน้าที่ของโปรตีน ได้แก่อิมมูโนฮิสโตเคมี การกลายพันธุ์ แบบกำหนดตำแหน่ง การตกผลึกด้วยรังสีเอกซ์ การเรโซแนนซ์แม่เหล็กนิวเคลียร์และแมสสเปกโทรเมตรีกิจกรรมและโครงสร้างของโปรตีนสามารถตรวจสอบได้ทั้งในหลอดทดลองในร่างกายและในคอมพิวเตอร์การ ศึกษา ใน หลอดทดลองของโปรตีนบริสุทธิ์ในสภาพแวดล้อมที่ควบคุมได้ นั้นมีประโยชน์สำหรับการเรียนรู้ว่าโปรตีนทำหน้าที่อย่างไร[ 69 ]ตัวอย่างเช่น การศึกษา จลนศาสตร์ของเอนไซม์จะสำรวจกลไกทางเคมีของกิจกรรมเร่งปฏิกิริยาของเอนไซม์และความสัมพันธ์สัมพัทธ์กับโมเลกุลของสารตั้งต้นที่เป็นไปได้ต่างๆ[ 70 ]ในทางตรงกันข้าม การทดลอง ในร่างกายสามารถให้ข้อมูลเกี่ยวกับบทบาททางสรีรวิทยาของโปรตีนในบริบทของเซลล์หรือแม้แต่สิ่งมีชีวิต ทั้งหมด และมักจะให้ข้อมูลเพิ่มเติมเกี่ยวกับพฤติกรรมของโปรตีนในบริบทต่างๆ[ 71 ] การศึกษา ในคอมพิวเตอร์ใช้วิธีการคำนวณเพื่อศึกษาโปรตีน[ 72 ]
การทำให้โปรตีนบริสุทธิ์
โปรตีนอาจได้รับการทำให้บริสุทธิ์จากส่วนประกอบเซลล์อื่นๆ โดยใช้เทคนิคต่างๆ เช่นการปั่นเหวี่ยงความเร็วสูง การตกตะกอน อิ เล็กโทร โฟเรซิสและโครมาโทกราฟี [ 36 ] : 21–24 การเกิดขึ้นของวิศวกรรมพันธุกรรมทำให้มีวิธีการต่างๆ มากมายที่ช่วยอำนวยความสะดวกในการทำให้บริสุทธิ์[ 73 ]
ในการ วิเคราะห์ ในหลอดทดลองจำเป็นต้องแยกโปรตีนออกจากส่วนประกอบอื่นๆ ของเซลล์ กระบวนการนี้มักเริ่มต้นด้วยการสลายเซลล์ซึ่งเยื่อหุ้มเซลล์จะถูกทำลายและส่วนประกอบภายในเซลล์จะถูกปล่อยออกมาในสารละลายที่เรียกว่าไลเซตดิบส่วนผสมที่ได้สามารถทำให้บริสุทธิ์ได้โดยใช้การปั่นเหวี่ยงความเร็วสูงซึ่งจะแยกส่วนประกอบต่างๆ ของเซลล์ออกเป็นส่วนๆ ที่ประกอบด้วยโปรตีนที่ละลายได้ ไขมันและโปรตีน ในเยื่อหุ้มเซลล์ ออร์แกเนลล์ ของเซลล์ และกรดนิวคลีอิกการตกตะกอนด้วยวิธีที่เรียกว่าการเติมเกลือ สามารถเพิ่มความเข้มข้นของโปรตีนจากไลเซตนี้ได้ จากนั้นจึงใช้ โครมาโทกราฟีหลายประเภทเพื่อแยกโปรตีนที่สนใจโดยอาศัยคุณสมบัติต่างๆ เช่น น้ำหนักโมเลกุล ประจุสุทธิ และความสามารถในการจับตัว[ 36 ] : 21–24 ระดับการทำให้บริสุทธิ์สามารถตรวจสอบได้โดยใช้เจลอิเล็กโทรโฟเรซิส ประเภทต่างๆ หาก ทราบน้ำหนักโมเลกุลและจุดไอโซอิเล็กทริก ของโปรตีนที่ต้องการ โดยใช้ สเปกโทรสโกปีหากโปรตีนมีคุณสมบัติทางสเปกโทรสโกปีที่สามารถแยกแยะได้ หรือโดยการทดสอบเอนไซม์หากโปรตีนมีกิจกรรมของเอนไซม์ นอกจากนี้ โปรตีนยังสามารถแยกได้ตามประจุโดยใช้อิเล็กโทรโฟกัสซิง[ 74 ]
สำหรับโปรตีนธรรมชาติ อาจจำเป็นต้องมีขั้นตอนการทำให้บริสุทธิ์หลายขั้นตอนเพื่อให้ได้โปรตีนที่บริสุทธิ์เพียงพอสำหรับการใช้งานในห้องปฏิบัติการ เพื่อลดความซับซ้อนของกระบวนการนี้ มักใช้ พันธุวิศวกรรมในการเพิ่มคุณสมบัติทางเคมีให้กับโปรตีนเพื่อให้ง่ายต่อการทำให้บริสุทธิ์โดยไม่ส่งผลกระทบต่อโครงสร้างหรือกิจกรรมของโปรตีน ในที่นี้ "แท็ก" ที่ประกอบด้วยลำดับกรดอะมิโนเฉพาะ ซึ่งมักจะเป็นชุดของ สารตกค้าง ฮิสติดีน (" แท็กฮิส ") จะถูกติดไว้ที่ปลายด้านหนึ่งของโปรตีน ผลที่ได้คือ เมื่อไลเซตผ่านคอลัมน์โครมาโทกราฟีที่มีนิกเกิลสารตกค้างฮิสติดีนจะจับกับนิกเกิลและยึดติดกับคอลัมน์ ในขณะที่ส่วนประกอบที่ไม่มีแท็กของไลเซตจะผ่านไปได้โดยไม่ติดขัด มีการพัฒนาแท็กจำนวนมากเพื่อช่วยให้นักวิจัยทำให้โปรตีนเฉพาะบริสุทธิ์จากสารผสมที่ซับซ้อน[ 73 ]
ตำแหน่งภายในเซลล์
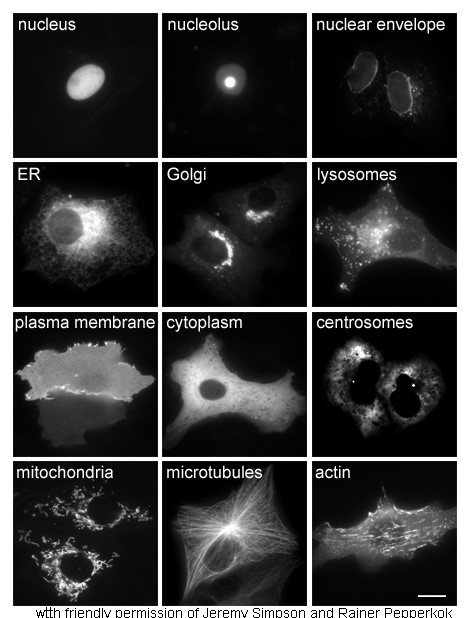
การศึกษาโปรตีนในร่างกายมักเกี่ยวข้องกับการสังเคราะห์และตำแหน่งของโปรตีนภายในเซลล์ แม้ว่าโปรตีนภายในเซลล์จำนวนมากจะถูกสังเคราะห์ในไซโตพลาสซึมและโปรตีนที่ยึดติดกับเยื่อหุ้มเซลล์หรือโปรตีนที่หลั่งออกมาจะถูกสังเคราะห์ในเอนโดพลาสมิกเรติคูลัม แต่ รายละเอียดเกี่ยวกับวิธีการ กำหนดเป้าหมายของโปรตีนไปยังออร์แกเนลล์หรือโครงสร้างเซลล์ที่เฉพาะเจาะจงนั้นมักไม่ชัดเจน เทคนิคที่มีประโยชน์สำหรับการประเมินตำแหน่งของเซลล์คือการใช้พันธุวิศวกรรมเพื่อแสดงออกในเซลล์โปรตีนลูกผสมหรือไคเมราที่ประกอบด้วยโปรตีนธรรมชาติที่สนใจซึ่งเชื่อมโยงกับ " ตัวรายงาน " เช่นโปรตีนเรืองแสงสีเขียว (GFP) [ 75 ]จากนั้นตำแหน่งของโปรตีนลูกผสมภายในเซลล์สามารถมองเห็นได้อย่างชัดเจนและมีประสิทธิภาพโดยใช้กล้องจุลทรรศน์[ 76 ]
วิธีการอื่นๆ ในการระบุตำแหน่งของโปรตีนในเซลล์จำเป็นต้องใช้เครื่องหมายแบ่งส่วนที่ทราบแล้วสำหรับบริเวณต่างๆ เช่น ER, Golgi, ไลโซโซมหรือแวคิวโอล, ไมโตคอนเดรีย, คลอโรพลาสต์, เยื่อหุ้มเซลล์ เป็นต้น การใช้เครื่องหมายที่มีการติดแท็กด้วยฟลูออเรสเซนต์หรือแอนติบอดีต่อเครื่องหมายที่ทราบแล้วจะทำให้การระบุตำแหน่งของโปรตีนที่สนใจทำได้ง่ายขึ้นมาก ตัวอย่างเช่น การตรวจ ภูมิคุ้มกันด้วยฟลูออเรสเซนต์ทางอ้อมจะช่วยให้สามารถระบุตำแหน่งร่วมกันของฟลูออเรสเซนต์และแสดงตำแหน่งได้ สีย้อมฟลูออเรสเซนต์ใช้ในการติดฉลากส่วนต่างๆ ของเซลล์เพื่อวัตถุประสงค์ที่คล้ายกัน[ 77 ]
ยังมีวิธีอื่นๆ อีก ตัวอย่างเช่นการตรวจทางอิมมูโนฮิสโตเคมีมักใช้แอนติบอดีต่อโปรตีนที่สนใจอย่างน้อยหนึ่งชนิดที่เชื่อมต่อกับเอนไซม์ ทำให้เกิดสัญญาณเรืองแสงหรือสัญญาณสีที่สามารถเปรียบเทียบระหว่างตัวอย่างได้ ทำให้ได้ข้อมูลตำแหน่ง[ 78 ]เทคนิคที่ใช้ได้อีกอย่างหนึ่งคือการแยกส่วนร่วมในเกรเดียนต์ซูโครส (หรือวัสดุอื่นๆ) โดยใช้การปั่นเหวี่ยงแบบไอโซพิคนิค [ 79 ] แม้ว่าเทคนิคนี้จะไม่สามารถพิสูจน์การอยู่ร่วมกันของช่องที่มีความหนาแน่นที่ทราบและโปรตีนที่สนใจได้ แต่ก็บ่งชี้ถึงความเป็นไปได้ที่เพิ่มขึ้น[ 79 ]
สุดท้าย วิธีการมาตรฐานทองคำในการระบุตำแหน่งเซลล์คือกล้องจุลทรรศน์อิเล็กตรอนภูมิคุ้มกันเทคนิคนี้ใช้แอนติบอดีต่อโปรตีนที่สนใจ ร่วมกับเทคนิคกล้องจุลทรรศน์อิเล็กตรอนแบบคลาสสิก ตัวอย่างจะถูกเตรียมสำหรับการตรวจสอบด้วยกล้องจุลทรรศน์อิเล็กตรอนตามปกติ จากนั้นจึงทำการบำบัดด้วยแอนติบอดีต่อโปรตีนที่สนใจซึ่งเชื่อมต่อกับวัสดุที่มีความหนาแน่นทางไฟฟ้าสูงมาก โดยปกติคือทองคำ วิธีนี้ช่วยให้สามารถระบุตำแหน่งทั้งรายละเอียดโครงสร้างระดับอัลตราและโปรตีนที่สนใจได้[ 80 ]
ด้วยการประยุกต์ใช้ทางวิศวกรรมพันธุกรรมอีกวิธีหนึ่งที่เรียกว่าการกลายพันธุ์แบบกำหนดตำแหน่งนักวิจัยสามารถเปลี่ยนแปลงลำดับโปรตีนและโครงสร้าง ตำแหน่งในเซลล์ และความไวต่อการควบคุมได้ เทคนิคนี้ยังช่วยให้สามารถรวมกรดอะมิโนที่ไม่เป็นธรรมชาติเข้าไปในโปรตีนได้โดยใช้ tRNA ที่ดัดแปลง[ 81 ]และอาจช่วยให้สามารถออกแบบโปรตีนใหม่ที่มีคุณสมบัติแปลกใหม่ได้อย่าง มีเหตุผล [ 82 ]
โปรตีโอมิกส์
โปรตีนทั้งหมดที่มีอยู่ในเซลล์หรือเซลล์ประเภทหนึ่ง ณ เวลาใดเวลาหนึ่ง เรียกว่าโปรตีโอม (proteome ) และการศึกษาชุดข้อมูลขนาดใหญ่ดังกล่าวได้กำหนดขอบเขตของสาขาโปรตีโอมิกส์ (proteomics)ซึ่งตั้งชื่อตามสาขาที่เกี่ยวข้องคือ จี โนมิก ส์ (genomics ) เทคนิคการทดลองที่สำคัญในโปรตีโอมิกส์ ได้แก่อิเล็กโทรโฟเรซิสแบบ 2 มิติ [ 83 ]ซึ่งช่วยให้สามารถแยกโปรตีนได้หลายชนิด แมสสเปก โทรเมตรี[ 84 ] ซึ่งช่วยให้สามารถระบุโปรตีนและลำดับของเป ปไทด์ได้อย่างรวดเร็วและมีประสิทธิภาพสูง (ส่วนใหญ่มักทำหลังจากการย่อยในเจล ) ไมโครอาร์เรย์โปรตีนซึ่งช่วยให้สามารถตรวจจับระดับสัมพัทธ์ของโปรตีนต่างๆ ที่มีอยู่ในเซลล์ และการคัดกรองแบบทูไฮบริดซึ่งช่วยให้สามารถสำรวจปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนได้ อย่างเป็นระบบ [ 85 ]ปฏิสัมพันธ์ทางชีวภาพที่เป็นไปได้ทั้งหมดเรียกว่าอินเตอร์แอคโตม (interactome ) [ 86 ]ความพยายามอย่างเป็นระบบในการกำหนดโครงสร้างของโปรตีนที่แสดงถึงการพับทุกรูปแบบที่เป็นไปได้ เรียกว่า จีโนมิกส์เชิงโครงสร้าง (structural genomics ) [ 87 ]
การกำหนดโครงสร้าง
การค้นพบโครงสร้างระดับตติยภูมิของโปรตีน หรือโครงสร้างระดับจตุรภูมิของสารประกอบเชิงซ้อน สามารถให้เบาะแสสำคัญเกี่ยวกับวิธีการทำงานของโปรตีนและผลกระทบที่อาจเกิดขึ้น เช่น ในการออกแบบยาเนื่องจากโปรตีนมีขนาดเล็กเกินกว่าจะมองเห็นได้ภายใต้กล้องจุลทรรศน์แบบใช้แสงจึงต้องใช้วิธีการอื่นในการกำหนดโครงสร้างของโปรตีน วิธีการทดลองทั่วไป ได้แก่การตกผลึกด้วยรังสีเอกซ์และการวิเคราะห์สเปกตรัมด้วยคลื่นแม่เหล็กไฟฟ้า (NMR)ซึ่งทั้งสองวิธีสามารถให้ข้อมูลโครงสร้างที่ ความละเอียดระดับ อะตอมได้ อย่างไรก็ตาม การทดลอง NMR สามารถให้ข้อมูลที่สามารถประมาณระยะห่างระหว่างอะตอมคู่ต่างๆ ได้ และรูปร่างสุดท้ายที่เป็นไปได้ของโปรตีนจะถูกกำหนดโดยการแก้ปัญหาทางเรขาคณิตของระยะทางการแทรกสอดแบบโพลาไรเซชันคู่เป็นวิธีการวิเคราะห์เชิงปริมาณสำหรับการวัดรูปร่างโดยรวมของโปรตีนและการเปลี่ยนแปลงรูปร่างเนื่องจากปฏิกิริยาหรือสิ่งกระตุ้นอื่นๆการดูดกลืนแสงแบบวงกลมเป็นอีกเทคนิคหนึ่งในห้องปฏิบัติการสำหรับการกำหนดองค์ประกอบภายในของแผ่นเบต้า/เกลียวอัลฟาของโปรตีนกล้องจุลทรรศน์อิเล็กตรอนแบบไครโอใช้ในการสร้างข้อมูลโครงสร้างที่มีความละเอียดต่ำเกี่ยวกับโปรตีนเชิงซ้อนขนาดใหญ่มาก รวมถึงไวรัสที่ประกอบขึ้น[ 41 ] : 340–41รูป แบบที่เรียกว่าผลึกศาสตร์อิเล็กตรอนสามารถสร้างข้อมูลที่มีความละเอียดสูงได้ในบางกรณี โดยเฉพาะอย่างยิ่งสำหรับผลึกสองมิติของโปรตีนเยื่อหุ้มเซลล์[ 88 ]โครงสร้างที่แก้ไขแล้วมักจะถูกฝากไว้ในProtein Data Bank (PDB) ซึ่งเป็นแหล่งข้อมูลที่เข้าถึงได้ฟรี ซึ่งสามารถรับข้อมูลโครงสร้างเกี่ยวกับโปรตีนหลายพันชนิดในรูปแบบพิกัดคาร์ทีเซียนสำหรับแต่ละอะตอมในโปรตีนได้[ 89 ]
ลำดับยีนเป็นที่รู้จักมากกว่าโครงสร้างโปรตีนมาก นอกจากนี้ ชุดโครงสร้างที่ได้รับการแก้ไขยังเอนเอียงไปทางโปรตีนที่สามารถนำไปใช้ภายใต้เงื่อนไขที่จำเป็นในการตกผลึกด้วยรังสีเอกซ์ ได้ง่าย ซึ่งเป็นหนึ่งในวิธีการกำหนดโครงสร้างหลัก โดยเฉพาะอย่างยิ่ง โปรตีนทรงกลมนั้นค่อนข้างง่ายต่อการตกผลึกเพื่อเตรียมสำหรับการตกผลึกด้วยรังสีเอกซ์ ในทางตรงกันข้าม โปรตีนเยื่อหุ้มเซลล์และโปรตีนเชิงซ้อนขนาดใหญ่นั้นยากต่อการตกผลึกและมีจำนวนน้อยใน PDB [ 90 ] โครงการ จีโนมิกส์เชิงโครงสร้างได้พยายามแก้ไขข้อบกพร่องเหล่านี้โดยการแก้ปัญหาโครงสร้างที่เป็นตัวแทนของกลุ่มพับหลักอย่างเป็นระบบ วิธี การทำนายโครงสร้างโปรตีนพยายามที่จะสร้างโครงสร้างที่เป็นไปได้สำหรับโปรตีนที่โครงสร้างยังไม่ได้รับการกำหนดโดยการทดลอง[ 91 ]
การทำนายโครงสร้าง

การทำนายโครงสร้างโปรตีนซึ่งเป็นส่วนเสริมของสาขาจีโนมิกส์เชิงโครงสร้างนั้นพัฒนาแบบจำลองทางคณิตศาสตร์ ที่มีประสิทธิภาพ ของโปรตีนเพื่อทำนายการก่อตัวระดับโมเลกุลในทางทฤษฎีโดยใช้การคำนวณ แทนที่จะตรวจจับโครงสร้างด้วยการสังเกตในห้องปฏิบัติการ[ 92 ]การทำนายโครงสร้างที่ประสบความสำเร็จมากที่สุด ซึ่งรู้จักกันในชื่อ การสร้างแบบจำลองความคล้ายคลึง กัน (homology modeling ) อาศัยการมีอยู่ของโครงสร้าง "แม่แบบ" ที่มีความคล้ายคลึงกันของลำดับกับโปรตีนที่กำลังสร้างแบบจำลอง เป้าหมายของจีโนมิกส์เชิงโครงสร้างคือการให้การแสดงผลที่เพียงพอในโครงสร้างที่แก้ไขแล้วเพื่อสร้างแบบจำลองโครงสร้างส่วนใหญ่ที่เหลืออยู่[ 93 ]แม้ว่าการสร้างแบบจำลองที่แม่นยำยังคงเป็นความท้าทายเมื่อมีเพียงโครงสร้างแม่แบบที่เกี่ยวข้องห่างไกลเท่านั้น แต่ก็มีการเสนอแนะว่าการจัดเรียงลำดับเป็นคอขวดในกระบวนการนี้ เนื่องจากสามารถสร้างแบบจำลองที่ค่อนข้างแม่นยำได้หากทราบการจัดเรียงลำดับที่ "สมบูรณ์แบบ" [ 94 ]วิธีการทำนายโครงสร้างหลายวิธีได้ถูกนำมาใช้เพื่อแจ้งให้ทราบถึงสาขาวิศวกรรมโปรตีน ที่กำลังเกิดขึ้นใหม่ ซึ่งมีการออกแบบโครงสร้างพับโปรตีนแบบใหม่แล้ว[ 95 ]โปรตีนจำนวนมาก (ในยูคาริโอตประมาณ 33%) ประกอบด้วยส่วนที่ไม่มีโครงสร้างขนาดใหญ่แต่มีฟังก์ชันทางชีวภาพ และสามารถจัดประเภทเป็นโปรตีนที่มีโครงสร้างไม่เป็นระเบียบโดยเนื้อแท้การทำนายและวิเคราะห์ความไม่เป็นระเบียบของโปรตีนเป็นส่วนสำคัญของการกำหนดลักษณะโครงสร้างของโปรตีน[ 96 ]
การจำลองกระบวนการพลวัตในคอมพิวเตอร์
ปัญหาการคำนวณที่ซับซ้อนกว่าคือการทำนายปฏิสัมพันธ์ระหว่างโมเลกุล เช่น ในการเชื่อมต่อโมเลกุล [ 97 ] การ พับโปรตีนปฏิสัมพันธ์ระหว่างโปรตีนกับโปรตีนและปฏิกิริยาทางเคมี แบบจำลองทางคณิตศาสตร์เพื่อจำลองกระบวนการไดนามิกเหล่านี้เกี่ยวข้องกับกลศาสตร์โมเลกุลโดยเฉพาะอย่างยิ่งพลศาสตร์โมเลกุลในส่วนนี้ การจำลอง ในคอมพิวเตอร์ได้ค้นพบการพับของโดเมนโปรตีน α-helical ขนาดเล็ก เช่นส่วนหัวของวิลลิน[ 98 ]โปรตีนเสริมของ HIV [ 99 ]และวิธีการแบบไฮบริดที่ผสมผสานพลศาสตร์โมเลกุลมาตรฐานกับ คณิตศาสตร์ กลศาสตร์ควอนตัมได้สำรวจสถานะอิเล็กตรอนของโรดอปซิน[ 100 ]
นอกเหนือจากพลศาสตร์โมเลกุลแบบคลาสสิกแล้ว วิธี การพลศาสตร์ควอนตัมยังช่วยให้สามารถจำลองโปรตีนในรายละเอียดระดับอะตอมด้วยคำอธิบายที่แม่นยำของผลกระทบทางกลศาสตร์ควอนตัม ตัวอย่างเช่น วิธี การ Hartree แบบหลายชั้นหลายการกำหนดค่าที่ขึ้นอยู่กับเวลาและวิธีการสมการการเคลื่อนที่แบบลำดับชั้นซึ่งถูกนำไปใช้กับคริปโตโครมของพืช[ 101 ]และคอมเพล็กซ์การเก็บเกี่ยวแสงของแบคทีเรีย[ 102 ]ตามลำดับ การจำลองระบบขนาดชีวภาพทั้งแบบควอนตัมและแบบคลาสสิกนั้นต้องการการคำนวณอย่างมาก ดังนั้น โครงการริเริ่ม การคำนวณแบบกระจายเช่น โครงการ Folding@homeจึงอำนวยความสะดวกในการสร้างแบบจำลองโมเลกุลโดยใช้ประโยชน์จากความก้าวหน้าในการประมวลผลแบบขนานGPU และ เทคนิคMonte Carlo [ 103 ] [ 104 ]
การวิเคราะห์ทางเคมี
ปริมาณไนโตรเจนทั้งหมดของสารอินทรีย์ส่วนใหญ่เกิดจากกลุ่มอะมิโนในโปรตีนไนโตรเจนเคเจลดาลทั้งหมด (TKN) เป็นการวัดไนโตรเจนที่ใช้กันอย่างแพร่หลายในการวิเคราะห์น้ำเสีย ดิน อาหาร อาหารสัตว์ และสารอินทรีย์โดยทั่วไป ดังที่ชื่อบ่งบอกวิธีการเคเจลดาลถูกนำมาใช้ มีวิธีการที่ละเอียดอ่อนกว่านี้[ 105 ] [ 106 ]
การย่อยอาหาร

ในกรณีที่ไม่มีตัวเร่งปฏิกิริยา โปรตีนจะไฮโดรไลซิสได้ ช้า [ 107 ]การสลายโปรตีนเป็นเปปไทด์ขนาดเล็กและกรดอะมิโน ( โปรตีโอไลซิส ) เป็นขั้นตอนหนึ่งในการย่อยอาหารผลิตภัณฑ์จากการสลายเหล่านี้จะถูกดูดซึมในลำไส้เล็ก[ 108 ]การไฮโดรไลซิสของโปรตีนอาศัยเอนไซม์ที่เรียกว่าโปรตีเอสหรือเปปติเดส โปรตีเอสซึ่งเป็นโปรตีนชนิดหนึ่ง มีหลายประเภทตามพันธะเปปไทด์ที่พวกมันตัด รวมถึงแนวโน้มที่จะตัดพันธะเปปไทด์ที่ปลายของโปรตีน (เอ็กโซเปปติเดส) เทียบกับพันธะเปปไทด์ที่อยู่ภายในของโปรตีน (เอนโดเปปติเดส) [ 109 ]เปปซินเป็นเอนโดเปปติเดสในกระเพาะอาหาร หลังจากกระเพาะอาหาร ตับอ่อนจะหลั่งโปรตีเอสอื่นๆ เพื่อให้การไฮโดรไลซิสเสร็จสมบูรณ์ ซึ่งรวมถึงทริปซินและไคโมทริปซิน[ 110 ]
การไฮโดรไลซิสโปรตีนถูกนำมาใช้ในเชิงพาณิชย์เพื่อผลิตกรดอะมิโนจากแหล่งโปรตีนจำนวนมาก เช่น เลือดป่น ขนสัตว์ เคราติน วัสดุเหล่านี้จะถูกบำบัดด้วยกรดไฮโดรคลอริก ร้อน ซึ่งส่งผลให้เกิดการไฮโดรไลซิสของพันธะเปปไทด์[ 111 ]
คุณสมบัติทางกล
คุณสมบัติเชิงกลของโปรตีนมีความหลากหลายสูงและมักเป็นหัวใจสำคัญของหน้าที่ทางชีวภาพ เช่น ในกรณีของโปรตีนอย่างเคราตินและคอลลาเจน [ 112 ] ตัวอย่างเช่น ความสามารถของเนื้อเยื่อกล้ามเนื้อในการขยายและหดตัวอย่างต่อเนื่องนั้นเชื่อมโยงโดยตรงกับคุณสมบัติความยืดหยุ่นขององค์ประกอบโปรตีนพื้นฐาน[ 113 ] [ 114 ]นอกเหนือจากโปรตีนเส้นใยแล้ว พลวัตเชิงโครงสร้างของเอนไซม์[ 115 ]และโครงสร้างของเยื่อหุ้มชีวภาพรวมถึงหน้าที่ทางชีวภาพอื่นๆ ล้วนถูกควบคุมโดยคุณสมบัติเชิงกลของโปรตีน นอกเหนือจากบริบททางชีวภาพแล้ว คุณสมบัติเชิงกลที่เป็นเอกลักษณ์ของโปรตีนหลายชนิด พร้อมกับความยั่งยืนเมื่อเทียบกับพอลิเมอร์สังเคราะห์ทำให้โปรตีนเป็นเป้าหมายที่น่าสนใจสำหรับการออกแบบวัสดุรุ่นต่อไป[ 116 ] [ 117 ]
โมดูลัสของยัง (Young's modulus ) Eคำนวณจากความเค้นตามแนวแกนσ หาร ด้วยความเครียดที่เกิดขึ้นεเป็นตัววัดความแข็ง สัมพัทธ์ ของวัสดุ ในบริบทของโปรตีน ความแข็งนี้มักมีความสัมพันธ์โดยตรงกับหน้าที่ทางชีวภาพ ตัวอย่างเช่นคอลลาเจนซึ่งพบในเนื้อเยื่อเกี่ยวพันกระดูกและกระดูกอ่อนและเคราติน ซึ่งพบในเล็บ กรงเล็บ และเส้นผม มีความแข็งที่สังเกตได้สูงกว่า อีลาสติ น หลายลำดับ[ 118 ]ซึ่งเชื่อกันว่าทำให้โครงสร้างต่างๆ เช่นหลอดเลือดเนื้อเยื่อปอดและเนื้อเยื่อกระเพาะปัสสาวะ มีความยืดหยุ่น [ 119 ] [ 120 ]เมื่อเปรียบเทียบกับสิ่งนี้โปรตีนทรงกลมเช่นอัลบูมินในซีรั่มวัวซึ่งลอยอยู่ในไซโตซอล ได้ค่อนข้างอิสระและ มักทำหน้าที่เป็นเอนไซม์ (และจึงมีการเปลี่ยนแปลงโครงสร้างบ่อยครั้ง) มีค่าโมดูลัสของยังที่ต่ำกว่ามาก[ 121 ] [ 122 ]
โมดูลัสของยัง (Young's modulus) ของโปรตีนเดี่ยวสามารถหาได้จาก การจำลอง พลศาสตร์โมเลกุลโดยใช้ฟิลด์แรงอะตอม เช่นCHARMMหรือGROMOSหรือฟิลด์แรงแบบหยาบ เช่น Martini [ 123 ]โมเลกุลโปรตีนเดี่ยวสามารถยืดได้ด้วยแรงแกนเดียว ในขณะที่การยืดที่เกิดขึ้นจะถูกบันทึกเพื่อคำนวณความเครียด[ 124 ] [ 125 ]ในทางทดลอง วิธีการต่างๆ เช่นกล้องจุลทรรศน์แรงอะตอมสามารถนำมาใช้เพื่อให้ได้ข้อมูลที่คล้ายกัน[ 126 ]พลศาสตร์ภายในของโปรตีนเกี่ยวข้องกับการเสียรูปยืดหยุ่นและพลาสติกที่ละเอียดอ่อนซึ่งเกิดจาก แรง หนืดซึ่งสามารถตรวจสอบได้ด้วยเทคนิค นา โนรีโอโลยี[ 127 ]
ในระดับมหภาค ค่าโมดูลัสของยัง (Young's modulus) ของโครงข่ายโปรตีนที่เชื่อมโยงกันสามารถหาได้จากการทดสอบทางกลแบบ ดั้งเดิม ค่าที่สังเกตได้จากการทดลองสำหรับโปรตีนบางชนิดแสดงไว้ด้านล่าง
| โปรตีน | โปรตีนคลาส | โมดูลัสของยัง |
|---|---|---|
| เคราติน (แบบเชื่อมโยง) | เส้นใย | 1.5–10 GPa [ 128 ] |
| อีลาสติน (แบบเชื่อมโยงข้าม) | เส้นใย | 1 MPa [ 118 ] |
| ไฟบริน (แบบเชื่อมโยง) | เส้นใย | 1–10 MPa [ 118 ] |
| คอลลาเจน (แบบเชื่อมโยงข้าม) | เส้นใย | 5–7.5 GPa [ 118 ] [ 129 ] |
| เรซิลิน (แบบเชื่อมโยง) | เส้นใย | 1–2 MPa [ 118 ] |
| อัลบูมินจากซีรั่มวัว (แบบเชื่อมโยงข้าม) | ทรงกลม | 2.5–15 kPa [ 121 ] |
| โปรตีนเยื่อหุ้มเซลล์ชั้นนอกแบบ β-barrel | เมมเบรน | 20–45 GPa [ 130 ] |
ดูเพิ่มเติม
- การกำจัดโปรตีน – เทคนิคในการวิจัยวัสดุที่มีชีวิต
- โปรตีนที่จับกับดีเอ็นเอ – โปรตีนที่จับกับดีเอ็นเอ
- ดัชนีบทความที่เกี่ยวข้องกับโปรตีน
- อินทีน
- รายชื่อโปรตีน
- โมเลกุลขนาด ใหญ่ – โมเลกุลที่มีขนาดใหญ่มาก
- วิวัฒนาการของโปรตีน – การศึกษาการเปลี่ยนแปลงของดีเอ็นเอและอาร์เอ็นเอเมื่อเวลาผ่านไป
- พื้นที่ลำดับโปรตีน – การแสดงลำดับทางพันธุกรรมที่เป็นไปได้
- ซูเปอร์แฟมิลีโปรตีน – การจัดกลุ่มโปรตีน
- ภาวะโปรตีนเป็นพิษ – การสะสมของของเสียจากกระบวนการเผาผลาญเนื่องจากความผิดปกติของไต
- โรคที่เกิดจากโครงสร้างโปรตีน ผิดปกติ – โรคที่เกิดจากโครงสร้างโปรตีนที่ผิดปกติ
- โปรทีโอพีเดีย – สารานุกรมสามมิติของโปรตีนและโมเลกุลอื่นๆ
อ่านเพิ่มเติม
- ตำราเรียน
- Branden C, Tooze J (1999). บทนำเกี่ยวกับโครงสร้างโปรตีน . นิวยอร์ก: สำนักพิมพ์ Garland. ISBN 978-0-8153-2305-1.
- Murray RF, Harper HW, Granner DK, Mayes PA, Rodwell VW (2006). Harper's Illustrated Biochemistry . นิวยอร์ก: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9.
- Van Holde KE, Mathews CK (1996). ชีวเคมี . เมนโลพาร์ค รัฐแคลิฟอร์เนีย: Benjamin/Cummings Pub. Co., Inc. ISBN 978-0-8053-3931-4.
- ประวัติศาสตร์
- Tanford C, Reynolds JA (2001). หุ่นยนต์แห่งธรรมชาติ: ประวัติศาสตร์ของโปรตีน . อ็อกซ์ฟอร์ด นิวยอร์ก: สำนักพิมพ์มหาวิทยาลัยอ็อกซ์ฟอร์ด สหรัฐอเมริกา. ISBN 978-0-19-850466-5.
ลิงก์ภายนอก
 คำจำกัดความจากวิกิพีเดีย
คำจำกัดความจากวิกิพีเดีย สื่อจากคอมมอนส์
สื่อจากคอมมอนส์
ฐานข้อมูลและโครงการ
- ฐานข้อมูลโปรตีน NCBI Entrez
- ฐานข้อมูลโครงสร้างโปรตีน NCBI
- ฐานข้อมูลอ้างอิงโปรตีนของมนุษย์
- โปรตีนของมนุษย์
- Folding@Home (มหาวิทยาลัยสแตนฟอร์ด) เก็บถาวรเมื่อ 2012-09-08 ที่Wayback Machine
- ฐานข้อมูลโปรตีนในยุโรป (ดูเพิ่มเติมที่PDBeQuipsบทความสั้น ๆ และบทช่วยสอนเกี่ยวกับโครงสร้าง PDB ที่น่าสนใจ)
- ศูนย์วิจัยความร่วมมือด้านชีวสารสนเทศเชิงโครงสร้าง (ดูเพิ่มเติมที่โมเลกุลประจำเดือนที่เก็บถาวรเมื่อวันที่ 24 กรกฎาคม 2020 ที่Wayback Machineซึ่งนำเสนอเรื่องราวสั้น ๆ เกี่ยวกับโปรตีนที่คัดเลือกจาก PDB)
- Proteopedia – ชีวิตในรูปแบบ 3 มิติ : โมเดล 3 มิติที่หมุนได้และซูมได้ พร้อมคำอธิบายประกอบแบบวิกิสำหรับโครงสร้างโมเลกุลของโปรตีนทุกชนิดที่รู้จัก
- UniProt แหล่งข้อมูลโปรตีนสากล
บทเรียนและเว็บไซต์เพื่อการศึกษา
- "ความรู้เบื้องต้นเกี่ยวกับโปรตีน"จากHOPES (โครงการเผยแพร่ความรู้เกี่ยวกับโรคฮันติงตันแห่งมหาวิทยาลัยสแตนฟอร์ด)
- โปรตีน: กระบวนการสร้างจนถึงการสลายตัว – ห้องสมุดเสมือนจริงด้านชีวเคมีและชีววิทยาของเซลล์
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ โปรตีน
โปรตีน เป็น โมเลกุลชีวภาพ ขนาดใหญ่ และ โมเลกุลขนาดมหึมา ที่ประกอบด้วยสายโซ่ยาวของกรดอะมิโนหนึ่งสายหรือมากกว่า โปรตีน ทำ หน้าที่มากมายในสิ่งมีชีวิต รวมถึง การเร่งปฏิกิริยาทางเม...
การค้นพบและการศึกษาเบื้องต้น
โปรตีนได้รับการศึกษาและรู้จักมาตั้งแต่ศตวรรษที่ 17 โดย Antoine Fourcroy และคนอื่นๆ [ 1 ] [ 2 ] ซึ่งมักเรียกโดยรวมว่า " อัลบูมิน " หรือ "สารอัลบูมิน" ( Eiweisskörper ในภาษาเยอรมัน) [ 2 ] ตัวอย่างเช่น กลูเตน...
โพลีเปปไทด์
ความเข้าใจเกี่ยวกับโปรตีนในฐานะพอ ลิเปปไทด์ หรือสายโซ่ของกรดอะมิโน เกิดขึ้นจากผลงานของ Franz Hofmeister และ Hermann Emil Fischer ในปี พ.ศ.
โครงสร้าง
ด้วยการพัฒนาของ ผลึกศาสตร์รังสีเอกซ์ ทำให้สามารถกำหนดโครงสร้างโปรตีนรวมถึงลำดับของโปรตีนได้ [ 25 ] โครงสร้างโปรตีน แรกที่ได้รับการแก้ไขคือ ฮีโมโกลบิน โดย Max Perutz และ ไมโอโกลบิน โดย John Kendrew ในปี 1958 [ 26 ] [ 27 ]...