สารประกอบแอนะล็อกของกรดนิวคลีอิกคือสารประกอบที่มีโครงสร้างคล้ายคลึงกับอาร์เอ็นเอและดีเอ็นเอ ที่เกิดขึ้นตามธรรมชาติ โดยการแทนที่ส่วนประกอบของน้ำตาล ฟอสเฟต และนิวคลีโอเบส สารประกอบแอนะล็อกเหล่านี้ถูกนำมาใช้ในทางการแพทย์และการวิจัยทางชีววิทยาโมเลกุล
ภาพรวม
{kind=link}

กรดนิวคลีอิกคือสายโซ่ของนิวคลีโอไทด์ ซึ่งประกอบด้วยสามส่วน ได้แก่ แกนหลักที่ประกอบด้วยน้ำตาลเพนโทสของไรโบสหรือดีออกซีไรโบสซึ่งเชื่อมต่อกันด้วย หมู่ ฟอสเฟตและหนึ่งในสี่นิวคลีโอเบส อะนาล็อกอาจมีส่วนใดส่วนหนึ่งที่เปลี่ยนแปลงไปโดยทั่วไป นิวคลีโอเบสอะนาล็อกจะมีคุณสมบัติการจับคู่เบสและการเรียงซ้อนเบสที่แตกต่างกัน ตัวอย่างเช่น เบสสากล ซึ่งสามารถจับคู่กับเบสแคโนนิคัลทั้งสี่ และอะนาล็อกแกนหลักฟอสเฟต-น้ำตาล เช่นPNAซึ่งมีผลต่อคุณสมบัติของสายโซ่ (PNA สามารถสร้างเกลียวสามชั้น ได้ )
อะนาล็อกกระดูกสันหลัง
อะนาล็อกของกรดนิวคลีอิกอาจมีการทดแทนโครงสร้างหลักให้กับส่วนประกอบของน้ำตาลหรือฟอสเฟตหรือทั้งสองอย่าง
ทฤษฎีโพลีอิเล็กโทรไลต์ของยีนเสนอว่าโมเลกุลทางพันธุกรรมต้องมีโครงสร้างหลักที่มีประจุจึงจะทำงานได้
น้ำตาลอะนาล็อก
หมู่ไฮด รอกซี 2' ของไรโบสทำปฏิกิริยากับหมู่ไฮดรอกซี 3' ที่เชื่อมกับฟอสเฟต ทำให้อาร์เอ็นเอไม่เสถียรจนไม่สามารถนำไปใช้หรือสังเคราะห์ได้อย่างน่าเชื่อถือ เพื่อแก้ไขปัญหานี้ จึงสามารถใช้อะนาล็อกของไรโบสได้ อะนาล็อกของอาร์เอ็นเอที่พบมากที่สุด ได้แก่ อาร์เอ็นเอที่แทนที่ด้วย 2'-O-เมทิล, กรดนิวคลีอิกแบบล็อก (LNA) หรือกรดนิวคลีอิกแบบบริดจ์ (BNA) และกรดนิวคลีอิกแบบเพปไทด์ (PNA) รวมถึงกรดนิวคลีอิกไกลคอล (GNA) กรดนิวคลีอิกแบบทรีส (TNA) และ กรดนิวคลีอิกแบบเฮกซิ ทอล (HNA)
แม้ว่าโอลิโกนิวคลีโอไทด์เหล่านี้จะมีโครงสร้างน้ำตาลที่แตกต่างกัน หรือในกรณีของ PNA จะมีกรดอะมิโนตกค้างแทนที่ไรโบสฟอสเฟต แต่พวกมันยังคงจับกับ RNA หรือ DNA ตามการจับคู่ของวัตสันและคริก ในขณะที่ยังคงต้านทานต่อกิจกรรมของนิวคลีเอส พวกมันไม่สามารถสังเคราะห์ด้วยเอนไซม์ได้ และสามารถทำได้โดยการสังเคราะห์โดยใช้กลยุทธ์ฟอสโฟราไมไดต์หรือสำหรับ PNA ใช้วิธีสังเคราะห์เปปไทด์อื่น ๆ เท่านั้น
ไดดีออกซีนิวคลีโอไทด์ถูกนำมาใช้ในการหาลำดับเบสนิวคลีโอไซด์ไตรฟอสเฟตเหล่านี้มีน้ำตาลไดดีออกซีไรโบสที่ไม่ใช่น้ำตาลปกติ ซึ่งขาดหมู่ไฮดรอกซิล 3' ซึ่งปกติพบในดีเอ็นเอ ดังนั้นจึงไม่สามารถจับกับเบสตัวถัดไปได้ การขาดหมู่ไฮดรอกซิล 3' ทำให้ปฏิกิริยาลูกโซ่สิ้นสุดลง เนื่องจากดีเอ็นเอโพลีเมอเรสเข้าใจผิดว่าเป็นดีออกซีไรโบนิวคลีโอไทด์ทั่วไป
อะนาล็อกที่สิ้นสุดสายอีกชนิดหนึ่งที่ไม่มี 3' ไฮดรอกซิลและเลียนแบบอะดีโนซีนเรียกว่าคอร์ไดเซพินคอร์ไดเซพินเป็นยาต้านมะเร็งที่มุ่งเป้าไปที่ การจำลองแบบของ อาร์เอ็นเอ อะนาล็อกอีกชนิดหนึ่งในการหาลำดับเบสคือ 7-ดีซา-จีทีพี ซึ่งเป็นอะนาล็อกของนิวคลีโอเบส และใช้ในการหาลำดับเบสของบริเวณที่มี CG สูง แทนที่จะใช้ 7-ดีซา-เอทีพี เรียกว่าทูเบอร์ซิดินซึ่งเป็นยาปฏิชีวนะ
กรดนิวคลีอิกของสัตว์ถือเป็นเสาหลักประการหนึ่งของชีววิทยาสัตว์ซึ่งเป็นการออกแบบรูปแบบชีวิตใหม่ตามธรรมชาติโดยอาศัยชีวเคมีทางเลือก
อะนาล็อกฟอสเฟต
สารอนาล็อกฟอสเฟต ได้แก่กรดนิวคลีอิกฟอสโฟโรไทโอเอตและกรดนิวคลีอิกฟอส โฟโรไดอะมิเดต
ทั้งคู่
{kind=link}

กรดนิวคลีอิก มอร์โฟลิโนจะแทนที่น้ำตาลด้วย ส่วนประกอบของ มอร์โฟลีนและแทนที่ฟอสเฟตด้วยส่วนประกอบของฟอสโฟโรไดอะมิเดต
Miravirsenเป็นยาต้านความรู้สึก ที่มีทั้ง ส่วนประกอบน้ำตาล กรดนิวคลีอิกที่ถูกล็อกและกลุ่มฟอสโฟโรไทโอเอต
อะนาล็อกฐาน
เบสที่เกิดขึ้นตามธรรมชาติสามารถแบ่งออกได้เป็น 2 ประเภทตามโครงสร้าง:
- ไพริมิดีนเป็นเฮเทอโรไซคลิกหกเหลี่ยมที่มีอะตอมไนโตรเจนอยู่ในตำแหน่งที่ 1 และ 3
- พิวรีนเป็นสารแบบไบไซคลิก ประกอบด้วยไพริมิดีนที่เชื่อมกับวงแหวนอิมีดาโซล
นิวคลีโอไทด์เทียม ( คู่เบสที่ไม่เป็นธรรมชาติ (UBPs) ชื่อd5SICS UBPและdNaM UBP ) ถูกแทรกเข้าไปในดีเอ็นเอของแบคทีเรีย แต่ยีนเหล่านี้ไม่ได้เป็นแม่แบบของ mRNA หรือกระตุ้นการสังเคราะห์โปรตีน นิวคลีโอไทด์เทียมมีวงแหวนอะโรมาติกสองวงที่เชื่อมติดกัน ซึ่งก่อตัวเป็นสารประกอบเชิงซ้อน (d5SICS–dNaM) ที่เลียนแบบคู่เบสตามธรรมชาติ (dG–dC)
สารก่อกลายพันธุ์
หนึ่งในเบสอะนาล็อกที่พบมากที่สุดคือ5-โบรโมราซิล (5BU) ซึ่งเป็นเบสผิดปกติที่พบใน BrdU ซึ่งเป็นเบสอะนาล็อกของนิวคลีโอไทด์ที่ก่อให้เกิดการกลายพันธุ์ เมื่อนิวคลีโอไทด์ที่มี 5-โบรโมราซิลถูกรวมเข้ากับดีเอ็นเอ มันมีแนวโน้มที่จะจับคู่กับอะดีนีน อย่างไรก็ตาม มันสามารถเปลี่ยนไปเป็นไอโซเมอร์ อื่น ที่จับคู่กับ นิวคลี โอเบส อื่นได้เอง คือกัวนีนหากสิ่งนี้เกิดขึ้นในระหว่างการจำลองดีเอ็นเอ กัวนีนจะถูกแทรกเข้าไปเป็นเบสอะนาล็อกตรงข้าม และในการจำลองดีเอ็นเอครั้งต่อไป กัวนีนนั้นจะจับคู่กับไซโทซีน ส่งผลให้เกิดการเปลี่ยนแปลงในคู่เบสหนึ่งของดีเอ็นเอ โดยเฉพาะอย่างยิ่งการกลายพันธุ์แบบทรานซิชัน [
นอกจากนี้ กรดไนตรัส (HNO 2 ) ยังเป็นสารก่อกลายพันธุ์ที่มีฤทธิ์รุนแรงซึ่งออกฤทธิ์ต่อดีเอ็นเอทั้งแบบจำลองและไม่จำลอง กรดไนตรัสสามารถทำให้เกิดการดีอะมิเนชันของหมู่อะมิโนของอะดีนีน กัวนีน และไซโทซีนได้ อะดีนีนจะถูกดีอะมิเนชันไปเป็นไฮโปแซนทีน ซึ่งเบสจะจับคู่กับไซโทซีนแทนที่จะเป็นไทมีน ไซโทซีนจะถูกดีอะมิเนชันไปเป็นยูราซิล ซึ่งเบสจะจับคู่กับอะดีนีนแทนที่จะเป็นกัวนีน การดีอะมิเนชันของกัวนีนไม่ก่อให้เกิดการกลายพันธุ์ การกลายพันธุ์ที่เกิดจากกรดไนตรัสยังสามารถเหนี่ยวนำให้กลายพันธุ์กลับไปเป็นไวลด์ไทป์ได้อีกด้วย
ฟลูออโรโฟร์
{kind=link}

โดยทั่วไปฟลูออโรฟอร์ (เช่นโรดามีนหรือฟลูออเรสซีน ) จะเชื่อมต่อกับวงแหวนที่เชื่อมต่อกับน้ำตาล (ในพารา) ผ่านแขนที่ยืดหยุ่น ซึ่งสันนิษฐานว่ายื่นออกมาจากร่องหลักของเกลียว เนื่องจากนิวคลีโอไทด์ที่เชื่อมต่อกับแอดดักต์ขนาดใหญ่ เช่น ฟลูออโรฟอร์ มีความสามารถในการประมวลผลต่ำ โดย [Taq polymerase] ลำดับนี้จึงมักถูกคัดลอกโดยใช้นิวคลีโอไทด์ที่มีแขน และต่อมาถูกจับคู่กับฟลูออโรฟอร์ที่มีปฏิกิริยา (การติดฉลากทางอ้อม):
- ปฏิกิริยาอะมีน: นิวคลีโอไทด์อะมิโนอัลลิลประกอบด้วยหมู่อะมีนหลักบนตัวเชื่อมที่ทำปฏิกิริยากับสีย้อมที่ไวต่ออะมิโน เช่น สี ย้อมไซยานีนหรืออเล็กซาฟลูออร์ซึ่งมีหมู่อะมิโนที่ไวต่อปฏิกิริยา เช่น ซักซินิมิดิลเอสเทอร์ (NHS) หมู่อะมิโนที่จับคู่เบสจะไม่ได้รับผลกระทบ
- ไทออลที่มีปฏิกิริยา: นิวคลีโอไทด์ที่มีไทออลจะทำปฏิกิริยากับสารฟลูออโรโฟร์ที่เชื่อมโยงกับกลุ่มปฏิกิริยาที่ออกจากกลุ่ม เช่น มาเลอิไมด์
- นิวคลีโอไทด์ที่เชื่อมโยงกับ ไบโอตินอาศัยหลักการติดฉลากทางอ้อมแบบเดียวกัน (และสเตรปตาปิดินเรืองแสง) และใช้ใน ชิป Affymetrix DNA
สารเรืองแสงมีการใช้งานที่หลากหลายในทางการแพทย์และชีวเคมี
2-อะมิโนพิวรีน (2-AP) ซึ่งเป็นสารอนาล็อกเบสเรืองแสงที่ใช้กันทั่วไปและหาซื้อได้ในท้องตลาด มีปริมาณผลผลิตควอนตัมฟลูออเรสเซนซ์สูงในสารละลาย (0.68) ซึ่งลดลงอย่างมาก (ประมาณ 100 เท่าแต่ขึ้นอยู่กับลำดับเบสอย่างมาก) เมื่อรวมเข้ากับกรดนิวคลี อิก ความไวในการปล่อยของ 2-AP ต่อสภาพแวดล้อมโดยรอบนั้นมีข้อดีและน่าสนใจอื่นๆ เช่น 3-MI, 6-MI, 6-MAP, ไพร์โรโล-ดีซี (มีจำหน่ายในท้องตลาดเช่นกัน), อนุพันธ์ของไพร์โรโล-ดีซีที่ดัดแปลงและปรับปรุงแล้ว, เบสที่ดัดแปลงด้วยฟิวแร น และอื่นๆ อีกมากมาย (ดูบทวิจารณ์ล่าสุด) ความไวต่อสภาพแวดล้อมจุลภาคนี้ถูกนำมาใช้ในการศึกษาโครงสร้างและพลวัตของทั้ง DNA และ RNA พลวัตและจลนพลศาสตร์ของปฏิสัมพันธ์ระหว่าง DNA และโปรตีน และการถ่ายโอนอิเล็กตรอนภายใน DNA
กลุ่มของสารอะนาล็อกเบสเรืองแสงที่พัฒนาขึ้นใหม่และน่าสนใจมากซึ่งมีผลผลิตควอนตัมการเรืองแสงที่แทบไม่ไวต่อสภาพแวดล้อมโดยรอบคือกลุ่มไซโตซีนไตรไซคลิก 1,3-Diaza-2-oxophenothiazine, tC มีผลผลิตควอนตัมการเรืองแสงประมาณ 0.2 ทั้งในสายเดี่ยวและสายคู่โดยไม่คำนึงถึงเบสโดยรอบนอกจากนี้ ออกโซโฮโมล็อกของ tC ที่เรียกว่า tC2O ทั้งคู่มีจำหน่ายในเชิงพาณิชย์) 1,3-diaza-2-oxophenoxazine มีผลผลิตควอนตัมที่ 0.2 ในระบบสายคู่อย่างไรก็ตาม มันค่อนข้างไวต่อเบสโดยรอบในสายเดี่ยว (ผลผลิตควอนตัมที่ 0.14–0.41) ผลผลิตควอนตัมที่สูงและเสถียรของเบสอะนาล็อกเหล่านี้ทำให้มีความสว่างมาก และเมื่อรวมกับคุณสมบัติของเบสอะนาล็อกที่ดี (ทำให้โครงสร้างและความเสถียรของดีเอ็นเอแทบจะไม่ถูกรบกวน) เบสอะนาล็อกเหล่านี้จึงมีประโยชน์อย่างยิ่งในการวัดค่าแอนไอโซทรอปิกของฟลูออเรสเซนซ์และ FRET ซึ่งเป็นบริเวณที่เบสอะนาล็อกเรืองแสงอื่นๆ มีความแม่นยำน้อยกว่า นอกจากนี้ ในตระกูลไซโทซีนอะนาล็อกเดียวกันนี้ยังได้พัฒนา เบสอะนาล็อกตัวรับ FRET คือ tC nitro เมื่อรวมกับ tC ในฐานะตัวให้ FRET นี่ถือเป็นเบสอะนาล็อกกรดนิวคลีอิกคู่แรก FRET ที่พัฒนาขึ้น ตัวอย่างเช่น ตระกูล tC ถูกนำมาใช้ในการศึกษาที่เกี่ยวข้องกับกลไกการจับดีเอ็นเอของพอลิเมอเรสและการเกิดพอลิเมอไรเซชันของดีเอ็นเอ
ฐานธรรมชาติที่ไม่ใช่แบบมาตรฐาน
ในเซลล์มีเบสที่ไม่ใช่แบบแคนนอนิคัลอยู่หลายชนิด ได้แก่ หมู่ CpG ในดีเอ็นเอ (มักถูกเมทิลเลชัน) mRNA ที่เป็นยูคาริโอตทั้งหมด (ปิดท้ายด้วยเมทิล-7-กัวโนซีน) และเบสหลายชนิดของ rRNA (ถูกเมทิลเลชัน) บ่อยครั้งที่ tRNA ถูกดัดแปลงอย่างมากหลังการถอดรหัสเพื่อปรับปรุงโครงสร้างหรือการจับคู่เบส โดยเฉพาะอย่างยิ่งในหรือใกล้กับแอนติโคดอน: อิโนซีนสามารถจับคู่เบสกับ C, U และแม้กระทั่งกับ A ได้ ในขณะที่ไทโอยูริดีน (กับ A) มีความจำเพาะมากกว่ายูราซิล (กับพิวรีน) การดัดแปลงเบส tRNA อื่นๆ ที่พบบ่อย ได้แก่ ซูโดยูริดีน (ซึ่งเป็นที่มาของชื่อวง TΨC ), ไดไฮโดรยูริดีน (ซึ่งไม่เรียงซ้อนกันเนื่องจากไม่ใช่อะโรมาติก), ควีโอซีน, ไวโอซีน และอื่นๆ อย่างไรก็ตาม สิ่งเหล่านี้ล้วนเป็นการดัดแปลงเบสปกติและไม่ได้ถูกกำหนดโดยพอลิเมอเรส
การจับคู่เบส
เบสแคโนนิคัลอาจมีหมู่คาร์บอนิลหรือหมู่เอมีนอยู่บนคาร์บอนที่ล้อมรอบอะตอมไนโตรเจนซึ่งอยู่ไกลจากพันธะไกลโคซิดิกมากที่สุด ซึ่งทำให้เบสเหล่านี้จับคู่กัน (การจับคู่เบสแบบวัตสัน-คริก) ผ่านพันธะไฮโดรเจน (เอมีนกับคีโตน, พิวรีนกับไพริมิดีน) อะดีนีนและ 2-อะมิโนอะดีนีนมีหมู่เอมีนหนึ่งหรือสองหมู่ ในขณะที่ไทมีนมีหมู่คาร์บอนิลสองหมู่ และไซโทซีนและกัวนีนเป็นเอมีนและคาร์บอนิลผสมกัน (กลับด้านกัน)
| เบสคู่ธรรมชาติ | |
|---|---|

|

|
| เบสคู่ GC: พิวรีนคาร์บอนิล/เอมีนสร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสามพันธะ กับไพริมิดีนเอมีน/คาร์บอนิล | เบสคู่ AT: พิวรีนเอมีน/- สร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสองพันธะ กับไพริมิดีนคาร์บอนิล/คาร์บอนิล |
{kind=link}
{kind=link}
เหตุผลที่แน่ชัดว่าทำไมจึงมีนิวคลีโอไทด์เพียงสี่ตัวนั้นยังคงเป็นที่ถกเถียงกันอยู่ แต่ยังมีความเป็นไปได้หลายอย่างที่ยังไม่ได้นำมาใช้ นอกจากนี้ อะดีนีนยังไม่ใช่ตัวเลือกที่เสถียรที่สุดสำหรับการจับคู่เบส ใน Cyanophage S-2L จะใช้ ไดอะมิโนพิวรีน (DAP) แทนอะดีนีนเบสไดอะมิโนพิวรีนจับคู่กับไทมีนได้อย่างสมบูรณ์แบบ เนื่องจากมีคุณสมบัติเหมือนกับอะดีนีน แต่มีหมู่เอมีนที่ตำแหน่ง 2 ซึ่งสร้างพันธะไฮโดรเจนภายในโมเลกุล 3 พันธะ ทำให้ไม่มีความแตกต่างที่สำคัญระหว่างเบสคู่ทั้งสองประเภทนี้ (AT อ่อนเทียบกับ CG แข็ง) ความเสถียรที่เพิ่มขึ้นนี้ส่งผลต่อปฏิกิริยาการจับกับโปรตีนที่อาศัยความแตกต่างเหล่านี้ การผสมผสานอื่นๆ ได้แก่:
- ไอโซกัวนีนและไอโซไซโทซีน ซึ่งมีเอมีนและคีโตนที่กลับด้านเมื่อเทียบกับกัวนีนและไซโทซีนมาตรฐาน อาจไม่มีการใช้ไอโซไซโทเมอร์เนื่องจากทอโทเมอร์เป็นปัญหาสำหรับการจับคู่เบส แต่ไอโซไซโทซีนและไอโซไซโทซีนสามารถเพิ่มปริมาณได้อย่างถูกต้องด้วย PCR แม้จะมีเบสหลัก 4 ตัว
- ไดอะมิโนไพริมิดีนและแซนทีน ซึ่งจับตัวกันคล้ายกับ 2-อะมิโนอะดีนีนและไทมีน แต่มีโครงสร้างแบบกลับด้าน ไม่ใช้คู่นี้เนื่องจากแซนทีนเป็นสารดีอะมิเนชัน
| การจัดเตรียมเบสแพร์ที่ไม่ได้ใช้ | ||
|---|---|---|

|
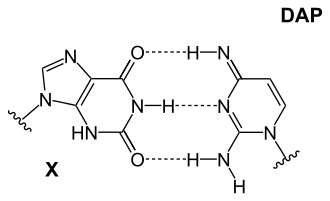
|

|
| เบส DAP-T: พิวรีนเอมีน/เอมีนสร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสามพันธะ กับไพริมิดีนคีโตน/คีโตน | เบส X-DAP: พิวรีนคีโตน/คีโตนสร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสามพันธะ กับไพริมิดีนเอมีน/เอมีน | เบส iG-iC: พิวรีนเอมีน/คีโตนสร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสามพันธะ กับไพริมิดีนคีโตน/เอมีน |
{kind=link}
{kind=link}
{kind=link}
อย่างไรก็ตาม โครงสร้าง DNA ที่ถูกต้องสามารถเกิดขึ้นได้แม้เบสจะไม่จับคู่กันผ่านพันธะไฮโดรเจน กล่าวคือ เบสจับคู่กันเนื่องจากคุณสมบัติไม่ชอบน้ำ ดังที่การศึกษาได้แสดงให้เห็นกับไอโซสเตอเรส ของ DNA (อะนาล็อกที่มีจำนวนอะตอมเท่ากัน) เช่น อะนาล็อกของไทมีน 2,4-ไดฟลูออโรโทลูอีน (F) หรืออะนาล็อกของอะดีนีน 4-เมทิลเบนซิมิดาโซล (Z) คู่ที่ไม่ชอบน้ำอีกทางเลือกหนึ่งอาจเป็นไอโซควิโนลีนและไพร์โรโล[2,3-b]ไพริดีน
เบสคู่อื่น ๆ ที่น่าสังเกต:
- เบสเรืองแสงหลายชนิดก็ถูกสร้างขึ้นเช่นกัน เช่น คู่เบส 2-อะมิโน-6-(2-ไทอีนิล)พิวรีนและไพรโรล-2-คาร์บาลดีไฮด์
- เบสที่ประสานงานกับโลหะ เช่น การจับคู่ระหว่างไพริดีน-2,6-ไดคาร์บอกซิเลต (ลิแกนด์ไตรเดนเตต) และไพริดีน (ลิแกนด์โมโนเดนเตต) ผ่านการประสานงานแบบระนาบสี่เหลี่ยมไปยังไอออนทองแดงที่อยู่ตรงกลาง
- เบสสากลอาจจับคู่กับเบสอื่น ๆ ได้อย่างไม่มีการแบ่งแยก แต่โดยทั่วไปแล้วเบสเหล่านี้จะลดอุณหภูมิการหลอมเหลวของลำดับเบสลงอย่างมาก ตัวอย่างเช่น อนุพันธ์ 2'-ดีออกซีอิโนซีน (ไฮโปแซนทีนดีออกซีนิวคลีโอไทด์) อะนาล็อกไนโตรอะโซล และเบสอะโรมาติกแบบไม่มีพันธะไฮโดรเจนที่ไม่ชอบน้ำ (ผลกระทบจากการเรียงซ้อนที่รุนแรง) เบสเหล่านี้ใช้เป็นหลักฐานแนวคิด และโดยทั่วไปจะไม่ถูกนำมาใช้ในไพรเมอร์ที่เสื่อมสภาพ (ซึ่งเป็นส่วนผสมของไพรเมอร์)
- จำนวนคู่เบสที่เป็นไปได้จะเพิ่มขึ้นเป็นสองเท่าเมื่อ พิจารณา xDNA xDNA ประกอบด้วยเบสที่ขยายออก ซึ่งมีการเพิ่มวงเบนซีนเข้าไป ซึ่งอาจจับคู่กับเบสมาตรฐาน ส่งผลให้มีคู่เบสที่เป็นไปได้เพิ่มเติมอีกสี่คู่ (xA-T, xT-A, xC-G, xG-C) โดยมีเบสแปดตัว (หรือ 16 ตัว หากใช้การจัดเรียงที่ไม่ได้ใช้) อีกรูปแบบหนึ่งของเบสที่เติมเบนซีนเข้าไปคือ yDNA ซึ่งเบสจะถูกขยายออกโดยเบนซีน
| เบสคู่ใหม่ที่มีคุณสมบัติพิเศษ | ||
|---|---|---|

|

|

|
| เบส FZ: เมทิลเบนซิมิดาโซลไม่สร้างพันธะไฮโดรเจน ระหว่างโมเลกุล กับโทลูอีน F/F | เบส S-Pa: พิวรีนไทเอนิล/เอมีนสร้างพันธะไฮโดรเจน ระหว่างโมเลกุลสามพันธะ กับไพร์โรล-/คาร์บัลดีไฮด์ | ฐาน xA-T: พันธะเดียวกับ AT |
{kind=link}
{kind=link}
{kind=link}
เบสคู่โลหะ
ในการจับคู่เบสโลหะ พันธะไฮโดรเจนวัตสัน-คริกจะถูกแทนที่ด้วยอันตรกิริยาระหว่างไอออนโลหะกับนิวคลีโอไซด์ที่ทำหน้าที่เป็นลิแกนด์ รูปทรงที่เป็นไปได้ของโลหะที่เอื้อต่อการก่อตัวแบบดูเพล็กซ์โดยมีนิวคลีโอไซด์ไบเดนเตตสองอันล้อมรอบอะตอมโลหะกลาง ได้แก่เตตระฮีดรัล โดเดคาฮีดรัลและสี่เหลี่ยมระนาบ การเกิดสารประกอบเชิงซ้อนของโลหะกับดีเอ็นเอสามารถเกิดขึ้นได้โดยการสร้างคู่เบสที่ไม่ใช่แบบคาโนนิคัลจากนิวคลีโอเบสธรรมชาติที่มีไอออนโลหะเข้าร่วม และโดยการแลกเปลี่ยนอะตอมไฮโดรเจนซึ่งเป็นส่วนหนึ่งของการจับคู่เบสวัตสัน-คริกโดยไอออนโลหะการนำไอออนโลหะเข้าสู่คู่ดีเอ็นเอแสดงให้เห็นว่ามีศักยภาพทางแม่เหล็กหรือคุณสมบัติการนำไฟฟ้ารวมถึงความเสถียรที่เพิ่มขึ้น
การเกิดสารประกอบเชิงซ้อนของโลหะแสดงให้เห็นว่าเกิดขึ้นระหว่างนิวคลีโอเบส ตามธรรมชาติ ตัวอย่างที่มีการบันทึกไว้เป็นอย่างดีคือการก่อตัวของ T-Hg-T ซึ่งเกี่ยวข้องกับ นิว คลีโอ เบสไทมีนที่ถูกดีโปรตอนสองตัว ที่ถูกเชื่อมต่อกันด้วย Hg และก่อตัวเป็นคู่โลหะ-เบสที่เชื่อมต่อกันโมทีฟนี้ไม่รองรับ Hg ที่ซ้อนกัน ในดูเพล็กซ์ เนื่องจากกระบวนการสร้างกิ๊บติดผมภายในสายซึ่งเป็นที่นิยมมากกว่าการสร้างดูเพล็กซ์ไทมีนสองตัวที่อยู่ตรงข้ามกันจะไม่สร้างคู่เบสวัตสัน-คริกในดูเพล็กซ์ นี่เป็นตัวอย่างที่ความไม่ตรงกันของคู่เบสวัตสัน-คริกถูกทำให้เสถียรโดยการก่อตัวของคู่โลหะ-เบส อีกตัวอย่างหนึ่งของการเกิดสารประกอบเชิงซ้อนของโลหะกับนิวคลีโอเบสตามธรรมชาติคือการก่อตัวของ A-Zn-T และ G-Zn-C ที่ pH สูง Co และ Ni ก็ก่อตัวเป็นสารเชิงซ้อนเหล่านี้เช่นกัน คู่เบสวัตสัน-คริก ซึ่งไอออนบวกที่มีประจุบวกสองประจุจะประสานกับนิวคลีโอเบส การผูกมัดที่แน่นอนยังคงเป็นที่ถกเถียงกันอยู่
มีการพัฒนานิวคลีโอเบสเทียมหลากหลายชนิดเพื่อใช้เป็นคู่เบสโลหะ นิวคลีโอเบสที่ผ่านการดัดแปลงเหล่านี้แสดงคุณสมบัติทางอิเล็กทรอนิกส์ ขนาด และความสัมพันธ์ในการจับที่ปรับได้ ซึ่งสามารถปรับให้เหมาะสมกับโลหะเฉพาะชนิดได้ ตัวอย่างเช่น นิวคลีโอไซด์ที่ผ่านการดัดแปลงด้วยไพริดีน-2,6-ไดคาร์บอกซิเลต พบว่าสามารถจับกับ Cu ได้อย่างแน่นหนา ในขณะที่ไอออนที่มีประจุสองประจุอื่นๆ จะจับได้เพียงหลวมๆ ลักษณะของไตรเดนเตตมีส่วนทำให้เกิดการเลือกปฏิบัตินี้ ตำแหน่งโคออร์ดิเนชันที่สี่บนทองแดงอิ่มตัวด้วยนิวคลีโอเบสไพริดีนที่จัดเรียงตรงข้ามกันระบบการจับคู่เบสโลหะแบบอสมมาตรตั้งฉากกับคู่เบสวัตสัน-คริก อีกตัวอย่างหนึ่งของนิวคลีโอเบสเทียมคือนิวคลีโอเบสไฮดรอกซีไพริโดน ซึ่งสามารถจับกับ Cu ภายในดูเพล็กซ์ของดีเอ็นเอ คู่เบสทองแดง-ไฮดรอกซีไพริโดนที่ต่อเนื่องกันห้าคู่ถูกรวมเข้าเป็นสายคู่ ซึ่งขนาบข้างด้วยนิวคลีโอเบสธรรมชาติเพียงตัวเดียวที่ปลายทั้งสองด้าน ข้อมูล EPR แสดงให้เห็นว่าระยะห่างระหว่างศูนย์กลางของทองแดงนั้นประมาณอยู่ที่ 3.7 ± 0.1 Å ในขณะที่ DNA ดูเพล็กซ์ชนิด B ตามธรรมชาตินั้นมีขนาดใหญ่กว่าเพียงเล็กน้อยเท่านั้น (3.4 Å) ข้อดีของการเรียงซ้อนไอออนของโลหะภายใน DNA ดูเพล็กซ์คือความหวังที่จะได้สายโลหะที่สามารถประกอบตัวเองได้ในระดับนาโน แม้ว่าสิ่งนี้จะยังไม่เกิดขึ้นจริงก็ตาม
คู่เบสที่ไม่เป็นธรรมชาติ (UBP)
คู่เบสที่ไม่เป็นธรรมชาติ (UBP) คือหน่วยย่อย (หรือนิวคลีโอเบส ) ที่ได้รับการออกแบบของDNAที่สร้างขึ้นในห้องปฏิบัติการและไม่เกิดขึ้นในธรรมชาติ ในปี 2012 กลุ่มนักวิทยาศาสตร์ชาวอเมริกันที่นำโดย Floyd Romesberg นักชีววิทยาเคมีที่Scripps Research Instituteใน San Diego, California ได้ตีพิมพ์ว่าทีมของเขาได้ออกแบบคู่เบสที่ไม่เป็นธรรมชาติสองคู่ชื่อว่าd5SICSและdNaM [ ในทางเทคนิคแล้ว นิวคลีโอไทด์เทียมเหล่านี้ซึ่งมีนิวคลีโอเบสที่ไม่ชอบน้ำจะ มี วงแหวนอะโรมาติกสองวงที่หลอมรวมกันซึ่งก่อตัวเป็นคอมเพล็กซ์ d5SICS–dNaM หรือคู่เบสใน DNA ในปี 2014 ทีมเดียวกันรายงานว่าพวกเขาได้สังเคราะห์พลาสมิดที่มีคู่เบส TA และ CG ตามธรรมชาติพร้อมกับ UBP ที่มีประสิทธิภาพดีที่สุด ห้องปฏิบัติการของ Romesberg ได้ออกแบบและใส่เข้าไปในเซลล์ของแบคทีเรียทั่วไปE. coliซึ่งประสบความสำเร็จในการจำลองคู่เบสที่ไม่เป็นธรรมชาติผ่านหลายชั่วอายุคนนี่เป็นตัวอย่างแรกที่ทราบกันดีของสิ่งมีชีวิตที่ถ่ายทอดรหัสพันธุกรรมที่ขยายออกไปยังรุ่นต่อๆ มาความสำเร็จนี้ส่วนหนึ่งเกิดจากการเพิ่มยีนสาหร่ายที่สนับสนุนซึ่งแสดง ตัวขนส่งนิว คลีโอไทด์ไตรฟอสเฟตซึ่งนำเข้าไตรฟอสเฟตของทั้ง d5SICSTP และ dNaMTP เข้าสู่แบคทีเรียอีโคไล ได้อย่างมีประสิทธิภาพ พลาสมิดที่มี d5SICS–dNaM ได้อย่างแม่นยำ
ความสำเร็จในการรวมคู่เบสที่สามเข้าด้วยกันถือเป็นความก้าวหน้าครั้งสำคัญที่นำไปสู่เป้าหมายในการเพิ่มจำนวนกรดอะมิโนที่ดีเอ็นเอสามารถเข้ารหัสได้อย่างมาก จากกรดอะมิโนที่มีอยู่ 20 ชนิด เป็น 172 ชนิดที่เป็นไปได้ในทางทฤษฎี ซึ่งจะช่วยเพิ่มศักยภาพของสิ่งมีชีวิตในการผลิตโปรตีน ชนิดใหม่ ก่อนหน้านี้ สายดีเอ็นเอเทียมไม่ได้เข้ารหัสอะไรเลย แต่นักวิทยาศาสตร์คาดการณ์ว่าสายดีเอ็นเอเทียมอาจถูกออกแบบมาเพื่อผลิตโปรตีนชนิดใหม่ที่อาจนำไปใช้ในอุตสาหกรรมหรือเภสัชกรรมได้[ การถอดรหัสดีเอ็นเอที่มีคู่เบสที่ไม่เป็นธรรมชาติและการแปลรหัส mRNA ที่สอดคล้องกันนั้น ประสบความสำเร็จเมื่อไม่นานมานี้ ในเดือนพฤศจิกายน 2017 ทีมวิจัยเดียวกันจากสถาบันวิจัย Scripps ซึ่งเป็นทีมแรกที่นำนิวคลีโอเบสเพิ่มเติมอีกสองตัวเข้าไปในดีเอ็นเอของแบคทีเรีย รายงานว่าได้สร้างแบคทีเรีย E. coliกึ่งสังเคราะห์ ที่สามารถสร้างโปรตีนโดยใช้ดีเอ็นเอดังกล่าวได้ ดีเอ็นเอของแบคทีเรียประกอบด้วย นิ วคลีโอ เบสที่แตกต่างกันหกชนิด ได้แก่ นิว คลีโอเบสแบบแคนนอนิคัลสี่ชนิด และ dNaM และ dTPT3 ที่เติมเข้าไปอีกสองชนิด (ทั้งสองชนิดนี้ประกอบกันเป็นคู่) แบคทีเรียมีเบส RNA ที่สอดคล้องกันสองตัวที่รวมอยู่ในโคดอนใหม่สองตัว tRNA เพิ่มเติมที่จดจำโคดอนใหม่เหล่านี้ (tRNA เหล่านี้ยังมีเบส RNA ใหม่สองตัวภายในแอนติโคดอนของมันด้วย) และกรดอะมิโนเพิ่มเติม ทำให้แบคทีเรียสามารถสังเคราะห์โปรตีนที่ "ไม่เป็นธรรมชาติ" ได้
การสาธิต UBP อีกครั้งหนึ่งทำได้โดยกลุ่มของ Ichiro Hirao ที่ สถาบัน RIKENในประเทศญี่ปุ่น ในปี 2002 พวกเขาพัฒนาคู่เบสที่ไม่เป็นธรรมชาติระหว่าง 2-amino-8-(2-thienyl)purine (s) และ pyridine-2-one (y) ซึ่งทำงานในหลอดทดลองในการถอดรหัสและการแปลรหัส เพื่อการรวมกรดอะมิโนที่ไม่ได้มาตรฐานเข้ากับโปรตีนที่ตำแหน่งเฉพาะในปี 2006 พวกเขาได้สร้าง 7-(2-thienyl)imidazo[4,5-b]pyridine (Ds) และ pyrrole-2-carbaldehyde (Pa) เป็นคู่เบสที่สามสำหรับการจำลองแบบและการถอดรหัสหลังจากนั้น Ds และ 4-[3-(6-aminohexanamido)-1-propynyl]-2-nitropyrrole (Px) ถูกค้นพบว่าเป็นคู่เบสที่มีความเที่ยงตรงสูงในการขยาย PCR ในปี 2013 พวกเขาได้นำคู่ Ds-Px มาใช้กับการสร้าง DNA แอปทาเมอร์โดย การคัดเลือก ในหลอดทดลอง (SELEX) และแสดงให้เห็นว่าการขยายตัวอักษรทางพันธุกรรมช่วยเพิ่มความสัมพันธ์ของ DNA แอปทาเมอร์กับโปรตีนเป้าหมายได้อย่างมีนัยสำคัญ
แอปพลิเคชัน
ยา
อะนาล็อกของนิวคลีโอไซด์หลายชนิดถูกใช้เป็นสารต้านไวรัสหรือสารต้านมะเร็ง โพลิ เมอเรส ของไวรัสจะรวมสารประกอบเหล่านี้เข้ากับเบสที่ไม่ใช่เบสหลัก สารประกอบเหล่านี้จะถูกกระตุ้นในเซลล์โดยการแปลงเป็นนิวคลีโอไทด์ และจะถูกให้ในรูปนิวคลีโอไซด์ เนื่องจากนิวคลีโอไทด์ที่มีประจุไม่สามารถผ่านเยื่อหุ้มเซลล์ได้ง่าย
ชีววิทยาโมเลกุล
{kind=link}

อนาล็อกของกรดนิวคลีอิกถูกนำมาใช้ในทางชีววิทยาโมเลกุลเพื่อวัตถุประสงค์หลายประการ:
- การตรวจสอบสถานการณ์ที่เป็นไปได้ของต้นกำเนิดของสิ่งมีชีวิต: โดยการทดสอบอะนาล็อกที่แตกต่างกัน นักวิจัยพยายามตอบคำถามว่าการใช้ DNA และ RNA ของสิ่งมีชีวิตนั้นถูกเลือกโดยอาศัยข้อดีของมันเมื่อเวลาผ่านไปหรือถูกเลือกโดยโอกาสโดยพลการ
- ในฐานะเครื่องมือสำหรับตรวจจับลำดับเฉพาะ: XNA สามารถใช้เพื่อแท็กและระบุส่วนประกอบ DNA และ RNA ที่หลากหลายด้วยความจำเพาะและความแม่นยำสูง
- เนื่องจากเป็นเอนไซม์ที่ออกฤทธิ์กับสารตั้งต้นของ DNA, RNA และ XNA - XNA ได้รับการพิสูจน์แล้วว่าสามารถตัดและเชื่อม DNA, RNA และโมเลกุล XNA อื่นๆ ได้คล้ายกับการทำงานของไรโบไซม์ ของ RNA [
- เป็นเครื่องมือที่มีความต้านทานต่อการ ไฮโดรไลซิ สRNA
- การศึกษากลไกการใช้เอนไซม์ และ
- การศึกษาลักษณะโครงสร้างของกรดนิวคลีอิก
สารตั้งต้นของโลกอาร์เอ็นเอ
มีการเสนอแนะว่าโลกของ RNAอาจมีมาก่อน "โลกที่คล้าย RNA" ซึ่งมีกรดนิวคลีอิกอื่นๆ ที่มีกระดูกสันหลังที่แตกต่างกัน เช่นGNA , PNAและTNAมีอยู่ อย่างไรก็ตาม หลักฐานสำหรับสมมติฐานนี้ถูกเรียกว่า "ไม่แน่นอน"
ระบบตั้งฉาก
มีการเสนอและศึกษาความเป็นไปได้ทั้งในเชิงทฤษฎีและเชิงทดลองในการนำระบบตั้งฉากมาใช้ภายในเซลล์ที่ไม่ขึ้นอยู่กับสารพันธุกรรมของเซลล์ เพื่อสร้างระบบที่ปลอดภัยอย่างสมบูรณ์โดยอาจเพิ่มศักยภาพในการเข้ารหัสได้ กลุ่มต่างๆ หลายกลุ่มได้มุ่งเน้นไปที่ด้านต่างๆ:
- โครงกระดูกและคู่เบสใหม่ตามที่ได้กล่าวข้างต้น
- การจำลองเทียม ของ XNAและการถอดรหัสโพลีเมอเรสโดยทั่วไปเริ่มต้นจากโพลีเมอเรส RNA T7 [
- ( ลำดับไรโบโซม 16Sที่มีลำดับ anti- Shine-Dalgarno ที่เปลี่ยนแปลงไป ทำให้สามารถแปล mRNA ออร์โธโกนัลได้เฉพาะกับลำดับ Shine-Dalgarno ที่เปลี่ยนแปลงไปเท่านั้น; และ
- tRNA ใหม่ที่เข้ารหัสกรดอะมิ โนที่ไม่ใช่ธรรมชาติสำหรับรหัสพันธุกรรมที่ขยายออก
ในเดือนพฤษภาคม 2557 นักวิจัยประกาศว่าพวกเขาประสบความสำเร็จในการนำนิวคลีโอไทด์ เทียมใหม่สองชนิด เข้าสู่ดีเอ็นเอของแบคทีเรีย และด้วยการนำนิวคลีโอไทด์เทียมแต่ละชนิดมาใส่ในอาหารเลี้ยงเชื้อ ทำให้สามารถผ่านแบคทีเรียได้ 24 ครั้ง พวกเขาไม่ได้สร้าง mRNA หรือโปรตีนที่สามารถใช้นิวคลีโอไทด์เทียมได้ นิวคลีโอไทด์เทียมเหล่านี้มีวงแหวนอะโรมาติก 2 วงที่เชื่อมติดกัน
ดูเพิ่มเติม
- ไบโอติน
- ยาดับกลิ่นมืด
- ดีออกซีไรโบไซม์
- รหัสพันธุกรรมที่ขยายออกไป
- ฟลูออโรโฟร์
- พันธุศาสตร์
- ชีววิทยาโมเลกุล
- กรดนิวคลีอิก
- นิวคลีโอเบส
- นิวคลีโอไซด์
- นิวคลีโอไทด์
- การสังเคราะห์โอลิโกนิวคลีโอไทด์
- ไรโบไซม์
- ชีววิทยาสังเคราะห์
- ชีววิทยาต่างดาว
- เอ็กซ์ดีเอ็นเอ
- ดีเอ็นเอของฮาจิโมจิ
- ระบบสารสนเทศทางพันธุกรรมที่ขยายโดยเทียม (AEGIS)
- กรดนิวคลีอิกซีโน
