อ่าน 17 นาที
ศักย์เยื่อหุ้มเซลล์
ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ (หรือ ศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์ หรือ แรงดันไฟฟ้าของเยื่อหุ้มเซลล์ ) คือความแตกต่างของ ศักย์ไฟฟ้า ภายในและภายนอกของ เซลล์ ชีวภาพ
ศักย์เยื่อหุ้มเซลล์

ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ (หรือศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์หรือแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ ) คือความแตกต่างของศักย์ไฟฟ้าภายในและภายนอกของเซลล์ ชีวภาพ
ตามธรรมเนียมจะเขียนว่า V m =V ภายใน −V ภายนอกดังนั้นศักย์เยื่อหุ้มเซลล์ที่เป็นลบหมายความว่าภายในเซลล์เป็นลบเมื่อเทียบกับภายนอก[ 1 ]โดยพื้นฐานแล้ว มันเท่ากับศักย์ภายในลบด้วยศักย์ภายนอก นี่คือพลังงานต่อประจุที่จำเป็นในการเคลื่อนย้ายประจุบวกขนาดเล็กมากด้วยความเร็วคงที่ผ่านเยื่อหุ้มเซลล์จากภายนอกสู่ภายใน อย่างไรก็ตาม โปรดทราบว่าหากอนุญาตให้ประจุเปลี่ยนความเร็ว จะต้องคำนึงถึงการเปลี่ยนแปลงของพลังงานจลน์และการผลิตรังสีด้วย[ 1 ]
ค่าทั่วไปของศักย์เยื่อหุ้มเซลล์มักจะระบุเป็นหน่วยมิลลิโวลต์และใช้สัญลักษณ์ mV ในเซลล์สัตว์หลายชนิด Vm โดยทั่วไปจะมีค่าอยู่ในช่วงหลายสิบมิลลิโวลต์ โดยทั่วไปจะอยู่ในช่วงประมาณ −20 mV ถึง −200 mV ขึ้นอยู่กับชนิดและสภาวะของเซลล์[ 1 ]สำหรับศักย์เยื่อหุ้มเซลล์ที่เป็นลบโดยทั่วไปเช่นนี้ ต้องใช้พลังงานบวกในการเคลื่อนย้ายประจุบวกจากภายในสู่ภายนอก อย่างไรก็ตามพลังงานจลน์ความร้อนช่วยให้ไอออนสามารถเอาชนะความแตกต่างของศักย์ได้ สำหรับเยื่อหุ้มเซลล์ที่เลือกผ่านได้สิ่งนี้จะทำให้เกิดการไหลสุทธิสวนทางกับความชัน
แม้ว่าภายในเซลล์ทั้งหมดจะถือว่ามีประจุลบ แต่ของเหลวภายในเซลล์นั้นเป็นกลางทางไฟฟ้า การมีอยู่ของแรงดันไฟฟ้าเกิดจากชั้นประจุบางๆ ที่สะสมอยู่ที่พื้นผิวของเยื่อหุ้มเซลล์ เยื่อหุ้มเซลล์มีความหนาเพียงไม่กี่นาโนเมตร ดังนั้นแม้แรงดันไฟฟ้า 100 mV ที่เกิดขึ้นทั่วความหนานี้จะสร้างสนามไฟฟ้าที่แรงมากบนเยื่อหุ้มเซลล์[ 1 ]
คำอธิบาย
เซลล์สัตว์ทุกเซลล์ถูกล้อมรอบด้วยเยื่อหุ้มเซลล์ซึ่งประกอบด้วยชั้นไขมันสองชั้นที่มีโปรตีนฝังอยู่ภายใน เยื่อหุ้มเซลล์ทำหน้าที่ทั้งเป็นตัวเก็บประจุและเป็นตัวกั้นการแพร่ของไอออนปั๊มไอออนจะผลักไอออนผ่านเยื่อหุ้มเซลล์อย่างแข็งขันและสร้างความแตกต่างของความเข้มข้นข้ามเยื่อหุ้มเซลล์ ในขณะที่ช่องไอออนจะยอมให้ไอออนเคลื่อนที่ผ่านเยื่อหุ้มเซลล์ตามความแตกต่างของศักย์ไฟฟ้า ช่องไอออนและปั๊มไอออนเป็นโปรตีนที่อยู่ภายในเยื่อหุ้มเซลล์และมีลักษณะทางไฟฟ้าคล้ายกับตัวต้านทานและแหล่งกำเนิดกระแสไฟฟ้า (แบตเตอรี่) และมีส่วนช่วยสร้างแรงดันไฟฟ้าข้ามสองด้านของเยื่อหุ้มเซลล์
เยื่อหุ้มพลาสมาทั้งหมดมีศักย์ไฟฟ้าคร่อมอยู่ โดยด้านในมักจะมีประจุลบเมื่อเทียบกับด้านนอก[ 2 ]ศักย์ไฟฟ้าของเยื่อหุ้มมีหน้าที่พื้นฐานสองประการ ประการแรก ช่วยให้เซลล์ทำงานเหมือนแบตเตอรี่ โดยให้พลังงานเพื่อใช้งาน "อุปกรณ์ระดับโมเลกุล" ต่างๆ ที่ฝังอยู่ในเยื่อหุ้ม[ 3 ]ประการที่สอง ในเซลล์ที่สามารถกระตุ้น ด้วยไฟฟ้าได้ เช่นเซลล์ประสาทและเซลล์กล้ามเนื้อ ศักย์ ไฟฟ้า นี้ใช้สำหรับการส่งสัญญาณระหว่างส่วนต่างๆ ของเซลล์
สัญญาณในเซลล์ประสาทและเซลล์กล้ามเนื้อ
ในเซลล์ที่ไวต่อการกระตุ้น สัญญาณจะเกิดขึ้นจากการเปิดหรือปิดของช่องไอออน ณ จุดหนึ่งในเยื่อหุ้มเซลล์ ทำให้เกิดการเปลี่ยนแปลงศักย์ไฟฟ้าเฉพาะที่ในเยื่อหุ้มเซลล์ การเปลี่ยนแปลงของสนามไฟฟ้าดังกล่าวสามารถรับรู้ได้อย่างรวดเร็วโดยช่องไอออนที่อยู่ใกล้เคียงหรืออยู่ห่างออกไปในเยื่อหุ้มเซลล์ จากนั้นช่องไอออนเหล่านั้นจะเปิดหรือปิดตามการเปลี่ยนแปลงของศักย์ไฟฟ้า ทำให้เกิดสัญญาณขึ้นใหม่
ในเซลล์ที่ไม่สามารถถูกกระตุ้นได้ และในเซลล์ที่สามารถถูกกระตุ้นได้ในสภาวะพื้นฐาน ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จะคงอยู่ที่ค่าที่ค่อนข้างคงที่ เรียกว่าศักย์พัก (resting potential ) สำหรับเซลล์ประสาท ศักย์พักจะถูกกำหนดให้มีค่าตั้งแต่ -80 ถึง -70 มิลลิโวลต์ นั่นคือ ภายในเซลล์มีแรงดันไฟฟ้าพื้นฐานเป็นลบเล็กน้อย น้อยกว่าหนึ่งในสิบของโวลต์ การเปิดและปิดของช่องไอออนสามารถทำให้เกิดการเปลี่ยนแปลงจากศักย์พักได้ หากแรงดันไฟฟ้าภายในเซลล์ลดลงเป็นลบน้อยลง (เช่น จาก -70 มิลลิโวลต์ เป็น -60 มิลลิโวลต์) เรียกว่าการลดศักย์ ( depolarization ) หรือหากแรงดันไฟฟ้าภายในเซลล์เพิ่มขึ้นเป็นลบมากขึ้น (เช่น จาก -70 มิลลิโวลต์ เป็น -80 มิลลิโวลต์) เรียกว่า การเพิ่มศักย์ (hyperpolarization) ในเซลล์ที่สามารถถูกกระตุ้นได้ การลดศักย์ที่มากพอสามารถกระตุ้นให้เกิดศักย์ การกระทำ ( action potential ) ซึ่งศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จะเปลี่ยนแปลงอย่างรวดเร็วและมีนัยสำคัญในช่วงเวลาสั้นๆ (ประมาณ 1 ถึง 100 มิลลิวินาที) และมักจะเปลี่ยนขั้ว ศักย์ไฟฟ้าแอ็กชันเกิดขึ้นจากการกระตุ้นของช่องไอออนที่ควบคุมด้วยแรงดันไฟฟ้าบาง ชนิด
ในเซลล์ประสาท ปัจจัยที่มีอิทธิพลต่อศักย์เยื่อหุ้มเซลล์นั้นมีหลากหลาย รวมถึงช่องไอออนหลายประเภท บางชนิดถูกควบคุมด้วยสารเคมี และบางชนิดถูกควบคุมด้วยแรงดันไฟฟ้า เนื่องจากช่องไอออนที่ควบคุมด้วยแรงดันไฟฟ้าถูกควบคุมโดยศักย์เยื่อหุ้มเซลล์ ในขณะที่ศักย์เยื่อหุ้มเซลล์เองก็ได้รับอิทธิพลจากช่องไอออนเหล่านี้เช่นกัน จึงเกิดวงจรป้อนกลับที่ทำให้เกิดพลวัตเชิงเวลาที่ซับซ้อน รวมถึงการสั่นและการเกิดใหม่ เช่น ศักย์การกระทำ (action potential)
การไล่ระดับความเข้มข้นของไอออน
ความแตกต่างของความเข้มข้นของไอออนที่ด้านตรงข้ามของเยื่อหุ้มเซลล์ทำให้เกิดแรงดันไฟฟ้าที่เรียกว่าศักย์เยื่อหุ้มเซลล์[ 4 ]
ไอออนหลายชนิดมีระดับความเข้มข้นแตกต่างกันข้ามเยื่อหุ้มเซลล์ รวมถึง โพแทสเซียม (K + ) ซึ่งมีความเข้มข้นสูงภายในเซลล์และความเข้มข้นต่ำภายนอกเซลล์ ส่วน โซเดียม (Na + ) และคลอไรด์ (Cl− )มีความเข้มข้นสูงใน บริเวณ นอกเซลล์และความเข้มข้นต่ำใน บริเวณ ภายในเซลล์ ความแตกต่างของความเข้มข้นเหล่านี้ก่อให้เกิดพลังงานศักย์เพื่อขับเคลื่อนการเกิดศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ แรงดันไฟฟ้านี้เกิดขึ้นเมื่อเยื่อหุ้มเซลล์สามารถซึมผ่านไอออนหนึ่งชนิดหรือมากกว่าได้
ในกรณีที่ง่ายที่สุด ดังแสดงในแผนภาพด้านบน ("ความแตกต่างของความเข้มข้นของไอออน") หากเยื่อหุ้มเซลล์ยอมให้โพแทสเซียมผ่านได้แบบเลือกสรร ไอออนที่มีประจุบวกเหล่านี้จะสามารถแพร่ไปตามความแตกต่างของความเข้มข้นไปยังภายนอกเซลล์ ทำให้เหลือประจุลบที่ไม่ได้รับการชดเชยไว้ การแยกตัวของประจุนี้เองที่เป็นสาเหตุให้เกิดศักย์เยื่อหุ้มเซลล์
โดยรวมแล้ว ระบบนี้มีสภาพเป็นกลางทางไฟฟ้า ประจุบวกที่ไม่ได้รับการชดเชยภายนอกเซลล์ และประจุลบที่ไม่ได้รับการชดเชยภายในเซลล์ จะเรียงตัวกันบนพื้นผิวของเยื่อหุ้มเซลล์และดึงดูดซึ่งกันและกันผ่านชั้นไขมันสองชั้นดังนั้น ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จึงอยู่เฉพาะบริเวณใกล้เคียงกับเยื่อหุ้มเซลล์เท่านั้น การแยกตัวของประจุเหล่านี้ข้ามเยื่อหุ้มเซลล์เป็นพื้นฐานของแรงดันไฟฟ้าของเยื่อหุ้มเซลล์
แผนภาพด้านบนเป็นเพียงการประมาณค่าการมีส่วนร่วมของไอออนต่อศักยภาพของเยื่อหุ้มเซลล์ ไอออนอื่นๆ รวมถึงโซเดียม คลอไรด์ แคลเซียม และอื่นๆ มีบทบาทน้อยกว่า แม้ว่าจะมีระดับความเข้มข้นสูงก็ตาม เนื่องจากมีการซึมผ่านที่จำกัดกว่าโพแทสเซียม[ 5 ]
เหตุใดเซลล์จึงมีศักย์เยื่อหุ้มเซลล์?
แหล่งที่มา: [ 6 ]
- เยื่อหุ้มเซลล์เป็นฉนวนไฟฟ้าสำหรับไอออน:เยื่อหุ้มพลาสมาเป็นชั้นไขมันสองชั้นที่มีความต้านทานต่อการผ่านของไอออนสูงมาก ดังนั้นไอออนจึงไม่สามารถแพร่ผ่านเยื่อหุ้มเซลล์ได้อย่างอิสระเหมือนกับที่แพร่ผ่านน้ำ เซลล์จึงแก้ปัญหานี้โดยใช้โปรตีนในเยื่อหุ้มเซลล์ เช่น ช่องทาง ตัวขนส่ง และปั๊ม เพื่อควบคุมการผ่านของไอออน
- เซลล์สร้างความแตกต่างของความเข้มข้นของไอออน (พลังงานสะสม):เซลล์ใช้พลังงานเพื่อรักษาระดับความเข้มข้นของไอออนภายในและภายนอกเซลล์ให้แตกต่างกัน ตัวอย่างที่รู้จักกันดีที่สุดคือปั๊มโซเดียม-โพแทสเซียม ซึ่งโดยปกติจะขนส่งโซเดียม 3 ไอออนออกและโพแทสเซียม 2 ไอออนเข้าต่อโมเลกุล ATP 1 โมเลกุล ทำให้เกิดความเข้มข้นของโซเดียมสูงภายนอกและความเข้มข้นของโพแทสเซียมสูงภายใน ซึ่งเป็นการเก็บสะสมพลังงานเคมีในความแตกต่างของความเข้มข้นนี้
- การซึมผ่านแบบเลือกสรรเปลี่ยนความแตกต่างของความเข้มข้นให้เป็นศักย์ไฟฟ้า:ในสภาวะพัก เซลล์ส่วนใหญ่จะซึมผ่านโพแทสเซียมได้มากกว่าโซเดียมมาก เนื่องจากมีช่องรั่วไหลของโพแทสเซียม ไอออนของโพแทสเซียมจะแพร่กระจายไปตามความเข้มข้นของมันออกจากเซลล์ ทำให้เกิดไอออนที่มีประจุลบอยู่ภายในเซลล์ ซึ่งก่อให้เกิดประจุลบของศักย์เยื่อหุ้มเซลล์
เหตุใดศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จึงมีความสำคัญ?
แหล่งที่มา: [ 1 ]
- การส่งสัญญาณไฟฟ้า (เซลล์ที่ไวต่อการกระตุ้น) : เซลล์ประสาทและเซลล์กล้ามเนื้อใช้ศักย์ไฟฟ้าในการส่งผ่านข้อมูล
- การขับเคลื่อนการขนส่งและการเผาผลาญ:แรงดันไฟฟ้าของเยื่อหุ้มเซลล์เก็บพลังงานที่เซลล์สามารถนำไปใช้ในกระบวนการขนส่งได้
- การผลิตพลังงานในไมโทคอนเดรีย:ศักย์ไฟฟ้ามีความสำคัญต่อการไล่ระดับโปรตอนที่ขับเคลื่อนการสังเคราะห์ ATP ทั่วเยื่อหุ้มไมโทคอนเดรียชั้นใน
- พฤติกรรมของเซลล์นอกเหนือจากเนื้อเยื่อ "ไฟฟ้า":การเปลี่ยนแปลงของศักย์เยื่อหุ้มเซลล์สามารถส่งผลต่อคุณสมบัติของเซลล์ เช่น กลไกและโครงสร้าง (ตัวอย่างเช่น เซลล์เยื่อบุผนังหลอดเลือดจะแข็งตัวขึ้นเมื่อเกิดการลดศักย์)
พื้นฐานทางกายภาพ
ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เกิดจากปัจจัยสองประการหลัก ได้แก่ แรงไฟฟ้าและการแพร่ แรงไฟฟ้าเกิดจากแรงดึงดูดระหว่างอนุภาคที่มีประจุไฟฟ้าตรงข้ามกัน (บวกและลบ) และแรงผลักระหว่างอนุภาคที่มีประจุชนิดเดียวกัน (บวกทั้งคู่หรือลบทั้งคู่) ส่วนการแพร่เกิดจากแนวโน้มทางสถิติของอนุภาคที่จะกระจายตัวจากบริเวณที่มีความเข้มข้นสูงไปยังบริเวณที่มีความเข้มข้นต่ำ
แรงดันไฟฟ้า

แรงดันไฟฟ้า ซึ่งมีความหมายเหมือนกับความแตกต่างของศักย์ไฟฟ้าคือความสามารถในการขับเคลื่อนกระแสไฟฟ้าผ่านความต้านทาน ที่จริงแล้ว นิยามที่ง่ายที่สุดของแรงดันไฟฟ้าคือกฎของโอห์ม : V=IR โดยที่ V คือแรงดันไฟฟ้า I คือกระแสไฟฟ้า และ R คือความต้านทาน หากแหล่งกำเนิดแรงดันไฟฟ้า เช่น แบตเตอรี่ ถูกวางไว้ในวงจรไฟฟ้า แรงดันไฟฟ้าของแหล่งกำเนิดยิ่งสูง กระแสไฟฟ้าที่ไหลผ่านความต้านทานก็จะยิ่งมากขึ้น ความสำคัญเชิงฟังก์ชันของแรงดันไฟฟ้าอยู่ที่ความแตกต่าง ของศักย์ไฟฟ้า ระหว่างสองจุดในวงจรเท่านั้น แนวคิดของแรงดันไฟฟ้าที่จุดเดียวไม่มีความหมาย ในทางอิเล็กทรอนิกส์ เป็นเรื่องปกติที่จะกำหนดแรงดันไฟฟ้าเป็นศูนย์ให้กับองค์ประกอบใดองค์ประกอบหนึ่งในวงจรที่เลือกไว้โดยพลการ จากนั้นจึงกำหนดแรงดันไฟฟ้าสำหรับองค์ประกอบอื่นๆ โดยวัดเทียบกับจุดศูนย์นั้น การเลือกองค์ประกอบใดเป็นจุดศูนย์นั้นไม่มีความสำคัญ การทำงานของวงจรขึ้นอยู่กับความแตกต่างของศักย์ไฟฟ้าเท่านั้น ไม่ใช่แรงดันไฟฟ้าโดยตรงอย่างไรก็ตาม ในกรณีส่วนใหญ่และตามธรรมเนียมปฏิบัติ ระดับศูนย์มักจะถูกกำหนดให้กับส่วนของวงจรที่สัมผัสกับกราวด์
หลักการเดียวกันนี้ใช้ได้กับแรงดันไฟฟ้าในชีววิทยาของเซลล์ ในเนื้อเยื่อที่มีการทำงานทางไฟฟ้า ความต่างศักย์ระหว่างสองจุดใดๆ สามารถวัดได้โดยการเสียบอิเล็กโทรดที่แต่ละจุด เช่น จุดหนึ่งอยู่ภายในเซลล์และอีกจุดหนึ่งอยู่ภายนอกเซลล์ แล้วเชื่อมต่ออิเล็กโทรดทั้งสองเข้ากับสายของเครื่องวัดแรงดันไฟฟ้าชนิดพิเศษ ตามธรรมเนียมแล้ว ค่าศักย์ศูนย์จะถูกกำหนดให้เป็นด้านนอกของเซลล์ และเครื่องหมายของความต่างศักย์ระหว่างด้านนอกและด้านในจะถูกกำหนดโดยศักย์ของด้านในเมื่อเทียบกับค่าศูนย์ภายนอก
ในทางคณิตศาสตร์ นิยามของแรงดันไฟฟ้าเริ่มต้นด้วยแนวคิดของสนามไฟฟ้าEซึ่งเป็นสนามเวกเตอร์ที่กำหนดขนาดและทิศทางให้กับแต่ละจุดในอวกาศ ในหลายสถานการณ์ สนามไฟฟ้าเป็นสนามอนุรักษ์ซึ่งหมายความว่าสามารถแสดงได้ในรูปของเกรเดียนต์ของฟังก์ชันสเกลาร์Vนั่นคือE = −∇ Vสนามสเกลาร์V นี้ เรียกว่าการกระจายแรงดันไฟฟ้า นิยามนี้อนุญาตให้มีค่าคงที่ของการอินทิเกรตตามอำเภอใจ ซึ่งเป็นเหตุผลว่าทำไมค่าสัมบูรณ์ของแรงดันไฟฟ้าจึงไม่มีความหมาย โดยทั่วไป สนามไฟฟ้าจะถือว่าเป็นสนามอนุรักษ์ได้ก็ต่อเมื่อสนามแม่เหล็กไม่มีอิทธิพลต่อสนามไฟฟ้าอย่างมีนัยสำคัญ แต่เงื่อนไขนี้มักใช้ได้ดีกับเนื้อเยื่อทางชีวภาพ
เนื่องจากสนามไฟฟ้าคือความชันของการกระจายแรงดันไฟฟ้า การเปลี่ยนแปลงอย่างรวดเร็วของแรงดันไฟฟ้าภายในบริเวณเล็กๆ บ่งชี้ถึงสนามไฟฟ้าที่แรง ในทางกลับกัน หากแรงดันไฟฟ้าคงที่โดยประมาณในบริเวณกว้าง สนามไฟฟ้าในบริเวณนั้นจะต้องอ่อน สนามไฟฟ้าที่แรง ซึ่งเทียบเท่ากับความชันของแรงดันไฟฟ้าที่แรง หมายความว่ามีแรงมหาศาลกระทำต่ออนุภาคที่มีประจุใดๆ ที่อยู่ในบริเวณนั้น
ไอออนและแรงที่ขับเคลื่อนการเคลื่อนที่ของพวกมัน

โดยทั่วไปแล้ว สัญญาณไฟฟ้าภายในสิ่งมีชีวิตจะถูกขับเคลื่อนด้วยไอออน [ 8 ] ไอออนบวกที่สำคัญที่สุดสำหรับศักย์ไฟฟ้าแอคชั่นคือโซเดียม ( Na + ) และโพแทสเซียม (K + ) [ 9 ]ทั้งสองเป็น ไอออน บวกโมโนวาเลนต์ที่มีประจุบวกหนึ่งตัว ศักย์ไฟฟ้าแอคชั่นยังสามารถเกี่ยวข้องกับแคลเซียม (Ca 2+ ) [ 10 ]ซึ่งเป็นไอออนบวกไดวาเลนต์ที่มีประจุบวกสองตัวไอออนคลอไรด์ (Cl − ) มีบทบาทสำคัญในศักย์ไฟฟ้าแอคชั่นของสาหร่ายบางชนิด[ 11 ] แต่มีบทบาทน้อยมากในศักย์ไฟฟ้าแอคชั่นของสัตว์ส่วนใหญ่[ 12 ]
ไอออนเคลื่อนที่ผ่านเยื่อหุ้มเซลล์ภายใต้อิทธิพลสองประการ ได้แก่การแพร่และสนามไฟฟ้าตัวอย่างง่ายๆ ที่สารละลายสองชนิด—A และ B—ถูกแยกออกจากกันด้วยสิ่งกีดขวางที่มีรูพรุน จะแสดงให้เห็นว่าการแพร่จะทำให้สารละลายทั้งสองผสมกันในที่สุดจนมีความเข้มข้นเท่ากัน การผสมนี้เกิดขึ้นเนื่องจากความแตกต่างของความเข้มข้น บริเวณที่มีความเข้มข้นสูงจะแพร่ไปยังบริเวณที่มีความเข้มข้นต่ำ เพื่อขยายตัวอย่าง สมมติว่าสารละลาย A มีไอออนโซเดียม 30 ตัวและไอออนคลอไรด์ 30 ตัว และสารละลาย B มีไอออนโซเดียม 20 ตัวและไอออนคลอไรด์ 20 ตัว สมมติว่าสิ่งกีดขวางยอมให้ไอออนทั้งสองชนิดผ่านได้ สภาวะสมดุลจะเกิดขึ้นโดยที่สารละลายทั้งสองมีไอออนโซเดียม 25 ตัวและไอออนคลอไรด์ 25 ตัว อย่างไรก็ตาม หากสิ่งกีดขวางที่มีรูพรุนนั้นเลือกเฉพาะไอออนที่จะผ่านได้ การแพร่เพียงอย่างเดียวจะไม่สามารถกำหนดสารละลายที่ได้ กลับไปที่ตัวอย่างก่อนหน้านี้ ตอนนี้เรามาสร้างสิ่งกีดขวางที่ยอมให้เฉพาะไอออนโซเดียมผ่านได้ ในตอนนี้ โซเดียมเท่านั้นที่สามารถแพร่ผ่านสิ่งกีดขวางจากสารละลาย A ที่มีความเข้มข้นสูงกว่าไปยังสารละลาย B ที่มีความเข้มข้นต่ำกว่าได้ ซึ่งจะส่งผลให้มีการสะสมของไอออนโซเดียมมากกว่าไอออนคลอไรด์ในสารละลาย B และมีการสะสมของไอออนโซเดียมน้อยกว่าไอออนคลอไรด์ในสารละลาย A
นี่หมายความว่าสารละลาย B มีประจุบวกสุทธิเนื่องจากความเข้มข้นของไอออนโซเดียมที่มีประจุบวกสูงกว่าไอออนคลอไรด์ที่มีประจุลบ ในทำนองเดียวกัน สารละลาย A มีประจุลบสุทธิเนื่องจากความเข้มข้นของไอออนคลอไรด์ที่มีประจุลบสูงกว่าไอออนโซเดียมที่มีประจุบวก เนื่องจากประจุตรงข้ามดึงดูดกันและประจุเหมือนกันผลักกัน ไอออนจึงได้รับอิทธิพลจากสนามไฟฟ้าเช่นเดียวกับแรงแพร่กระจาย ดังนั้นไอออนโซเดียมที่มีประจุบวกจึงมีโอกาสน้อยที่จะเคลื่อนที่ไปยังสารละลาย B ที่มีประจุบวกมากกว่า และจะคงอยู่ในสารละลาย A ที่มีประจุลบมากกว่า จุดที่แรงของสนามไฟฟ้าหักล้างกับแรงแพร่กระจายอย่างสมบูรณ์เรียกว่าศักย์สมดุล ณ จุดนี้ การไหลสุทธิของไอออนเฉพาะ (ในกรณีนี้คือโซเดียม) จะเป็นศูนย์
เยื่อหุ้มพลาสมา

เซลล์ทุกเซลล์ถูกห่อหุ้มด้วยเยื่อหุ้ม เซลล์ ซึ่งมีโครงสร้างเป็นชั้นไขมันสองชั้นที่มีโมเลกุลขนาดใหญ่หลายชนิดฝังอยู่ภายใน เนื่องจากทำจากโมเลกุลไขมัน เยื่อหุ้มเซลล์จึงมีความต้านทานไฟฟ้าสูงโดยธรรมชาติ กล่าวอีกนัยหนึ่งคือมีการซึมผ่านของไอออนต่ำโดยธรรมชาติ อย่างไรก็ตาม โมเลกุลบางชนิดที่ฝังอยู่ในเยื่อหุ้มเซลล์สามารถขนส่งไอออนจากด้านหนึ่งของเยื่อหุ้มเซลล์ไปยังอีกด้านหนึ่งได้ หรือสร้างช่องทางให้ไอออนเคลื่อนที่ผ่านได้[ 13 ]
ในทางไฟฟ้า เยื่อหุ้มเซลล์ทำหน้าที่เสมือนตัวต้านทานและตัวเก็บประจุ รวมกัน ความต้านทานเกิดจากข้อเท็จจริงที่ว่าเยื่อหุ้มเซลล์ขัดขวางการเคลื่อนที่ของประจุ ส่วนความจุเกิดจากข้อเท็จจริงที่ว่าชั้นไขมันสองชั้นนั้นบางมาก การสะสมของอนุภาคที่มีประจุอยู่ด้านหนึ่งจึงก่อให้เกิดแรงทางไฟฟ้าที่ดึงอนุภาคที่มีประจุตรงข้ามไปยังอีกด้านหนึ่ง ความจุของเยื่อหุ้มเซลล์นั้นแทบจะไม่ได้รับผลกระทบจากโมเลกุลที่ฝังอยู่ภายใน ดังนั้นจึงมีค่าคงที่โดยประมาณที่ 2 μF/cm² (ความจุรวมของเยื่อหุ้มเซลล์ส่วนหนึ่งเป็นสัดส่วนกับพื้นที่) ในทางกลับกัน การนำไฟฟ้าของชั้นไขมันสองชั้นบริสุทธิ์นั้นต่ำมาก จนในสถานการณ์ทางชีววิทยา การนำไฟฟ้าของชั้นไขมันจึงมักถูกครอบงำด้วยการนำไฟฟ้าของเส้นทางอื่นที่เกิดจากโมเลกุลที่ฝังอยู่ภายใน ดังนั้น ความจุของเยื่อหุ้มเซลล์จึงค่อนข้างคงที่ แต่ความต้านทานนั้นแปรผันได้สูง
ความหนาของเยื่อหุ้มเซลล์คาดว่าอยู่ที่ประมาณ 7-8 นาโนเมตร เนื่องจากเยื่อหุ้มเซลล์บางมาก จึงไม่จำเป็นต้องใช้แรงดันไฟฟ้าข้ามเยื่อหุ้มเซลล์สูงมากนักเพื่อสร้างสนามไฟฟ้าที่แรงภายในเยื่อหุ้มเซลล์ แรงดันไฟฟ้าของเยื่อหุ้มเซลล์โดยทั่วไปในเซลล์สัตว์อยู่ที่ประมาณ 100 มิลลิโวลต์ (นั่นคือหนึ่งในสิบของโวลต์) แต่การคำนวณแสดงให้เห็นว่าแรงดันไฟฟ้านี้สร้างสนามไฟฟ้าใกล้เคียงกับค่าสูงสุดที่เยื่อหุ้มเซลล์สามารถทนได้—มีการคำนวณว่าความแตกต่างของแรงดันไฟฟ้าที่มากกว่า 200 มิลลิโวลต์มากอาจทำให้เกิดการแตกตัวของฉนวนไฟฟ้านั่นคือการเกิดประกายไฟข้ามเยื่อหุ้มเซลล์
การแพร่และการขนส่งแบบอำนวยความสะดวก

เยื่อไขมันบริสุทธิ์มีความต้านทานสูงมากต่อการผ่านของไอออน แต่โครงสร้างที่ฝังอยู่ในเยื่อหุ้มเซลล์สามารถช่วยเพิ่มการเคลื่อนที่ของไอออนได้อย่างมาก ทั้งแบบแอคทีฟและแบบพาสซีฟ โดยเกิดขึ้นผ่านกลไกการขนส่งแบบแอคทีฟและการแพร่แบบอำนวยความสะดวกโครงสร้างสองประเภทที่มีบทบาทสำคัญที่สุดคือ ช่องไอออนและปั๊มไอออนซึ่งโดยทั่วไปแล้วมักเกิดจากการรวมตัวของโมเลกุลโปรตีน ช่องไอออนเป็นทางผ่านที่ไอออนสามารถเคลื่อนที่ได้ ในกรณีส่วนใหญ่ ช่องไอออนจะยอมให้เฉพาะไอออนบางชนิดผ่านได้ (เช่น โซเดียมและโพแทสเซียม แต่ไม่ใช่คลอไรด์หรือแคลเซียม) และบางครั้งการซึมผ่านจะแตกต่างกันไปขึ้นอยู่กับทิศทางการเคลื่อนที่ของไอออน ปั๊มไอออน หรือที่เรียกว่าตัวขนส่งไอออนหรือโปรตีนพาหะ จะขนส่งไอออนบางชนิดจากด้านหนึ่งของเยื่อหุ้มเซลล์ไปยังอีกด้านหนึ่งอย่างแอคทีฟ บางครั้งใช้พลังงานที่ได้จากกระบวนการเมตาบอลิซึมในการทำเช่นนั้น
ปั๊มไอออน

ปั๊มไอออนเป็นโปรตีนเมมเบรนแบบบูรณาการที่ทำหน้าที่ขนส่งแบบแอคทีฟ กล่าวคือ ใช้พลังงานเซลล์ (ATP) เพื่อ "ปั๊ม" ไอออนสวนทางกับความเข้มข้นของไอออน[ 14 ]ปั๊มไอออนดังกล่าวจะรับไอออนจากด้านหนึ่งของเมมเบรน (ทำให้ความเข้มข้นลดลง) และปล่อยไอออนออกไปอีกด้านหนึ่ง (ทำให้ความเข้มข้นเพิ่มขึ้น)
ปั๊มไอออนที่เกี่ยวข้องกับศักยภาพการกระทำมากที่สุดคือปั๊มโซเดียม-โพแทสเซียมซึ่งขนส่งไอออนโซเดียม 3 ตัวออกจากเซลล์และไอออนโพแทสเซียม 2 ตัวเข้าไป[ 15 ] [ 16 ] ผลที่ตามมาคือ ความเข้มข้นของไอออนโพแทสเซียม K +ภายในเซลล์ประสาทจะมากกว่าความเข้มข้นภายนอกประมาณ 30 เท่า ในขณะที่ความเข้มข้นของโซเดียมภายนอกจะมากกว่าภายในประมาณ 5 เท่า[ 16 ] [ 17 ] [ 18 ] ในทำนองเดียวกัน ไอออนอื่นๆ ก็มีความ เข้มข้นที่แตกต่างกันทั้งภายในและภายนอกเซลล์ประสาท เช่นแคลเซียมคลอไรด์และแมกนีเซียม[ 18 ]
หากจำนวนของไอออนแต่ละชนิดเท่ากัน ปั๊มโซเดียม-โพแทสเซียมจะเป็นกลางทางไฟฟ้า แต่เนื่องจากการแลกเปลี่ยนแบบสามต่อสอง ทำให้เกิดการเคลื่อนที่สุทธิของประจุบวกหนึ่งตัวจากภายในเซลล์ไปยังภายนอกเซลล์ในแต่ละรอบ ซึ่งส่งผลให้เกิดความต่างศักย์ไฟฟ้าเป็นบวก ปั๊มนี้มีผลสามประการ: (1) ทำให้ความเข้มข้นของโซเดียมสูงในพื้นที่ภายนอกเซลล์และต่ำในพื้นที่ภายในเซลล์ (2) ทำให้ความเข้มข้นของโพแทสเซียมสูงในพื้นที่ภายในเซลล์และต่ำในพื้นที่ภายนอกเซลล์ (3) ทำให้พื้นที่ภายในเซลล์มีศักย์ไฟฟ้าเป็นลบเมื่อเทียบกับพื้นที่ภายนอกเซลล์
ปั๊มโซเดียม-โพแทสเซียมทำงานค่อนข้างช้า หากเซลล์เริ่มต้นด้วยความเข้มข้นของโซเดียมและโพแทสเซียมที่เท่ากันทุกจุด ปั๊มจะต้องใช้เวลาหลายชั่วโมงในการสร้างสมดุล ปั๊มทำงานอย่างต่อเนื่อง แต่ประสิทธิภาพจะลดลงเรื่อยๆ เมื่อความเข้มข้นของโซเดียมและโพแทสเซียมที่พร้อมสำหรับการสูบฉีดลดลง
ปั๊มไอออนมีอิทธิพลต่อศักยภาพการกระทำโดยการสร้างอัตราส่วนสัมพัทธ์ของความเข้มข้นของไอออนภายในเซลล์และภายนอกเซลล์เท่านั้น ศักยภาพการกระทำเกี่ยวข้องกับการเปิดและปิดช่องไอออนเป็นหลัก ไม่ใช่ปั๊มไอออน หากปั๊มไอออนถูกปิดโดยการกำจัดแหล่งพลังงาน หรือโดยการเพิ่มสารยับยั้ง เช่นโอวาเบนแอกซอนยังคงสามารถสร้างศักยภาพการกระทำได้หลายแสนครั้งก่อนที่แอมพลิจูดจะเริ่มลดลงอย่างมีนัยสำคัญ[ 14 ] โดยเฉพาะอย่างยิ่ง ปั๊มไอออนไม่มีบทบาทสำคัญในการคืนสภาพขั้วของเยื่อหุ้มเซลล์หลังจากศักยภาพการกระทำ[ 9 ]
ปั๊มไอออนอีกตัวที่มีความสำคัญต่อการทำงานคือตัวแลกเปลี่ยนโซเดียม-แคลเซียมปั๊มนี้ทำงานในลักษณะที่คล้ายคลึงกับปั๊มโซเดียม-โพแทสเซียม แต่ในแต่ละรอบจะแลกเปลี่ยน Na + สามไอออน จากพื้นที่ภายนอกเซลล์กับ Ca ++ หนึ่ง ไอออนจากพื้นที่ภายในเซลล์ เนื่องจากกระแสประจุสุทธิไหลเข้า ปั๊มนี้จึงทำงาน "ลงเนิน" ในทางปฏิบัติ และดังนั้นจึงไม่ต้องการแหล่งพลังงานใดๆ ยกเว้นแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ ผลที่สำคัญที่สุดของมันคือการสูบแคลเซียมออกไปภายนอกเซลล์ มันยังช่วยให้โซเดียมไหลเข้าภายในเซลล์ด้วย ซึ่งเป็นการต้านการทำงานของปั๊มโซเดียม-โพแทสเซียม แต่เนื่องจากความเข้มข้นโดยรวมของโซเดียมและโพแทสเซียมสูงกว่าความเข้มข้นของแคลเซียมมาก ผลกระทบนี้จึงค่อนข้างไม่สำคัญ ผลลัพธ์สุทธิของตัวแลกเปลี่ยนโซเดียม-แคลเซียมคือ ในสภาวะพัก ความเข้มข้นของแคลเซียมภายในเซลล์จะต่ำมาก
ช่องไอออน

ช่องไอออนเป็นโปรตีนเมมเบรนแบบฝังตัวที่มีรูพรุนซึ่งไอออนสามารถเดินทางผ่านระหว่างพื้นที่นอกเซลล์และภายในเซลล์ได้ ช่องส่วนใหญ่มีความจำเพาะ (เลือก) สำหรับไอออนหนึ่งชนิด ตัวอย่างเช่น ช่องโพแทสเซียมส่วนใหญ่มีอัตราส่วนการเลือก 1000:1 สำหรับโพแทสเซียมเหนือโซเดียม แม้ว่าไอออนโพแทสเซียมและโซเดียมจะมีประจุเดียวกันและแตกต่างกันเพียงเล็กน้อยในรัศมี รูพรุนของช่องมักมีขนาดเล็กมากจนไอออนต้องผ่านในลำดับเรียงเดี่ยว[ 20 ] รูพรุนของช่องสามารถเปิดหรือปิดเพื่อให้ไอออนผ่านได้ แม้ว่าช่องจำนวนหนึ่งจะแสดงระดับการนำไฟฟ้าย่อยต่างๆ เมื่อช่องเปิด ไอออนจะซึมผ่านรูพรุนของช่องตามความชันของความเข้มข้นข้ามเยื่อหุ้มเซลล์สำหรับไอออนนั้นๆ อัตราการไหลของไอออนผ่านช่อง เช่น แอมพลิจูดกระแสของช่องเดี่ยว จะถูกกำหนดโดยการนำไฟฟ้าสูงสุดของช่องและแรงขับทางเคมีไฟฟ้าสำหรับไอออนนั้น ซึ่งเป็นความแตกต่างระหว่างค่าทันทีของศักย์เยื่อหุ้มเซลล์และค่าของศักย์ย้อนกลับ[ 21 ]
ช่องอาจมีสถานะที่แตกต่างกันได้หลายสถานะ (ซึ่งสอดคล้องกับโครงสร้าง ที่แตกต่างกัน ของโปรตีน) แต่แต่ละสถานะดังกล่าวจะเป็นแบบเปิดหรือปิด โดยทั่วไป สถานะปิดจะสอดคล้องกับการหดตัวของรูพรุน ทำให้ไอออนไม่สามารถผ่านได้ หรือส่วนแยกของโปรตีนที่ปิดกั้นรูพรุน ตัวอย่างเช่น ช่องโซเดียมที่ขึ้นอยู่กับแรงดันไฟฟ้าจะเกิดการปิดใช้งานซึ่งส่วนหนึ่งของโปรตีนจะแกว่งเข้าไปในรูพรุนและปิดผนึก[ 22 ] การปิดใช้งานนี้จะปิดกระแสโซเดียมและมีบทบาทสำคัญในศักยภาพการกระทำ
ช่องไอออนสามารถจำแนกได้ตามการตอบสนองต่อสิ่งแวดล้อม[ 23 ]ตัวอย่างเช่น ช่องไอออนที่เกี่ยวข้องกับศักยภาพการกระทำเป็นช่องที่ไวต่อแรงดันไฟฟ้าพวกมันจะเปิดและปิดเพื่อตอบสนองต่อแรงดันไฟฟ้าข้ามเยื่อหุ้มเซลล์ช่องที่ควบคุมด้วยลิแกนด์เป็นอีกกลุ่มหนึ่งที่สำคัญ ช่องไอออนเหล่านี้จะเปิดและปิดเพื่อตอบสนองต่อการจับตัวของโมเลกุลลิแกนด์เช่นสารสื่อประสาทช่องไอออนอื่นๆ จะเปิดและปิดด้วยแรงทางกล ช่องไอออนอื่นๆ เช่น ช่องของเซลล์ประสาทรับความรู้สึกจะเปิดและปิดเพื่อตอบสนองต่อสิ่งเร้าอื่นๆ เช่น แสง อุณหภูมิ หรือความดัน
ช่องทางการรั่วไหล
ช่องรั่วไหลเป็นช่องไอออนชนิดที่ง่ายที่สุด เนื่องจากความสามารถในการซึมผ่านค่อนข้างคงที่ ช่องรั่วไหลที่มีความสำคัญมากที่สุดในเซลล์ประสาทคือช่องโพแทสเซียมและช่องคลอไรด์ แม้แต่ช่องเหล่านี้ก็ยังไม่คงที่อย่างสมบูรณ์แบบในคุณสมบัติของมัน ประการแรก ช่องส่วนใหญ่ขึ้นอยู่กับแรงดันไฟฟ้า กล่าวคือมันนำไฟฟ้าได้ดีกว่าในทิศทางหนึ่งมากกว่าอีกทิศทางหนึ่ง (กล่าวอีกนัยหนึ่งคือ มันเป็นตัวเรียงกระแส ) ประการที่สอง บางช่องสามารถถูกปิดได้ด้วยสารเคมีที่จับกับช่อง แม้ว่ามันจะไม่จำเป็นต้องมีสารที่จับกับช่องเพื่อการทำงานก็ตาม
ช่องที่ควบคุมด้วยลิแกนด์

ช่องไอออนที่ควบคุมด้วยลิแกนด์คือช่องที่มีความสามารถในการซึมผ่านเพิ่มขึ้นอย่างมากเมื่อลิแกนด์ทางเคมีบางชนิดจับกับโครงสร้างโปรตีน เซลล์สัตว์มีช่องเหล่านี้หลายร้อยหรือหลายพันชนิด กลุ่มย่อยจำนวนมากทำหน้าที่เป็นตัวรับสารสื่อประสาทโดยจะพบที่ บริเวณ หลังไซแนปส์และลิแกนด์ทางเคมีที่ควบคุมช่องเหล่านี้จะถูกปล่อยออกมาจากปลายแอกซอน ก่อนไซแนปส์ ตัวอย่างหนึ่งของช่องประเภทนี้คือตัวรับ AMPAซึ่งเป็นตัวรับสารสื่อประสาทกลูตาเมตเมื่อถูกกระตุ้นจะยอมให้ไอออนโซเดียมและโพแทสเซียมผ่านได้ อีกตัวอย่างหนึ่งคือตัวรับGABA A ซึ่ง เป็นตัวรับสารสื่อประสาทGABAเมื่อถูกกระตุ้นจะยอมให้ไอออนคลอไรด์ผ่านได้
ตัวรับสารสื่อประสาทจะถูกกระตุ้นโดยลิแกนด์ที่ปรากฏในบริเวณภายนอกเซลล์ แต่ยังมีช่องสัญญาณที่ควบคุมโดยลิแกนด์ประเภทอื่น ๆ ซึ่งถูกควบคุมโดยปฏิกิริยาที่เกิดขึ้นภายในเซลล์
ช่องสัญญาณที่ขึ้นอยู่กับแรงดันไฟฟ้า
ช่องไอออนที่ควบคุมด้วยแรงดันไฟฟ้าหรือที่เรียกว่าช่องไอออนที่ขึ้นอยู่กับแรงดันไฟฟ้าคือช่องที่มีการซึมผ่านได้ถูกควบคุมโดยศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ ช่องไอออนกลุ่มนี้เป็นกลุ่มใหญ่มากอีกกลุ่มหนึ่ง โดยแต่ละกลุ่มมีคุณสมบัติในการเลือกไอออนและมีความขึ้นอยู่กับแรงดันไฟฟ้าเฉพาะตัว หลายช่องยังขึ้นอยู่กับเวลาด้วย กล่าวคือ ช่องไอออนเหล่านี้จะไม่ตอบสนองต่อการเปลี่ยนแปลงแรงดันไฟฟ้าทันที แต่จะตอบสนองหลังจากช่วงเวลาหนึ่งผ่านไปแล้ว
หนึ่งในสมาชิกที่สำคัญที่สุดของกลุ่มนี้คือช่องโซเดียมแบบควบคุมด้วยแรงดันไฟฟ้าชนิดหนึ่ง ซึ่งเป็นพื้นฐานของศักย์ไฟฟ้าแอ็กชัน—บางครั้งเรียกว่าช่องโซเดียมฮอดจ์กิน-ฮักซ์ลีย์เนื่องจากได้รับการจำแนกลักษณะครั้งแรกโดยอลัน ลอยด์ ฮอดจ์กินและแอนดรูว์ ฮักซ์ลีย์ในงานวิจัยที่ได้รับรางวัลโนเบลเกี่ยวกับสรีรวิทยาของศักย์ไฟฟ้าแอ็กชัน ช่องนี้จะปิดอยู่ที่ระดับแรงดันไฟฟ้าขณะพัก แต่จะเปิดอย่างฉับพลันเมื่อแรงดันไฟฟ้าเกินเกณฑ์ที่กำหนด ทำให้ไอออนโซเดียมจำนวนมากไหลเข้ามา ส่งผลให้เกิดการเปลี่ยนแปลงอย่างรวดเร็วมากในศักย์เยื่อหุ้มเซลล์ การฟื้นตัวจากศักย์ไฟฟ้าแอ็กชันขึ้นอยู่กับช่องโพแทสเซียมแบบควบคุมด้วยแรงดันไฟฟ้าชนิดหนึ่ง ซึ่งจะปิดอยู่ที่ระดับแรงดันไฟฟ้าขณะพัก แต่จะเปิดออกเนื่องจากการเปลี่ยนแปลงแรงดันไฟฟ้าอย่างมากที่เกิดขึ้นระหว่างศักย์ไฟฟ้าแอ็กชัน
ศักยภาพย้อนกลับ
ศักย์ย้อนกลับ (หรือศักย์สมดุล ) ของไอออน คือค่าของแรงดันไฟฟ้าข้ามเยื่อหุ้มเซลล์ที่แรงแพร่และแรงไฟฟ้าหักล้างกัน ทำให้ไม่มีการไหลสุทธิของไอออนผ่านเยื่อหุ้มเซลล์ นั่นหมายความว่าแรงดันไฟฟ้าข้ามเยื่อหุ้มเซลล์จะต้านแรงแพร่ของไอออนอย่างพอดี ทำให้กระแสสุทธิของไอออนผ่านเยื่อหุ้มเซลล์เป็นศูนย์และไม่เปลี่ยนแปลง ศักย์ย้อนกลับมีความสำคัญเพราะมันให้ค่าแรงดันไฟฟ้าที่กระทำต่อช่องทางที่ไอออนนั้นสามารถผ่านได้ กล่าวอีกนัยหนึ่งคือ มันให้ค่าแรงดันไฟฟ้าที่เกิดจากความแตกต่างของความเข้มข้นของไอออนเมื่อมันทำหน้าที่เหมือน แบตเตอรี่
ศักยภาพสมดุลของไอออนเฉพาะมักจะถูกกำหนดด้วยสัญลักษณ์E ionศักยภาพสมดุลสำหรับไอออนใดๆ สามารถคำนวณได้โดยใช้สมการเนิร์นสต์ [ 24 ] ตัวอย่างเช่น ศักยภาพย้อนกลับสำหรับไอออนโพแทสเซียมจะเป็นดังนี้:
![{\displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}\ln {\frac {[K^{+}]_{o}}{[K^{+}]_{i}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/f898ec5e5474736ecfe73462fdcb4b4d92a21618)
ที่ไหน
- E eq,K + = ศักย์สมดุลของโพแทสเซียม วัดเป็นโวลต์
- R = ค่าคงที่ของแก๊สสากลเท่ากับ 8.314 จูล ·K −1 ·mol −1
- T = อุณหภูมิสัมบูรณ์วัดเป็นเคลวิน (= K = องศาเซลเซียส + 273.15)
- z = จำนวนประจุพื้นฐานของไอออนที่เกี่ยวข้องในปฏิกิริยา
- F = ค่าคงที่ฟาราเดย์เท่ากับ 96,485 คูลอมบ์ ·โมล⁻¹หรือ จูล·โวลต์⁻¹ ·โมล⁻¹
- [K + ] o = ความเข้มข้นของโพแทสเซียมภายนอกเซลล์ วัดในหน่วย mol ·m −3หรือ mmol·l −1
- [K + ] i = ความเข้มข้นของโพแทสเซียมภายในเซลล์
แม้ว่าไอออนสองชนิดที่แตกต่างกันจะมีประจุเดียวกัน (เช่น K +และ Na + ) แต่ก็ยังสามารถมีศักย์สมดุลที่แตกต่างกันมากได้ หากความเข้มข้นภายนอกและ/หรือภายในแตกต่างกัน ตัวอย่างเช่น ศักย์สมดุลของโพแทสเซียมและโซเดียมในเซลล์ประสาท ศักย์สมดุลของโพแทสเซียมE Kคือ −84 mV เมื่อมีโพแทสเซียม 5 mM ภายนอกและ 140 mM ภายใน ในทางกลับกัน ศักย์สมดุลของโซเดียม E Naคือประมาณ +66 mV เมื่อมีโซเดียมประมาณ 12 mM ภายในและ 140 mM ภายนอก[หมายเหตุ 1 ]
การเปลี่ยนแปลงของศักย์เยื่อหุ้มเซลล์ระหว่างการพัฒนา
ศักย์เยื่อหุ้มเซลล์ขณะพักของเซลล์ประสาทจะเปลี่ยนแปลงไปในระหว่างการพัฒนาของสิ่งมีชีวิต เพื่อให้เซลล์ประสาทสามารถทำหน้าที่ได้อย่างเต็มที่ในวัยผู้ใหญ่ ศักย์ของเซลล์ประสาทจะต้องได้รับการควบคุมอย่างเข้มงวดในระหว่างการพัฒนา เมื่อสิ่งมีชีวิตพัฒนาไปเรื่อยๆ ศักย์เยื่อหุ้มเซลล์ขณะพักก็จะยิ่งเป็นลบมากขึ้น[ 25 ]เซลล์เกลียก็มีการเปลี่ยนแปลงและเพิ่มจำนวนขึ้นตามการพัฒนาของสมอง[ 26 ] การเพิ่มจำนวนของเซลล์เกลียเหล่านี้ทำให้สิ่งมีชีวิตมีความสามารถในการควบคุมโพแทสเซียม ภายนอกเซลล์ได้มากขึ้น การลดลงของโพแทสเซียมภายนอกเซลล์อาจทำให้ศักย์เยื่อหุ้มเซลล์ลดลงถึง 35 mV [ 27 ]
ความสามารถในการกระตุ้นของเซลล์
ความตื่นตัวของเซลล์คือการเปลี่ยนแปลงของศักยภาพเยื่อหุ้มเซลล์ซึ่งจำเป็นต่อการตอบสนองของเซลล์ในเนื้อเยื่อต่างๆ ความตื่นตัวของเซลล์เป็นคุณสมบัติที่ถูกเหนี่ยวนำในช่วงต้นของการเกิดตัวอ่อน[ 28 ]ความตื่นตัวของเซลล์ยังถูกกำหนดให้เป็นความง่ายในการกระตุ้นการตอบสนอง[ 29 ] ศักยภาพ ขณะพักและศักยภาพเกณฑ์เป็นพื้นฐานของความตื่นตัวของเซลล์ และกระบวนการเหล่านี้เป็นพื้นฐานสำหรับการสร้างศักยภาพแบบไล่ระดับและศักยภาพการกระทำ
ตัวควบคุมที่สำคัญที่สุดของการกระตุ้นเซลล์คือ ความเข้มข้นของอิเล็ก โทรไลต์ ภายนอกเซลล์ (เช่น Na + , K + , Ca2 + , Cl− , Mg2 + )และโปรตีนที่เกี่ยวข้อง โปรตีนสำคัญที่ควบคุมการกระตุ้นเซลล์ ได้แก่ช่องไอออนที่ควบคุมด้วยแรง ดันไฟฟ้า ตัวขนส่งไอออน (เช่นNa+/K+-ATPase , ตัวขนส่งแมกนีเซียม , ตัวขนส่งกรด-เบส ) ตัวรับเมมเบรนและช่องที่ควบคุมด้วยนิวคลีโอไทด์แบบวงจรที่กระตุ้นด้วยไฮเปอร์โพลาไรเซชัน [ 30 ] ตัวอย่างเช่นช่องโพแทสเซียมและตัวรับที่รับรู้แคลเซียมเป็นตัวควบคุมที่สำคัญของการกระตุ้นในเซลล์ประสาทเซลล์กล้ามเนื้อหัวใจและเซลล์ที่กระตุ้นได้อื่นๆ อีกมากมาย เช่นแอสโทรไซต์[ 31 ]ไอออนแคลเซียมยังเป็นตัวส่งสัญญาณรองที่ สำคัญที่สุด ในการส่งสัญญาณของเซลล์ ที่กระตุ้นได้ การกระตุ้นตัวรับไซแนปส์จะเริ่มต้นการเปลี่ยนแปลงที่ยั่งยืนในการกระตุ้นของเซลล์ประสาท[ 32 ]ฮอร์โมนไทรอยด์ต่อมหมวกไตและฮอร์โมนอื่นๆ ยังควบคุมการกระตุ้นของเซลล์ด้วย ตัวอย่างเช่นโปรเจสเตอโรนและเอสโตรเจนจะปรับการกระตุ้นของ เซลล์กล้ามเนื้อเรียบของมดลูก
เซลล์หลายประเภทถือว่ามีเยื่อหุ้มเซลล์ที่สามารถกระตุ้นได้ เซลล์ที่สามารถกระตุ้นได้ ได้แก่ เซลล์ ประสาท เซลล์กล้ามเนื้อ ( กล้าม เนื้อหัวใจ กล้ามเนื้อโครงร่าง กล้ามเนื้อเรียบ ) เซลล์บุ ผนังหลอดเลือด เซลล์เพอริไซต์ เซลล์จุกซ์ตาโก ล เม อรูลา ร์ เซลล์ อินเตอร์สติเชีย ลของคาฮาลเซลล์เยื่อบุผิวหลายประเภท(เช่นเซลล์เบต้าเซลล์อัลฟา เซลล์เดลต้าเซลล์เอ็นเทอโรเอนโดครีน เซลล์ นิวโรเอนโดครีนในปอด เซลล์ ไพ เนียล ) เซลล์ เกลีย (เช่น แอสโทร ไซต์) เซลล์ ตัวรับแรงกล (เช่นเซลล์ขนและเซลล์เมอร์เคล ) เซลล์ ตัวรับสารเคมี (เช่นเซลล์กลอมัสตัวรับรส ) เซลล์พืช บางชนิด และอาจรวมถึงเซลล์ภูมิคุ้มกันด้วย[ 33 ]แอสโทรไซต์แสดงการกระตุ้นที่ไม่ใช่ไฟฟ้าโดยอาศัยการเปลี่ยนแปลงของแคลเซียมภายในเซลล์ที่เกี่ยวข้องกับการแสดงออกของตัวรับหลายชนิด ซึ่งพวกมันสามารถตรวจจับสัญญาณไซแนปส์ได้ ในเซลล์ประสาท มีคุณสมบัติของเยื่อหุ้มเซลล์ที่แตกต่างกันในบางส่วนของเซลล์ ตัวอย่างเช่น ความสามารถในการกระตุ้นของเดนไดรต์ทำให้เซลล์ประสาทมีความสามารถในการตรวจจับการเกิดขึ้นพร้อมกันของอินพุตที่แยกจากกันในเชิงพื้นที่[ 34 ]
วงจรสมมูล

นักสรีรวิทยาไฟฟ้าจำลองผลกระทบของความแตกต่างของความเข้มข้นของไอออน ช่องไอออน และความจุของเยื่อหุ้มเซลล์ในรูปของวงจรสมมูลซึ่งมีจุดประสงค์เพื่อแสดงคุณสมบัติทางไฟฟ้าของเยื่อหุ้มเซลล์ส่วนเล็กๆ วงจรสมมูลประกอบด้วยตัวเก็บประจุขนานกับเส้นทางสี่เส้น แต่ละเส้นประกอบด้วยแบตเตอรี่อนุกรมที่มีค่าการนำไฟฟ้าแปรผันได้ ค่าความจุถูกกำหนดโดยคุณสมบัติของชั้นไขมันสองชั้น และถือว่าคงที่ เส้นทางขนานทั้งสี่เส้นมาจากไอออนหลักตัวใดตัวหนึ่ง ได้แก่ โซเดียม โพแทสเซียม คลอไรด์ และแคลเซียม แรงดันไฟฟ้าของแต่ละเส้นทางไอออนถูกกำหนดโดยความเข้มข้นของไอออนในแต่ละด้านของเยื่อหุ้มเซลล์ ดู ส่วน ศักย์ย้อนกลับด้านบน ค่าการนำไฟฟ้าของแต่ละเส้นทางไอออน ณ จุดใดๆ ในเวลาใดๆ ถูกกำหนดโดยสถานะของช่องไอออนทั้งหมดที่อาจยอมให้ไอออนนั้นผ่านได้ รวมถึงช่องรั่วไหล ช่องที่ควบคุมโดยลิแกนด์ และช่องไอออนที่ควบคุมโดยแรงดันไฟฟ้า

สำหรับความเข้มข้นของไอออนคงที่และค่าการนำไฟฟ้าของช่องไอออนคงที่ วงจรสมมูลสามารถลดทอนลงได้อีกโดยใช้สมการของโกลด์แมนดังที่อธิบายไว้ด้านล่าง ให้เป็นวงจรที่มีตัวเก็บประจุขนานกับแบตเตอรี่และตัวนำ ในทางไฟฟ้า นี่คือวงจรRC (วงจรความต้านทาน-ตัวเก็บประจุ) และคุณสมบัติทางไฟฟ้าของมันนั้นง่ายมาก เริ่มต้นจากสถานะเริ่มต้นใดๆ กระแสที่ไหลผ่านตัวนำหรือตัวเก็บประจุจะลดลงตามการเปลี่ยนแปลงของเวลาแบบเอกซ์โปเนนเชียล โดยมีค่าคงที่เวลาτ = RCโดยที่Cคือความจุของเยื่อหุ้มเซลล์ และR = 1/g netคือความต้านทานสุทธิ สำหรับสถานการณ์จริง ค่าคงที่เวลามักจะอยู่ในช่วง 1–100 มิลลิวินาที ในกรณีส่วนใหญ่ การเปลี่ยนแปลงของการนำไฟฟ้าของช่องไอออนเกิดขึ้นในระยะเวลาที่เร็วกว่า ดังนั้นวงจร RC จึงไม่ใช่การประมาณที่ดี อย่างไรก็ตาม สมการเชิงอนุพันธ์ที่ใช้ในการจำลองเยื่อหุ้มเซลล์มักจะเป็นสมการวงจร RC ที่ดัดแปลงแล้ว
ศักย์พัก

เมื่อศักย์เยื่อหุ้มเซลล์คงที่อยู่เป็นเวลานานโดยไม่เปลี่ยนแปลงอย่างมีนัยสำคัญ จะเรียกว่าศักย์พักหรือแรงดันไฟฟ้าพัก คำนี้ใช้สำหรับศักย์เยื่อหุ้มเซลล์ของเซลล์ที่ไม่สามารถถูกกระตุ้นได้ แต่ยังใช้สำหรับศักย์เยื่อหุ้มเซลล์ของเซลล์ที่สามารถถูกกระตุ้นได้ในกรณีที่ไม่มีการกระตุ้น ในเซลล์ที่สามารถถูกกระตุ้นได้ สถานะอื่นๆ ที่เป็นไปได้คือ ศักย์เยื่อหุ้มเซลล์แบบไล่ระดับ (ที่มีแอมพลิจูดแปรผันได้) และศักย์การกระทำ ซึ่งเป็นการเพิ่มขึ้นของศักย์เยื่อหุ้มเซลล์อย่างมากแบบทั้งหมดหรือไม่มีเลย ซึ่งมักจะเกิดขึ้นตามช่วงเวลาที่แน่นอน เซลล์ที่สามารถถูกกระตุ้นได้ ได้แก่เซลล์ประสาทเซลล์กล้ามเนื้อ และเซลล์หลั่งบางชนิดในต่อมต่างๆอย่างไรก็ตาม แม้ในเซลล์ประเภทอื่นๆ แรงดันไฟฟ้าของเยื่อหุ้มเซลล์ก็สามารถเปลี่ยนแปลงได้เพื่อตอบสนองต่อสิ่งเร้าจากสิ่งแวดล้อมหรือภายในเซลล์ ตัวอย่างเช่น การลดศักย์ของเยื่อหุ้มพลาสมาดูเหมือนจะเป็นขั้นตอนสำคัญในการตายของเซลล์แบบโปรแกรม[ 35 ]
ปฏิสัมพันธ์ที่สร้างศักยภาพพักจะถูกจำลองโดยสมการโกลด์แมน[ 36 ] ซึ่งมีรูป แบบ คล้ายกับสมการเนิร์นสต์ที่แสดงไว้ข้างต้น โดยอิงจากประจุของไอออนที่เกี่ยวข้อง รวมถึงความแตกต่างระหว่างความเข้มข้นภายในและภายนอก อย่างไรก็ตาม ยังคำนึงถึงการซึมผ่านสัมพัทธ์ของเยื่อหุ้มพลาสมาต่อไอออนแต่ละชนิดด้วย ในความเป็นจริง มีการซึมผ่านในระดับหนึ่งสำหรับไอออนหลายชนิดพร้อมกัน ดังนั้นศักยภาพเยื่อหุ้มเซลล์ขณะพักจึงไม่เหมือนกับศักยภาพเนิร์นสต์ใดๆ แต่จะอยู่ใกล้จุดสมดุลของการซึมผ่านสำหรับไอออนที่ซึมผ่านได้เด่นชัด (โดยปกติคือโพแทสเซียม โซเดียม และคลอไรด์) [ 37 ]สิ่งนี้สามารถแสดงได้ด้วยสมการแรงดันไฟฟ้าโกลด์แมน-ฮอดจ์กิน-แคทซ์ (GHK) ในเชิงแนวคิด: ไอออนที่มีการซึมผ่านได้มากกว่าจะส่งผลต่อ Vm มากขึ้น นี่คือเหตุผลที่ศักยภาพพักมักจะใกล้เคียงกับ EK (เนื่องจาก PK มักจะมีค่ามากที่สุดขณะพัก) แต่เป็นบวกเล็กน้อยเมื่อเทียบกับ ENa ขึ้นอยู่กับการรั่วไหลของโซเดียมและกิจกรรมทางไฟฟ้าอื่นๆ[ 6 ]
![{\displaystyle E_{m}={\frac {RT}{F}}\ln {\left({\frac {P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\mathrm {out} }+P_{\mathrm {Na} [\mathrm {Na} ^{+}]_{\mathrm {out} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{-}]_{\mathrm {in} }}{P_{\mathrm {K} }[\mathrm {K} ^{+}]_{\mathrm {in} }+P_{\mathrm {Na} }[\mathrm {Na} ^{+}]_{\mathrm {in} }+P_{\mathrm {Cl} }[\mathrm {Cl} ^{-}]_{\คณิตศาสตร์ {ออก} }}}\ขวา)}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/4b29151571b6fb807674bf342d01f178908a1a98)
ไอออนสามตัวที่ปรากฏในสมการนี้คือโพแทสเซียม (K + ) โซเดียม (Na + ) และคลอไรด์ (Cl − ) แคลเซียมถูกละเว้น แต่สามารถเพิ่มเข้าไปได้เพื่อจัดการกับสถานการณ์ที่แคลเซียมมีบทบาทสำคัญ[ 38 ] เนื่องจากคลอไรด์เป็นแอนไอออน จึงต้องได้รับการจัดการแตกต่างจากแคตไอออน โดยความเข้มข้นภายในเซลล์จะอยู่ในตัวเศษ และความเข้มข้นภายนอกเซลล์จะ อยู่ ในตัวส่วน ซึ่งกลับด้านจากแคตไอออนPiหมายถึงการซึมผ่านสัมพัทธ์ของไอออนชนิด i
โดยพื้นฐานแล้ว สูตรของโกลด์แมนแสดงศักยภาพของเยื่อหุ้มเซลล์เป็นค่าเฉลี่ยถ่วงน้ำหนักของศักยภาพย้อนกลับสำหรับไอออนแต่ละชนิด โดยถ่วงน้ำหนักด้วยค่าการซึมผ่าน (แม้ว่าศักยภาพของเยื่อหุ้มเซลล์จะเปลี่ยนแปลงประมาณ 100 mV ในระหว่างศักยภาพการกระทำ แต่ความเข้มข้นของไอออนภายในและภายนอกเซลล์จะไม่เปลี่ยนแปลงอย่างมีนัยสำคัญ พวกมันยังคงใกล้เคียงกับความเข้มข้นของพวกมันเมื่อเยื่อหุ้มเซลล์อยู่ที่ศักยภาพพัก) ในเซลล์สัตว์ส่วนใหญ่ ค่าการซึมผ่านของโพแทสเซียมจะสูงกว่าค่าการซึมผ่านของโซเดียมมากในสภาวะพัก ดังนั้น ศักยภาพพักจึงมักจะใกล้เคียงกับศักยภาพย้อนกลับของโพแทสเซียม[ 39 ] [ 40 ] ค่าการซึมผ่านของคลอไรด์อาจสูงพอที่จะมีนัยสำคัญ แต่ต่างจากไอออนอื่นๆ คลอไรด์ไม่ได้ถูกสูบอย่างแข็งขัน ดังนั้นจึงสมดุลที่ศักยภาพย้อนกลับที่ใกล้เคียงกับศักยภาพพักที่กำหนดโดยไอออนอื่นๆ มาก
ค่าศักย์เยื่อหุ้มเซลล์ขณะพักในเซลล์สัตว์ส่วนใหญ่มักจะแตกต่างกันระหว่างศักย์ผกผันของโพแทสเซียม (โดยปกติประมาณ −80 mV) และประมาณ −40 mV ศักย์ขณะพักในเซลล์ที่ตื่นตัวได้ (สามารถสร้างศักย์การกระทำได้) มักจะอยู่ใกล้ −60 mV—แรงดันไฟฟ้าที่ลดลงมากกว่านี้จะทำให้เกิดศักย์การกระทำขึ้นเอง เซลล์ที่ยังไม่เจริญเต็มที่หรือยังไม่แตกต่างจะแสดงค่าแรงดันไฟฟ้าขณะพักที่แปรผันได้สูง โดยปกติจะมีค่าบวกมากกว่าในเซลล์ที่แตกต่างอย่างมีนัยสำคัญ[ 41 ]ในเซลล์ดังกล่าว ค่าศักย์ขณะพักจะสัมพันธ์กับระดับความแตกต่าง: ในบางกรณี เซลล์ที่ยังไม่แตกต่างอาจไม่แสดงความแตกต่างของแรงดันไฟฟ้าข้ามเยื่อหุ้มเซลล์เลย
การรักษาระดับศักยภาพพักตัวอาจมีค่าใช้จ่ายทางเมตาบอลิซึมสูงสำหรับเซลล์ เนื่องจากต้องใช้การปั๊มไอออนอย่างต่อเนื่องเพื่อชดเชยการสูญเสียเนื่องจากช่องทางการรั่วไหล ค่าใช้จ่ายจะสูงที่สุดเมื่อการทำงานของเซลล์ต้องการค่าแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ที่ลดลงเป็นพิเศษ ตัวอย่างเช่น ศักยภาพพักตัวในเซลล์รับแสงของ แมลงวันหัวเขียว ( Calliphora vicina ) ที่ปรับตัวเข้ากับแสงแดด อาจสูงถึง −30 mV [ 42 ] ศักยภาพ ของเยื่อหุ้ม เซลล์ที่สูงขึ้นนี้ทำให้เซลล์สามารถตอบสนองต่อสิ่งเร้าทางสายตาได้อย่างรวดเร็วมาก ค่าใช้จ่ายคือการรักษาระดับศักยภาพพักตัวอาจ ใช้ATPในเซลล์ทั้งหมดมากกว่า 20% [ 43 ]
ในทางกลับกัน ศักยภาพการพักตัวที่สูงในเซลล์ที่ยังไม่แยกความแตกต่างนั้น ไม่จำเป็นต้องก่อให้เกิดต้นทุนการเผาผลาญที่สูงเสมอไป ความขัดแย้งที่เห็นได้ชัดนี้ได้รับการแก้ไขโดยการตรวจสอบที่มาของศักยภาพการพักตัวนั้น เซลล์ที่แยกความแตกต่างน้อยนั้นมีลักษณะเฉพาะคือความต้านทานอินพุตที่สูงมาก[ 41 ]ซึ่งหมายความว่ามีช่องทางการรั่วไหลน้อยมากในระยะนี้ของชีวิตเซลล์ ผลที่เห็นได้ชัดคือ การซึมผ่านของโพแทสเซียมจะคล้ายกับของไอออนโซเดียม ซึ่งทำให้ศักยภาพการพักตัวอยู่ระหว่างศักยภาพย้อนกลับของโซเดียมและโพแทสเซียมดังที่ได้กล่าวไว้ข้างต้น กระแสการรั่วไหลที่ลดลงยังหมายความว่าไม่จำเป็นต้องมีการสูบฉีดอย่างแข็งขันเพื่อชดเชย ดังนั้นจึงมีต้นทุนการเผาผลาญต่ำ
การวัดศักย์เยื่อหุ้มเซลล์
วิธีการแบบดั้งเดิมคือการบันทึกภายในเซลล์: วางอิเล็กโทรดหนึ่งตัวไว้ภายในเซลล์และอิเล็กโทรดอ้างอิงจะอยู่ภายนอก ทำให้สามารถวัด V m Perkins KL ได้โดยตรง เทคนิคการบันทึกและการกระตุ้นด้วยแรงดันและกระแสไฟฟ้าที่ติดกับเซลล์ในชิ้นส่วนสมอง[ 44 ]
สรีรวิทยาไฟฟ้าสมัยใหม่มักใช้แนวทางแพทช์แคลมป์ โดยใช้ไมโครปิเปตแก้วขนาดเล็กกดลงบนเยื่อหุ้มเซลล์เพื่อสร้างซีลที่มีความต้านทานสูงมาก (กิกะซีล) ซีลนี้จะแยกเยื่อหุ้มเซลล์ส่วนเล็กๆ ออกมา ทำให้นักวิจัยสามารถบันทึกสัญญาณไฟฟ้าเฉพาะได้ และขึ้นอยู่กับการกำหนดค่า สามารถควบคุมได้ว่าด้านใดของเยื่อหุ้มเซลล์จะสัมผัสกับสารละลายที่แตกต่างกัน[ 44 ]
การทดลองแบบ Patch-clamp มักจะดำเนินการในสองโหมด ได้แก่ โหมด current-clamp ซึ่งผู้ทำการทดลองจะฉีดกระแสไฟฟ้าที่กำหนดไว้และวัดว่าศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ (V m ) เปลี่ยนแปลงไปอย่างไรเมื่อเวลาผ่านไป และโหมด voltage-clamp ซึ่งผู้ทำการทดลองจะรักษาระดับแรงดันไฟฟ้าของเยื่อหุ้มเซลล์ไว้ที่ระดับที่เลือกไว้และวัดกระแสไอออนที่จำเป็นในการรักษาระดับนั้นไว้ โหมด voltage-clamp มีประโยชน์สำหรับการแยกกระแสโซเดียม โพแทสเซียม แคลเซียม หรือกระแสไซแนปส์[ 44 ]
การกำหนดค่าหลักของการวัดกระแสไฟฟ้าผ่านแผ่นเชื่อมต่อ (patch-clamp)
การบันทึกแบบเซลล์ต่อเซลล์:หลังจากเกิดกิกะซีลแล้ว เยื่อหุ้มเซลล์ยังคงสมบูรณ์ วิธีนี้มักใช้สำหรับการบันทึกแบบช่องสัญญาณเดียว[ 45 ]
การบันทึกเซลล์ทั้งหมด:เริ่มต้นจากซีลที่ติดกับเซลล์ แผ่นเมมเบรนขนาดเล็กใต้ปลายปิเปตจะแตกออก (โดยการดูดหรือพัลส์ไฟฟ้าสั้นๆ) ซึ่งทำให้ปิเปตสามารถเข้าถึงภายในเซลล์ได้ ทำให้ผู้วิจัยสามารถบันทึกกระแสไฟฟ้าของเซลล์ทั้งหมด (voltage-clamp) หรือการตอบสนองแรงดันไฟฟ้าของเซลล์ทั้งหมด (current-clamp) ได้[ 45 ]
แพทช์แบบกลับด้าน:จากซีลที่ติดกับเซลล์ ปิเปตจะถูกดึงออกเพื่อให้แพทช์ขนาดเล็กถูกตัดออก และด้านภายในเซลล์ของเยื่อหุ้มเซลล์จะถูกเปิดออกสู่สารละลาย วิธีนี้มีประโยชน์อย่างยิ่งสำหรับการทดสอบว่าช่องตอบสนองต่อลิแกนด์ภายในเซลล์อย่างไร เนื่องจากสามารถควบคุมสารละลายที่ด้านไซโตพลาสมิกได้[ 45 ]
แพทช์แบบนอกเซลล์:หลังจากเข้าสู่โหมดเซลล์ทั้งหมดแล้ว จะนำปิเปตกลับและปิดผนึกเยื่อหุ้มเซลล์ที่ปลายอีกครั้งเพื่อให้พื้นผิวนอกเซลล์สัมผัสกับสารละลาย วิธีนี้ใช้สำหรับการศึกษาช่องสัญญาณที่ควบคุมด้วยลิแกนด์และตัวรับไซแนปส์ เนื่องจากสามารถเพิ่มสารสื่อประสาทลงบนพื้นผิวด้านนอกได้อย่างรวดเร็วและวัดกระแสไฟฟ้าที่เกิดขึ้นได้[ 45 ]
แผ่นแปะแบบมีรูพรุน:แทนที่จะทำลายแผ่นเยื่อหุ้มเซลล์ทั้งหมด การใช้แผ่นแปะแบบมีรูพรุนจะใช้สารประกอบที่สร้างรูพรุน (เช่น ไนสตาตินหรือแอมโฟเทอริซิน บี) เพื่อเชื่อมต่อปิเปตกับเซลล์ด้วยไฟฟ้า ในขณะเดียวกันก็ลดการชะล้างสารภายในเซลล์ให้น้อยที่สุด[ 45 ]
การบันทึกแบบแพทช์หลวม:ในบางสถานการณ์ จะใช้ซีลที่มีความต้านทานต่ำกว่าเพื่อให้การบันทึกไม่รบกวนมากนัก วิธีนี้มักใช้สำหรับการบันทึกศักยภาพการกระทำหรือกิจกรรมเฉพาะที่โดยไม่รบกวนเยื่อหุ้มเซลล์อย่างมีนัยสำคัญ[ 45 ]
ศักยภาพที่แบ่งระดับ
ดังที่ได้อธิบายไว้ข้างต้น ศักยภาพ ณ จุดใด ๆ ในเยื่อหุ้มเซลล์ถูกกำหนดโดยความแตกต่างของความเข้มข้นของไอออนระหว่างบริเวณภายในและภายนอกเซลล์ และโดยการซึมผ่านของเยื่อหุ้มเซลล์ต่อไอออนแต่ละชนิด ความเข้มข้นของไอออนโดยปกติจะไม่เปลี่ยนแปลงอย่างรวดเร็ว (ยกเว้น Ca²⁺ ซึ่งความเข้มข้นพื้นฐานภายในเซลล์ต่ำมากจนแม้การไหลเข้าเพียงเล็กน้อยก็อาจเพิ่มขึ้นได้หลายเท่า) แต่การซึมผ่านของไอออนสามารถเปลี่ยนแปลงได้ในเวลาเพียงเสี้ยววินาที อันเป็นผลมาจากการกระตุ้นช่องไอออนที่ควบคุมโดยลิแกนด์ การเปลี่ยนแปลงของศักยภาพเยื่อหุ้มเซลล์อาจมีขนาดใหญ่หรือเล็ก ขึ้นอยู่กับจำนวนช่องไอออนที่ถูกกระตุ้นและชนิดของช่องเหล่านั้น และอาจยาวหรือสั้น ขึ้นอยู่กับระยะเวลาที่ช่องเหล่านั้นเปิดอยู่ การเปลี่ยนแปลงประเภทนี้เรียกว่าศักยภาพแบบค่อยเป็นค่อยไป (graded potentials ) ซึ่งแตกต่างจากศักยภาพการกระทำ (action potentials) ซึ่งมีแอมพลิจูดและระยะเวลาคงที่
จาก สมการของโกลด์แมน ที่แสดงไว้ข้างต้น จะเห็นได้ว่าผลของการเพิ่มการซึมผ่านของเยื่อหุ้มเซลล์ต่อไอออนชนิดใดชนิดหนึ่ง จะทำให้ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เคลื่อนไปทางศักย์ย้อนกลับของไอออนนั้น ดังนั้น การเปิดช่อง Na +จะทำให้ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เคลื่อนไปทางศักย์ย้อนกลับของ Na +ซึ่งโดยปกติจะอยู่ที่ประมาณ +100 mV ในทำนองเดียวกัน การเปิดช่อง K +จะทำให้ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เคลื่อนไปทางประมาณ −90 mV และการเปิดช่อง Cl− จะ ทำให้ศักย์ไฟฟ้า เคลื่อนไปทางประมาณ −70 mV (ศักย์พักของเยื่อหุ้มเซลล์ส่วนใหญ่) ดังนั้น ช่อง Na +จะทำให้ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เคลื่อนไปในทิศทางบวก ช่อง K + จะ ทำให้ศักย์ไฟฟ้าเคลื่อนไปในทิศทางลบ (ยกเว้นเมื่อเยื่อหุ้มเซลล์มีศักย์ไฟฟ้าสูงกว่าศักย์ย้อนกลับของ K + ) และช่อง Cl− มีแนวโน้มที่จะทำให้ศักย์ไฟฟ้าเคลื่อนไปทางศักย์พัก
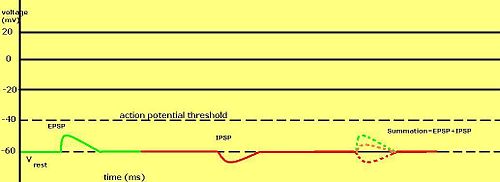
ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์แบบค่อยเป็นค่อยไปมีความสำคัญอย่างยิ่งในเซลล์ประสาทโดยเกิดขึ้นจากไซแนปส์ — การเปลี่ยนแปลงชั่วคราวของศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ที่เกิดจากการกระตุ้นไซแนปส์ด้วยศักย์ไฟฟ้าแบบค่อยเป็นค่อยไปหรือศักย์ไฟฟ้าแอคชั่นเพียงครั้งเดียว เรียกว่าศักย์ไฟฟ้าหลัง ไซแน ปส์ สารสื่อประสาทที่ทำหน้าที่เปิดช่อง Na +มักทำให้ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์เป็นบวกมากขึ้น ในขณะที่สารสื่อประสาทที่กระตุ้นช่อง K +มักทำให้ศักย์ไฟฟ้าเป็นลบมากขึ้น ส่วนสารสื่อประสาทที่ยับยั้งช่องเหล่านี้มักจะมีผลตรงกันข้าม
การที่ศักย์หลังไซแนปส์จะถูกพิจารณาว่าเป็นแบบกระตุ้นหรือแบบยับยั้งนั้น ขึ้นอยู่กับศักย์ย้อนกลับของไอออนในกระแสนั้น และเกณฑ์ที่เซลล์จะสร้างศักย์การกระทำ (ประมาณ −50 mV) กระแสหลังไซแนปส์ที่มีศักย์ย้อนกลับสูงกว่าเกณฑ์ เช่น กระแส Na + ทั่วไป จะถูกพิจารณาว่าเป็นแบบกระตุ้น กระแสที่มีศักย์ย้อนกลับต่ำกว่าเกณฑ์ เช่น กระแส K + ทั่วไป จะถูกพิจารณาว่าเป็นแบบยับยั้ง กระแสที่มีศักย์ย้อนกลับสูงกว่าศักย์พัก แต่ต่ำกว่าเกณฑ์ จะไม่กระตุ้นให้เกิดศักย์การกระทำด้วยตัวเอง แต่จะทำให้เกิดการแกว่งของศักย์เยื่อหุ้ม เซลล์ที่ต่ำกว่าเกณฑ์ ดังนั้น สารสื่อประสาทที่ทำหน้าที่เปิดช่อง Na + จะสร้าง ศักย์หลังไซแนปส์แบบกระตุ้นหรือ EPSP ในขณะที่สารสื่อประสาทที่ทำหน้าที่เปิดช่อง K +หรือ Cl− มัก จะสร้าง ศักย์หลังไซแนปส์แบบยับยั้งหรือ IPSP เมื่อช่องหลายประเภทเปิดพร้อมกันในช่วงเวลาเดียวกัน ศักย์หลังไซแนปส์ของช่องเหล่านั้นจะรวมกัน (บวกเข้าด้วยกัน)
ค่าอื่นๆ
จากมุมมองของชีวฟิสิกส์ ศักย์เยื่อหุ้มเซลล์ ขณะพักเป็นเพียงศักย์เยื่อหุ้มเซลล์ที่เกิดจากค่าการซึมผ่านของเยื่อหุ้มเซลล์ซึ่งเป็นค่าหลักเมื่อเซลล์อยู่ในสภาวะพัก สมการค่าเฉลี่ยถ่วงน้ำหนักข้างต้นใช้ได้เสมอ แต่แนวทางต่อไปนี้อาจมองเห็นภาพได้ง่ายกว่า ในช่วงเวลาใดเวลาหนึ่ง มีปัจจัยสองประการสำหรับไอออนที่กำหนดว่าไอออนนั้นจะมีอิทธิพลต่อศักย์เยื่อหุ้มเซลล์มากน้อยเพียงใด:
- แรงขับเคลื่อนของไอออนนั้น
- การซึมผ่านของไอออนนั้น
ถ้าแรงผลักดันสูง ไอออนก็จะถูก "ผลัก" ผ่านเยื่อหุ้มเซลล์ แต่ถ้าการซึมผ่านสูง ไอออนก็จะแพร่ผ่านเยื่อหุ้มเซลล์ได้ง่ายขึ้น
- แรงขับเคลื่อนคือแรงทางไฟฟ้าสุทธิที่มีอยู่เพื่อเคลื่อนไอออนนั้นผ่านเยื่อหุ้มเซลล์ คำนวณได้จากผลต่างระหว่างแรงดันไฟฟ้าที่ไอออน "ต้องการ" อยู่ (ศักย์สมดุล) กับศักย์เยื่อหุ้มเซลล์จริง ( E <sub>m</sub> ) ดังนั้น ในเชิงสูตร แรงขับเคลื่อนสำหรับไอออน = E <sub>m</sub> − E<sub> ion</sub>
- ตัวอย่างเช่น ที่ศักย์พักที่เราคำนวณไว้ก่อนหน้านี้ที่ −73 mV แรงขับเคลื่อนของโพแทสเซียมคือ 7 mV : (−73 mV) − (−80 mV) = 7 mV ส่วนแรงขับเคลื่อนของโซเดียมจะเป็น (−73 mV) − (60 mV) = −133 mV
- การซึมผ่านได้เป็นตัววัดว่าไอออนสามารถผ่านเยื่อหุ้มเซลล์ได้ง่ายเพียงใด โดยปกติจะวัดเป็นค่าการนำไฟฟ้า และหน่วยคือซีเมนส์ซึ่งสอดคล้องกับ 1 C·s −1 ·V −1นั่นคือ 1 คูลอมบ์ต่อวินาทีต่อศักย์ไฟฟ้า 1 โวลต์
ดังนั้น ในเยื่อหุ้มเซลล์ที่อยู่ในสภาวะพัก แรงขับเคลื่อนของโพแทสเซียมต่ำ แต่การซึมผ่านของมันกลับสูงมาก ในขณะที่โซเดียมมีแรงขับเคลื่อนมหาศาล แต่การซึมผ่านในสภาวะพักแทบไม่มีเลย ในกรณีนี้ โพแทสเซียมจะนำกระแสไฟฟ้าได้มากกว่าโซเดียมประมาณ 20 เท่า และด้วยเหตุนี้จึงมีอิทธิพลต่อE mมากกว่าโซเดียม ถึง 20 เท่า
อย่างไรก็ตาม ลองพิจารณากรณีอื่น นั่นคือจุดสูงสุดของศักย์ไฟฟ้าแอ ค ชั่น ในกรณีนี้ การซึมผ่านของ Na สูง และการซึมผ่านของ K ค่อนข้างต่ำ ดังนั้น เยื่อหุ้มเซลล์จึงเคลื่อนที่เข้าใกล้E Naและห่างจากE K
ยิ่งมีไอออนที่ผ่านเยื่อหุ้มเซลล์มากเท่าไร การทำนายศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ก็จะยิ่งซับซ้อนมากขึ้นเท่านั้น อย่างไรก็ตาม สามารถทำได้โดยใช้สมการโกลด์แมน-ฮอดจ์กิน-แคทซ์หรือสมการค่าเฉลี่ยถ่วงน้ำหนัก โดยการแทนค่าความแตกต่างของความเข้มข้นและความสามารถในการผ่านของไอออน ณ เวลาใดเวลาหนึ่ง ก็สามารถกำหนดศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ ณ ขณะนั้นได้ สมการ GHK หมายความว่า ณ เวลาใดเวลาหนึ่ง ค่าของศักย์ไฟฟ้าของเยื่อหุ้มเซลล์จะเป็นค่าเฉลี่ยถ่วงน้ำหนักของศักย์ไฟฟ้าสมดุลของไอออนที่ผ่านเยื่อหุ้มเซลล์ทั้งหมด โดย "การถ่วงน้ำหนัก" คือความสามารถในการผ่านสัมพัทธ์ของไอออนผ่านเยื่อหุ้มเซลล์
ผลกระทบและนัยยะต่างๆ
เซลล์ใช้พลังงานในการขนส่งไอออนและสร้างศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์ ในขณะเดียวกันก็ใช้ศักย์ไฟฟ้านี้ในการขนส่งไอออนและสารเมตาบอไลต์อื่นๆ เช่น น้ำตาล ศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์ของไมโทคอนเดรียเป็นตัวขับเคลื่อนการผลิตATPซึ่งเป็นหน่วยพลังงานพื้นฐานทางชีวภาพ
เซลล์อาจดึงพลังงานที่สะสมไว้ในศักย์พักมาใช้เพื่อขับเคลื่อนศักย์การกระทำหรือการกระตุ้นรูปแบบอื่นๆ การเปลี่ยนแปลงของศักย์เยื่อหุ้มเซลล์เหล่านี้ช่วยให้เกิดการสื่อสารกับเซลล์อื่นๆ (เช่นเดียวกับศักย์การกระทำ) หรือเริ่มต้นการเปลี่ยนแปลงภายในเซลล์ ซึ่งเกิดขึ้นในไข่เมื่อได้รับ การปฏิสนธิจากอสุจิ
การเปลี่ยนแปลงคุณสมบัติไดอิเล็กทริกของเยื่อหุ้มพลาสมาอาจทำหน้าที่เป็นตัวบ่งชี้ของสภาวะพื้นฐาน เช่น โรคเบาหวานและภาวะไขมันในเลือดสูง[ 46 ]
ในเซลล์ประสาท การเกิดศักย์ไฟฟ้าเริ่มต้นจากการไหลของไอออนโซเดียมเข้าสู่เซลล์ผ่านช่องโซเดียม ส่งผลให้เกิดการลดศักย์ไฟฟ้า ในขณะที่การฟื้นตัวเกี่ยวข้องกับการไหลของไอออนโพแทสเซียมออกไปภายนอกเซลล์ผ่านช่องโพแทสเซียม การไหลทั้งสองนี้เกิดขึ้นโดยการแพร่แบบพาสซีฟ
ปริมาณเกลืออาจกระตุ้นเซลล์ประสาทที่ยังทำงานอยู่ของเนื้อสดให้ทำงาน ทำให้เกิดอาการกล้ามเนื้อหดเกร็ง[ 47 ] [ 48 ] [ 49 ] [ 50 ] [ 51 ]
ดูเพิ่มเติม
- ชีวไฟฟ้าเคมี
- ศักยภาพเคมีออสโมติก
- ศักยภาพทางไฟฟ้าเคมี
- สมการโกลด์แมน
- ชีวฟิสิกส์ของเยื่อหุ้มเซลล์
- อาร์เรย์ไมโครอิเล็กโทรด
- การนำไฟฟ้าแบบกระโดด
- ศักยภาพพื้นผิว
- ผลกระทบของกิบบส์-ดอนแนน
- ศักยภาพของไซแนปส์
หมายเหตุ
- ^เครื่องหมายของ E Naและ E Kตรงข้ามกัน เนื่องจากความชันของความเข้มข้นของโพแทสเซียมมีทิศทางออกจากเซลล์ ในขณะที่ความชันของความเข้มข้นของโซเดียมมีทิศทางเข้าสู่เซลล์ ศักย์ไฟฟ้าของเยื่อหุ้มเซลล์ถูกกำหนดโดยเทียบกับภายนอกเซลล์ ดังนั้น ศักย์ไฟฟ้า −70 mV หมายความว่าภายในเซลล์มีประจุลบเมื่อเทียบกับภายนอกเซลล์
อ่านเพิ่มเติม
- Alberts และคณะ. ชีววิทยาระดับโมเลกุลของเซลล์ . สำนักพิมพ์ Garland; ฉบับพิมพ์ครั้งที่ 4 (มีนาคม 2545). ISBN 0-8153-3218-1ระดับปริญญาตรี
- กายตัน, อาร์เธอร์ ซี., จอห์น อี. ฮอลล์. ตำราสรีรวิทยาทางการแพทย์ . บริษัท ดับเบิลยูบี ซอนเดอร์ส; ฉบับที่ 10 (15 สิงหาคม 2543). ISBN 0-7216-8677-Xระดับปริญญาตรี
- Hille, B. ช่องไอออนของเยื่อหุ้มเซลล์ที่ไวต่อการกระตุ้น Sinauer Associates, Sunderland, MA, USA; ฉบับพิมพ์ครั้งที่ 1, 1984. ISBN 0-87893-322-0
- Nicholls, JG, Martin, AR และ Wallace, BG จากเซลล์ประสาทสู่สมองสำนักพิมพ์ Sinauer Associates, Inc. ซันเดอร์แลนด์, แมสซาชูเซตส์, สหรัฐอเมริกา ฉบับพิมพ์ครั้งที่ 3, 1992. ISBN 0-87893-580-0
- Ove-Sten Knudsen. เยื่อชีวภาพ: ทฤษฎีการขนส่ง ศักยภาพ และแรงกระตุ้นทางไฟฟ้า . สำนักพิมพ์มหาวิทยาลัยเคมบริดจ์ (26 กันยายน 2545). ISBN 0-521-81018-3ระดับบัณฑิตศึกษา
- ชุดหนังสือเรียนทางการแพทย์ระดับชาติสำหรับการศึกษาด้วยตนเองสรีรวิทยา สำนักพิมพ์ Lippincott Williams & Wilkins ฟิลาเดลเฟีย รัฐเพนซิลเวเนีย สหรัฐอเมริกา ฉบับพิมพ์ครั้งที่ 4 ปี 2001 ISBN 0-683-30603-0
ลิงก์ภายนอก
- หน้าที่ของเยื่อหุ้มเซลล์
- เครื่องจำลองสมการ Nernst/Goldman
- เครื่องคำนวณสมการเนิร์นสต์
- เครื่องคำนวณสมการโกลด์แมน-ฮอดจ์กิน-แคทซ์
- เครื่องคำนวณแรงขับเคลื่อนทางไฟฟ้าเคมี
- ที่มาของศักย์เยื่อหุ้มเซลล์ขณะพัก - บทเรียนแบบโต้ตอบออนไลน์ (Flash)