อ่าน 43 นาที
การจัดลำดับดีเอ็นเอ
การจัดลำดับดีเอ็นเอเป็นกระบวนการกำหนดลำดับกรดนิวคลี อิก – ลำดับของนิวคลีโอไทด์ในดีเอ็นเอซึ่งรวมถึงวิธีการหรือเทคโนโลยีใดๆ ที่ใช้ในการกำหนดลำดับของเบสทั้งสี่
การจัดลำดับดีเอ็นเอ
| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| พันธุศาสตร์ |
|---|
 |
|
การจัดลำดับดีเอ็นเอเป็นกระบวนการกำหนดลำดับกรดนิวคลี อิก – ลำดับของนิวคลีโอไทด์ในดีเอ็นเอซึ่งรวมถึงวิธีการหรือเทคโนโลยีใดๆ ที่ใช้ในการกำหนดลำดับของเบสทั้งสี่ ได้แก่อะดีนีนไทมีนไซโตซีนและกัวนีนการพัฒนาวิธีการจัดลำดับดีเอ็นเอที่รวดเร็วได้เร่งการวิจัยและการค้นพบทางชีววิทยาและการแพทย์อย่างมาก[ 1 ] [ 2 ]
ความรู้เกี่ยวกับลำดับดีเอ็นเอกลายเป็นสิ่งจำเป็นอย่างยิ่งสำหรับการวิจัยทางชีววิทยาขั้นพื้นฐานโครงการจีโนกราฟิกดีเอ็นเอและในสาขาประยุกต์ต่างๆ มากมาย เช่นการวินิจฉัยทางการแพทย์เทคโนโลยีชีวภาพนิติ ชีววิทยา ไวรัสวิทยาและอนุกรมวิธาน ทางชีววิทยา การเปรียบเทียบลำดับดีเอ็นเอที่ปกติและกลายพันธุ์สามารถวินิจฉัยโรคต่างๆ ได้ รวมถึงมะเร็งหลายชนิด[ 3 ]ระบุลักษณะของแอนติบอดี[ 4 ]และสามารถใช้เป็นแนวทางในการรักษาผู้ป่วยได้[ 5 ]การมีวิธีที่รวดเร็วในการจัดลำดับดีเอ็นเอช่วยให้สามารถให้การดูแลทางการแพทย์ที่รวดเร็วและเป็นรายบุคคลมากขึ้น และสามารถระบุและจัดทำรายการสิ่งมีชีวิตได้มากขึ้น[ 4 ]
ความก้าวหน้าอย่างรวดเร็วในเทคโนโลยีการถอดรหัสลำดับดีเอ็นเอมีบทบาทสำคัญอย่างยิ่งในการถอดรหัสจีโนมทั้งหมดของมนุษย์ รวมถึงสัตว์ พืช และจุลินทรีย์อีกหลายชนิด

ลำดับดีเอ็นเอชุดแรกได้มาในช่วงต้นทศวรรษ 1970 โดยนักวิจัยทางวิชาการโดยใช้วิธีการที่ยุ่งยากซึ่งอาศัยโครมาโทกราฟีแบบสองมิติหลังจากการพัฒนาวิธีการจัดลำดับโดยใช้ฟลูออเรสเซนซ์ด้วยเครื่องจัดลำดับดีเอ็นเอ[ 6 ] การจัดลำดับดีเอ็นเอจึงง่ายขึ้นและเร็วขึ้นหลายเท่า[ 7 ] [ 8 ]
แอปพลิเคชัน
การจัดลำดับดีเอ็นเอสามารถใช้เพื่อกำหนดลำดับของยีน แต่ละตัว บริเวณทางพันธุกรรมที่ใหญ่กว่า (เช่น กลุ่มยีนหรือโอเปรอน ) โครโมโซมทั้งหมด หรือจีโนมทั้งหมดของสิ่งมีชีวิตใดๆ การจัดลำดับดีเอ็นเอยังเป็นวิธีที่มีประสิทธิภาพที่สุดในการจัดลำดับอาร์เอ็นเอหรือโปรตีน ทางอ้อม (ผ่านกรอบการอ่านแบบเปิด ) อัน ที่ จริง การจัดลำดับดีเอ็นเอได้กลายเป็นเทคโนโลยีสำคัญในหลายสาขาของชีววิทยาและวิทยาศาสตร์อื่นๆ เช่น การแพทย์นิติวิทยาศาสตร์และมานุษยวิทยา
ชีววิทยาโมเลกุล
การจัดลำดับดีเอ็นเอเป็นเทคนิคที่ใช้ในชีววิทยาระดับโมเลกุลเพื่อศึกษาจีโนมและโปรตีนที่เข้ารหัสโดยจีโนมเหล่านั้น ข้อมูลที่ได้จากการจัดลำดับดีเอ็นเอช่วยให้นักวิจัยสามารถระบุการเปลี่ยนแปลงในยีนและดีเอ็นเอที่ไม่เข้ารหัส (รวมถึงลำดับควบคุม) ความสัมพันธ์กับโรคและลักษณะทางฟีโนไทป์ และระบุเป้าหมายยาที่มีศักยภาพได้
ชีววิทยาเชิงวิวัฒนาการ
เนื่องจาก DNA เป็นโมเลกุลขนาดใหญ่ ที่มีข้อมูล ในแง่ของการส่งต่อจากรุ่นสู่รุ่น การจัดลำดับ DNA จึงถูกนำมาใช้ในชีววิทยาวิวัฒนาการ เพื่อศึกษาความสัมพันธ์ระหว่างสิ่งมีชีวิตต่าง ๆ และวิวัฒนาการของพวกมัน ในเดือนกุมภาพันธ์ พ.ศ. 2564 นักวิทยาศาสตร์ได้รายงานการจัดลำดับ DNAจากซากสัตว์เป็นครั้งแรก ซึ่ง ในกรณีนี้คือ แมมมอธที่มีอายุมากกว่าหนึ่งล้านปี นับเป็น DNA ที่เก่าแก่ที่สุดที่ได้รับการจัดลำดับจนถึงปัจจุบัน[ 9 ] [ 10 ]
เมตาจีโนมิกส์
สาขาเมตาจีโนมิกส์เกี่ยวข้องกับการระบุสิ่งมีชีวิตที่พบในแหล่งน้ำน้ำเสียดิน เศษสิ่งสกปรกที่กรองจากอากาศ หรือตัวอย่างที่เก็บจากสิ่งมีชีวิต การทราบว่ามีสิ่งมีชีวิตชนิดใดอยู่ในสภาพแวดล้อมใดนั้นมีความสำคัญอย่างยิ่งต่อการวิจัยในด้านนิเวศวิทยาระบาดวิทยาจุลชีววิทยาและสาขาอื่นๆ การจัดลำดับดีเอ็นเอช่วยให้นักวิจัยสามารถระบุชนิดของจุลินทรีย์ที่อาจมีอยู่ในไมโครไบโอมได้ เป็นต้น
ไวรัสวิทยา
เนื่องจากไวรัสส่วนใหญ่มีขนาดเล็กเกินกว่าจะมองเห็นได้ด้วยกล้องจุลทรรศน์แบบใช้แสง การจัดลำดับจึงเป็นหนึ่งในเครื่องมือหลักในวิทยาไวรัสเพื่อระบุและศึกษาไวรัส[ 11 ]จีโนมของไวรัสอาจมีพื้นฐานมาจาก DNA หรือ RNA ไวรัส RNA มีความไวต่อเวลามากกว่าสำหรับการจัดลำดับจีโนม เนื่องจากเสื่อมสภาพได้เร็วกว่าในตัวอย่างทางคลินิก[ 12 ]การจัดลำดับแบบ Sangerแบบดั้งเดิมและการจัดลำดับรุ่นใหม่ถูกนำมาใช้ในการจัดลำดับไวรัสในการวิจัยพื้นฐานและทางคลินิก รวมถึงการวินิจฉัยการติดเชื้อไวรัสอุบัติใหม่ระบาดวิทยาโมเลกุลของเชื้อไวรัส และการทดสอบความต้านทานยา มีลำดับไวรัสที่ไม่ซ้ำกันมากกว่า 2.3 ล้านลำดับในGenBank [ 11 ] ในปี 2019 NGS ได้แซงหน้า Sanger แบบดั้งเดิมในฐานะวิธีการที่ได้รับความนิยมมากที่สุดสำหรับการสร้างจีโนมไวรัส[ 11 ]
ในระหว่างการระบาดของไข้หวัดนกในปี 1997การจัดลำดับไวรัสพบว่าสายพันธุ์ย่อยของไข้หวัดใหญ่มีต้นกำเนิดมาจากการผสมผสานระหว่างนกกระทาและสัตว์ปีก ซึ่งนำไปสู่การออกกฎหมายในฮ่องกงที่ห้ามขายนกกระทาและสัตว์ปีกที่มีชีวิตร่วมกันในตลาด การจัดลำดับไวรัสยังสามารถใช้เพื่อประมาณเวลาที่การระบาดของไวรัสเริ่มต้นขึ้นโดยใช้เทคนิคนาฬิกาโมเลกุล ได้อีกด้วย [ 12 ]
ยา
ช่างเทคนิคทางการแพทย์อาจจัดลำดับยีน (หรือในทางทฤษฎีคือจีโนมทั้งหมด) จากผู้ป่วยเพื่อตรวจสอบว่ามีความเสี่ยงต่อโรคทางพันธุกรรมหรือไม่ นี่เป็นรูปแบบหนึ่งของการทดสอบทางพันธุกรรมแม้ว่าการทดสอบทางพันธุกรรมทั้งหมดจะไม่เกี่ยวข้องกับการจัดลำดับดีเอ็นเอจีโนมทั้งหมดก็ตาม[ 13 ]
นับตั้งแต่ปี 2013 การจัดลำดับดีเอ็นเอถูกนำมาใช้มากขึ้นในการวินิจฉัยและรักษาโรคหายาก เมื่อมีการระบุยีนที่ก่อให้เกิดโรคทางพันธุกรรมที่หายากมากขึ้นเรื่อยๆ การวินิจฉัยทางโมเลกุลสำหรับผู้ป่วยจึงกลายเป็นกระแสหลักมากขึ้น การจัดลำดับดีเอ็นเอช่วยให้แพทย์สามารถระบุโรคทางพันธุกรรม ปรับปรุงการจัดการโรค ให้คำปรึกษาด้านการสืบพันธุ์ และให้การรักษาที่มีประสิทธิภาพมากขึ้น[ 14 ]แผงการจัดลำดับยีนใช้เพื่อระบุสาเหตุทางพันธุกรรมที่เป็นไปได้หลายประการของความผิดปกติที่สงสัย[ 15 ]
นอกจากนี้ การจัดลำดับดีเอ็นเออาจมีประโยชน์ในการระบุแบคทีเรียชนิดเฉพาะ เพื่อให้สามารถรักษาด้วยยาปฏิชีวนะได้อย่างแม่นยำ ยิ่งขึ้น ซึ่งจะช่วยลดความเสี่ยงในการเกิดการดื้อยาต้านจุลชีพในประชากรแบคทีเรีย[ 16 ] [ 17 ] [ 18 ] [ 19 ] [ 20 ] [ 21 ]
การสืบสวนทางนิติวิทยาศาสตร์
การจัดลำดับดีเอ็นเออาจใช้ร่วมกับวิธีการสร้างโปรไฟล์ดีเอ็นเอ สำหรับ การระบุตัวตนทางนิติวิทยาศาสตร์[ 22 ]และการทดสอบความเป็นพ่อการทดสอบดีเอ็นเอมีการพัฒนาอย่างมากในช่วงไม่กี่ทศวรรษที่ผ่านมาเพื่อเชื่อมโยงลายนิ้วมือดีเอ็นเอกับสิ่งที่กำลังตรวจสอบ รูปแบบดีเอ็นเอในลายนิ้วมือ น้ำลาย รูขุมขน ฯลฯ แยกสิ่งมีชีวิตแต่ละชนิดออกจากกันอย่างเป็นเอกลักษณ์ การทดสอบดีเอ็นเอเป็นเทคนิคที่สามารถตรวจจับจีโนมเฉพาะในสายดีเอ็นเอเพื่อสร้างรูปแบบเฉพาะบุคคลและเป็นเอกลักษณ์
ฐานหลักทั้งสี่
โครงสร้างมาตรฐานของ DNA มีเบสสี่ชนิด ได้แก่ไทมีน (T), อะดีนีน (A), ไซโตซีน (C) และกัวนีน (G) การจัดลำดับ DNA คือการกำหนดลำดับทางกายภาพของเบสเหล่านี้ในโมเลกุลของ DNA อย่างไรก็ตาม อาจมีเบสอื่นๆ อีกมากมายในโมเลกุล ในไวรัสบางชนิด (โดยเฉพาะแบคทีริโอเฟจ ) ไซโตซีนอาจถูกแทนที่ด้วยไฮดรอกซีเมทิลไซโตซีนหรือไฮดรอกซีเมทิลกลูโคสไซโตซีน[ 23 ]ใน DNA ของสัตว์เลี้ยงลูกด้วยนมอาจพบ เบสที่แตกต่างกันที่มี หมู่เมทิล หรือ ฟอสโฟซัลเฟต[ 24 ] [ 25 ]ขึ้นอยู่กับเทคนิคการจัดลำดับ การดัดแปลงเฉพาะอย่าง เช่น 5mC ( 5-เมทิลไซโตซีน ) ที่พบได้ทั่วไปในมนุษย์ อาจตรวจพบได้หรือไม่ก็ได้[ 26 ]
ในสิ่งมีชีวิตเกือบทั้งหมด DNA จะถูกสังเคราะห์ในร่างกายโดยใช้เบสมาตรฐาน 4 ตัวเท่านั้น การดัดแปลงที่เกิดขึ้นหลังการจำลองแบบจะสร้างเบสอื่นๆ เช่น 5-เมทิล C อย่างไรก็ตาม แบคทีริโอเฟจบางชนิดสามารถรวมเบสที่ไม่เป็นมาตรฐานได้โดยตรง[ 27 ]
นอกจากการปรับเปลี่ยนแล้ว DNA ยังถูกโจมตีอย่างต่อเนื่องจากตัวแทนสิ่งแวดล้อม เช่น รังสี UV และอนุมูลอิสระของออกซิเจน ในปัจจุบัน วิธีการจัดลำดับ DNA ส่วนใหญ่ไม่สามารถตรวจพบการมีอยู่ของเบสที่เสียหายดังกล่าวได้ แม้ว่า PacBio จะได้ตีพิมพ์เรื่องนี้แล้วก็ตาม[ 28 ]
ประวัติศาสตร์
การค้นพบโครงสร้างและหน้าที่ของดีเอ็นเอ
กรดดีออกซีไรโบ nucléique ( DNA ) ถูกค้นพบและแยกออกมาเป็นครั้งแรกโดยฟรีดริช มีเชอร์ในปี 1869 แต่กลับไม่ได้รับการศึกษาอย่างเพียงพอเป็นเวลาหลายทศวรรษ เนื่องจาก เชื่อกันว่า โปรตีนต่างหากที่เป็นตัวนำพาพิมพ์เขียวทางพันธุกรรมของสิ่งมีชีวิต ไม่ใช่ DNA สถานการณ์นี้เปลี่ยนไปหลังจากปี 1944 อันเป็นผลมาจากการทดลองของออสวาลด์ เอเวอรี , โคลิน แมคลีโอและแมคลิน แมคคาร์ตีที่แสดงให้เห็นว่า DNA ที่บริสุทธิ์สามารถเปลี่ยนสายพันธุ์ของแบคทีเรียสายพันธุ์หนึ่งไปเป็นอีกสายพันธุ์หนึ่งได้ นี่เป็นครั้งแรกที่มีการแสดงให้เห็นว่า DNA สามารถเปลี่ยนแปลงคุณสมบัติของเซลล์ได้
ในปี พ.ศ. 2496 เจมส์ วัตสันและฟรานซิส คริกได้เสนอ แบบ จำลองเกลียวคู่ของดีเอ็นเอ โดยอิงจาก โครงสร้าง ผลึกเอ็กซ์เรย์ที่โรซาลินด์ แฟรงคลินกำลังศึกษาตามแบบจำลองนี้ ดีเอ็นเอประกอบด้วยนิวคลีโอไทด์สองสายที่ขดตัวรอบกัน โดยเชื่อมต่อกันด้วยพันธะไฮโดรเจนและวิ่งไปในทิศทางตรงกันข้าม แต่ละสายประกอบด้วยนิวคลีโอไทด์ที่เสริมกันสี่ตัว ได้แก่ อะดีนีน (A) ไซโตซีน (C) กัวนีน (G) และไทมีน (T) โดย A บนสายหนึ่งจะจับคู่กับ T บนอีกสายหนึ่งเสมอ และ C จะจับคู่กับ G เสมอ พวกเขาเสนอว่าโครงสร้างดังกล่าวทำให้แต่ละสายสามารถใช้สร้างอีกสายหนึ่งขึ้นใหม่ได้ ซึ่งเป็นแนวคิดสำคัญในการส่งต่อข้อมูลทางพันธุกรรมระหว่างรุ่น[ 29 ]

รากฐานสำหรับการจัดลำดับโปรตีนนั้นวางโดยงานของFrederick Sangerซึ่งในปี 1955 เขาได้จัดลำดับกรดอะมิโนทั้งหมดในอินซูลินซึ่งเป็นโปรตีนขนาดเล็กที่หลั่งจากตับอ่อน นี่เป็นหลักฐานที่ชัดเจนครั้งแรกว่าโปรตีนเป็นสารเคมีที่มีรูปแบบโมเลกุลที่เฉพาะเจาะจง ไม่ใช่ส่วนผสมแบบสุ่มของวัสดุที่แขวนลอยอยู่ในของเหลว ความสำเร็จของ Sanger ในการจัดลำดับอินซูลินกระตุ้นให้นักผลึกศาสตร์รังสีเอกซ์ รวมถึง Watson และ Crick ซึ่งในขณะนั้นกำลังพยายามทำความเข้าใจว่า DNA ควบคุมการสร้างโปรตีนภายในเซลล์ได้อย่างไร ไม่นานหลังจากเข้าร่วมการบรรยายชุดหนึ่งที่ Frederick Sanger บรรยายในเดือนตุลาคม 1954 Crick เริ่มพัฒนาทฤษฎีที่กล่าวว่าการจัดเรียงนิวคลีโอไทด์ใน DNA กำหนดลำดับของกรดอะมิโนในโปรตีน ซึ่งในทางกลับกันช่วยกำหนดหน้าที่ของโปรตีน เขาตีพิมพ์ทฤษฎีนี้ในปี 1958 [ 30 ]
การจัดลำดับอาร์เอ็นเอ
การจัดลำดับ RNAเป็นหนึ่งในรูปแบบแรกสุดของการจัดลำดับนิวคลีโอไทด์ ความสำเร็จที่สำคัญของการจัดลำดับ RNA คือการจัดลำดับยีนที่สมบูรณ์ตัวแรกและจีโนมที่สมบูรณ์ของแบคทีริโอเฟจ MS2ซึ่งระบุและตีพิมพ์โดยWalter Fiersและเพื่อนร่วมงานของเขาที่มหาวิทยาลัย Ghent ( Ghentประเทศเบลเยียม ) ในปี 1972 [ 31 ]และ 1976 [ 32 ]วิธีการจัดลำดับ RNA แบบดั้งเดิมจำเป็นต้องสร้าง โมเลกุล cDNAซึ่งจะต้องได้รับการจัดลำดับ[ 33 ]
วิธีการลำดับดีเอ็นเอในยุคแรก
วิธีแรกในการกำหนดลำดับดีเอ็นเอเกี่ยวข้องกับกลยุทธ์การขยายไพรเมอร์เฉพาะตำแหน่งที่คิดค้นโดยRay Wuนักพันธุศาสตร์ที่มหาวิทยาลัยคอร์เนลล์ในปี 1970 [ 34 ]การเร่งปฏิกิริยาของดีเอ็นเอพอลิเมอเรสและการติดฉลากนิวคลีโอไทด์เฉพาะ ซึ่งทั้งสองอย่างนี้มีบทบาทสำคัญในแผนการจัดลำดับในปัจจุบัน ถูกนำมาใช้ในการจัดลำดับปลายที่เชื่อมต่อกันของดีเอ็นเอ ของ แลมบ์ดาฟาจ[ 35 ] [ 36 ] [ 37 ]ระหว่างปี 1970 ถึง 1973 Wu นักวิทยาศาสตร์ Radha Padmanabhan และเพื่อนร่วมงานได้แสดงให้เห็นว่าวิธีการนี้สามารถนำมาใช้ในการกำหนดลำดับดีเอ็นเอใดๆ ก็ได้โดยใช้ไพรเมอร์เฉพาะตำแหน่งสังเคราะห์[ 38 ] [ 39 ] [ 8 ]
วอลเตอร์ กิลเบิร์ตนักชีวเคมี และอัลลัน แม็กซัมนักพันธุศาสตร์โมเลกุล ที่มหาวิทยาลัยฮาร์ วาร์ด ได้พัฒนาวิธีการจัดลำดับดีเอ็นเอ รวมถึงวิธีการ "จัดลำดับดีเอ็นเอโดยการย่อยสลายทางเคมี" [ 40 ] [ 41 ]ในปี พ.ศ. 2516 กิลเบิร์ตและแม็กซัมได้รายงานลำดับเบส 24 คู่โดยใช้วิธีที่เรียกว่าการวิเคราะห์จุดเคลื่อนที่[ 42 ]ความก้าวหน้าในการจัดลำดับได้รับการสนับสนุนจากการพัฒนา เทคโนโลยี ดีเอ็นเอลูกผสมควบคู่ กันไป ทำให้สามารถแยกตัวอย่างดีเอ็นเอจากแหล่งอื่นที่ไม่ใช่ไวรัสได้[ 43 ]
สองปีต่อมาในปี 1975 เฟรเดอริค แซงเกอร์นักชีวเคมี และอลัน คูลสันนักวิทยาศาสตร์จีโนม ได้พัฒนาวิธีการจัดลำดับดีเอ็นเอ[ 44 ]เทคนิคนี้เรียกว่าวิธี "บวกและลบ" ซึ่งเกี่ยวข้องกับการจัดหาส่วนประกอบทั้งหมดของดีเอ็นเอ แต่ไม่รวมปฏิกิริยาของเบสหนึ่งในสี่ตัวที่จำเป็นในการสร้างดีเอ็นเอให้สมบูรณ์[ 45 ]
ในปี พ.ศ. 2519 กิลเบิร์ตและแม็กซัมได้คิดค้นวิธีการจัดลำดับดีเอ็นเออย่างรวดเร็วขณะอยู่ที่มหาวิทยาลัยฮาร์วาร์ด ซึ่งรู้จักกันในชื่อการจัดลำดับแบบแม็กซัม-กิลเบิร์ต[ 46 ]เทคนิคนี้เกี่ยวข้องกับการบำบัดดีเอ็นเอที่ติดฉลากด้วยรังสีด้วยสารเคมีและใช้เจลโพลีอะคริลาไมด์เพื่อกำหนดลำดับ[ 47 ]
ในปี พ.ศ. 2520 Sanger ได้นำกลยุทธ์การขยายไพรเมอร์มาใช้เพื่อพัฒนาวิธีการจัดลำดับดีเอ็นเอที่รวดเร็วยิ่งขึ้นที่ศูนย์ MRCเมืองเคมบริดจ์สหราชอาณาจักร เทคนิคนี้คล้ายกับกลยุทธ์ "บวกและลบ" ของเขา อย่างไรก็ตาม เทคนิคนี้อาศัยการรวมไดดีออกซีนิวคลีโอไทด์ (ddNTPs) ที่ยุติสายโซ่แบบเลือกโดยดีเอ็นเอพอลิเมอเรสใน ระหว่าง การจำลองดีเอ็นเอในหลอดทดลอง[ 48 ] [ 47 ] [ 49 ] Sanger ได้ตีพิมพ์วิธีการนี้ในปีเดียวกัน[ 50 ]
การจัดลำดับจีโนมทั้งหมด

จีโนม DNA ที่สมบูรณ์ชุดแรกที่ได้รับการจัดลำดับคือของแบคทีริโอเฟจ φX174ในปี 1977 [ 51 ] นักวิทยาศาสตร์ ของสภาวิจัยทางการแพทย์ถอดรหัสลำดับ DNA ที่สมบูรณ์ของไวรัส Epstein-Barrในปี 1984 พบว่ามีนิวคลีโอไทด์ 172,282 ตัว การจัดลำดับที่สมบูรณ์นี้ถือเป็นจุดเปลี่ยนที่สำคัญในการจัดลำดับ DNA เนื่องจากทำได้โดยไม่มีความรู้เกี่ยวกับโปรไฟล์ทางพันธุกรรมของไวรัสมาก่อน[ 52 ] [ 8 ]
เฮอร์เบิร์ต โพห์ลและเพื่อนร่วมงานได้พัฒนาวิธีการที่ไม่ใช้สารกัมมันตรังสีสำหรับการถ่ายโอนโมเลกุล DNA ของส่วนผสมปฏิกิริยาการจัดลำดับไปยังเมทริกซ์ตรึงระหว่างอิเล็กโทรโฟเร ซิสในช่วงต้นทศวรรษ 1980 [ 53 ] [ 54 ]ตามมาด้วยการวางจำหน่ายเครื่องจัดลำดับ DNA "Direct-Blotting-Electrophoresis-System GATC 1500" โดยGATC Biotechซึ่งถูกนำมาใช้อย่างเข้มข้นในกรอบของโครงการจัดลำดับจีโนมของสหภาพยุโรป ส่งผลให้ได้ลำดับ DNA ที่สมบูรณ์ของโครโมโซม II ของยีสต์ Saccharomyces cerevisiae [ 55 ] ห้องปฏิบัติการของLeroy E. Hood ที่ สถาบันเทคโนโลยีแคลิฟอร์เนียประกาศเปิดตัวเครื่องจัดลำดับดีเอ็นเอแบบกึ่งอัตโนมัติเครื่องแรกในปี 1986 [ 56 ]ตามมาด้วยApplied Biosystemsที่วางจำหน่ายเครื่องจัดลำดับแบบอัตโนมัติเต็มรูปแบบเครื่องแรก คือ ABI 370 ในปี 1987 และ Genesis 2000 ของ Dupont [ 57 ]ซึ่งใช้เทคนิคการติดฉลากด้วยฟลูออเรสเซนต์แบบใหม่ ทำให้สามารถระบุได ดีออก ซีนิวคลีโอไทด์ ทั้งสี่ชนิดได้ในเลนเดียว ภายในปี 1990 สถาบันสุขภาพแห่งชาติ ของสหรัฐอเมริกา (NIH) ได้เริ่มการทดลองจัดลำดับขนาดใหญ่กับMycoplasma capricolum , Escherichia coli , Caenorhabditis elegansและSaccharomyces cerevisiaeในราคา 0.75 ดอลลาร์สหรัฐต่อเบส ในขณะเดียวกัน การจัด ลำดับลำดับcDNAของมนุษย์ ที่เรียกว่า แท็กแสดงลำดับเริ่มต้นขึ้นในห้องปฏิบัติการของCraig Venter ซึ่งเป็นความพยายามที่จะจับส่วนที่เข้ารหัสของจีโน มมนุษย์[ 58 ]ในปี 1995 Venter, Hamilton Smithและเพื่อนร่วมงานที่สถาบันวิจัยจีโนม (TIGR) ได้ตีพิมพ์จีโนมที่สมบูรณ์ครั้งแรกของสิ่งมีชีวิตอิสระ คือแบคทีเรียHaemophilus influenzaeโครโมโซมวงกลมประกอบด้วยเบส 1,830,137 เบส และการตีพิมพ์ในวารสาร Science [ 59 ]ถือเป็นการใช้การจัดลำดับจีโนมแบบช็อตกันครั้งแรกที่ได้รับการตีพิมพ์ ซึ่งช่วยลดความจำเป็นในการทำแผนที่เบื้องต้น
ในปี 2003 วิธีการจัดลำดับแบบช็อตกันของโครงการจีโนมมนุษย์ได้ถูกนำมาใช้เพื่อสร้างลำดับร่างของจีโนมมนุษย์ ซึ่งมีความแม่นยำ 92% [ 60 ] [ 61 ] [ 62 ]ในปี 2022 นักวิทยาศาสตร์ประสบความสำเร็จในการจัดลำดับจีโนมมนุษย์ส่วนที่เหลืออีก 8% ยีนอ้างอิงมาตรฐานที่จัดลำดับอย่างสมบูรณ์เรียกว่า GRCh38.p14 และประกอบด้วยเบสคู่ 3.1 พันล้านคู่[ 62 ] [ 63 ]
วิธีการลำดับดีเอ็นเอความเร็วสูง (HTS)

ในช่วงกลางถึงปลายทศวรรษ 1990 มีการพัฒนาวิธีการจัดลำดับดีเอ็นเอแบบใหม่หลายวิธี และนำไปใช้ในเครื่องจัดลำดับดีเอ็นเอ เชิงพาณิชย์ ภายในปี 2000 วิธีการเหล่านี้เรียกรวมกันว่า วิธีการจัดลำดับดีเอ็นเอ "รุ่นต่อไป" หรือ "รุ่นที่สอง" (NGS) เพื่อแยกแยะออกจากวิธีการก่อนหน้านี้ รวมถึงการจัดลำดับแบบแซงเกอร์แตกต่างจากการจัดลำดับรุ่นแรก เทคโนโลยี NGS มีลักษณะเด่นคือ สามารถปรับขนาดได้สูง ทำให้สามารถจัดลำดับจีโนมทั้งหมดได้ในคราวเดียว โดยปกติแล้วจะทำได้โดยการแบ่งจีโนมออกเป็นชิ้นเล็กๆ สุ่มเลือกชิ้นส่วน และจัดลำดับโดยใช้เทคโนโลยีต่างๆ เช่นที่อธิบายไว้ด้านล่าง การจัดลำดับจีโนมทั้งหมดเป็นไปได้เพราะสามารถจัดลำดับชิ้นส่วนหลายชิ้นพร้อมกัน (จึงเรียกว่า การจัดลำดับแบบ "ขนานจำนวนมาก") ในกระบวนการอัตโนมัติ
เทคโนโลยี NGS ได้เพิ่มศักยภาพอย่างมหาศาลให้แก่นักวิจัยในการค้นหาข้อมูลเชิงลึกเกี่ยวกับสุขภาพ นักมานุษยวิทยาในการตรวจสอบต้นกำเนิดของมนุษย์ และเป็นตัวเร่งให้เกิดการเคลื่อนไหว " การแพทย์เฉพาะบุคคล " อย่างไรก็ตาม เทคโนโลยีนี้ยังเปิดโอกาสให้เกิดข้อผิดพลาดมากขึ้นด้วย มีเครื่องมือซอฟต์แวร์มากมายที่ใช้ในการวิเคราะห์ข้อมูล NGS ด้วยวิธีการคำนวณ ซึ่งมักจะรวบรวมไว้ในแพลตฟอร์มออนไลน์ เช่น CSI NGS Portal โดยแต่ละเครื่องมือมีอัลกอริทึมของตนเอง แม้แต่พารามิเตอร์ภายในแพ็กเกจซอฟต์แวร์เดียวกันก็สามารถเปลี่ยนแปลงผลลัพธ์ของการวิเคราะห์ได้ นอกจากนี้ ปริมาณข้อมูลจำนวนมากที่ได้จากการจัดลำดับดีเอ็นเอยังต้องการการพัฒนาวิธีการและโปรแกรมใหม่สำหรับการวิเคราะห์ลำดับอีกด้วย มีความพยายามหลายครั้งในการพัฒนามาตรฐานในสาขา NGS เพื่อแก้ไขปัญหาเหล่านี้ ซึ่งส่วนใหญ่เป็นความพยายามขนาดเล็กที่เกิดขึ้นจากห้องปฏิบัติการแต่ละแห่ง เมื่อไม่นานมานี้ ความพยายามขนาดใหญ่ที่เป็นระบบและได้รับการสนับสนุนจาก FDA ได้นำไปสู่มาตรฐานBioCompute [ 65 ]
เมื่อวันที่ 26 ตุลาคม พ.ศ. 2533 Roger Tsien , Pepi Ross, Margaret Fahnestock และ Allan J Johnston ได้ยื่นจดสิทธิบัตรที่อธิบายถึงการจัดลำดับแบบทีละเบส ("เบสต่อเบส") โดยใช้บล็อกเกอร์ 3' ที่ถอดออกได้บนอาร์เรย์ DNA (แผ่นและโมเลกุล DNA เดี่ยว) [ 66 ] ในปี พ.ศ. 2539 Pål NyrénและMostafa Ronaghi นักศึกษาของเขา ที่สถาบันเทคโนโลยีแห่งราชวงศ์ในสตอกโฮล์มได้ตีพิมพ์วิธีการไพโรซีเควนซิงของ พวกเขา [ 67 ]
เมื่อวันที่ 1 เมษายน พ.ศ. 2540 Pascal Mayerและ Laurent Farinelli ได้ยื่นจดสิทธิบัตรที่อธิบายถึงการจัดลำดับโคโลนี DNA [ 68 ]วิธีการเตรียมตัวอย่าง DNA และ วิธีการจัดเรียง ปฏิกิริยาลูกโซ่พอลิเมอเรส (PCR) บนพื้นผิวแบบสุ่มที่อธิบายไว้ในสิทธิบัตรนี้ ควบคู่กับวิธีการจัดลำดับแบบ "เบสต่อเบส" ของ Roger Tsien และคณะ ได้ถูกนำไปใช้ในเครื่องจัดลำดับจีโนม Hi-Seq ของ Illumina แล้ว
ในปี พ.ศ. 2541 Phil Green และ Brent Ewing จากมหาวิทยาลัยวอชิงตันได้อธิบายคะแนนคุณภาพ phred ของพวกเขา สำหรับการวิเคราะห์ข้อมูลซีเควนเซอร์[ 69 ]ซึ่งเป็นเทคนิคการวิเคราะห์ที่สำคัญที่ได้รับการยอมรับอย่างกว้างขวาง และยังคงเป็นตัวชี้วัดที่ใช้กันทั่วไปที่สุดสำหรับการประเมินความแม่นยำของแพลตฟอร์มการจัดลำดับ[ 70 ]
Lynx Therapeutics ได้เผยแพร่และทำการตลาดการจัดลำดับลายเซ็นแบบขนานขนาดใหญ่ (MPSS) ในปี 2000 วิธีนี้รวมเอาเทคโนโลยีการจัดลำดับแบบขนานโดยใช้อะแดปเตอร์/การเชื่อมต่อแบบใช้ลูกปัด และถือเป็นวิธีการจัดลำดับ "รุ่นต่อไป" ที่วางจำหน่ายในเชิงพาณิชย์เป็นครั้งแรก แม้ว่าจะไม่มี การขาย เครื่องจัดลำดับ DNAให้กับห้องปฏิบัติการอิสระก็ตาม[ 71 ]
วิธีการพื้นฐาน
การจัดลำดับแบบ Maxam-Gilbert
Allan MaxamและWalter Gilbertได้ตีพิมพ์วิธีการจัดลำดับดีเอ็นเอในปี 1977 โดยอาศัยการดัดแปลงทางเคมีของดีเอ็นเอและการตัดที่เบสเฉพาะ[ 40 ]วิธีนี้เรียกอีกอย่างว่าการจัดลำดับทางเคมี ซึ่งช่วยให้สามารถใช้ตัวอย่างดีเอ็นเอสองสายที่บริสุทธิ์ได้โดยไม่ต้องทำการโคลนนิ่งเพิ่มเติม การใช้การติดฉลากด้วยสารกัมมันตรังสีและความซับซ้อนทางเทคนิคของวิธีนี้ทำให้การใช้งานอย่างแพร่หลายเป็นไปได้ยากหลังจากมีการปรับปรุงวิธีการของ Sanger แล้ว
การจัดลำดับแบบ Maxam-Gilbert ต้องใช้การติดฉลากกัมมันตรังสีที่ปลาย 5' ด้านหนึ่งของ DNA และการทำให้บริสุทธิ์ของชิ้นส่วน DNA ที่จะจัดลำดับ จากนั้นการบำบัดทางเคมีจะสร้างรอยแตกที่สัดส่วนเล็กน้อยของเบสนิวคลีโอไทด์หนึ่งหรือสองตัวจากสี่ตัวในแต่ละปฏิกิริยาทั้งสี่ (G, A+G, C, C+T) ความเข้มข้นของสารเคมีที่ปรับเปลี่ยนจะถูกควบคุมเพื่อให้เกิดการปรับเปลี่ยนโดยเฉลี่ยหนึ่งครั้งต่อโมเลกุล DNA ดังนั้นจึงมีการสร้างชิ้นส่วนที่ติดฉลากเป็นชุดๆ จากปลายที่ติดฉลากกัมมันตรังสีไปยังตำแหน่ง "ตัด" แรกในแต่ละโมเลกุล ชิ้นส่วนในปฏิกิริยาทั้งสี่จะถูกแยกด้วยไฟฟ้าแบบเคียงข้างกันใน เจลอะคริลาไมด์แบบ ดีเนเจอร์ริ่ ง เพื่อแยกขนาด เพื่อให้เห็นภาพชิ้นส่วน เจลจะถูกนำไปฉายรังสีเอกซ์บนฟิล์มสำหรับการถ่ายภาพรังสีอัตโนมัติ ซึ่งจะได้แถบสีเข้มเป็นชุดๆ โดยแต่ละแถบจะสอดคล้องกับชิ้นส่วน DNA ที่ติดฉลากกัมมันตรังสี ซึ่งสามารถอนุมานลำดับได้จากแถบเหล่านี้[ 40 ]
วิธีการนี้ล้าสมัยไปแล้วเป็นส่วนใหญ่ตั้งแต่ปี 2023 [ 72 ]
วิธีการยุติลูกโซ่
วิธีการยุติสายโซ่ที่พัฒนาโดยFrederick Sangerและเพื่อนร่วมงานในปี 1977 ในไม่ช้าก็กลายเป็นวิธีการที่ได้รับเลือก เนื่องจากมีความง่ายและน่าเชื่อถือ[ 50 ] [ 73 ]เมื่อคิดค้นขึ้น วิธีการยุติสายโซ่ใช้สารเคมีที่เป็นพิษน้อยกว่าและมีปริมาณกัมมันตภาพรังสีน้อยกว่าวิธีการของ Maxam และ Gilbert เนื่องจากความง่ายในการเปรียบเทียบ วิธีการของ Sanger จึงถูกทำให้เป็นระบบอัตโนมัติในไม่ช้าและเป็นวิธีการที่ใช้ในเครื่องจัดลำดับดีเอ็นเอ รุ่น แรก
การจัดลำดับแบบแซงเกอร์เป็นวิธีการที่แพร่หลายตั้งแต่ทศวรรษ 1980 จนถึงกลางทศวรรษ 2000 ในช่วงเวลานั้น มีความก้าวหน้าอย่างมากในเทคนิคนี้ เช่น การติดฉลากด้วยฟลูออเรสเซนต์ อิเล็กโทรโฟเรซิสแบบเส้นเลือดฝอย และระบบอัตโนมัติทั่วไป การพัฒนาเหล่านี้ทำให้การจัดลำดับมีประสิทธิภาพมากขึ้น ส่งผลให้ต้นทุนลดลง วิธีการแซงเกอร์ในรูปแบบการผลิตจำนวนมากเป็นเทคโนโลยีที่สร้างจีโนมมนุษย์ตัวแรกในปี 2001 ซึ่งเป็นการเริ่มต้นยุคของจีโนมิกส์อย่างไรก็ตาม ในช่วงปลายทศวรรษนั้น วิธีการที่แตกต่างอย่างสิ้นเชิงได้เข้าสู่ตลาด ทำให้ต้นทุนต่อจีโนมลดลงจาก 100 ล้านดอลลาร์ในปี 2001 เหลือ 10,000 ดอลลาร์ในปี 2011 [ 74 ]
การจัดลำดับโดยการสังเคราะห์
วัตถุประสงค์ของการจัดลำดับแบบต่อเนื่องโดยการสังเคราะห์ (SBS) คือการกำหนดลำดับของ ตัวอย่าง DNAโดยการตรวจจับการรวมตัวของนิวคลีโอไทด์โดยDNA โพลีเมอเรส มีการใช้โพลีเมอเรสที่ได้รับการดัดแปลงทางวิศวกรรมเพื่อสังเคราะห์สำเนาของสาย DNA เดี่ยว และการรวมตัวของนิวคลีโอไทด์แต่ละตัวจะถูกตรวจสอบ หลักการของการจัดลำดับแบบเรียลไทม์โดยการสังเคราะห์ได้รับการอธิบายครั้งแรกในปี 1993 [ 75 ]โดยมีการปรับปรุงตีพิมพ์ในอีกหลายปีต่อมา[ 76 ]ส่วนประกอบหลักมีความคล้ายคลึงกันอย่างมากสำหรับ SBS ทุกรูปแบบ และประกอบด้วย (1) การขยาย DNA (เพื่อเพิ่มสัญญาณในภายหลัง) และการติด DNA ที่จะทำการเรียงลำดับเข้ากับตัวรองรับของแข็ง (ยกเว้นPacBio SMRT ) (2) การสร้าง DNA สายเดี่ยวบนตัวรองรับของแข็ง (3) การรวมนิวคลีโอไทด์โดยใช้พอลิเมอเรสที่ได้รับการดัดแปลง และ (4) การตรวจจับการรวมนิวคลีโอไทด์แบบเรียลไทม์ ขั้นตอนที่ 3-4 จะถูกทำซ้ำ และลำดับจะถูกประกอบขึ้นจากสัญญาณที่ได้รับในขั้นตอนที่ 4 หลักการของการเรียงลำดับแบบเรียลไทม์โดยการสังเคราะห์นี้ถูกนำมาใช้กับเครื่องมือเรียงลำดับแบบขนานขนาดใหญ่เกือบทั้งหมดรวมถึง454 , PacBio , IonTorrent , IlluminaและMGI [ 77 ]
การจัดลำดับโดยการขยายตัว
ในเดือนกุมภาพันธ์ พ.ศ. 2568 Rocheประกาศการพัฒนาวิธีการจัดลำดับแบบใหม่ที่เป็นกรรมสิทธิ์โดยอาศัยโมเลกุลขนาดเล็ก วิธีนี้ "เข้ารหัสลำดับของโมเลกุลกรดนิวคลีอิกเป้าหมาย (DNA หรือ RNA) ลงในพอลิเมอร์ทดแทนที่วัดได้ซึ่งเรียกว่า Xpandomer" และรวมการขยายเข้ากับการตรวจจับในภายหลังโดยแพลตฟอร์มนาโนพอเรที่มีกล้องCMOS [ 78 ] [ 79 ]
การจัดลำดับดีเอ็นเอขนาดใหญ่และการจัดลำดับดีเอ็นเอแบบ de novo

การจัดลำดับดีเอ็นเอขนาดใหญ่ มักมีเป้าหมายในการจัดลำดับชิ้นส่วนดีเอ็นเอที่มีความยาวมาก เช่นโครโมโซม ทั้งหมด แม้ว่าการจัดลำดับดีเอ็นเอขนาดใหญ่จะสามารถใช้สร้างลำดับสั้นๆ จำนวนมากได้เช่นกัน เช่นที่พบในเทคนิคฟาจดิสเพลย์สำหรับเป้าหมายที่มีความยาว เช่น โครโมโซม วิธีการทั่วไปประกอบด้วยการตัด (ด้วยเอนไซม์ตัดจำเพาะ ) หรือการเฉือน (ด้วยแรงทางกล) ชิ้นส่วนดีเอ็นเอขนาดใหญ่ให้เป็นชิ้นส่วนดีเอ็นเอที่สั้นลง จากนั้นดีเอ็นเอที่ถูกตัดแล้วอาจถูกโคลนลงในเวกเตอร์ดีเอ็นเอและเพิ่มจำนวนในโฮสต์แบคทีเรีย เช่นEscherichia coliชิ้นส่วนดีเอ็นเอสั้นๆ ที่บริสุทธิ์จากโคโลนีแบคทีเรียแต่ละตัวจะถูกจัดลำดับทีละชิ้นและประกอบเข้าด้วยกันทางอิเล็กทรอนิกส์เป็นลำดับยาวต่อเนื่องหนึ่งลำดับ การศึกษาแสดงให้เห็นว่าการเพิ่มขั้นตอนการคัดเลือกขนาดเพื่อรวบรวมชิ้นส่วนดีเอ็นเอที่มีขนาดสม่ำเสมอสามารถปรับปรุงประสิทธิภาพการจัดลำดับและความแม่นยำของการประกอบจีโนมได้ ในการศึกษาเหล่านี้ การกำหนดขนาดอัตโนมัติได้รับการพิสูจน์แล้วว่ามีความน่าเชื่อถือและแม่นยำกว่าการกำหนดขนาดเจลด้วยตนเอง[ 80 ] [ 81 ] [ 82 ]
คำว่า " การจัดลำดับดีเอ็นเอ แบบ de novo " หมายถึงวิธีการที่ใช้ในการกำหนดลำดับของดีเอ็นเอที่ไม่เคยมีลำดับที่ทราบมาก่อนde novoมาจากภาษาละติน แปลว่า "จากจุดเริ่มต้น" ช่องว่างในลำดับที่ประกอบขึ้นอาจถูกเติมเต็มโดยวิธีการเดินไพรเมอร์ (primer walking ) กลยุทธ์ที่แตกต่างกันมีข้อดีข้อเสียที่แตกต่างกันในด้านความเร็วและความแม่นยำวิธีการแบบ shotgunมักใช้สำหรับการจัดลำดับจีโนมขนาดใหญ่ แต่การประกอบลำดับนั้นซับซ้อนและยาก โดยเฉพาะอย่างยิ่งกับลำดับซ้ำที่มักทำให้เกิดช่องว่างในการประกอบจีโนม
วิธีการจัดลำดับส่วนใหญ่ใช้ วิธีการโคลนนิ่ง ในหลอดทดลองเพื่อขยายโมเลกุล DNA แต่ละตัว เนื่องจากวิธีการตรวจจับโมเลกุลไม่ไวพอสำหรับการจัดลำดับโมเลกุลเดี่ยว Emulsion PCR [ 83 ]แยกโมเลกุล DNA แต่ละตัวพร้อมกับลูกปัดที่เคลือบด้วยไพรเมอร์ในหยดน้ำภายในเฟสน้ำมัน จากนั้น ปฏิกิริยาลูกโซ่พอลิเมอเรส (PCR) จะเคลือบลูกปัดแต่ละเม็ดด้วยสำเนาโคลนของโมเลกุล DNA ตามด้วยการตรึงเพื่อการจัดลำดับในภายหลัง Emulsion PCR ถูกใช้ในวิธีการที่พัฒนาโดย Marguilis et al. (จำหน่ายโดย454 Life Sciences ), Shendure และ Porreca et al. (หรือที่รู้จักกันในชื่อ " polony sequencing ") และSOLiD sequencing (พัฒนาโดยAgencourtต่อมาคือApplied BiosystemsปัจจุบันคือLife Technologies ) [ 84 ] [ 85 ] [ 86 ] Emulsion PCR ยังใช้ในแพลตฟอร์ม GemCode และ Chromium ที่พัฒนาโดย10x Genomicsอีก ด้วย [ 87 ]
ลำดับปืนลูกซอง
การจัดลำดับแบบช็อตกัน (Shotgun sequencing) เป็นวิธีการจัดลำดับที่ออกแบบมาสำหรับการวิเคราะห์ลำดับ DNA ที่ยาวกว่า 1,000 คู่เบส ไปจนถึงโครโมโซมทั้งหมด วิธีนี้ต้องการให้ DNA เป้าหมายถูกแบ่งออกเป็นชิ้นส่วนแบบสุ่ม หลังจากจัดลำดับชิ้นส่วนแต่ละชิ้นโดยใช้วิธีการยุติสายโซ่แล้ว ลำดับเหล่านั้นสามารถประกอบขึ้นใหม่ได้โดยอาศัยบริเวณที่ทับซ้อนกัน[ 88 ]วิธีนี้ถูกนำมาใช้อย่างแพร่หลายใน โครงการจีโน มมนุษย์[ 89 ]
วิธีการที่มีประสิทธิภาพสูง

การจัดลำดับแบบความเร็วสูง ซึ่งรวมถึงวิธีการจัดลำดับแบบ "อ่านสั้น" รุ่นต่อไปและแบบ "อ่านยาว" รุ่นที่สาม[ nt 1 ]ใช้กับการจัดลำดับเอ็กโซมการจัดลำดับจีโนม การจัดลำดับจีโนมใหม่ การวิเคราะห์ โปรไฟล์ทรานสคริปโตม ( RNA-Seq ) ปฏิสัมพันธ์ระหว่าง DNA กับโปรตีน ( ChIP-sequencing ) และลักษณะเฉพาะของเอพิเจโนม[ 90 ]
ความต้องการการจัดลำดับดีเอ็นเอต้นทุนต่ำที่สูงได้ผลักดันการพัฒนาเทคโนโลยีการจัดลำดับดีเอ็นเอความเร็วสูง ซึ่ง ทำให้ กระบวนการจัดลำดับเป็นแบบขนาน และสามารถสร้างลำดับได้หลายพันหรือหลายล้านลำดับพร้อมกัน [ 91 ] [ 92 ] [ 93 ]เทคโนโลยีการจัดลำดับดีเอ็นเอความเร็วสูงมีจุดประสงค์เพื่อลดต้นทุนการจัดลำดับดีเอ็นเอให้ต่ำกว่าที่ทำได้ด้วยวิธีการย้อมสีแบบมาตรฐาน[ 94 ]ในการจัดลำดับดีเอ็นเอความเร็วสูงมาก สามารถดำเนินการจัดลำดับโดยการสังเคราะห์ได้มากถึง 500,000 ครั้งพร้อมกัน[ 95 ] [ 96 ] [ 97 ]เทคโนโลยีดังกล่าวทำให้สามารถจัดลำดับจีโนมมนุษย์ทั้งหมดได้ภายในเวลาเพียงหนึ่งวัน[ 98 ] ณ ปี 2019 บริษัทชั้นนำในการพัฒนาผลิตภัณฑ์การจัดลำดับดีเอ็นเอความเร็วสูง ได้แก่Illumina , QiagenและThermoFisher Scientific [ 98 ]
| วิธี | ความยาวในการอ่าน | ความแม่นยำ (การอ่านครั้งเดียว ไม่ใช่ฉันทามติ) | จำนวนการอ่านต่อการทำงาน | เวลาต่อรอบ | ต้นทุนต่อ 1 พันล้านฐาน (ในสกุลเงินดอลลาร์สหรัฐ) | ข้อดี | ข้อเสีย |
|---|---|---|---|---|---|---|---|
| การลำดับดีเอ็นเอแบบเรียลไทม์ระดับโมเลกุลเดี่ยว (Pacific Biosciences) | 30,000 bp ( N50 ); | ความแม่นยำในการอ่านดิบ 87% [ 104 ] | 4,000,000 ต่อเซลล์ SMRT ของ Sequel 2, 100–200 กิกะเบส[ 101 ] [ 105 ] [ 106 ] | 30 นาทีถึง 20 ชั่วโมง[ 101 ] [ 107 ] | 7.2-43.3 ดอลลาร์สหรัฐ | รวดเร็ว ตรวจจับ 4mC, 5mC, 6mA [ 108 ] | อัตราการผลิตปานกลาง อุปกรณ์อาจมีราคาสูงมาก |
| เซมิคอนดักเตอร์ไอออน (การจัดลำดับไอออนทอร์เรนต์) | มากถึง 600 bp [ 109 ] | 99.6% [ 110 ] | มากถึง 80 ล้าน | 2 ชั่วโมง | 66.8-950 ดอลลาร์สหรัฐ | อุปกรณ์ราคาถูกกว่า ทำงานได้เร็ว | ข้อผิดพลาดของโฮโมพอลิเมอร์ |
| ไพโรซีเควนซิง (454) | 700 bp | 99.9% | 1 ล้าน | 24 ชั่วโมง | 10,000 เหรียญสหรัฐ | ขนาดไฟล์อ่านใหญ่ เร็ว. | การผลิตจำนวนมากมีค่าใช้จ่ายสูง ข้อผิดพลาดจากโฮโมโพลิเมอร์ |
| การจัดลำดับดีเอ็นเอด้วยวิธีสังเคราะห์ (Illumina) | MiniSeq, NextSeq: 75–300 bp; MiSeq: 50–600 bp; HiSeq 2500: 50–500 bp; HiSeq 3/4000: 50–300 bp; HiSeq X: 300 bp | 99.9% (Phred30) | MiniSeq/MiSeq: 1–25 ล้าน; NextSeq: 130-00 ล้าน; HiSeq 2500: 300 ล้าน – 2 พันล้าน; HiSeq 3/4000 2.5 พันล้าน; HiSeq X: 3 พันล้าน | 1 ถึง 11 วัน ขึ้นอยู่กับซีเควนเซอร์และความยาวการอ่านที่ระบุ[ 111 ] | 5 ถึง 150 ดอลลาร์ | มีศักยภาพในการให้ผลผลิตลำดับสูง ขึ้นอยู่กับรุ่นของเครื่องจัดลำดับและแอปพลิเคชันที่ต้องการ | อุปกรณ์อาจมีราคาแพงมาก และต้องใช้ดีเอ็นเอที่มีความเข้มข้นสูง |
| การสังเคราะห์ตัวยึดโพรบแบบผสมผสาน (cPAS- BGI/MGI) | BGISEQ-50: 35-50bp; MGISEQ 200: 50-200bp; BGISEQ-500, MGISEQ-2000: 50-300bp [ 112 ] | 99.9% (Phred30) | BGISEQ-50: 160M; MGISEQ 200: 300 เมตร; BGISEQ-500: 1300M ต่อเซลล์การไหล; MGISEQ-2000: เซลล์ไหล FCS ขนาด 375M, เซลล์ไหล FCL ขนาด 1500M ต่อเซลล์ไหล | 1 ถึง 9 วัน ขึ้นอยู่กับเครื่องมือ ความยาวของการอ่าน และจำนวนเซลล์การไหลที่ทำงานพร้อมกันในแต่ละครั้ง | 5–120 ดอลลาร์ | ||
| การจัดลำดับดีเอ็นเอด้วยวิธีการเชื่อมต่อ (SOLiD sequencing) | 50+35 หรือ 50+50 bp | 99.9% | 1.2 ถึง 1.4 พันล้าน | 1 ถึง 2 สัปดาห์ | 60–130 เหรียญสหรัฐ | ต้นทุนต่อฐานต่ำ | ช้ากว่าวิธีอื่น มีปัญหาในการจัดลำดับพาลินโดรม[ 113 ] |
| การจัดลำดับนาโนพอเร | ขึ้นอยู่กับการเตรียมห้องสมุด ไม่ใช่ตัวอุปกรณ์ ดังนั้นผู้ใช้จึงเลือกความยาวการอ่าน (รายงานสูงสุด 2,272,580 bp [ 114 ] ) | ~92–97% การอ่านครั้งเดียว | ขึ้นอยู่กับความยาวของการอ่านที่ผู้ใช้เลือก | ข้อมูลถูกสตรีมแบบเรียลไทม์ เลือกช่วงเวลาได้ตั้งแต่ 1 นาที ถึง 48 ชั่วโมง | 7–100 เหรียญสหรัฐ | จำนวนผู้อ่านสูงสุด ชุมชนผู้ใช้เข้าถึงง่าย พกพาสะดวก (ขนาดเท่าฝ่ามือ) | อัตราการประมวลผลต่ำกว่าเครื่องอื่นๆ ความแม่นยำในการอ่านแต่ละครั้งอยู่ที่ประมาณ 90% |
| การจัดลำดับจีแนปซิส | ประมาณ 150 bp แบบปลายเดี่ยว | 99.9% (Phred30) | 1 ถึง 16 ล้าน | ประมาณ 24 ชั่วโมง | 667 เหรียญสหรัฐ | ต้นทุนของเครื่องมือต่ำ ($10,000) | |
| การยุติสายโซ่ (การจัดลำดับแบบแซงเกอร์) | 400 ถึง 900 bp | 99.9% | ไม่มีข้อมูล | 20 นาที ถึง 3 ชั่วโมง | 2,400,000 เหรียญสหรัฐ | มีประโยชน์สำหรับการใช้งานหลายประเภท | วิธีนี้มีราคาแพงกว่าและไม่เหมาะสมสำหรับโครงการลำดับดีเอ็นเอขนาดใหญ่ นอกจากนี้ยังต้องใช้ขั้นตอนการโคลนนิ่งพลาสมิดหรือ PCR ซึ่งใช้เวลานาน |
วิธีการจัดลำดับดีเอ็นเอแบบอ่านยาว
การจัดลำดับจีโนมแบบเรียลไทม์ระดับโมเลกุลเดี่ยว (SMRT)
การจัดลำดับดีเอ็นเอแบบ SMRT ใช้หลักการจัดลำดับโดยการสังเคราะห์ (sequencing by synthesis) โดยสังเคราะห์ดีเอ็นเอในตัวนำคลื่นแบบศูนย์โหมด (Zero-Mode Wave-Guides หรือ ZMWs) ซึ่งเป็นภาชนะขนาดเล็กคล้ายบ่อ โดยมีเครื่องมือจับยึดอยู่ที่ก้นบ่อ การจัดลำดับจะทำโดยใช้พอลิเมอเรสที่ไม่ถูกดัดแปลง (ติดอยู่กับก้น ZMW) และนิวคลีโอไทด์ที่ติดฉลากด้วยสารเรืองแสงซึ่งไหลเวียนอย่างอิสระในสารละลาย บ่อถูกสร้างขึ้นในลักษณะที่ตรวจจับเฉพาะแสงเรืองแสงที่เกิดขึ้นที่ก้นบ่อเท่านั้น สารเรืองแสงจะหลุดออกจากนิวคลีโอไทด์เมื่อมันถูกรวมเข้ากับสายดีเอ็นเอ ทำให้เหลือสายดีเอ็นเอที่ไม่ถูกดัดแปลง ตามข้อมูลจากPacific Biosciences (PacBio) ผู้พัฒนาเทคโนโลยี SMRT วิธีการนี้ช่วยให้สามารถตรวจจับการดัดแปลงนิวคลีโอไทด์ (เช่น การเมทิลเลชันของไซโตซีน) ได้ โดยการสังเกตจลนศาสตร์ของพอลิเมอเรส วิธีการนี้ช่วยให้สามารถอ่านลำดับนิวคลีโอไทด์ได้ 20,000 ตัวขึ้นไป โดยมีความยาวเฉลี่ย 5 กิโลเบส[ 105 ] [ 115 ]ในปี 2558 Pacific Biosciences ได้ประกาศเปิดตัวเครื่องมือจัดลำดับดีเอ็นเอรุ่นใหม่ชื่อ Sequel System ซึ่งมี ZMW จำนวน 1 ล้านตัว เมื่อเทียบกับ 150,000 ZMW ในเครื่องมือ PacBio RS II [ 116 ] [ 117 ]การจัดลำดับดีเอ็นเอแบบ SMRT เรียกว่า "การจัดลำดับรุ่นที่สาม " หรือ "การจัดลำดับแบบอ่านยาว"
การลำดับดีเอ็นเอด้วยนาโนพอเร
ดีเอ็นเอที่ผ่านรูนาโนจะทำให้กระแสไอออนเปลี่ยนแปลงไป การเปลี่ยนแปลงนี้ขึ้นอยู่กับรูปร่าง ขนาด และความยาวของลำดับดีเอ็นเอ นิวคลีโอไทด์แต่ละชนิดจะขัดขวางการไหลของไอออนผ่านรูเป็นระยะเวลาที่แตกต่างกัน วิธีนี้ไม่จำเป็นต้องใช้นิวคลีโอไทด์ที่ดัดแปลง และสามารถทำได้แบบเรียลไทม์ การจัดลำดับดีเอ็นเอด้วยนาโนพอเรเรียกว่า "การจัดลำดับรุ่นที่สาม " หรือ "การจัดลำดับแบบอ่านยาว" รวมถึงการจัดลำดับ SMRT ด้วย
การวิจัยทางอุตสาหกรรมในช่วงแรกเกี่ยวกับวิธีการนี้ใช้เทคนิคที่เรียกว่า 'การจัดลำดับเอ็กโซนิวคลีเอส' โดยการอ่านสัญญาณไฟฟ้าเกิดขึ้นเมื่อนิวคลีโอไทด์ผ่าน รูพรุน อัลฟา(α)-เฮโมไลซินที่ยึดด้วยพันธะโควาเลนต์กับไซโคลเดกซ์ทริน [ 118 ] อย่างไรก็ตามวิธีการเชิงพาณิชย์ที่ตามมาคือ 'การจัดลำดับสาย' ซึ่งจัดลำดับเบส DNA ในสายที่สมบูรณ์
การพัฒนาการจัดลำดับนาโนพอเรในสองด้านหลัก ได้แก่ การจัดลำดับนาโนพอเรแบบของแข็ง และการจัดลำดับนาโนพอเรแบบใช้โปรตีน การจัดลำดับนาโนพอเรแบบใช้โปรตีนใช้โปรตีนเชิงซ้อนของเยื่อหุ้มเซลล์ เช่น α-hemolysin, MspA ( Mycobacterium smegmatis Porin A) หรือ CssG ซึ่งมีแนวโน้มที่ดีเนื่องจากความสามารถในการแยกแยะระหว่างนิวคลีโอไทด์แต่ละตัวและกลุ่มของนิวคลีโอไทด์[ 119 ]ในทางตรงกันข้าม การจัดลำดับนาโนพอเรแบบของแข็งใช้วัสดุสังเคราะห์ เช่น ซิลิคอนไนไตรด์และอะลูมิเนียมออกไซด์ และเป็นที่นิยมเนื่องจากความสามารถเชิงกลที่เหนือกว่า ความเสถียรทางความร้อนและทางเคมี[ 120 ]วิธีการผลิตมีความสำคัญอย่างยิ่งสำหรับการจัดลำดับประเภทนี้ เนื่องจากอาร์เรย์นาโนพอเรสามารถมีรูพรุนหลายร้อยรูที่มีเส้นผ่านศูนย์กลางเล็กกว่าแปดนาโนเมตร[ 119 ]
แนวคิดนี้มีต้นกำเนิดมาจากความคิดที่ว่าโมเลกุล DNA หรือ RNA สายเดี่ยวสามารถถูกขับเคลื่อนด้วยไฟฟ้าในลำดับเชิงเส้นที่เข้มงวดผ่านรูพรุนทางชีวภาพที่มีขนาดเล็กกว่าแปดนาโนเมตร และสามารถตรวจจับได้เนื่องจากโมเลกุลปล่อยกระแสไอออนิกขณะเคลื่อนที่ผ่านรูพรุน รูพรุนประกอบด้วยบริเวณตรวจจับที่สามารถจดจำเบสต่างๆ ได้ โดยแต่ละเบสจะสร้างสัญญาณเฉพาะเวลาต่างๆ ที่สอดคล้องกับลำดับของเบสขณะที่พวกมันผ่านรูพรุน ซึ่งจะถูกประเมินในภายหลัง[ 120 ]การควบคุมการขนส่ง DNA ผ่านรูพรุนอย่างแม่นยำเป็นสิ่งสำคัญสำหรับความสำเร็จ เอนไซม์ต่างๆ เช่น เอ็กโซนิวคลีเอสและพอลิเมอเรสถูกนำมาใช้เพื่อควบคุมกระบวนการนี้โดยการวางตำแหน่งไว้ใกล้ทางเข้าของรูพรุน[ 121 ]
วิธีการจัดลำดับดีเอ็นเอแบบอ่านสั้น
การจัดลำดับลายเซ็นแบบขนานขนาดใหญ่ (MPSS)
เทคโนโลยีการจัดลำดับดีเอ็นเอความเร็วสูงรุ่นแรก คือการจัดลำดับแบบขนานขนาดใหญ่ (หรือ MPSS หรือที่เรียกว่าการจัดลำดับรุ่นใหม่) พัฒนาขึ้นในช่วงทศวรรษ 1990 ที่บริษัท Lynx Therapeutics ซึ่งก่อตั้งขึ้นในปี 1992 โดยSydney BrennerและSam Eletr MPSS เป็นวิธีการที่ใช้ลูกปัด (bead-based method) โดยใช้แนวทางที่ซับซ้อนในการเชื่อมต่ออะแดปเตอร์ (adapter ligation) ตามด้วยการถอดรหัสอะแดปเตอร์ (adapter decoding) โดยอ่านลำดับทีละสี่นิวคลีโอไทด์ วิธีนี้ทำให้เกิดความคลาดเคลื่อนจากอคติเฉพาะลำดับ (sequence-specific bias) หรือการสูญเสียลำดับเฉพาะได้ เนื่องจากเทคโนโลยีมีความซับซ้อนมาก MPSS จึงดำเนินการเฉพาะภายในบริษัท Lynx Therapeutics เท่านั้น และไม่มีการขายเครื่องจัดลำดับดีเอ็นเอให้กับห้องปฏิบัติการอิสระ Lynx Therapeutics ควบรวมกิจการกับ Solexa (ซึ่งต่อมาถูกซื้อกิจการโดยIllumina ) ในปี 2004 นำไปสู่การพัฒนาการจัดลำดับโดยการสังเคราะห์ (sequencing-by-synthesis) ซึ่งเป็นแนวทางที่ง่ายกว่าที่ได้มาจากManteia Predictive Medicineทำให้ MPSS ล้าสมัยไป อย่างไรก็ตาม คุณสมบัติที่สำคัญของเอาต์พุต MPSS นั้นเป็นแบบทั่วไปของประเภทข้อมูลที่มีปริมาณมากในภายหลัง ซึ่งรวมถึงลำดับ DNA สั้น ๆ หลายแสนรายการ ในกรณีของ MPSS โดยทั่วไปจะใช้สำหรับการจัดลำดับcDNAเพื่อวัดระดับการแสดงออกของยีน[ 71 ]
การจัดลำดับโพลอนี
วิธีการจัดลำดับโพลอนีซึ่งพัฒนาขึ้นในห้องปฏิบัติการของGeorge M. Churchที่มหาวิทยาลัยฮาร์วาร์ด เป็นหนึ่งในระบบการจัดลำดับที่มีปริมาณงานสูงระบบแรกๆ และถูกนำมาใช้ในการจัดลำดับ จีโนม E. coli ทั้งหมด ในปี 2548 [ 85 ]วิธีนี้รวมไลบรารีแบบจับคู่แท็กในหลอดทดลองเข้ากับ PCR แบบอิมัลชัน กล้องจุลทรรศน์อัตโนมัติ และเคมีจัดลำดับแบบใช้การเชื่อมต่อ เพื่อจัดลำดับ จีโนม E. coliด้วยความแม่นยำมากกว่า 99.9999% และมีต้นทุนประมาณ 1/9 ของการจัดลำดับแบบแซงเกอร์[ 85 ]เทคโนโลยีนี้ได้รับอนุญาตให้ Agencourt Biosciences ซึ่งต่อมาได้แยกตัวออกมาเป็น Agencourt Personal Genomics และในที่สุดก็ถูกรวมเข้ากับ แพลตฟอร์ม SOLiD ของ Applied Biosystemsต่อมา Applied Biosystems ถูกซื้อกิจการโดยLife Technologiesซึ่งปัจจุบันเป็นส่วนหนึ่งของThermo Fisher Scientific
ไพโรซีเควนซิง 454
454 Life Sciencesซึ่งต่อมาถูกซื้อกิจการโดยRoche Diagnosticsได้พัฒนา วิธี การไพโรซีเควนซิงแบบขนานขึ้นมาวิธีการนี้จะขยายดีเอ็นเอภายในหยดน้ำในสารละลายน้ำมัน (emulsion PCR) โดยแต่ละหยดน้ำจะมีแม่แบบดีเอ็นเอเพียงตัวเดียวที่ติดอยู่กับลูกปัดเคลือบไพรเมอร์เพียงตัวเดียว จากนั้นจึงสร้างโคโลนีโคลน เครื่องจัดลำดับดีเอ็นเอมี หลุมปริ มาตรพิโคลิตร จำนวนมาก โดยแต่ละหลุมจะมีลูกปัดและเอนไซม์จัดลำดับเพียงตัวเดียว ไพโรซีเควนซิงใช้ลูซิเฟอเรสในการสร้างแสงเพื่อตรวจจับนิวคลีโอไทด์แต่ละตัวที่เพิ่มเข้าไปในดีเอ็นเอที่เกิดขึ้นใหม่ และข้อมูลที่รวมกันจะถูกนำมาใช้ในการสร้างลำดับการอ่าน[ 84 ]เทคโนโลยีนี้ให้ความยาวการอ่านและราคาต่อเบสระดับกลางเมื่อเทียบกับการจัดลำดับแบบ Sanger ในด้านหนึ่ง และ Solexa และ SOLiD ในอีกด้านหนึ่ง[ 94 ]
การจัดลำดับดีเอ็นเอด้วยเครื่อง Illumina (Solexa)
Solexaซึ่งปัจจุบันเป็นส่วนหนึ่งของIlluminaก่อตั้งโดยShankar BalasubramanianและDavid Klenermanในปี 1998 และได้พัฒนาวิธีการจัดลำดับโดยใช้เทคโนโลยีตัวหยุดสีแบบย้อนกลับได้ และพอลิเมอเรสที่ได้รับการดัดแปลง[ 122 ]แนวคิดเคมีแบบหยุดย้อนกลับได้ถูกคิดค้นโดย Bruno Canard และ Simon Sarfati ที่สถาบัน Pasteur ในปารีส[ 123 ] [ 124 ]แนวคิดนี้ได้รับการพัฒนาขึ้นภายใน Solexa โดยผู้ที่ระบุชื่อไว้ในสิทธิบัตรที่เกี่ยวข้อง ในปี 2004 Solexa ได้เข้าซื้อกิจการบริษัทManteia Predictive Medicineเพื่อให้ได้เทคโนโลยีการจัดลำดับแบบขนานขนาดใหญ่ที่คิดค้นขึ้นในปี 1997 โดยPascal Mayerและ Laurent Farinelli [ 68 ]เทคโนโลยีนี้มีพื้นฐานมาจาก "กลุ่ม DNA" หรือ "อาณานิคม DNA" ซึ่งเกี่ยวข้องกับการขยาย DNA แบบโคลนบนพื้นผิว เทคโนโลยีคลัสเตอร์นี้ได้มาจากการร่วมซื้อกิจการกับ Lynx Therapeutics แห่งแคลิฟอร์เนีย ต่อมา Solexa Ltd. ได้ควบรวมกิจการกับ Lynx เพื่อก่อตั้งเป็น Solexa Inc.


ในวิธีการนี้ โมเลกุล DNA และไพรเมอร์จะถูกยึดติดกับสไลด์หรือเซลล์ไหลก่อน จากนั้นจึงขยายจำนวนด้วย พอ ลิเมอเรสทำให้เกิดกลุ่ม DNA โคลนเฉพาะที่ ซึ่งต่อมาเรียกว่า "คลัสเตอร์ DNA" ในการกำหนดลำดับ จะมีการเติมเบสเทอร์มิเนเตอร์แบบย้อนกลับได้ (RT-bases) สี่ชนิด และล้างนิวคลีโอไทด์ที่ไม่รวมตัวกันออกไป กล้องจะถ่ายภาพ นิวคลีโอไทด์ ที่ติดฉลากด้วยสารเรืองแสง จากนั้นสีย้อมพร้อมกับตัวบล็อก 3' ที่ปลายจะถูกกำจัดออกจาก DNA ด้วยกระบวนการทางเคมี ทำให้สามารถเริ่มรอบต่อไปได้ แตกต่างจากไพโรซีเควนซิง ตรงที่สาย DNA จะถูกขยายทีละนิวคลีโอไทด์ และการเก็บภาพสามารถทำได้ในเวลาที่ล่าช้า ทำให้สามารถจับภาพกลุ่ม DNA จำนวนมากได้ด้วยภาพต่อเนื่องจากกล้องตัวเดียว

การแยกปฏิกิริยาเอนไซม์และการจับภาพออกจากกันทำให้ได้ปริมาณงานที่เหมาะสมที่สุดและมีความสามารถในการจัดลำดับที่ไม่จำกัดในทางทฤษฎี ด้วยการกำหนดค่าที่เหมาะสมที่สุด ปริมาณงานของเครื่องมือที่สามารถเข้าถึงได้ในท้ายที่สุดจึงถูกกำหนดโดยอัตราการแปลงอนาล็อกเป็นดิจิทัลของกล้องเท่านั้น คูณด้วยจำนวนกล้องและหารด้วยจำนวนพิกเซลต่อกลุ่ม DNA ที่จำเป็นสำหรับการมองเห็นอย่างเหมาะสม (ประมาณ 10 พิกเซล/กลุ่ม) ในปี 2012 ด้วยกล้องที่ทำงานที่อัตราการแปลง A/D มากกว่า 10 MHz และเลนส์ ระบบของเหลว และเอนไซม์ที่มีอยู่ ปริมาณงานสามารถเป็นหลายเท่าของ 1 ล้านนิวคลีโอไทด์/วินาที ซึ่งสอดคล้องกับจีโนมมนุษย์เทียบเท่า 1 ตัวที่ความครอบคลุม 1x ต่อชั่วโมงต่อเครื่องมือ และการจัดลำดับจีโนมมนุษย์ใหม่ 1 ตัว (ที่ประมาณ 30x) ต่อวันต่อเครื่องมือ (ที่ติดตั้งกล้องตัวเดียว) [ 125 ]
การสังเคราะห์จุดยึดโพรบแบบผสมผสาน (cPAS)
วิธีการนี้เป็นการปรับปรุงแก้ไขเทคโนโลยีการเชื่อมต่อจุดยึดโพรบแบบผสมผสาน (cPAL) ที่อธิบายโดยComplete Genomics [ 126 ]ซึ่งต่อมาได้กลายเป็นส่วนหนึ่งของบริษัทจีโนมิกส์ของจีนBGIในปี 2013 [ 127 ]ทั้งสองบริษัทได้ปรับปรุงเทคโนโลยีนี้เพื่อให้สามารถอ่านข้อมูลได้ยาวขึ้น ลดเวลาปฏิกิริยา และได้ผลลัพธ์เร็วขึ้น นอกจากนี้ ข้อมูลยังถูกสร้างขึ้นเป็นข้อมูลการอ่านแบบเต็มความยาวต่อเนื่องในรูปแบบไฟล์ FASTQ มาตรฐาน และสามารถใช้งานได้โดยตรงในไปป์ไลน์การวิเคราะห์ข้อมูลชีวสารสนเทศแบบอ่านสั้นส่วนใหญ่[ 128 ] [ 129 ]
เทคโนโลยีสองอย่างที่เป็นพื้นฐานของเทคโนโลยีการจัดลำดับแบบความเร็วสูงนี้คือนาโนบอล DNA (DNB) และอาร์เรย์แบบมีลวดลายสำหรับการยึดนาโนบอลเข้ากับพื้นผิวแข็ง[ 126 ]นาโนบอล DNA ถูกสร้างขึ้นโดยการทำให้ไลบรารีแบบสายคู่ที่เชื่อมต่ออะแดปเตอร์แล้วเสียสภาพ และเชื่อมต่อเฉพาะสายไปข้างหน้ากับโอลิโกนิวคลีโอไทด์แบบสปลินต์เพื่อสร้างวงกลม ssDNA สำเนาที่ถูกต้องของวงกลมที่มี DNA แทรกจะถูกสร้างขึ้นโดยใช้การขยายแบบวงกลมแบบหมุน (Rolling Circle Amplification) ซึ่งสร้างสำเนาได้ประมาณ 300–500 ชุด สาย ssDNA ยาวจะพับทับกันเองเพื่อสร้างโครงสร้างนาโนบอลสามมิติที่มีเส้นผ่านศูนย์กลางประมาณ 220 นาโนเมตร การสร้าง DNB ช่วยลดความจำเป็นในการสร้างสำเนา PCR ของไลบรารีบนเซลล์ไหล และด้วยเหตุนี้จึงสามารถกำจัดลำดับการอ่านซ้ำ การเชื่อมต่ออะแดปเตอร์-อะแดปเตอร์ และข้อผิดพลาดที่เกิดจาก PCR ได้เป็นจำนวนมาก[ 128 ] [ 130 ]

อาร์เรย์ของจุดประจุบวกที่มีลวดลายถูกสร้างขึ้นผ่านเทคนิคโฟโตลิโทกราฟีและการกัดเซาะ ตามด้วยการดัดแปลงทางเคมีเพื่อสร้างเซลล์ไหลแบบลำดับ จุดแต่ละจุดบนเซลล์ไหลมีเส้นผ่านศูนย์กลางประมาณ 250 นาโนเมตร ห่างกัน 700 นาโนเมตร (จากจุดศูนย์กลางถึงจุดศูนย์กลาง) และช่วยให้สามารถยึด DNB ที่มีประจุลบเพียงตัวเดียวเข้ากับเซลล์ไหลได้ง่าย จึงช่วยลดการรวมกลุ่มที่น้อยเกินไปหรือมากเกินไปบนเซลล์ไหล[ 126 ] [ 131 ]
จากนั้นจึงทำการจัดลำดับโดยการเติมโพรบโอลิโกนิวคลีโอไทด์ที่ยึดติดกับตำแหน่งเฉพาะภายใน DNB โพรบทำหน้าที่เป็นตัวยึดเหนี่ยวที่ช่วยให้หนึ่งในสี่ของนิวคลีโอไทด์ที่ถูกทำให้ไม่ทำงานแบบย้อนกลับได้และติดฉลากสามารถจับได้หลังจากไหลผ่านเซลล์ไหล นิวคลีโอไทด์ที่ไม่จับจะถูกชะล้างออกไปก่อนที่จะกระตุ้นด้วยเลเซอร์ที่ฉลากที่จับอยู่ จากนั้นฉลากจะปล่อยแสงฟลูออเรสเซนต์และสัญญาณจะถูกจับโดยกล้องซึ่งจะถูกแปลงเป็นเอาต์พุตดิจิทัลสำหรับการระบุเบส เบสที่จับอยู่จะถูกตัดส่วนปลายและฉลากออกด้วยสารเคมีเมื่อสิ้นสุดวงจร วงจรจะถูกทำซ้ำด้วยการไหลของนิวคลีโอไทด์ที่ติดฉลากอิสระอีกครั้งผ่านเซลล์ไหลเพื่อให้นิวคลีโอไทด์ถัดไปจับและจับสัญญาณ กระบวนการนี้จะทำซ้ำหลายครั้ง (โดยปกติ 50 ถึง 300 ครั้ง) เพื่อกำหนดลำดับของชิ้นส่วน DNA ที่แทรกเข้าไปในอัตราประมาณ 40 ล้านนิวคลีโอไทด์ต่อวินาที ณ ปี 2018
การจัดลำดับ SOLiD

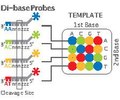
เทคโนโลยี SOLiD ของ Applied Biosystems (ปัจจุบันเป็น แบรนด์ ของ Life Technologies ) ใช้ การจัดลำดับโดยการเชื่อมต่อ (ligation ) โดยกลุ่มของโอลิโกนิวคลีโอไทด์ที่เป็นไปได้ทั้งหมดที่มีความยาวคงที่จะถูกติดฉลากตามตำแหน่งการจัดลำดับ โอลิโกนิวคลีโอไทด์จะถูกเชื่อมต่อและเชื่อมเข้าด้วยกัน การเชื่อมต่อแบบพิเศษโดย DNA ligaseสำหรับลำดับที่ตรงกันจะส่งผลให้เกิดสัญญาณที่ให้ข้อมูลเกี่ยวกับนิวคลีโอไทด์ในตำแหน่งนั้น แต่ละเบสในแม่แบบจะถูกจัดลำดับสองครั้ง และข้อมูลที่ได้จะถูกถอดรหัสตาม แผนการ เข้ารหัส 2 เบสที่ใช้ในวิธีนี้ ก่อนการจัดลำดับ DNA จะถูกขยายโดย emulsion PCR ลูกปัดที่ได้แต่ละเม็ดซึ่งมีสำเนาเดียวของโมเลกุล DNA เดียวกันจะถูกวางลงบนแผ่นกระจก[ 132 ]ผลลัพธ์ที่ได้คือลำดับที่มีปริมาณและความยาวเทียบเท่ากับการจัดลำดับของ Illumina [ 94 ] มีรายงานว่าวิธี การจัดลำดับโดยการเชื่อมต่อนี้มีปัญหาในการจัดลำดับลำดับพาลินโดรม[ 113 ]
การจัดลำดับเซมิคอนดักเตอร์ Ion Torrent
บริษัท Ion Torrent Systems Inc. (ปัจจุบันเป็นของLife Technologies ) ได้พัฒนาระบบที่ใช้เคมีลำดับมาตรฐาน แต่ใช้ระบบตรวจจับแบบใหม่ที่ใช้เซมิคอนดักเตอร์ วิธีการจัดลำดับนี้อาศัยการตรวจจับไอออนไฮโดรเจนที่ปล่อยออกมาในระหว่างการ พอ ลิเมอไรเซชันของDNAซึ่งแตกต่างจากวิธีการทางแสงที่ใช้ในระบบการจัดลำดับอื่นๆ ไมโครเวลล์ที่มีสาย DNA แม่แบบที่จะจัดลำดับจะถูกเติมด้วยนิวคลีโอไทด์ ชนิดเดียว หากนิวคลีโอไทด์ที่ใส่เข้าไปนั้นเป็นส่วนประกอบเสริมกับนิวคลีโอไทด์แม่แบบนำ มันจะถูกรวมเข้ากับสายเสริมที่กำลังเติบโต ซึ่งจะทำให้เกิดการปล่อยไอออนไฮโดรเจนที่กระตุ้นเซนเซอร์ไอออนที่มีความไวสูง ซึ่งบ่งชี้ว่าเกิดปฏิกิริยาขึ้น หาก มีการทำซ้ำ ของโฮโมพอลิเมอร์ในลำดับแม่แบบ นิวคลีโอไทด์หลายตัวจะถูกรวมเข้าในรอบเดียว ซึ่งนำไปสู่จำนวนไฮโดรเจนที่ปล่อยออกมาที่สอดคล้องกันและสัญญาณอิเล็กทรอนิกส์ที่สูงขึ้นตามสัดส่วน[ 133 ]

การจัดลำดับดีเอ็นเอนาโนบอล
การจัดลำดับดีเอ็นเอนาโนบอลเป็นเทคโนโลยีการจัดลำดับแบบความเร็วสูงชนิดหนึ่งที่ใช้ในการกำหนดลำดับจีโนม ทั้งหมด ของสิ่งมีชีวิต บริษัทComplete Genomicsใช้เทคโนโลยีนี้ในการจัดลำดับตัวอย่างที่ส่งมาจากนักวิจัยอิสระ วิธีนี้ใช้การจำลองแบบวงกลมเพื่อขยายชิ้นส่วนเล็กๆ ของดีเอ็นเอจีโนมให้เป็นดีเอ็นเอนาโนบอล จากนั้นจึงใช้การจัดลำดับแบบไม่ต่อเนื่องโดยการเชื่อมต่อเพื่อกำหนดลำดับนิวคลีโอไทด์[ 134 ]วิธีการจัดลำดับดีเอ็นเอแบบนี้ช่วยให้สามารถจัดลำดับดีเอ็นเอนาโนบอลจำนวนมากต่อรอบได้ และมี ต้นทุน สารเคมี ต่ำ เมื่อเทียบกับแพลตฟอร์มการจัดลำดับแบบความเร็วสูงอื่นๆ[ 135 ]อย่างไรก็ตาม จะมีการกำหนดลำดับดีเอ็นเอสั้นๆ จากดีเอ็นเอนาโนบอลแต่ละลูกเท่านั้น ซึ่งทำให้การจับคู่ลำดับสั้นๆ กับจีโนมอ้างอิงทำได้ยาก[ 134 ]
การจัดลำดับโมเลกุลเดี่ยว Heliscope
การจัดลำดับ Heliscope เป็นวิธีการจัดลำดับโมเลกุลเดี่ยวที่พัฒนาโดยHelicos Biosciencesโดยใช้ชิ้นส่วน DNA ที่มีอะแดปเตอร์หางโพลีเอที่เพิ่มเข้ามาซึ่งติดอยู่กับพื้นผิวของเซลล์ไหล ขั้นตอนต่อไปเกี่ยวข้องกับการจัดลำดับแบบขยายด้วยการล้างเซลล์ไหลเป็นวงจรด้วยนิวคลีโอไทด์ที่ติดฉลากด้วยฟลูออเรสเซนต์ (นิวคลีโอไทด์ชนิดเดียวในแต่ละครั้ง เช่นเดียวกับวิธี Sanger) การอ่านจะดำเนินการโดยเครื่องจัดลำดับ Heliscope [ 136 ] [ 137 ]การอ่านนั้นสั้น โดยเฉลี่ย 35 bp [ 138 ]สิ่งที่ทำให้เทคโนโลยีนี้มีความแปลกใหม่เป็นพิเศษคือ เป็นเทคโนโลยีแรกในกลุ่มที่จัดลำดับ DNA ที่ไม่ได้ขยาย จึงป้องกันข้อผิดพลาดในการอ่านที่เกี่ยวข้องกับขั้นตอนการขยาย[ 47 ]ในปี 2009 มีการจัดลำดับจีโนมมนุษย์โดยใช้ Heliscope อย่างไรก็ตาม ในปี 2012 บริษัทล้มละลาย[ 139 ]
ระบบไมโครฟลูอิดิก
ระบบไมโครฟลูอิดิกส์หลักๆ ที่ใช้ในการจัดลำดับดีเอ็นเอมีอยู่ 2 ระบบ ได้แก่ไมโครฟลูอิดิกส์แบบหยดและไมโครฟลูอิดิกส์แบบดิจิทัลอุปกรณ์ไมโครฟลูอิดิกส์ช่วยแก้ปัญหาข้อจำกัดหลายประการของระบบจัดลำดับดีเอ็นเอแบบอาร์เรย์ในปัจจุบัน
Abate และคณะได้ศึกษาการใช้อุปกรณ์ไมโครฟลูอิดิกแบบหยดสำหรับการจัดลำดับดีเอ็นเอ[ 4 ]อุปกรณ์เหล่านี้มีความสามารถในการสร้างและประมวลผลหยดขนาดพิโคลิตรในอัตราหลายพันหยดต่อวินาที อุปกรณ์เหล่านี้สร้างขึ้นจากโพลีไดเมทิลไซลอกเซน (PDMS)และใช้การถ่ายโอนพลังงานเรโซแนนซ์ของฟอร์สเตอร์ (FRET)เพื่ออ่านลำดับดีเอ็นเอที่อยู่ในหยด แต่ละตำแหน่งบนอาร์เรย์จะทดสอบลำดับเบส 15 ตัวที่เฉพาะเจาะจง[ 4 ]
Fair และคณะได้ใช้อุปกรณ์ไมโครฟลูอิดิกแบบดิจิทัลเพื่อศึกษาการจัดลำดับดีเอ็นเอแบบไพโรซีเควนซิง[ 140 ]ข้อดีที่สำคัญ ได้แก่ ความสะดวกในการพกพา ปริมาณรีเอเจนต์ ความเร็วในการวิเคราะห์ ความสามารถในการผลิตจำนวนมาก และปริมาณงานสูง การศึกษานี้ได้พิสูจน์แนวคิดที่แสดงให้เห็นว่าอุปกรณ์ดิจิทัลสามารถใช้สำหรับการจัดลำดับแบบไพโรซีเควนซิงได้ การศึกษานี้รวมถึงการใช้การสังเคราะห์ ซึ่งเกี่ยวข้องกับการขยายเอนไซม์และการเพิ่มนิวคลีโอไทด์ที่มีฉลาก[ 140 ]
Boles และคณะยังได้ศึกษาการจัดลำดับไพโรซีเควนซิงบนอุปกรณ์ไมโครฟลูอิดิกแบบดิจิทัลด้วย[ 141 ]พวกเขาใช้อุปกรณ์อิเล็กโทรเวตติ้งเพื่อสร้าง ผสม และแยกหยด การจัดลำดับใช้โปรโตคอลสามเอนไซม์และแม่แบบ DNA ที่ยึดด้วยลูกปัดแม่เหล็ก อุปกรณ์ได้รับการทดสอบโดยใช้สองโปรโตคอลและส่งผลให้มีความแม่นยำ 100% โดยพิจารณาจากระดับไพโรแกรมดิบ ข้อดีของอุปกรณ์ไมโครฟลูอิดิกแบบดิจิทัลเหล่านี้ ได้แก่ ขนาด ต้นทุน และระดับการบูรณาการการทำงานที่สามารถทำได้[ 141 ]
การวิจัยการจัดลำดับดีเอ็นเอโดยใช้ไมโครฟลูอิดิกส์ยังสามารถนำไปประยุกต์ใช้กับการจัดลำดับอาร์เอ็นเอโดยใช้เทคนิคไมโครฟลูอิดิกส์แบบหยดที่คล้ายกัน เช่น วิธีการ inDrops [ 142 ]ซึ่งแสดงให้เห็นว่าเทคนิคการจัดลำดับดีเอ็นเอเหล่านี้จำนวนมากจะสามารถนำไปประยุกต์ใช้ได้ต่อไปและใช้เพื่อทำความเข้าใจเกี่ยวกับจีโนมและทรานสคริปโตมได้มากขึ้น
วิธีการในการพัฒนา
วิธีการจัดลำดับดีเอ็นเอที่กำลังพัฒนาในปัจจุบัน ได้แก่ การอ่านลำดับขณะที่สายดีเอ็นเอเคลื่อนผ่านนาโนพอเรส (ซึ่งเป็นวิธีการที่ใช้ในเชิงพาณิชย์แล้ว แต่รุ่นต่อๆ ไป เช่น นาโนพอเรสแบบโซลิดสเตท ยังอยู่ในระหว่างการพัฒนา) [ 143 ] [ 144 ]และเทคนิคที่ใช้กล้องจุลทรรศน์ เช่นกล้องจุลทรรศน์แรงอะตอมหรือกล้องจุลทรรศน์อิเล็กตรอนแบบส่งผ่านซึ่งใช้ในการระบุตำแหน่งของนิวคลีโอไทด์แต่ละตัวภายในชิ้นส่วนดีเอ็นเอที่ยาว (>5,000 bp) โดยการติดฉลากนิวคลีโอไทด์ด้วยธาตุที่มีน้ำหนักมากกว่า (เช่น ฮาโลเจน) เพื่อการตรวจจับและบันทึกด้วยสายตา[ 145 ] [ 146 ]เทคโนโลยีรุ่นที่สามมีเป้าหมายเพื่อเพิ่มปริมาณงาน ลดเวลาในการได้ผลลัพธ์ และลดต้นทุนโดยการกำจัดความจำเป็นในการใช้สารเคมีมากเกินไป และใช้ประโยชน์จากกระบวนการทำงานของดีเอ็นเอพอลิเมอเรส[ 147 ]
กระแสอุโมงค์ การจัดลำดับดีเอ็นเอ
แนวทางอื่นใช้การวัดกระแสอุโมงค์ไฟฟ้าผ่าน DNA สายเดี่ยวขณะเคลื่อนที่ผ่านช่องทาง ขึ้นอยู่กับโครงสร้างอิเล็กตรอน เบสแต่ละตัวจะส่งผลต่อกระแสอุโมงค์แตกต่างกัน[ 148 ]ทำให้สามารถแยกแยะความแตกต่างระหว่างเบสต่างๆ ได้ [ 149 ]
การใช้กระแสอุโมงค์มีศักยภาพที่จะจัดลำดับได้เร็วกว่าวิธีการกระแสไอออนิกหลายเท่า และการจัดลำดับโอลิโกเมอร์ DNA และไมโครอาร์เอ็นเอหลายตัวก็ประสบความสำเร็จแล้ว[ 150 ]
การจัดลำดับโดยวิธีไฮบริดไดเซชัน
การจัดลำดับโดยวิธีไฮบริดไดเซชันเป็นวิธีการที่ไม่ใช้เอนไซม์โดยใช้ไมโครอาร์เรย์ DNAกลุ่ม DNA เดียวที่ต้องการกำหนดลำดับจะถูกติดฉลากด้วยสารเรืองแสงและไฮบริดไดเซชันกับอาร์เรย์ที่มีลำดับที่ทราบแล้ว สัญญาณไฮบริดไดเซชันที่แรงจากจุดใดจุดหนึ่งบนอาร์เรย์จะระบุลำดับของจุดนั้นใน DNA ที่กำลังจัดลำดับ [ 151 ]
วิธีการจัดลำดับนี้ใช้ลักษณะการจับของไลบรารีของโมเลกุล DNA สายเดี่ยวสั้นๆ (โอลิโกนิวคลีโอไทด์) หรือที่เรียกว่าโพรบ DNA เพื่อสร้างลำดับ DNA เป้าหมายขึ้นใหม่ ไฮบริดที่ไม่จำเพาะจะถูกกำจัดออกโดยการล้าง และ DNA เป้าหมายจะถูกชะออกมา[ 152 ]ไฮบริดจะถูกจัดเรียงใหม่เพื่อให้สามารถสร้างลำดับ DNA ขึ้นใหม่ได้ ข้อดีของการจัดลำดับประเภทนี้คือความสามารถในการจับเป้าหมายจำนวนมากด้วยความครอบคลุมที่สม่ำเสมอ[ 153 ]โดยปกติแล้วจะต้องใช้สารเคมีและ DNA เริ่มต้นจำนวนมาก อย่างไรก็ตาม ด้วยการเกิดขึ้นของการไฮบริดไดเซชันแบบใช้สารละลาย ทำให้ไม่จำเป็นต้องใช้อุปกรณ์และสารเคมีจำนวนมาก[ 152 ]
การจัดลำดับด้วยแมสสเปกโทรเมตรี
สเปกโทรเมตรีมวลอาจใช้เพื่อกำหนดลำดับดีเอ็นเอ สเปกโทรเมตรีมวลแบบ MALDI-TOF (Matrix-assisted laser desorption ionization time-of-flight mass spectrometry ) ได้รับการศึกษาเป็นพิเศษในฐานะวิธีการทางเลือกแทนเจลอิเล็กโทรโฟเรซิสสำหรับการมองเห็นชิ้นส่วนดีเอ็นเอ ด้วยวิธีนี้ ชิ้นส่วนดีเอ็นเอที่สร้างขึ้นโดยปฏิกิริยาการจัดลำดับแบบหยุดสายจะถูกเปรียบเทียบโดยมวลแทนที่จะเป็นขนาด มวลของนิวคลีโอไทด์แต่ละตัวจะแตกต่างจากตัวอื่น ๆ และความแตกต่างนี้สามารถตรวจจับได้ด้วยสเปกโทรเมตรีมวล การกลายพันธุ์ของนิวคลีโอไทด์เดี่ยวในชิ้นส่วนสามารถตรวจจับได้ง่ายกว่าด้วย MS มากกว่าเจลอิเล็กโทรโฟเรซิสเพียงอย่างเดียว MALDI-TOF MS สามารถตรวจจับความแตกต่างระหว่างชิ้นส่วน RNA ได้ง่ายกว่า ดังนั้นนักวิจัยอาจจัดลำดับดีเอ็นเอทางอ้อมด้วยวิธีการที่ใช้ MS โดยการแปลงเป็น RNA ก่อน[ 154 ]
ความละเอียดที่สูงขึ้นของชิ้นส่วน DNA ที่ได้จากวิธีการแบบ MS เป็นสิ่งที่นักวิจัยด้านนิติวิทยาศาสตร์ให้ความสนใจเป็นพิเศษ เนื่องจากพวกเขาอาจต้องการค้นหาโพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยวในตัวอย่าง DNA ของมนุษย์เพื่อระบุตัวบุคคล ตัวอย่างเหล่านี้อาจเสื่อมสภาพอย่างมาก ดังนั้นนักวิจัยด้านนิติวิทยาศาสตร์จึงมักเลือกใช้DNA ไมโทคอนเดรียเนื่องจากมีความเสถียรสูงกว่าและสามารถนำไปใช้ในการศึกษาลำดับวงศ์ตระกูลได้ วิธีการจัดลำดับแบบ MS ได้ถูกนำมาใช้เพื่อเปรียบเทียบลำดับของ DNA ไมโทคอนเดรียของมนุษย์จากตัวอย่างในฐานข้อมูลของสำนักงานสอบสวนกลาง[ 155 ]และจากกระดูกที่พบในหลุมฝังศพหมู่ของทหารในสงครามโลกครั้งที่ 1 [ 156 ]
วิธีการยุติสายโซ่และ TOF MS ในยุคแรกแสดงให้เห็นความยาวของการอ่านสูงสุดถึง 100 คู่เบส[ 157 ]นักวิจัยไม่สามารถเกินขนาดการอ่านเฉลี่ยนี้ได้ เช่นเดียวกับการจัดลำดับแบบยุติสายโซ่เพียงอย่างเดียว การจัดลำดับ DNA แบบ MS อาจไม่เหมาะสมสำหรับ โครงการจัดลำดับ de novo ขนาดใหญ่ ถึงกระนั้น การศึกษาในปี 2010 ก็ได้ใช้การอ่านลำดับสั้นและสเปกโทรสโกปีมวลเพื่อเปรียบเทียบโพลีมอร์ฟิซึมของนิวคลีโอไทด์เดี่ยวในสายพันธุ์Streptococcus ที่ก่อโรค [ 158 ]
การจัดลำดับดีเอ็นเอแบบแซงเกอร์ด้วยไมโครฟลูอิดิก
ในการจัดลำดับดีเอ็นเอแบบ Sanger ด้วยไมโครฟลูอิดิก การขยายดีเอ็นเอด้วยเทอร์โมไซคลิงทั้งหมด รวมถึงการแยกด้วยอิเล็กโทรโฟเรซิส จะทำบนแผ่นแก้วแผ่นเดียว (เส้นผ่านศูนย์กลางประมาณ 10 ซม.) ซึ่งช่วยลดการใช้สารเคมีและต้นทุน[ 159 ]ในบางกรณี นักวิจัยได้แสดงให้เห็นว่าพวกเขาสามารถเพิ่มปริมาณงานของการจัดลำดับแบบดั้งเดิมได้โดยใช้ไมโครชิป[ 160 ]ยังคงต้องมีการวิจัยเพิ่มเติมเพื่อให้การใช้เทคโนโลยีนี้มีประสิทธิภาพ
เทคนิคที่ใช้กล้องจุลทรรศน์
แนวทางนี้แสดงภาพลำดับของโมเลกุล DNA โดยตรงโดยใช้กล้องจุลทรรศน์อิเล็กตรอน การระบุคู่เบส DNA ครั้งแรกภายในโมเลกุล DNA ที่สมบูรณ์โดยการรวมเบสที่ดัดแปลงด้วยเอนไซม์ ซึ่งมีอะตอมที่มีเลขอะตอมเพิ่มขึ้น การแสดงภาพและการระบุเบสที่ติดฉลากแต่ละตัวโดยตรงภายในโมเลกุล DNA สังเคราะห์ 3,272 คู่เบส และจีโนมไวรัส 7,249 คู่เบส ได้รับการสาธิตแล้ว[ 161 ]
การจัดลำดับ RNAP
วิธีนี้ใช้เอนไซม์RNA polymerase (RNAP) ซึ่งติดอยู่กับ ลูกปัด โพลีสไตรีนปลายด้านหนึ่งของ DNA ที่ต้องการจัดลำดับจะติดอยู่กับลูกปัดอีกอัน โดยวางลูกปัดทั้งสองไว้ในกับดักแสง การเคลื่อนที่ของ RNAP ระหว่างการถอดรหัสจะทำให้ลูกปัดเข้าใกล้กันมากขึ้น และระยะห่างสัมพัทธ์ของลูกปัดจะเปลี่ยนแปลง ซึ่งสามารถบันทึกได้ที่ความละเอียดระดับนิวคลีโอไทด์เดี่ยว ลำดับจะถูกอนุมานจากค่าที่อ่านได้สี่ค่า โดยมีความเข้มข้นของนิวคลีโอไทด์ทั้งสี่ชนิดลดลง คล้ายกับวิธีของ Sanger [ 162 ]มีการเปรียบเทียบระหว่างภูมิภาค และข้อมูลลำดับจะถูกอนุมานโดยการเปรียบเทียบภูมิภาคที่มีลำดับที่ทราบกับภูมิภาคที่มีลำดับที่ไม่ทราบ[ 162 ]
การจัดลำดับจีโนมไวรัส ในหลอดทดลองด้วยความเร็วสูง
ได้มีการพัฒนาวิธีการวิเคราะห์ชุดปฏิสัมพันธ์ของโปรตีน ทั้งหมด โดยใช้การผสมผสานระหว่าง 454 pyrosequencing และ วิธี การแสดงผล mRNAของไวรัสในหลอดทดลองโดยเฉพาะอย่างยิ่ง วิธีนี้จะเชื่อมโยงโปรตีนที่สนใจเข้ากับ mRNA ที่เข้ารหัสโปรตีนเหล่านั้นด้วยพันธะโควาเลนต์ จากนั้นตรวจจับชิ้นส่วน mRNA โดยใช้PCR แบบย้อนกลับการถอดรหัส จากนั้น mRNA อาจถูกขยายและจัดลำดับ วิธีการผสมผสานนี้มีชื่อว่า IVV-HiTSeq และสามารถดำเนินการได้ภายใต้สภาวะที่ปราศจากเซลล์ แม้ว่าผลลัพธ์อาจไม่เป็นตัวแทนของสภาวะในร่างกาย ก็ตาม [ 163 ]
ส่วนแบ่งการตลาด
แม้ว่าจะมีวิธีการจัดลำดับดีเอ็นเอที่แตกต่างกันมากมาย แต่มีเพียงไม่กี่วิธีเท่านั้นที่ครองตลาด ในปี 2022 Illumina มีส่วนแบ่งการตลาดประมาณ 80% ส่วนที่เหลือของตลาดถูกครอบครองโดยผู้เล่นเพียงไม่กี่ราย (PacBio, Oxford, 454, MGI) [ 164 ]
การเตรียมตัวอย่าง
ความสำเร็จของกระบวนการจัดลำดับดีเอ็นเอใดๆ ขึ้นอยู่กับการสกัดและการเตรียมตัวอย่างดีเอ็นเอหรืออาร์เอ็นเอจากวัสดุชีวภาพที่สนใจ
- การสกัด DNA ที่ประสบความสำเร็จจะให้ตัวอย่าง DNA ที่มีสายยาวและไม่เสื่อมสภาพ
- การสกัด RNA ที่ประสบความสำเร็จจะให้ตัวอย่าง RNA ที่ควรแปลงเป็น DNA เสริม (cDNA) โดยใช้เอนไซม์รีเวอร์สทรานสคริปเทส— เอนไซม์รีเวอร์สทรานสคริปเทสจะสังเคราะห์ DNA เสริมแบบสองสายโดยอิงจากสาย RNA ที่มีอยู่ และกลุ่ม cDNA ที่ได้สามารถนำไปจัดลำดับได้[ 165 ]โดยทั่วไปปฏิกิริยานี้จะเริ่มต้นด้วยเฮกซาเมอร์แบบสุ่มหรือไพรเมอร์ที่กำหนดเป้าหมายไปที่ หาง โพลี(A) ของ mRNA จากนั้น cDNA สามารถนำไปประมวลผลได้ในลักษณะเดียวกับ DNA จีโนม
หลังจากสกัด DNA หรือ RNA แล้ว ตัวอย่างอาจต้องมีการเตรียมเพิ่มเติมขึ้นอยู่กับวิธีการจัดลำดับ สำหรับการจัดลำดับแบบ Sanger จำเป็นต้องมีขั้นตอนการโคลนหรือ PCR ก่อนการจัดลำดับ ในกรณีของวิธีการจัดลำดับรุ่นใหม่ จำเป็นต้องมีการเตรียมไลบรารีก่อนการประมวลผล[ 166 ]การประเมินคุณภาพและปริมาณของกรดนิวคลีอิกทั้งหลังการสกัดและหลังการเตรียมไลบรารีจะช่วยระบุตัวอย่างที่เสื่อมสภาพ แตกหัก และมีความบริสุทธิ์ต่ำ และให้ข้อมูลการจัดลำดับที่มีคุณภาพสูง[ 167 ]
โครงการพัฒนา

ในเดือนตุลาคม พ.ศ. 2549 มูลนิธิ X Prizeได้ริเริ่มโครงการเพื่อส่งเสริมการพัฒนาเทคโนโลยีการจัดลำดับจีโนมแบบสมบูรณ์ เรียกว่า Archon X Prizeโดยตั้งใจจะมอบเงิน 10 ล้านดอลลาร์สหรัฐให้แก่ "ทีมแรกที่สามารถสร้างอุปกรณ์และใช้งานเพื่อจัดลำดับจีโนมมนุษย์ 100 จีโนมภายใน 10 วันหรือน้อยกว่านั้น โดยมีความแม่นยำไม่เกิน 1 ข้อผิดพลาดในทุกๆ 100,000 เบสที่จัดลำดับ โดยมีลำดับที่ครอบคลุมจีโนมอย่างน้อย 98% อย่างแม่นยำ และมีค่าใช้จ่ายต่อเนื่องไม่เกิน 10,000 ดอลลาร์สหรัฐต่อจีโนม" [ 168 ]
ทุกปีสถาบันวิจัยจีโนมมนุษย์แห่งชาติหรือ NHGRI จะส่งเสริมการให้ทุนสำหรับการวิจัยและพัฒนาใหม่ๆ ในด้านจีโนมิกส์ทุนในปี 2010 และผู้สมัครในปี 2011 รวมถึงงานต่อเนื่องในวิธีการจัดลำดับไมโครฟลูอิดิก โพลอนี และเบสหนัก[ 169 ]
ความท้าทายในการคำนวณ
เทคโนโลยีการจัดลำดับที่อธิบายไว้ในที่นี้สร้างข้อมูลดิบที่ต้องนำมาประกอบกันเป็นลำดับที่ยาวขึ้น เช่น จีโนมที่สมบูรณ์ ( การประกอบลำดับ ) มีความท้าทายทางด้านการคำนวณมากมายในการบรรลุเป้าหมายนี้ เช่น การประเมินข้อมูลลำดับดิบซึ่งทำโดยโปรแกรมและอัลกอริธึม เช่นPhredและPhrapความท้าทายอื่นๆ คือการจัดการกับ ลำดับ ที่ซ้ำกันซึ่งมักจะขัดขวางการประกอบจีโนมที่สมบูรณ์ เนื่องจากเกิดขึ้นในหลายตำแหน่งของจีโนม ผลที่ตามมาคือ ลำดับจำนวนมากอาจไม่ถูกกำหนดให้กับโครโมโซม ที่เฉพาะเจาะจง การสร้างข้อมูลลำดับดิบเป็นเพียงจุดเริ่มต้นของการวิเคราะห์ทางชีวสารสนเทศ อย่างละเอียด [ 170 ]อย่างไรก็ตาม วิธีการใหม่สำหรับการจัดลำดับและการแก้ไขข้อผิดพลาดในการจัดลำดับได้รับการพัฒนาขึ้น[ 171 ]
อ่านการตัดแต่ง
บางครั้ง ข้อมูลดิบที่ได้จากการจัดลำดับดีเอ็นเอจะถูกต้องและแม่นยำเฉพาะในส่วนย่อยของความยาวเท่านั้น การใช้ข้อมูลทั้งหมดอาจทำให้เกิดสิ่งผิดปกติในการวิเคราะห์ขั้นต่อไป เช่น การประกอบจีโนม การระบุ SNP หรือการประมาณการแสดงออกของยีน มีการแนะนำโปรแกรมตัดแต่งสองประเภท โดยอิงตามอัลกอริทึมแบบใช้หน้าต่างหรือแบบผลรวมต่อเนื่อง[ 172 ]นี่คือรายการบางส่วนของอัลกอริทึมตัดแต่งที่มีอยู่ในปัจจุบัน โดยระบุประเภทของอัลกอริทึมที่อัลกอริทึมเหล่านั้นสังกัดอยู่:
| ชื่อของอัลกอริทึม | ประเภทของอัลกอริทึม |
|---|---|
| Cutadapt [ 173 ] | ผลรวมสะสม |
| ConDeTri [ 174 ] | อิงตามหน้าต่าง |
| ตัวกรอง ERNE [ 175 ] | ผลรวมสะสม |
| เครื่องตัดแต่งคุณภาพสูง FASTX | อิงตามหน้าต่าง |
| ปรินเซค[ 176 ] | อิงตามหน้าต่าง |
| Trimmomatic [ 177 ] | อิงตามหน้าต่าง |
| SolexaQA [ 178 ] | อิงตามหน้าต่าง |
| โซเลกซาคิวเอ-บีดับบลิวเอ | ผลรวมสะสม |
| เคียว | อิงตามหน้าต่าง |
ประเด็นด้านจริยธรรม
พันธุศาสตร์ของมนุษย์ถูกรวมอยู่ในสาขาจริยธรรมชีวภาพตั้งแต่ต้นทศวรรษ 1970 [ 179 ]และการเติบโตของการใช้การจัดลำดับดีเอ็นเอ (โดยเฉพาะการจัดลำดับแบบความเร็วสูง) ได้นำมาซึ่งประเด็นทางจริยธรรมหลายประการ ประเด็นสำคัญประการหนึ่งคือกรรมสิทธิ์ในดีเอ็นเอของแต่ละบุคคลและข้อมูลที่ได้จากการจัดลำดับดีเอ็นเอ[ 180 ]ในส่วนของโมเลกุลดีเอ็นเอเอง คดีทางกฎหมายที่สำคัญในหัวข้อนี้คือคดีMoore v. Regents of the University of California (1990) ได้ตัดสินว่าบุคคลไม่มีสิทธิ์ในทรัพย์สินใดๆ ต่อเซลล์ที่ถูกทิ้งหรือผลกำไรใดๆ ที่ได้จากการใช้เซลล์เหล่านี้ (เช่นเซลล์สายพันธุ์ ที่จดสิทธิบัตร ) อย่างไรก็ตาม บุคคลมีสิทธิ์ได้รับความยินยอมโดยแจ้งให้ทราบเกี่ยวกับการนำเซลล์ออกและการใช้เซลล์ ในส่วนของข้อมูลที่ได้จากการจัดลำดับดีเอ็นเอ คดีMooreไม่ได้ให้สิทธิ์ใดๆ แก่บุคคลในข้อมูลที่ได้จากดีเอ็นเอของพวกเขา[ 180 ]
เมื่อการจัดลำดับดีเอ็นเอแพร่หลายมากขึ้น การจัดเก็บ ความปลอดภัย และการแบ่งปันข้อมูลจีโนมก็มีความสำคัญมากขึ้นเช่นกัน[ 180 ] [ 181 ]ตัวอย่างเช่น ข้อกังวลประการหนึ่งคือ บริษัทประกันภัยอาจใช้ข้อมูลจีโนมของบุคคลเพื่อปรับเปลี่ยนราคาเบี้ยประกัน โดยขึ้นอยู่กับสุขภาพในอนาคตที่คาดการณ์ไว้ของบุคคลนั้นจากดีเอ็นเอของพวกเขา[ 181 ] [ 182 ]ในเดือนพฤษภาคม พ.ศ. 2551 พระราชบัญญัติการไม่เลือกปฏิบัติทางข้อมูลพันธุกรรม (GINA) ได้รับการลงนามในสหรัฐอเมริกา ซึ่งห้ามการเลือกปฏิบัติบนพื้นฐานของข้อมูลพันธุกรรมเกี่ยวกับการประกันสุขภาพและการจ้างงาน[ 183 ] [ 184 ]ในปี 2555 คณะกรรมการประธานาธิบดีสหรัฐฯ เพื่อการศึกษาประเด็นจริยธรรมชีวภาพได้รายงานว่ากฎหมายคุ้มครองความเป็นส่วนตัวที่มีอยู่สำหรับข้อมูลลำดับดีเอ็นเอ เช่น GINA และพระราชบัญญัติการพกพาและการรับผิดชอบด้านการประกันสุขภาพนั้นไม่เพียงพอ โดยระบุว่าข้อมูลลำดับจีโนมทั้งหมดมีความละเอียดอ่อนเป็นพิเศษ เนื่องจากสามารถนำไปใช้ระบุตัวตนได้ไม่เพียงแต่บุคคลที่สร้างข้อมูลนั้นขึ้นมาเท่านั้น แต่ยังรวมถึงญาติของบุคคลนั้นด้วย[ 185 ] [ 186 ]
ในสหรัฐอเมริกาส่วนใหญ่ ดีเอ็นเอที่ "ถูกทิ้ง" เช่น ดีเอ็นเอที่พบในแสตมป์หรือซองจดหมายที่ถูกเลีย ถ้วยกาแฟ บุหรี่ หมากฝรั่ง ขยะในครัวเรือน หรือเส้นผมที่ร่วงหล่นบนทางเท้าสาธารณะ สามารถเก็บรวบรวมและวิเคราะห์ลำดับได้โดยชอบด้วยกฎหมายโดยใครก็ได้ รวมถึงตำรวจ นักสืบเอกชน ฝ่ายตรงข้ามทางการเมือง หรือผู้ที่เกี่ยวข้องในข้อพิพาทเรื่องความเป็นพ่อ ณ ปี 2013 มี 11 รัฐที่มีกฎหมายที่สามารถตีความได้ว่าห้าม "การขโมยดีเอ็นเอ" [ 187 ]
ประเด็นด้านจริยธรรมยังถูกหยิบยกขึ้นมาจากการใช้การตรวจคัดกรองความแปรผันทางพันธุกรรมที่เพิ่มมากขึ้น ทั้งในทารกแรกเกิดและในผู้ใหญ่ โดยบริษัทต่างๆ เช่น23andMe [ 188 ] [ 189 ] มีการกล่าวอ้างว่าการตรวจคัดกรองความแปรผันทางพันธุกรรมอาจเป็นอันตรายและเพิ่มความวิตกกังวลในบุคคลที่พบว่ามีความเสี่ยงต่อโรคเพิ่มขึ้น[ 190 ]ตัวอย่างเช่น ในกรณีหนึ่งที่กล่าวถึงในTimeแพทย์ที่ตรวจคัดกรองความแปรผันทางพันธุกรรมในทารกที่ป่วยเลือกที่จะไม่แจ้งให้ผู้ปกครองทราบถึงความแปรผันที่ไม่เกี่ยวข้องซึ่งเชื่อมโยงกับภาวะสมองเสื่อมเนื่องจากเกรงว่าจะก่อให้เกิดอันตรายต่อผู้ปกครอง[ 191 ]อย่างไรก็ตาม การศึกษาในปี 2011 ในThe New England Journal of Medicineแสดงให้เห็นว่าบุคคลที่เข้ารับการประเมินความเสี่ยงของโรคไม่ได้แสดงระดับความวิตกกังวลที่เพิ่มขึ้น[ 190 ]นอกจากนี้ การพัฒนาเทคโนโลยีการจัดลำดับรุ่นต่อไป เช่น การจัดลำดับแบบ Nanopore ยังก่อให้เกิดข้อกังวลด้านจริยธรรมเพิ่มเติมอีกด้วย[ 192 ]
ดูเพิ่มเติม
- ชีวสารสนเทศศาสตร์ – การวิเคราะห์เชิงคำนวณของชุดข้อมูลทางชีวภาพขนาดใหญ่และซับซ้อน
- การถอดรหัสจีโนมมะเร็ง
- การจัดลำดับฉันทามติแบบวงกลม
- การคำนวณด้วยดีเอ็นเอ – การคำนวณโดยใช้ฮาร์ดแวร์ทางชีววิทยาระดับโมเลกุล
- ทรานซิสเตอร์สนามไฟฟ้าดีเอ็นเอ
- ทฤษฎีการเรียงลำดับดีเอ็นเอ – ทฤษฎีทางชีววิทยา
- เครื่องถอดรหัสลำดับดีเอ็นเอ – เครื่องมือทางวิทยาศาสตร์ที่ทำให้กระบวนการถอดรหัสลำดับดีเอ็นเอเป็นไปโดยอัตโนมัติ
- โครงการ Genographic – โครงการวิทยาศาสตร์ภาคประชาชน
- โครงการจีโนม – ความพยายามทางวิทยาศาสตร์เพื่อกำหนดลำดับจีโนมที่สมบูรณ์ของสิ่งมีชีวิต
- การจัดลำดับจีโนมของสัตว์ใกล้สูญพันธุ์ – การทดสอบดีเอ็นเอเพื่อประเมินความเสี่ยงต่อการสูญ พันธุ์
- การสุ่มตัวอย่างจีโนม – วิธีการจัดลำดับจีโนม
- IsoBase – ฐานข้อมูลสำหรับระบุโปรตีนที่มีความสัมพันธ์กันในเชิงหน้าที่
- การจัดลำดับแบบอ่านเชื่อมโยง
- ห้องสมุดกระโดด – ชุดรวมชิ้นส่วนดีเอ็นเอจีโนม
- ลำดับกรดนิวคลีอิก – ลำดับของนิวคลีโอไทด์ในกรดนิวคลีอิก
- การขยายสัญญาณโพรบแบบพึ่งพาการเชื่อมต่อหลายจุด – วิธีการตรวจจับทางชีวเคมี
- การแพทย์เฉพาะบุคคล – รูปแบบทางการแพทย์ที่ปรับวิธีการรักษาให้เหมาะสมกับผู้ป่วยแต่ละราย
- การจัดลำดับโปรตีน – การจัดลำดับการเรียงตัวของกรดอะมิโนในโปรตีน
- การขุดลำดับ – เทคนิคการขุดข้อมูล
- เครื่องมือวิเคราะห์ลำดับดีเอ็นเอ
- การจัดลำดับโดยวิธีไฮบริดไดเซชัน
- การจัดลำดับโดยการเชื่อมต่อ
- TIARA (ฐานข้อมูล) – ฐานข้อมูลข้อมูลจีโนมส่วนบุคคล
- การจัดลำดับดีเอ็นเอด้วยกล้องจุลทรรศน์อิเล็กตรอนแบบส่งผ่าน – เทคโนโลยีการจัดลำดับระดับโมเลกุลเดี่ยว
หมายเหตุ
- ^ คำว่า "รุ่นต่อไป" ยังคงถูกใช้อย่างแพร่หลายในปี 2019 ตัวอย่างเช่น Straiton J, Free T, Sawyer A, Martin J (กุมภาพันธ์ 2019) "จากการจัดลำดับแบบ Sanger ไปสู่ฐานข้อมูลจีโนมและอื่นๆ" BioTechniques 66 ( 2 ): 60– 63. doi : 10.2144/btn-2019-0011 . PMID 30744413 เทคโนโลยีการจัดลำดับรุ่นต่อไป (
NGS) ได้ปฏิวัติการวิจัยทางจีโนม (ประโยคเปิดของบทความ)
ลิงก์ภายนอก
- วิกิตำราเกี่ยวกับการเรียงลำดับรุ่นต่อไป
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การจัดลำดับดีเอ็นเอ
การจัดลำดับดีเอ็นเอเป็นกระบวนการกำหนดลำดับกรดนิวคลี อิก – ลำดับของนิวคลีโอไทด์ในดีเอ็นเอซึ่งรวมถึงวิธีการหรือเทคโนโลยีใดๆ ที่ใช้ในการกำหนดลำดับของเบสทั้งสี่
แอปพลิเคชัน
การจัดลำดับดีเอ็นเอสามารถใช้เพื่อกำหนดลำดับของ ยีน แต่ละตัว บริเวณทางพันธุกรรมที่ใหญ่กว่า (เช่น กลุ่มยีนหรือ โอเปรอน ) โครโมโซมทั้งหมด หรือ จีโนมทั้งหมด ของสิ่งมีชีวิตใดๆ การจัดลำดับดีเอ็นเอยังเป็นวิธีที่มีประสิทธิภาพที่สุดในการจัดลำดับ อาร์เอ็นเอ หรือ โปรตีน...
ชีววิทยาโมเลกุล
การจัดลำดับดีเอ็นเอเป็นเทคนิคที่ใช้ใน ชีววิทยาระดับโมเลกุล เพื่อศึกษาจีโนมและโปรตีนที่เข้ารหัสโดยจีโนมเหล่านั้น ข้อมูลที่ได้จากการจัดลำดับดีเอ็นเอช่วยให้นักวิจัยสามารถระบุการเปลี่ยนแปลงในยีนและดีเอ็นเอที่ไม่เข้ารหัส (รวมถึงลำดับควบคุม)...
ชีววิทยาเชิงวิวัฒนาการ
เนื่องจาก DNA เป็น โมเลกุลขนาดใหญ่ ที่มีข้อมูล ในแง่ของการส่งต่อจากรุ่นสู่รุ่น การจัดลำดับ DNA จึงถูกนำมาใช้ใน ชีววิทยาวิวัฒนาการ เพื่อศึกษาความสัมพันธ์ระหว่างสิ่งมีชีวิตต่าง ๆ และวิวัฒนาการของพวกมัน ในเดือนกุมภาพันธ์ พ.ศ.