อ่าน 25 นาที
เฆี่ยน
แฟ ลเจลลัม ( / f l ə ˈ dʒ ɛ l əm / ; พหูพจน์ : flagella ) ( ภาษาละติน แปลว่า 'แส้' หรือ 'แส้') คือ ระยางค์ คล้ายเส้นผมที่ยื่นออกมาจาก เซลล์สเปิร์ม ของ พืช และสัตว์ บางชนิด จาก...
เฆี่ยน
| เฆี่ยน | |
|---|---|
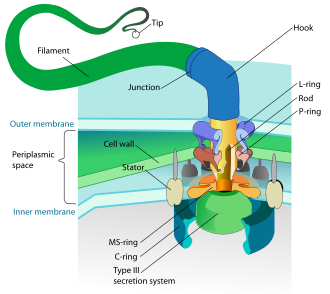 โครงสร้างของแฟลเจลลัมของแบคทีเรีย | |
 | |
| ตัวระบุ | |
| เมช | D005407 |
| ไทย | H1.00.01.1.01032 |
| เอฟเอ็มเอ | 67472 |
| ศัพท์ทางกายวิภาคศาสตร์ | |
แฟลเจลลัม ( / f l ə ˈ dʒ ɛ l əm / ; พหูพจน์ : flagella ) ( ภาษาละติน แปลว่า 'แส้' หรือ 'แส้') คือ ระยางค์คล้ายเส้นผมที่ยื่นออกมาจากเซลล์สเปิร์ม ของ พืชและสัตว์ บางชนิด จากสปอร์ของเชื้อรา ( ซูโอสปอร์ ) และจาก จุลินทรีย์หลากหลายชนิดเพื่อช่วยในการเคลื่อนที่[ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ]โปรติสต์หลายชนิดที่มีแฟลเจลลัมเรียกว่า แฟลเจ ล เลต
จุลินทรีย์อาจมีแฟลเจลลาตั้งแต่หนึ่งถึงหลายอันตัวอย่างเช่นแบคทีเรียแกรมลบHelicobacter pylori ใช้แฟลเจลลาในการเคลื่อนที่ผ่านกระเพาะอาหารไปยัง เยื่อบุผิวซึ่งอาจเข้าไปอาศัยอยู่ในเยื่อบุผิวและอาจทำให้เกิดโรคกระเพาะอักเสบและแผลในกระเพาะ อาหาร ซึ่งเป็นปัจจัยเสี่ยงต่อมะเร็งกระเพาะอาหาร [ 6 ] ในแบคทีเรียที่เคลื่อนที่เป็นกลุ่ม บางชนิด แฟลเจลลายังสามารถทำหน้าที่เป็นออร์แกเนลล์ รับ ความรู้สึก โดยมีความไวต่อความชื้นภายนอกเซลล์[ 7 ]
แฟ ลเจลลั ม ในสามโดเมนของแบคทีเรียอาร์เคียและยูคาริโอตามีโครงสร้าง องค์ประกอบโปรตีน และกลไกการขับเคลื่อนที่แตกต่างกัน แต่มีหน้าที่เดียวกันคือการทำให้เกิดการเคลื่อนที่คำภาษาละตินflagellumหมายถึง " แส้ " เพื่ออธิบายการเคลื่อนไหวในการว่ายน้ำที่คล้ายกับแส้ แฟลเจลลัมในอาร์เคียเรียกว่าarchaellumเพื่อสังเกตความแตกต่างจากแฟลเจลลัมของแบคทีเรีย[ 8 ] [ 9 ]
Eukaryotic flagella and cilia are identical in structure but have different lengths and functions.[10]Prokaryoticfimbriae and pili are smaller, and thinner appendages, with different functions. Surface-attached cilia and flagella are used to swim or move fluid from one region to another.[11]
Types
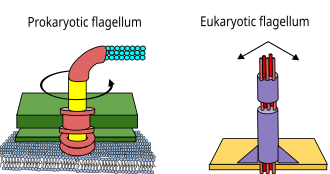
The three types of flagella are bacterial, archaeal, and eukaryotic.
The flagella in eukaryotes have dynein and microtubules that move with a bending mechanism. Bacteria and archaea do not have dynein or microtubules in their flagella, and they move using a rotary mechanism.[13]
Other differences among these three types are:
- Bacterial flagella[2] are helical filaments, each with a rotary motor at its base which can turn clockwise or counterclockwise.[14][15][16] They provide two of several kinds of bacterial motility.[17][18]
- Archaeal flagella (archaella) are superficially similar to bacterial flagella in that it also has a rotary motor, but are different in many details and considered non-homologous.[19][20][21]
- Eukaryotic flagella—those of animal, plant, and protist cells—are complex cellular projections that lash back and forth. Eukaryotic flagella and motile cilia are identical in structure, but have different lengths, waveforms, and functions. Primary cilia are immotile, and have a structurally different9+0 axoneme rather than the 9+2 axoneme found in both flagella and motile cilia.
Bacterial flagella
Structure and composition
การประกอบและการทำงานของแฟลเจลลาของแบคทีเรียต้องอาศัยโปรตีนมากกว่า 50 ชนิดที่ทำงานร่วมกัน[ 22 ]แฟลเจลลัมของแบคทีเรียประกอบด้วย หน่วยย่อย โปรตีนของแฟลเจลลิน [ 13 ] รูปร่างของมันคือท่อกลวงหนา 20 นาโนเมตร มี ลักษณะเป็นเกลียวและมีส่วนโค้งแหลมอยู่ด้านนอกเยื่อหุ้มเซลล์ชั้นนอก "ตะขอ" นี้ทำให้แกนของเกลียวชี้ออกไปจากเซลล์โดยตรง มีแกนวิ่งระหว่างตะขอและฐานของแฟลเจลลัม โดยผ่านวงแหวนโปรตีนในเยื่อหุ้มเซลล์ที่ทำหน้าที่เป็นแบริ่งจุลินทรีย์แกรมบวกมีวงแหวนฐานของแฟลเจลลัมสองวง วงหนึ่งอยู่ใน ชั้น เพปติโดไกลแคนและอีกวงหนึ่งอยู่ในเยื่อหุ้มพลาสมาจุลินทรีย์แกรมลบมีวงแหวนดังกล่าวสี่วง ได้แก่วงแหวน Lเกี่ยวข้องกับลิโปโพลีแซ คคาไรด์ วงแหวนPเกี่ยวข้องกับ ชั้น เพปติโดไกลแคนวงแหวน M ฝังอยู่ในเยื่อหุ้มพลาสมาและวงแหวน S ติดอยู่กับไซโตพลาส ซึมโดยตรง เส้นใยจะสิ้นสุดด้วยโปรตีนปิดปลาย[ 23 ] [ 24 ]
เส้นใยแฟลเจลลาเป็นเกลียวยาวที่ขับเคลื่อนแบคทีเรียเมื่อหมุนโดยมอเตอร์ผ่านตะขอ ในแบคทีเรียส่วนใหญ่ที่ได้รับการศึกษา รวมถึงแบคทีเรียแกรมลบEscherichia coli , Salmonella typhimurium , Caulobacter crescentusและVibrio alginolyticusเส้นใยประกอบด้วยโปรโตฟิลาเมนต์ 11 เส้นที่ขนานกับแกนของเส้นใยโดยประมาณ โปรโตฟิลาเมนต์แต่ละเส้นเป็นชุดของสายโปรตีนที่เรียงต่อกัน อย่างไรก็ตามCampylobacter jejuniมีโปรโตฟิลาเมนต์เจ็ดเส้น[ 25 ]
โครงสร้างฐานของแฟลเจลลามีลักษณะหลายอย่างที่คล้ายคลึงกับรูหลั่งสาร บางชนิด เช่น "ปลั๊ก" รูปทรงแท่งกลวงตรงกลางที่ยื่นออกมาผ่านเยื่อหุ้มเซลล์ ความคล้ายคลึงกันระหว่างโครงสร้างและโปรตีนของแฟลเจลลาแบคทีเรียและระบบหลั่งสารของแบคทีเรียเป็นหลักฐานทางวิทยาศาสตร์ที่สนับสนุนทฤษฎีที่ว่าแฟลเจลลาแบคทีเรียวิวัฒนาการมาจากระบบหลั่งสารประเภทที่สาม (TTSS)
โครงสร้างอะตอมของทั้งแฟลเจลลาของแบคทีเรียและอินเจกติโซม ของ TTSS ได้รับการอธิบายอย่างละเอียด โดยเฉพาะอย่างยิ่งด้วยการพัฒนาของกล้องจุลทรรศน์อิเล็กตรอนแบบไครโอส่วนที่เข้าใจได้ดีที่สุดคือส่วนที่อยู่ระหว่างเยื่อหุ้ม ชั้นในและชั้นนอก นั่นคือ วงแหวนโครงสร้างของเยื่อหุ้มชั้นใน (IM) คู่โครงสร้างของเยื่อหุ้มชั้นนอก (OM) และส่วนแท่ง/เข็ม (อินเจกติโซม) หรือแท่ง/ตะขอ (แฟลเจลลัม) [ 26 ]
มอเตอร์

แฟลเจลลัมของแบคทีเรียถูกขับเคลื่อนด้วยเครื่องยนต์หมุน ( คอมเพล็กซ์ Mot ) ที่ประกอบด้วยโปรตีน ซึ่งตั้งอยู่ที่จุดยึดของแฟลเจลลัมบนเยื่อหุ้มเซลล์ชั้นใน เครื่องยนต์นี้ขับเคลื่อนด้วยแรงขับเคลื่อนโปรตอนกล่าวคือ ด้วยการไหลของโปรตอน (ไอออนไฮโดรเจน) ข้ามเยื่อหุ้มเซลล์ของแบคทีเรียเนื่องจากความแตกต่างของความเข้มข้นที่สร้างขึ้นโดยกระบวนการเผาผลาญของเซลล์ ( สายพันธุ์ Vibrioมีแฟลเจลลัมสองชนิด คือ ด้านข้างและขั้ว และบางชนิดถูกขับเคลื่อนด้วยปั๊มไอออน โซเดียม แทนที่จะเป็นปั๊มโปรตอน[ 28 ] ) โรเตอร์จะขนส่งโปรตอนข้ามเยื่อหุ้มเซลล์และหมุนไปในกระบวนการ โรเตอร์เพียงอย่างเดียวสามารถทำงานได้ที่ 6,000 ถึง 100,000 รอบต่อนาที[ 29 ]แต่เมื่อมีเส้นใยแฟลเจลลัมติดอยู่ มักจะถึงเพียง 200 ถึง 1000 รอบต่อนาทีเท่านั้น ทิศทางการหมุนสามารถเปลี่ยนแปลงได้เกือบจะในทันทีโดยสวิตช์มอเตอร์แฟลเจลลัมซึ่งเกิดจากการเปลี่ยนแปลงเล็กน้อยในตำแหน่งของโปรตีนFliGในโรเตอร์[ 30 ]หน่วยสร้างแรงบิดคือสเตเตอร์ ซึ่งเป็นมอเตอร์หมุนที่ขับเคลื่อนด้วยโปรตอนซึ่งขับเคลื่อนมอเตอร์แฟลเจลลาหลักของแบคทีเรีย จากสเตเตอร์ การหมุนของเพนทาเมอร์ MotA5 รอบไดเมอร์ MotB2 จะยึดติดกับผนังเซลล์ซึ่งเชื่อมโยงโดยตรงกับการหมุนของวงแหวน C ดังนั้น มอเตอร์สเตเตอร์สร้างแรงบิดแต่ละตัวและโรเตอร์มอเตอร์แฟลเจลลาจึงทำหน้าที่เหมือนเฟืองที่ขบกันด้วยอัตราส่วนเกียร์ 6.2 [ 31 ] [ 32 ]มีการสังเกตพบมอเตอร์สเตเตอร์ระหว่าง 11 ถึง 16 ตัวที่สัมพันธ์โดยตรงกับโรเตอร์มอเตอร์แฟลเจลลาในแบคทีเรียต่างๆ[ 33 ]แรงบิดจะถูกถ่ายโอนจาก MotAB ไปยังเกลียวแรงบิดบนโดเมน D5 ของ FliG และเมื่อความต้องการแรงบิดหรือความเร็วเพิ่มขึ้น จะมีการใช้ MotAB มากขึ้น[ 27 ]เนื่องจากมอเตอร์แฟลเจลลัมไม่มีสวิตช์เปิด-ปิด โปรตีน epsE จึงถูกใช้เป็นคลัตช์เชิงกลเพื่อแยกมอเตอร์ออกจากโรเตอร์ ทำให้แฟลเจลลัมหยุดทำงานและช่วยให้แบคทีเรียคงอยู่ในตำแหน่งเดิม[ 34 ]
| ส่วนหนึ่งของชุดบทความเกี่ยวกับ |
| การเคลื่อนที่ของจุลินทรีย์และไมโครบอท |
|---|
 |
| ไมโครสวิมเมอร์ |
|
การผลิตและการหมุนของแฟลเจลลัมอาจใช้พลังงานมากถึง 10% ของ งบประมาณพลังงานของเซลล์ Escherichia coliซึ่งให้ประโยชน์ในระยะยาว เนื่องจากสิ่งใดก็ตามที่ก่อให้เกิดข้อเสียเปรียบด้านความเหมาะสม อาจถูกกำจัดออกไปเมื่อเวลาผ่านไป[ 22 ]การทำงานของมันก่อให้เกิดอนุมูลออกซิเจนที่ทำให้เกิดอัตราการกลายพันธุ์สูงขึ้น[ 22 ]
รูปทรงกระบอกของแฟลเจลลานั้นเหมาะสมกับการเคลื่อนที่ของสิ่งมีชีวิตขนาดเล็ก สิ่งมีชีวิตเหล่านี้ทำงานที่เลขเรย์โนลด์ ต่ำ ซึ่งความหนืดของน้ำโดยรอบมีความสำคัญมากกว่ามวลหรือความเฉื่อย[ 35 ]
ความเร็วในการหมุนของแฟลเจลลาจะแปรผันตามความเข้มของแรงขับเคลื่อนโปรตอน ซึ่งทำให้สามารถควบคุมความเร็วได้ในบางรูปแบบ และยังทำให้แบคทีเรียบางชนิดมีความเร็วที่น่าทึ่งเมื่อเทียบกับขนาดของมัน บางชนิดมีความเร็วประมาณ 60 เท่าของความยาวเซลล์ต่อวินาที ด้วยความเร็วเช่นนี้ แบคทีเรียจะใช้เวลาประมาณ 245 วันในการเดินทาง 1 กิโลเมตร แม้ว่าอาจดูช้า แต่แนวคิดจะเปลี่ยนไปเมื่อมีการนำแนวคิดเรื่องขนาดเข้ามาพิจารณา เมื่อเปรียบเทียบกับสิ่งมีชีวิตขนาดใหญ่แล้ว ถือว่าเร็วมากเมื่อแสดงในแง่ของจำนวนความยาวลำตัวต่อวินาที ตัวอย่างเช่น เสือชีตาห์มีความเร็วเพียงประมาณ 25 เท่าของความยาวลำตัวต่อวินาที[ 36 ]
แบคทีเรียสามารถเคลื่อนที่อย่างรวดเร็วไปยังสารดึงดูดและหนีจากสารขับไล่โดยใช้แฟลเจลลา โดยอาศัยการเดินแบบสุ่มที่มีอคติโดยการวิ่งและการหมุนตัวเกิดขึ้นจากการหมุนแฟลเจลลัมทวนเข็ม นาฬิกา และ ตาม เข็มนาฬิกาตามลำดับ ทิศทางการหมุนทั้งสองไม่เหมือนกัน (ในแง่ของการเคลื่อนที่ของแฟลเจลลัม) และถูกเลือกโดยสวิตช์ระดับโมเลกุล[ 37 ]การหมุนตามเข็มนาฬิกาเรียกว่าโหมดแรงดึงโดยที่ตัวแบคทีเรียจะตามแฟลเจลลัมไป การหมุนทวนเข็มนาฬิกาเรียกว่าโหมดแรงขับโดยที่แฟลเจลลัมจะล้าหลังตัวแบคทีเรีย[ 38 ]
การประกอบ
ระหว่างการประกอบแฟลเจลลัม ส่วนประกอบของแฟลเจลลัมจะผ่านแกนกลวงของฐานและเส้นใยที่กำลังก่อตัว ระหว่างการประกอบ ส่วนประกอบโปรตีนจะถูกเพิ่มที่ปลายแฟลเจลลัมแทนที่จะเป็นที่ฐาน[ 39 ]ในหลอดทดลองเส้นใยแฟลเจลลัมจะประกอบขึ้นเองในสารละลายที่มีแฟลเจลลินบริสุทธิ์เป็นโปรตีนเพียงชนิดเดียว[ 40 ]
วิวัฒนาการ
อย่างน้อย 10 ส่วนประกอบโปรตีนของแฟลเจลลัมของแบคทีเรียมีโปรตีนที่คล้ายคลึงกันกับระบบการหลั่งแบบที่สาม (T3SS) ที่พบในแบคทีเรียแกรมลบหลายชนิด[ 41 ]ดังนั้นจึงเป็นไปได้ว่าระบบหนึ่งวิวัฒนาการมาจากอีกระบบหนึ่ง เนื่องจาก T3SS มีจำนวนส่วนประกอบใกล้เคียงกับอุปกรณ์แฟลเจลลัม (ประมาณ 25 โปรตีน) จึงเป็นการยากที่จะระบุว่าระบบใดวิวัฒนาการมาก่อน อย่างไรก็ตาม ระบบแฟลเจลลัมดูเหมือนจะมีโปรตีนโดยรวมมากกว่า รวมถึงตัวควบคุมและชาเปอโรนต่างๆ ดังนั้นจึงมีการโต้แย้งว่าแฟลเจลลัมวิวัฒนาการมาจาก T3SS อย่างไรก็ตาม ยังมีข้อเสนอแนะอีกด้วย[ 42 ]ว่าแฟลเจลลัมอาจวิวัฒนาการมาก่อนหรือโครงสร้างทั้งสองอาจวิวัฒนาการไปพร้อมๆ กัน ความต้องการ การเคลื่อนที่ (ความคล่องตัว) ของสิ่งมีชีวิตเซลล์เดียวในยุคแรกๆ สนับสนุนว่าแฟลเจลลาที่เคลื่อนที่ได้คล่องตัวกว่าจะถูกคัดเลือกโดยวิวัฒนาการก่อน [ 42 ]แต่ T3SS ที่วิวัฒนาการมาจากแฟลเจลลัมสามารถมองได้ว่าเป็น 'วิวัฒนาการแบบลดทอน' และไม่ได้รับการสนับสนุนทางโทโพโลยีจากแผนภูมิวิวัฒนาการ[ 43 ]สมมติฐานที่ว่าโครงสร้างทั้งสองวิวัฒนาการแยกจากกันจากบรรพบุรุษร่วมกัน อธิบายถึงความคล้ายคลึงกันของโปรตีนระหว่างโครงสร้างทั้งสอง รวมถึงความหลากหลายทางหน้าที่ของพวกมันด้วย[ 44 ]
แฟลเจลลาและขบวนการออกแบบอัจฉริยะ
ผู้เขียนบางคนโต้แย้งว่าแฟลเจลลาไม่สามารถวิวัฒนาการได้ โดยสันนิษฐานว่าแฟลเจลลาจะทำงานได้อย่างถูกต้องก็ต่อเมื่อโปรตีนทั้งหมดอยู่ในตำแหน่งที่ถูกต้องเท่านั้น กล่าวอีกนัยหนึ่งคือ อุปกรณ์แฟลเจลลาเป็น " โครงสร้างที่ซับซ้อนอย่างไม่อาจลดทอนได้ " [ 45 ]อย่างไรก็ตาม โปรตีนหลายชนิดสามารถถูกลบหรือกลายพันธุ์ได้ และแฟลเจลลัมก็ยังคงทำงานได้ แม้บางครั้งจะมีประสิทธิภาพลดลง[ 46 ]ยิ่งไปกว่านั้น ด้วยโปรตีนหลายชนิดที่มีลักษณะเฉพาะในบางสายพันธุ์ ความหลากหลายขององค์ประกอบแฟลเจลลาของแบคทีเรียจึงสูงกว่าที่คาดไว้[ 47 ] ดังนั้น อุปกรณ์แฟลเจลลาจึงมีความยืดหยุ่นมากในแง่ของวิวัฒนาการ และสามารถสูญเสียหรือได้รับส่วนประกอบของโปรตีนได้อย่างสมบูรณ์ ตัวอย่างเช่น พบการกลายพันธุ์จำนวนหนึ่งที่เพิ่มความสามารถในการเคลื่อนที่ของ E. coli [ 48 ]หลักฐานเพิ่มเติมสำหรับการวิวัฒนาการของแฟลเจลลาของแบคทีเรีย ได้แก่ การมีอยู่ของแฟลเจลลาที่เหลืออยู่ รูปแบบแฟลเจลลาขั้นกลาง และรูปแบบความคล้ายคลึงกันในลำดับโปรตีนแฟลเจลลา รวมถึงการสังเกตว่าโปรตีนแฟลเจลลาหลักเกือบทั้งหมดมีความคล้ายคลึงกับโปรตีนที่ไม่ใช่แฟลเจลลา[ 41 ]นอกจากนี้ ยังมีการระบุถึงกระบวนการหลายอย่างที่มีบทบาทสำคัญในการวิวัฒนาการของแฟลเจลลา ซึ่งรวมถึงการประกอบตัวเองของหน่วยย่อยที่ซ้ำกันอย่างง่าย การจำลองยีนพร้อมกับการแยกตัวในภายหลัง การดึงเอาองค์ประกอบจากระบบอื่นมาใช้ ('การประกอบโมเลกุล') และการรวมตัวกันใหม่[ 49 ]
การจัดเรียงแฟลเจลลา
แบคทีเรียสายพันธุ์ต่าง ๆ มีจำนวนและการจัดเรียงของแฟลเจลลาที่แตกต่างกัน[ 50 ] [ 51 ]ซึ่งตั้งชื่อโดยใช้คำว่าtrichoจากภาษากรีกtrichosที่แปลว่าขน[ 52 ]
- แบคทีเรีย โมโนทริคัสเช่นVibrio cholerae มี แฟลเจลลัมขั้วเดียว[ 53 ]
- แบคทีเรียแอม ฟิทริคัสมีแฟลเจลลัมเพียงเส้นเดียวที่ปลายทั้งสองข้างตรงข้ามกัน (เช่นCampylobacter jejuniหรือAlcaligenes faecalis ) โดยแฟลเจลลัมทั้งสองจะหมุนแต่ทำงานประสานกันเพื่อสร้างแรงผลักดันที่สอดคล้องกัน
- แบคทีเรียLophotrichous ( lophoเป็นคำผสมภาษากรีกที่หมายถึงยอดหรือกระจุก ) [ 54 ]มีแฟลเจลลาหลายเส้นอยู่ที่จุดเดียวกันบนพื้นผิวของแบคทีเรีย เช่นHelicobacter pylori ซึ่งทำงานร่วมกันเพื่อขับเคลื่อนแบคทีเรียไปในทิศทางเดียว ในหลายกรณี ฐานของแฟลเจลลาหลายเส้นจะถูกล้อมรอบด้วยบริเวณพิเศษของเยื่อหุ้มเซลล์ ที่เรียกว่าออร์แกเนลล์ขั้ว
- แบคทีเรียชนิด เพอริทริคัสจะมีแฟลเจลลาที่ยื่นออกไปในทุกทิศทาง (เช่นอี. โคไล )
ในทางตรงกันข้าม สไปโรเคตมีแฟลเจลลาที่เรียกว่าเอนโดแฟลเจล ลา ซึ่งเกิดขึ้นจากขั้วตรงข้ามของเซลล์ และตั้งอยู่ในช่องว่างเพริพลาส มิก ดังที่แสดงโดยการแตกของเยื่อหุ้มชั้นนอกและโดยกล้องจุลทรรศน์อิเล็กตรอนไครโอโทโมกรา ฟี [ 55 ]การหมุนของเส้นใยเมื่อเทียบกับตัวเซลล์ทำให้แบคทีเรียทั้งหมดเคลื่อนที่ไปข้างหน้าในลักษณะคล้ายเกลียว แม้กระทั่งผ่านวัสดุที่มีความหนืดมากพอที่จะป้องกันการผ่านของแบคทีเรียที่มีแฟลเจลลาตามปกติ
ในSelenomonas บางชนิดที่มีขนาดใหญ่ จะมีแฟลเจลลามากกว่า 30 เส้นเรียงตัวอยู่นอกตัวเซลล์ โดยพันกันเป็นเกลียว形成โครงสร้างหนา (มองเห็นได้ง่ายด้วยกล้องจุลทรรศน์แบบใช้แสง) เรียกว่า " ฟาสซิเคิล "
ในVibrio spp. บางชนิด (โดยเฉพาะVibrio parahaemolyticus [ 56 ] ) และแบคทีเรีย ที่เกี่ยวข้อง เช่นAeromonasจะมีระบบแฟลเจลลาสองระบบอยู่ร่วมกัน โดยใช้ชุดยีนที่แตกต่างกันและระดับความเข้มข้นของไอออนที่แตกต่างกันสำหรับพลังงาน แฟลเจลลาขั้วจะถูกแสดงออกอย่างต่อเนื่องและให้การเคลื่อนที่ในของเหลวปริมาณมาก ในขณะที่แฟลเจลลาด้านข้างจะถูกแสดงออกเมื่อแฟลเจลลาขั้วพบกับแรงต้านมากเกินไปจนไม่สามารถหมุนได้[ 57 ] [ 58 ] [ 59 ] [ 60 ] [ 61 ] [ 62 ]สิ่งเหล่านี้ให้การเคลื่อนที่แบบรวมกลุ่มบนพื้นผิวหรือในของเหลวหนืด
การรวมกลุ่ม
การรวมกลุ่ม (Bundling) เป็นปรากฏการณ์ในเซลล์ที่มีแฟลเจลลาหลายเส้น โดยแฟลเจลลาจะรวมกลุ่มกันและหมุนอย่างพร้อมเพรียงกันเพื่อช่วยในการเคลื่อนที่ แฟลเจลลาเหล่านี้มีลักษณะโครงสร้างเป็นเกลียวซ้าย จะเรียงตัวและหมุนเป็นหน่วยเดียวกันเมื่อตัวหมุนโมเลกุลหมุนทวนเข็มนาฬิกา ทำให้เซลล์เคลื่อนที่ไปข้างหน้าผ่านสภาพแวดล้อม เมื่อตัวหมุนกลับทิศทางเป็นตามเข็มนาฬิกา แฟลเจลลาจะคลายตัวออกจากกลุ่ม ทำให้การเคลื่อนที่ที่ประสานกันหยุดชะงัก การคลายตัวนี้กระตุ้นพฤติกรรมที่เรียกว่าการพลิกตัว (Tumbling) ซึ่งเซลล์จะหยุดการเคลื่อนที่ไปข้างหน้าและกระตุกหรือสั่นอยู่กับที่ การพลิกตัวส่งผลให้เซลล์เปลี่ยนทิศทางแบบสุ่ม ซึ่งเป็นการเปลี่ยนทิศทางการว่ายน้ำไปข้างหน้าในครั้งต่อไป กระบวนการนี้มีความสำคัญอย่างยิ่งสำหรับแบคทีเรีย เช่นเอสเชอริเชีย โคไลทำให้พวกมันสามารถนำทางในสภาพแวดล้อมที่ซับซ้อนได้โดยการสลับระหว่างการว่ายน้ำแบบมีทิศทางและการเปลี่ยนทิศทางแบบสุ่ม กลไกของการรวมกลุ่มและการพลิกตัวเน้นให้เห็นถึงกลยุทธ์การเคลื่อนที่ที่ซับซ้อนของจุลินทรีย์ ทำให้พวกมันสามารถตอบสนองต่อสิ่งเร้าจากสิ่งแวดล้อมได้ พฤติกรรมนี้ได้รับการศึกษาอย่างละเอียดในกระบวนการเคโมแท็กซิสของแบคทีเรีย ซึ่งเซลล์จะปรับเส้นทางการเคลื่อนที่ของตนเองไปยังสภาวะที่เหมาะสม
ในแบบจำลองที่อธิบายถึงเคโมแท็กซิส ("การเคลื่อนที่อย่างมีจุดประสงค์") การหมุนตามเข็มนาฬิกาของแฟลเจลลัมจะถูกยับยั้งโดยสารประกอบทางเคมีที่เป็นประโยชน์ต่อเซลล์ (เช่น อาหาร) เมื่อเคลื่อนที่ไปในทิศทางที่เหมาะสม ความเข้มข้นของสารดึงดูดทางเคมีดังกล่าวจะเพิ่มขึ้น ดังนั้นการหมุนจึงถูกยับยั้งอย่างต่อเนื่อง ทำให้เซลล์เคลื่อนที่ไปข้างหน้าได้ ในทำนองเดียวกัน เมื่อทิศทางการเคลื่อนที่ของเซลล์ไม่เอื้ออำนวย (เช่น เคลื่อนที่ออกห่างจากสารดึงดูดทางเคมี) การหมุนจะไม่ถูกยับยั้งอีกต่อไปและเกิดขึ้นบ่อยขึ้นมาก โดยมีโอกาสที่เซลล์จะปรับทิศทางไปในทิศทางที่ถูกต้องได้
แม้ว่าแฟลเจลลาทั้งหมดจะหมุนตามเข็มนาฬิกา แต่บ่อยครั้งที่ไม่สามารถรวมตัวกันเป็นมัดได้เนื่องจากเหตุผลทางเรขาคณิตและอุทกพลศาสตร์[ 63 ] [ 64 ]
กลไกโมเลกุลของการสลับการหมุน
มอเตอร์แฟลเจลลาประกอบด้วยโปรตีนทั้งหมด สเตเตอร์เป็นหน่วยสร้างแรงบิดและเป็นมอเตอร์หมุนที่ขับเคลื่อนด้วยโปรตอนซึ่งขับเคลื่อนมอเตอร์แฟลเจลลาหลักของแบคทีเรีย จากสเตเตอร์ การหมุนของเพนทาเมอร์ MotA5 รอบไดเมอร์ MotB2 จะยึดติดอยู่กับผนังเซลล์เหมือนล้อเฟือง ซึ่งเชื่อมต่อโดยตรงกับการหมุนของวงแหวน C และด้วยเหตุนี้จึงทำหน้าที่เป็นตัวสร้างแรงโดยการโต้ตอบกับวงแหวน C ของโรเตอร์[ 31 ]เนื่องจากวงแหวน C สามารถหมุนได้สองทิศทาง ในขณะที่คอมเพล็กซ์สเตเตอร์เองหมุนได้ทิศทางเดียว วงแหวน C จึงต้องมีการจัดเรียงภายในใหม่ ในระหว่างการจัดเรียงใหม่นี้ วงแหวน C จะไม่แตกสลายและสร้างใหม่ แต่โดเมนโปรตีนจะมีการเปลี่ยนแปลงโครงสร้างที่ประสานกัน[ 65 ]
วงแหวน C มีตำแหน่งสำหรับจับสัญญาณสวิตช์ เช่น โปรตีน CheY ที่ถูกฟอสฟอริเลต ซึ่งจะเริ่มต้นกระบวนการจัดเรียงตัวภายในใหม่หลายขั้นตอน โดเมนโปรตีนหลายโดเมนภายในวงแหวน C จะมีการจัดเรียงตัวใหม่ รวมถึงโดเมน FliGN และ FliGM ซึ่งหมุนและเปลี่ยนตำแหน่งไปพร้อมกับโดเมน FliGC การพลิกกลับของโดเมนที่ประสานกันนี้ทำให้สเตเตอร์เปลี่ยนตำแหน่งสัมพันธ์กับวงแหวน C เมื่อสเตเตอร์ออกแรงจากด้านนอกของวงแหวน C โรเตอร์จะหมุนทวนเข็มนาฬิกา (การรวมกลุ่ม) เมื่อสเตเตอร์เปลี่ยนตำแหน่งเพื่อออกแรงกับด้านในของวงแหวน C โรเตอร์จะหมุนตามเข็มนาฬิกา (การพลิกคว่ำ) แม้ว่ากลไกการหมุนของสเตเตอร์จะคงที่ แต่ทิศทางที่สเตเตอร์ออกแรงกับวงแหวน C จะกำหนดทิศทางการหมุนของโรเตอร์ ผลการค้นพบเหล่านี้อิงตามโครงสร้างของมอเตอร์แฟลเจลลาของ Salmonella Typhimuriumแม้ว่าส่วนประกอบมอเตอร์หลักจะได้รับการอนุรักษ์ไว้อย่างดีในสายพันธุ์แบคทีเรีย แต่ก็ยังไม่ชัดเจนว่ากลไกการสลับเฉพาะนี้ได้รับการอนุรักษ์ไว้โดยทั่วไปหรือไม่ หรือว่าสายพันธุ์แบคทีเรียที่แตกต่างกันใช้กลยุทธ์นี้ในรูปแบบที่แตกต่างกัน [ 65 ]
แฟลเจลลาของยูคาริโอต

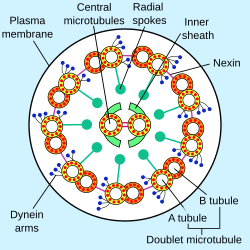

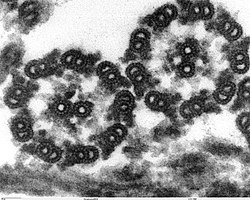
ศัพท์เฉพาะ
เพื่อเน้นย้ำความแตกต่างระหว่างแฟลเจลลาของแบคทีเรียและซิเลียและแฟลเจลลาของยูคาริโอต ผู้เขียนบางคนพยายามแทนที่ชื่อของโครงสร้างยูคาริโอตทั้งสองนี้ด้วย " อันดูลิโพเดีย " (เช่น บทความทั้งหมดของMargulisตั้งแต่ปี 1970 เป็นต้นมา) [ 66 ]หรือ "ซิเลีย" สำหรับทั้งสอง (เช่น Hülsmann, 1992; [ 67 ] Adl et al., 2012; [ 68 ]บทความส่วนใหญ่ของCavalier-Smith ) โดยคงคำว่า "แฟลเจลลา" ไว้สำหรับโครงสร้างของแบคทีเรีย อย่างไรก็ตาม การใช้คำว่า "ซิเลีย" และ "แฟลเจลลา" เพื่อแยกแยะยูคาริโอตที่นำมาใช้ในบทความนี้ (ดู§ แฟลเจลลาเทียบกับซิเลียด้านล่าง) ยังคงเป็นเรื่องปกติ (เช่น Andersen et al., 1991; [ 69 ] Leadbeater et al., 2000) [ 70 ]
โครงสร้างภายใน
แกนกลางของแฟลเจลลัมในยูคาริโอต หรือที่เรียกว่าแอ็กโซนีมคือกลุ่มของไมโครทิวบูล 9 คู่ที่เชื่อมต่อกัน ( เรียกว่าดับเบิล เล็ต) ล้อมรอบไมโครทิวบูลเดี่ยว 2 เส้นตรงกลาง ( ซิงเกิลเล็ต ) แอ็กโซนีมแบบ 9+2 นี้ เป็นลักษณะเฉพาะของแฟลเจลลัมในยูคาริโอต ที่ฐานของแฟลเจลลัมในยูคาริโอตคือเบซัลบอดี "เบลฟาโรพลาสต์" หรือคิเนโทโซม ซึ่งเป็นศูนย์กลางการจัดระเบียบไมโครทิวบูลของแฟลเจลลัม และมีความยาวประมาณ 500 นาโนเมตร โครงสร้างของเบซัลบอดีเหมือนกับเซนทริโอล แฟ ลเจ ลลัมถูกห่อหุ้มด้วยเยื่อหุ้มเซลล์ ทำให้ภายในของแฟลเจลลัมสามารถเข้าถึงได้โดย ไซโตพลาสซึมของ เซลล์
นอกจากแอ็กโซนีมและฐานร่างกายซึ่งมีรูปร่างค่อนข้างคงที่แล้ว โครงสร้างภายในอื่นๆ ของอุปกรณ์แฟลเจลลา ได้แก่ โซนเปลี่ยนผ่าน (บริเวณที่แอ็กโซนีมและฐานร่างกายมาบรรจบกัน) และระบบราก (โครงสร้างไมโครทิวบูลาร์หรือไฟบริลาร์ที่ยื่นออกมาจากฐานร่างกายเข้าไปในไซโตพลาสซึม) ซึ่งมีความแปรผันมากกว่าและมีประโยชน์ในฐานะตัวบ่งชี้ความสัมพันธ์ทางวิวัฒนาการของยูคาริโอต โครงสร้างอื่นๆ ที่พบได้น้อยกว่า ได้แก่ แท่งพาราแฟลเจลลาร์ (หรือพาราแอ็กเซียล พาราแอ็กโซนีม) เส้นใย R และเส้นใย S [ 71 ] : 63–84 สำหรับโครงสร้างพื้นผิว โปรดดูด้านล่าง
กลไก
ไมโครทูบูลคู่ 9 คู่ด้านนอกแต่ละคู่จะยื่น แขน ไดเนอิน (แขน "ด้านใน" และแขน "ด้านนอก") ไปยังไมโครทูบูลที่อยู่ติดกัน ซึ่งจะสร้างแรงผ่านการไฮโดรไลซิสของ ATP แกนแอกโซนีมของแฟลเจลลายังประกอบด้วยซี่รัศมีซึ่งเป็นคอมเพล็กซ์โพลีเปปไทด์ที่ยื่นออกมาจากไมโครทูบูลคู่ 9 คู่ด้านนอกแต่ละคู่ไปยังคู่กลาง โดยที่ "หัว" ของซี่รัศมีหันเข้าด้านใน เชื่อกันว่าซี่รัศมีมีส่วนเกี่ยวข้องในการควบคุมการเคลื่อนที่ของแฟลเจลลา แม้ว่าหน้าที่และวิธีการทำงานที่แน่นอนของมันยังไม่เป็นที่เข้าใจ[ 72 ]
แฟลเจลลาเทียบกับซิเลีย

รูปแบบการเต้นปกติของซีเลียและแฟลเจลลาของยูคาริโอตสร้างการเคลื่อนไหวในระดับเซลล์ ตัวอย่างมีตั้งแต่การขับเคลื่อนของเซลล์เดี่ยว เช่น การว่ายน้ำของสเปิร์มไปจนถึงการขนส่งของเหลวไปตามชั้นเซลล์ที่อยู่กับที่ เช่น ในทางเดินหายใจ [ 73 ]
แม้ว่าซีเลียและแฟลเจลลาของยูคาริโอตจะเหมือนกันในที่สุด แต่บางครั้งก็มีการจัดประเภทตามรูปแบบการเคลื่อนไหว ซึ่งเป็นธรรมเนียมปฏิบัติมาตั้งแต่ก่อนที่จะทราบโครงสร้างของพวกมัน ในกรณีของแฟลเจลลา การเคลื่อนไหวมักจะเป็นระนาบและเป็นคลื่น ในขณะที่ซีเลียที่เคลื่อนไหวได้มักจะเคลื่อนไหวแบบสามมิติที่ซับซ้อนกว่า โดยมีจังหวะการออกแรงและการคืนตัว[ 73 ]อีกรูปแบบหนึ่งของการจำแนกแบบดั้งเดิมคือจำนวนออร์แกเนลล์ 9+2 บนเซลล์[ 72 ]
การขนส่งภายในแฟลเจลลัม
การขนส่งภายในแฟลเจลลัมซึ่งเป็นกระบวนการที่ซับยูนิตแอ็กโซเนมตัวรับทรานส์เมมเบรนและโปรตีนอื่นๆ เคลื่อนที่ขึ้นและลงตามความยาวของแฟลเจลลัม เป็นสิ่งจำเป็นสำหรับการทำงานที่เหมาะสมของแฟลเจลลัม ทั้งในด้านการเคลื่อนที่และการส่งสัญญาณ[ 74 ]
วิวัฒนาการและการเกิดขึ้น
แฟลเจลลาหรือซิเลียของยูคาริโอต ซึ่งอาจเป็นลักษณะดั้งเดิม[ 75 ]แพร่หลายในเกือบทุกกลุ่มของยูคาริโอต ในรูปแบบที่ค่อนข้างถาวร หรือในรูปแบบระยะวงจรชีวิตที่มีแฟลเจลลา (เช่นซอยด์แกมีตซูโอสปอร์ซึ่งอาจผลิตอย่างต่อเนื่องหรือไม่ก็ได้) [ 76 ] [ 77 ] [ 68 ]
สถานการณ์แรกพบได้ในเซลล์เฉพาะของสิ่งมีชีวิตหลายเซลล์ (เช่นโคอาโนไซต์ของฟองน้ำหรือเยื่อบุผิว ที่มีขน ของสัตว์หลายเซลล์ ) เช่นเดียวกับในซีลิเอตและยูคาริโอตจำนวนมากที่มี "สภาวะแฟลเจลเลต" (หรือ " ระดับการจัดระเบียบ แบบโมนาดอยด์ " ดูFlagellataซึ่งเป็นกลุ่มที่สร้างขึ้น)
ระยะวงจรชีวิตที่มีแฟลเจลลาพบได้ในหลายกลุ่ม เช่นสาหร่ายสีเขียว หลายชนิด (ซูโอสปอร์และแกมีตตัวผู้), ไบ รโอไฟต์ (แกมีตตัวผู้), เทอริโดไฟต์ (แกมีตตัวผู้), พืช เมล็ดเปลือยบางชนิด( ไซแค ดและแปะก๊วยเป็นแกมีตตัวผู้), ไดอะตอม แบบเซนทริก (แกมีตตัวผู้), สาหร่ายสีน้ำตาล (ซูโอสปอร์และแกมีต), โอโอไมซีต (ซูโอ สปอร์และแกมีตแบบไม่อาศัยเพศ), ไฮโฟไคทริด (ซูโอสปอร์), ลาบิรินทูโลไมซีต ( ซูโอสปอร์), อะพิคอมเพล็ กซานบางชนิด (แกมีต), เรดิโอลาเรียนบางชนิด(น่าจะเป็นแกมีต), [ 78 ]ฟอรามินิเฟอแร น (แกมีต), พลาสโมดิโอโฟโรไมซีต (ซูโอสปอร์และแกมีต), ไมโซแกสท ริด (ซูโอสปอร์), เมตาโซแอน (แกมีตตัวผู้) และ เชื้อรา ไคทริด (ซูโอสปอร์และแกมีต)
แฟลเจลลาหรือซีเลียหายไปอย่างสมบูรณ์ในบางกลุ่ม อาจเนื่องมาจากการสูญเสียมากกว่าที่จะเป็น สภาวะ ดั้งเดิมการสูญเสียซีเลียเกิดขึ้นในสาหร่ายสีแดงสาหร่ายสีเขียวบางชนิด ( Zygnematophyceae ) พืชเมล็ด เปลือย ยกเว้นไซแคดและแปะก๊วยพืช ดอก ไดอะตอมเพนเนต อะพิคอม เพล็กซาน บางชนิดอะมีโบโซแอนบางชนิดในสเปิร์มของสัตว์ หลายเซลล์บาง ชนิด [ 79 ] และในเชื้อรา (ยกเว้นไคทริด )
ประเภท
มีการใช้คำศัพท์หลายคำที่เกี่ยวข้องกับแฟลเจลลาหรือซิเลียเพื่อจำแนกลักษณะของยูคาริโอต[ 77 ] [ 80 ] [ 71 ] : 60–63 [ 81 ] [ 82 ]ตามโครงสร้างพื้นผิวที่มีอยู่ แฟลเจลลาอาจเป็น:
- แฟลเจลลาแบบแส้ (= แฟลเจลลาเรียบ ไม่มีขน): ไม่มีขน เช่น ในOpisthokonta
- แฟลเจลลาที่มีขน (= แฟลเจลลาแบบมีขน, แฟลเจลลาแบบมีประกาย, แฟลเจลลาแบบพลูโรเนมาติก): มีขน (= มาสติโกนีมเซนซู ลาโต ) แบ่งออกเป็น:
- มีขนละเอียด (= ขนที่ไม่เป็นท่อ หรือขนแบบเรียบง่าย): พบในEuglenophyceae , DinoflagellataและHaptophyceae บางชนิด ( Pavlovales )
- มีขนแข็ง (= ขนรูปท่อ, เรโทรนีม, มาสติโกนีมในความหมายแคบ ) แบ่งออกเป็น:
- ขนแบบสองส่วน: มีสองบริเวณ พบในCryptophyceae , PrasinophyceaeและHeterokonta บางชนิด
- ขนไตรภาค (= straminipilous): ประกอบด้วยสามส่วน (ฐาน แกนกลาง และขนปลายสุดหนึ่งเส้นหรือมากกว่า) พบในHeterokonta ส่วนใหญ่
- แฟลเจลลาแบบสติโคนีมาติก: มีขนเรียงเป็นแถวเดียว
- แฟลเจลลาแบบแพนโทนีมาติก: มีขนสองแถว
- acronematic: แฟลเจลลาที่มีมาสติโกนีมหรือขนแฟลเจลลาเพียงเส้นเดียวที่ปลาย (เช่นโบโดนิด ) [ 83 ]ผู้เขียนบางคนใช้คำนี้เป็นคำพ้องความหมายของ whiplash
- มีเกล็ด เช่นPrasinophyceae
- มีหนาม: เช่นสาหร่ายสีน้ำตาล บางชนิด
- มีเยื่อหุ้มเซลล์เป็นคลื่น เช่นคิเนโตพลาส ติดบางชนิด และพาราบาซาลิด บางชนิด
- มีงวง (ส่วนที่ยื่นออกมาคล้ายลำต้นของเซลล์): เช่น อะปูโซโมนาดส์ บอโดนิดส์บางชนิด[ 84 ]
ตามจำนวนแฟลเจลลา เซลล์อาจเป็น: (โปรดจำไว้ว่าผู้เขียนบางคนใช้คำว่า "มีซีเลีย" แทนคำว่า "มีแฟลเจลลา") [ 68 ] [ 85 ]
- ไม่มีการติดธง: เช่นOpisthokonta ส่วนใหญ่
- มีแฟลเจลลาสองเส้น: เช่นไดโนแฟลเจลลาตา ทั้งหมด แกมีตของชาโรไฟซี ของไบรโอ ไฟต์ส่วนใหญ่และของเมตาโซแอน บางชนิด [ 79 ]
- มีแฟลเจลลา 3 เส้น: เช่น แกมีตของฟอรามินิเฟอรา บางชนิด
- มีแฟลเจลลาสี่เส้น: เช่นPrasinophyceae บางชนิด , Collodictyonidae
- มีแฟลเจลลา 8 เส้น: เช่นDiplomonada บางชนิด , Prasinophyceae บางชนิด
- มีแฟลเจลลาหลายเส้น เช่นOpalinata , Ciliophora , Stephanopogon , Parabasalida , Hemimastigophora , Caryoblastea , Multicilia , แกมีต (หรือซอยด์ ) ของOedogoniales ( Chlorophyta ), เฟิร์น บางชนิด และพืชเมล็ดเปลือย บางชนิด
ตามตำแหน่งการแทรกของแฟลเจลลา: [ 86 ]
- โอพิสโทคอนต์: เซลล์ที่มีแฟลเจลลาแทรกอยู่ทางด้านหลัง เช่น ในโอพิสโทคอนตา (วิสเชอร์, 1945) ในแฮปโทไฟซี แฟลเจลลาจะแทรกอยู่ทางด้านข้างหรือด้านปลาย แต่จะหันไปทางด้านหลังเมื่อว่ายน้ำอย่างรวดเร็ว[ 87 ]
- อะโครคอนต์: เซลล์ที่มีแฟลเจลลาอยู่บริเวณส่วนปลาย
- subakrokont: เซลล์ที่มีแฟลเจลลาแทรกอยู่ใต้ส่วนปลาย
- เพลอโรคอนต์: เซลล์ที่มีแฟลเจลลาแทรกอยู่ด้านข้าง
ตามรูปแบบการเต้น:
- การร่อน: แฟลเจลลัมที่ลากไปตามพื้นผิว[ 84 ]
- เฮเทอโรไดนามิก: แฟลเจลลาที่มีรูปแบบการตีที่แตกต่างกัน (โดยปกติแฟลเจลลัมหนึ่งทำหน้าที่ในการจับอาหารและอีกแฟลเจลลัมหนึ่งทำหน้าที่ในการร่อน การยึดเกาะ การขับเคลื่อน หรือ "การควบคุมทิศทาง") [ 88 ]
- ไอโซไดนามิก: แฟลเจลลาโบยบินด้วยรูปแบบเดียวกัน
คำศัพท์อื่นๆ ที่เกี่ยวข้องกับประเภทของแฟลเจลลา:
- ไอโซคอนต์: เซลล์ที่มีแฟลเจลลาความยาวเท่ากัน ในอดีตเคยใช้เรียกกลุ่มคลอโรไฟตา ด้วย
- อะนิโซคอนต์: เซลล์ที่มีแฟลเจลลาความยาวไม่เท่ากัน เช่นยูเกลโนไฟซีและพราซิโนไฟซี บางชนิด
- เฮเทอโรคอนต์ (Heterokonta): คำที่ลูเธอร์ (Luther, 1899) นำมาใช้เรียกกลุ่มสาหร่ายXanthophyceaeเนื่องจากมีแฟลเจลลาคู่หนึ่งที่มีความยาวไม่เท่ากัน ต่อมาคำนี้มีความหมายเฉพาะเจาะจงในการอ้างถึงเซลล์ที่มีแฟลเจลลาด้านหน้าเป็นแบบสตรามินิพิลัส (มีมาสติโกนีมแบบสามส่วน เรียงเป็นหนึ่งหรือสองแถว) และแฟลเจลลาด้านหลังมักจะเรียบ นอกจากนี้ยังใช้เรียกกลุ่มอนุกรมวิธานว่า เฮเทอโรคอนตา(Heterokonta ) ด้วย
- สเตฟาโนคอนต์ (stephanokont): เซลล์ที่มีกลุ่มแฟลเจลลาเรียงตัวเป็นมงกุฎอยู่ใกล้ส่วนปลายด้านหน้า เช่น แกมีตและสปอร์ของOedogoniales และ สปอร์ของBryopsidales บางชนิด คำนี้ถูกนำมาใช้โดย Blackman & Tansley (1902) เพื่อ ใช้เรียกOedogoniales
- akont: เซลล์ที่ไม่มีแฟลเจลลา นอกจากนี้ยังใช้เพื่ออ้างถึงกลุ่มอนุกรมวิธาน เช่น Aconta หรือ Akonta ได้แก่ZygnematophyceaeและBacillariophyceae (Oltmanns, 1904) หรือRhodophyceae (Christensen, 1962)
แฟลเจลลาของอาร์เคีย
อาร์เคเอลลัมที่พบในอาร์เคียบางสปีชีส์มีลักษณะคล้ายคลึงกับแฟลเจลลัมของแบคทีเรีย ในช่วงทศวรรษ 1980 เชื่อกันว่าทั้งสองชนิดมีความคล้ายคลึงกันโดยพิจารณาจากรูปร่างและพฤติกรรมโดย รวม [ 89 ]ทั้งแฟลเจลลาและอาร์เคเอลลาประกอบด้วยเส้นใยที่ยื่นออกมานอกเซลล์และหมุนเพื่อขับเคลื่อนเซลล์ แฟลเจลลาของอาร์เคียมีโครงสร้างที่เป็นเอกลักษณ์ซึ่งไม่มีช่องตรงกลาง คล้ายกับไพลินชนิด IV ของแบคทีเรีย โปรตีนของอาร์เคีย (อาร์เคเอลลิน) สร้างขึ้นจากเปปไทด์สัญญาณคลาส 3 และถูกประมวลผลโดยเอนไซม์คล้ายเพปติเดสพรีไพลินชนิด IV โดยทั่วไปอาร์เคเอลลินจะถูกดัดแปลงโดยการเพิ่มไกลแคนแบบ N-linked ซึ่งจำเป็นสำหรับการประกอบหรือการทำงานที่เหมาะสม[ 4 ]
การค้นพบในช่วงทศวรรษ 1990 เผยให้เห็นความแตกต่างโดยละเอียดมากมายระหว่างแฟลเจลลาของอาร์เคียและแบคทีเรีย ซึ่งรวมถึง:
- การหมุนของแฟลเจลลาของแบคทีเรียได้รับพลังงานจากแรงขับเคลื่อนโปรตอนซึ่งเป็นการไหลของไอออนH +หรือบางครั้งอาจได้รับพลังงานจากแรงขับเคลื่อนโซเดียมซึ่งเป็นการไหลของ ไอออน Na +การหมุนของแฟลเจลลาของอาร์เคียได้รับพลังงานจากATP [ 90 ]
- ในขณะที่เซลล์แบคทีเรียมักจะมีเส้นใยแฟลเจลลัมจำนวนมาก ซึ่งแต่ละเส้นจะหมุนอย่างอิสระ แต่แฟลเจลลัมของอาร์เคียนั้นประกอบด้วยมัดของเส้นใยจำนวนมากที่หมุนเป็นชุดเดียว (อ้างอิง? ดูเหมือนว่าจะไม่เป็นเช่นนั้นในPyrococcus furiosus [ 91 ] )
- แฟลเจลลาของแบคทีเรียเจริญเติบโตโดยการเพิ่มหน่วยย่อยแฟลเจลลินที่ปลาย ในขณะที่แฟลเจลลาของอาร์เคียเจริญเติบโตโดยการเพิ่มหน่วยย่อยที่ฐาน
- แฟลเจลลาของแบคทีเรียมีความหนากว่าของอาร์เคียลลา และเส้นใยของแบคทีเรียมี "ท่อ" กลวงขนาดใหญ่พอที่ซับยูนิตแฟลเจลลินจะไหลขึ้นไปภายในเส้นใยและถูกเพิ่มเข้าไปที่ปลาย ในขณะที่อาร์เคียลลามีขนาดบางเกินไป (12-15 นาโนเมตร) ที่จะทำให้เกิดสิ่งนี้ได้[ 92 ]
- ส่วนประกอบหลายอย่างของแฟลเจลลาแบคทีเรียมีความคล้ายคลึงกันในลำดับกับส่วนประกอบของระบบการหลั่งแบบประเภท IIIแต่ส่วนประกอบของแฟลเจลลาแบคทีเรียและอาร์เคียลลาไม่มีความคล้ายคลึงกันในลำดับ ในทางกลับกัน ส่วนประกอบบางอย่างของอาร์เคียลลามีความคล้ายคลึงกันในลำดับและรูปร่างกับส่วนประกอบของพิไลแบบประเภท IVซึ่งประกอบขึ้นผ่านการทำงานของระบบการหลั่งแบบประเภท II (การตั้งชื่อพิไลและระบบการหลั่งโปรตีนไม่สอดคล้องกัน) [ 92 ]
ความแตกต่างเหล่านี้สนับสนุนทฤษฎีที่ว่าแฟลเจลลาของแบคทีเรียและอาร์เคเอลลาเป็นกรณีคลาสสิกของความคล้ายคลึง ทางชีวภาพ หรือวิวัฒนาการแบบบรรจบกันมากกว่าความเหมือนกันทาง โครงสร้าง [ 93 ] [ 94 ] [ 95 ]การวิจัยเกี่ยวกับโครงสร้างของอาร์เคเอลลามีความก้าวหน้าอย่างมากตั้งแต่ช่วงต้นทศวรรษ 2010 โดยมีโครงสร้างความละเอียดระดับอะตอมแรกของโปรตีนอาร์เคเอลลา การค้นพบหน้าที่เพิ่มเติมของอาร์เคเอลลา และรายงานแรกของอาร์เคเอลลาใน Nanoarchaeota และ Thaumarchaeota [ 96 ] [ 97 ]
เชื้อรา
เชื้อราเพียงชนิดเดียวที่มีแฟลเจลลัมเดี่ยวบนสปอร์คือ เชื้อรากลุ่มไค ทริ ด ในBatrachochytrium dendrobatidisแฟลเจลลัมมีความยาว 19–20 μm [ 98 ]เซนทริโอลที่ไม่ทำงานอยู่ติดกับไคเนโตโซมแท่งเชื่อมต่อเก้าแท่ง ยึดไคเนโตโซมเข้ากับ พลาสมาเลมมาและมีแผ่นปลายอยู่ในโซนเปลี่ยนผ่าน มีการสังเกตโครงสร้างคล้ายวงแหวนด้านในที่ติดอยู่กับท่อของแฟลเจลลัมคู่ภายในโซนเปลี่ยนผ่านในส่วนตัดขวาง[ 98 ]
รูปภาพเพิ่มเติม
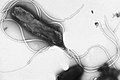 แฟลเจลลาหลายเส้นเรียงตัวแบบลอโฟทริคัสบนพื้นผิวของแบคทีเรีย Helicobacter pylori
แฟลเจลลาหลายเส้นเรียงตัวแบบลอโฟทริคัสบนพื้นผิวของแบคทีเรีย Helicobacter pylori แบบจำลองทางกายภาพของแฟลเจลลัมของแบคทีเรีย
แบบจำลองทางกายภาพของแฟลเจลลัมของแบคทีเรีย


ดูเพิ่มเติม
อ่านเพิ่มเติม
- Berg, Howard C. (มกราคม 2000). "พฤติกรรมการเคลื่อนที่ของแบคทีเรีย" . Physics Today . 53 (1): 24– 29. Bibcode : 2000PhT....53a..24B . doi : 10.1063/1.882934 . S2CID 178516210 .
- ลินเดมันน์, ชาร์ลส์ (4 เมษายน 2551). "กลไกการเคลื่อนที่ของอสุจิ" . มหาวิทยาลัยโอ๊คแลนด์. เก็บถาวรจากต้นฉบับเมื่อ 16 พฤษภาคม 2551. สืบค้นเมื่อ18 พฤษภาคม 2551 .
- Purcell, EM (1977). "ชีวิตที่เลขเรย์โนลด์ต่ำ" (PDF) . American Journal of Physics . 45 (1): 3– 11. Bibcode : 1977AmJPh..45....3P . doi : 10.1119/1.10903 . hdl : 2433/226838 . เก็บถาวรจากต้นฉบับ(PDF)เมื่อวันที่ 5 มิถุนายน 2011 . สืบค้นเมื่อ19 ตุลาคม 2009 .
- Matzke, NJ (10 พฤศจิกายน 2003). "วิวัฒนาการในอวกาศ (บราวน์เนียน): แบบจำลองสำหรับต้นกำเนิดของแฟลเจลลัมของแบคทีเรีย" talkdesign.org.
ลิงก์ภายนอก
- คลังภาพเซลล์ - แฟลเจลลา
![]() บทความนี้ได้นำข้อความจากสิ่งพิมพ์ที่อยู่ในสาธารณสมบัติ มาใช้ : Chambers, Ephraim , ed. (1728). Cyclopædia, or an Universal Dictionary of Arts and Sciences (1st ed.). James and John Knapton, et al.
บทความนี้ได้นำข้อความจากสิ่งพิมพ์ที่อยู่ในสาธารณสมบัติ มาใช้ : Chambers, Ephraim , ed. (1728). Cyclopædia, or an Universal Dictionary of Arts and Sciences (1st ed.). James and John Knapton, et al.{{cite encyclopedia}}: ข้อมูลหายไปหรือว่างเปล่า|title=( ขอความช่วยเหลือ )
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เฆี่ยน
แฟ ลเจลลัม ( / f l ə ˈ dʒ ɛ l əm / ; พหูพจน์ : flagella ) ( ภาษาละติน แปลว่า 'แส้' หรือ 'แส้') คือ ระยางค์ คล้ายเส้นผมที่ยื่นออกมาจาก เซลล์สเปิร์ม ของ พืช และสัตว์ บางชนิด จาก...
Types
The three types of flagella are bacterial, archaeal, and eukaryotic.
Bacterial flagella
การประกอบและการทำงานของแฟลเจลลาของแบคทีเรียต้องอาศัยโปรตีนมากกว่า 50 ชนิดที่ทำงานร่วมกัน [ 22 ] แฟลเจลลัมของแบคทีเรียประกอบด้วย หน่วยย่อย โปรตีน ของ แฟลเจลลิน [ 13 ] รูปร่าง ของมันคือท่อกลวงหนา 20 นาโนเมตร มี ลักษณะเป็นเกลียว...
แฟลเจลลาของยูคาริโอต
เพื่อเน้นย้ำความแตกต่างระหว่างแฟลเจลลาของแบคทีเรียและซิเลียและแฟลเจลลาของยูคาริโอต ผู้เขียนบางคนพยายามแทนที่ชื่อของโครงสร้างยูคาริโอตทั้งสองนี้ด้วย " อันดูลิโพเดีย " (เช่น บทความทั้งหมดของ Margulis ตั้งแต่ปี 1970 เป็นต้นมา) [ 66 ] หรือ "ซิเลีย" สำหรับทั้งสอง...