อ่าน 15 นาที
การควบคุมการแสดงออกของยีน
การควบคุมการแสดงออกของยีนหรือการควบคุมยีนครอบคลุมกลไกที่หลากหลายซึ่งเซลล์ใช้เพื่อเพิ่มหรือลดการผลิตผลิตภัณฑ์ยีน เฉพาะ ( โปรตีนหรือRNA ) โปรแกรมการแสดงออกของยีน ที่ซับซ้อน...
การควบคุมการแสดงออกของยีน

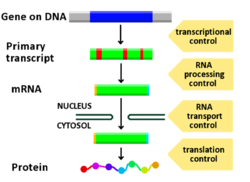
การควบคุมการแสดงออกของยีนหรือการควบคุมยีน[ 1 ]ครอบคลุมกลไกที่หลากหลายซึ่งเซลล์ใช้เพื่อเพิ่มหรือลดการผลิตผลิตภัณฑ์ยีน เฉพาะ ( โปรตีนหรือRNA ) โปรแกรมการแสดงออกของยีน ที่ซับซ้อน พบเห็นได้ทั่วไปในทางชีววิทยา เช่น เพื่อกระตุ้นเส้นทางการพัฒนา ตอบสนองต่อสิ่งเร้าจากสิ่งแวดล้อม หรือปรับตัวให้เข้ากับแหล่งอาหารใหม่ แทบทุกขั้นตอนของการแสดงออกของยีนสามารถถูกปรับเปลี่ยนได้ ตั้งแต่การเริ่มต้นการถอดรหัสไปจนถึงการประมวลผล RNAและ การดัดแปลง โปรตีนหลังการแปลรหัส บ่อยครั้งที่ตัวควบคุมยีนตัวหนึ่งควบคุมอีกตัวหนึ่ง และเป็นเช่นนั้น เรื่อย ไปใน เครือข่ายควบคุมยีน
การควบคุมยีนมีความสำคัญอย่างยิ่งต่อไวรัสโปรคาริโอตและยูคาริโอตเนื่องจากช่วยเพิ่มความหลากหลายและความสามารถในการปรับตัวของสิ่งมีชีวิตโดยทำให้เซลล์สามารถสร้างโปรตีนได้เมื่อจำเป็น แม้ว่าในปี 1951 บาร์บารา แมคคลินท็อกจะแสดงให้เห็นถึงปฏิสัมพันธ์ระหว่างยีนสองตำแหน่ง คือ Activator ( Ac ) และ Dissociator ( Ds ) ในการสร้างสีของเมล็ดข้าวโพด แต่การค้นพบระบบควบคุมยีนครั้งแรกนั้นถือกันอย่างกว้างขวางว่าเป็นการระบุโอเปรอนlac ในปี 1961 ซึ่งค้นพบโดยฟรองซัวส์ จาคอบและฌาคส์ โมโนด์ โดยโอเปรอน lac นี้จะสร้างเอนไซม์บางชนิดที่เกี่ยวข้องกับ การเผาผลาญแล คโตสในE. coliเฉพาะเมื่อมีแลคโตสและไม่มีกลูโคสเท่านั้น
ในสิ่งมีชีวิตหลายเซลล์ การควบคุมยีนเป็นตัวขับเคลื่อนการแบ่งแยกเซลล์และการสร้างรูปร่างในตัวอ่อน นำไปสู่การสร้างเซลล์ประเภทต่างๆ ที่มีรูปแบบการแสดงออกของยีนที่แตกต่างกันจาก ลำดับ จีโนม เดียวกัน แม้ว่าสิ่งนี้จะไม่สามารถอธิบายถึงที่มาของการควบคุมยีนได้ แต่นักชีววิทยาวิวัฒนาการก็รวมสิ่งนี้ไว้เป็นคำอธิบายบางส่วนของวิธี การทำงาน ของวิวัฒนาการในระดับโมเลกุลและเป็นหัวใจสำคัญของวิทยาศาสตร์ชีววิทยาการพัฒนาเชิงวิวัฒนาการ ("evo-devo")
ขั้นตอนที่ถูกควบคุมของการแสดงออกของยีน
ทุกขั้นตอนของการแสดงออกของยีนสามารถถูกควบคุมได้ ตั้งแต่การส่งสัญญาณ การ ถอดรหัส ไปจนถึง การ ดัดแปลง โปรตีน หลังการสังเคราะห์ต่อไปนี้คือรายการขั้นตอนที่การแสดงออกของยีนถูกควบคุม โดยจุดที่ถูกนำมาใช้มากที่สุดคือการเริ่มต้นการถอดรหัส ซึ่งเป็นขั้นตอนแรกของการถอดรหัส:
- การส่งสัญญาณ
- โครมาติน , การปรับโครงสร้างโครมาติน , โดเมนโครมาติน
- การถอดเสียง
- การดัดแปลงหลังการถอดรหัส
- การขนส่ง RNA
- การแปล
- การย่อยสลาย mRNA
การดัดแปลงดีเอ็นเอ

ในยูคาริโอต การเข้าถึงบริเวณขนาดใหญ่ของดีเอ็นเออาจขึ้นอยู่กับ โครงสร้าง โครมาตินซึ่งสามารถเปลี่ยนแปลงได้อันเป็นผลมาจาก การดัดแปลง ฮิสโตนที่ควบคุมโดยการเมทิลเลชันของดีเอ็นเอ อา ร์เอ็นเอที่ไม่เข้ารหัสหรือโปรตีนที่จับกับดีเอ็นเอดังนั้นการดัดแปลงเหล่านี้อาจควบคุมการแสดงออกของยีนเพิ่มขึ้นหรือลดลง การดัดแปลงบางอย่างที่ควบคุมการแสดงออกของยีนนั้นสามารถถ่ายทอดทางพันธุกรรมได้ และเรียกว่าการควบคุมทางเอพิเจเนติกส์
โครงสร้าง
การถอดรหัส DNA ถูกกำหนดโดยโครงสร้างของมัน โดยทั่วไป ความหนาแน่นของการบรรจุจะบ่งบอกถึงความถี่ของการถอดรหัส โปรตีนเชิงซ้อนแปดหน่วยที่เรียกว่าฮิสโตนร่วมกับส่วนของ DNA ที่พันรอบโปรตีนฮิสโตนทั้งแปด (เรียกรวมกันว่านิวคลีโอโซม) มีหน้าที่รับผิดชอบต่อปริมาณการขดตัวของ DNA และเชิงซ้อนเหล่านี้สามารถถูกดัดแปลงชั่วคราวโดยกระบวนการต่างๆ เช่นการฟอสโฟรีเลชันหรือถูกดัดแปลงอย่างถาวรมากขึ้นโดยกระบวนการต่างๆ เช่นการเมทิลเลชันการดัดแปลงดังกล่าวถือเป็นสาเหตุของการเปลี่ยนแปลงระดับการแสดงออกของยีนอย่างถาวรมากหรือน้อย[ 2 ]
เคมี
การเติมหมู่เมทิลลงบน DNAเป็นวิธีการปิดกั้นยีนที่พบได้ทั่วไป โดยทั่วไป DNA จะถูกเติมหมู่เมทิลโดยเอนไซม์เมทิลทรานสเฟอเรสบนนิวคลีโอไทด์ไซโตซีนในลำดับไดนิวคลีโอไทด์ CpG (เรียกอีกอย่างว่า " เกาะ CpG " เมื่อมีการรวมกลุ่มกันอย่างหนาแน่น) การวิเคราะห์รูปแบบการเติมหมู่เมทิลในบริเวณที่กำหนดของ DNA (ซึ่งอาจเป็นโปรโมเตอร์) สามารถทำได้โดยวิธีการที่เรียกว่าการทำแผนที่ด้วยไบซัลไฟต์ หมู่ไซโตซีนที่ถูกเติมหมู่เมทิลจะไม่เปลี่ยนแปลงจากการบำบัด ในขณะที่หมู่ที่ไม่ถูกเติมหมู่เมทิลจะเปลี่ยนเป็นยูราซิล ความแตกต่างจะถูกวิเคราะห์โดยการจัดลำดับ DNA หรือโดยวิธีการที่พัฒนาขึ้นเพื่อหาปริมาณ SNP เช่นPyrosequencing ( Biotage ) หรือ MassArray ( Sequenom ) โดยการวัดปริมาณสัมพัทธ์ของ C/T ที่ไดนิวคลีโอไทด์ CG รูปแบบการเติมหมู่เมทิลที่ผิดปกติเชื่อว่าเกี่ยวข้องกับการเกิดมะเร็ง[ 3 ]
การอะเซทิเลชันของฮิสโตนก็เป็นกระบวนการสำคัญในการถอดรหัสพันธุกรรมเช่นกัน เอนไซม์ ฮิสโตนอะเซทิลทรานสเฟอ เรส (HATs) เช่นโปรตีนที่จับกับ CREBยังทำหน้าที่แยก DNA ออกจากคอมเพล็กซ์ฮิสโตน ทำให้การถอดรหัสพันธุกรรมดำเนินต่อไปได้ บ่อยครั้งที่การเมทิลเลชันของ DNA และการดีอะเซทิเลชันของฮิสโตนทำงานร่วมกันในการปิดการทำงานของยีนการรวมกันของทั้งสองดูเหมือนจะเป็นสัญญาณให้ DNA ถูกจัดเรียงอย่างหนาแน่นมากขึ้น ส่งผลให้การแสดงออกของยีนลดลง
การควบคุมการถอดรหัส

การควบคุมการถอดรหัสจึงควบคุมว่าการถอดรหัสจะเกิดขึ้นเมื่อใดและสร้าง RNA มากน้อยเพียงใด การถอดรหัสยีนโดยRNA polymeraseสามารถถูกควบคุมได้ด้วยกลไกหลายอย่าง ปัจจัยจำเพาะจะเปลี่ยนแปลงความจำเพาะของ RNA polymerase ต่อโปรโมเตอร์หรือชุดของโปรโมเตอร์ที่กำหนด ทำให้มีโอกาสจับกับโปรโมเตอร์มากขึ้นหรือน้อยลง (เช่นปัจจัยซิกมาที่ใช้ในการถอดรหัสในโปรคาริโอต ) ตัวยับยั้งจะจับกับตัวดำเนินการซึ่งเป็นลำดับการเข้ารหัสบนสาย DNA ที่อยู่ใกล้หรือทับซ้อนกับบริเวณโปรโมเตอร์ ขัดขวางการเคลื่อนที่ของ RNA polymerase ไปตามสาย DNA จึงขัดขวางการแสดงออกของยีน ภาพด้านขวาแสดงให้เห็นถึงการควบคุมโดยตัวยับยั้งใน lac operon ปัจจัยการถอดรหัสทั่วไปจะวางตำแหน่ง RNA polymerase ที่จุดเริ่มต้นของลำดับการเข้ารหัสโปรตีน จากนั้นจึงปล่อย polymerase เพื่อถอดรหัส mRNA ตัว กระตุ้นจะเพิ่มปฏิสัมพันธ์ระหว่าง RNA polymerase และโปรโมเตอร์ เฉพาะ ส่งเสริมการแสดงออกของยีน ตัวกระตุ้นจะทำเช่นนี้โดยการเพิ่มแรงดึงดูดของ RNA polymerase ต่อโปรโมเตอร์ ผ่านการโต้ตอบกับหน่วยย่อยของ RNA polymerase หรือโดยอ้อมโดยการเปลี่ยนโครงสร้างของ DNA ตัวเร่งปฏิกิริยาคือไซต์บนเกลียว DNA ที่ถูกจับโดยตัวกระตุ้นเพื่อสร้างวง DNA นำโปรโมเตอร์เฉพาะไปยังคอมเพล็กซ์เริ่มต้น ตัวเร่งปฏิกิริยาพบได้บ่อยในยูคาริโอตมากกว่าโปรคาริโอต ซึ่งมีตัวอย่างเพียงไม่กี่ตัวอย่าง (จนถึงปัจจุบัน) [ 4 ]ตัวยับยั้งคือบริเวณของลำดับ DNA ที่เมื่อถูกจับโดยปัจจัยการถอดรหัสเฉพาะ จะสามารถยับยั้งการแสดงออกของยีนได้
การควบคุมโดย RNA
RNA เป็นตัวควบคุมกิจกรรมของยีนที่สำคัญ เช่นไมโคร RNA (miRNA), แอนติเซนส์ RNAหรือลอง นอนโค้ดดิ้ง RNA (lncRNA) lncRNA แตกต่างจากmRNAตรงที่พวกมันมีตำแหน่งและหน้าที่เฉพาะในเซลล์ เดิมทีพบว่า lncRNA อยู่ในนิวเคลียสและโครมาตินและปัจจุบันตำแหน่งและหน้าที่ของพวกมันมีความหลากหลายมาก บางชนิดยังคงอยู่ในโครมาตินและมีปฏิสัมพันธ์กับโปรตีน ในขณะที่ lncRNA บางชนิดส่งผลต่อการแสดงออกของยีนในโรคทางระบบประสาท เช่นโรคพาร์กินสันโรคฮันติงตันและโรคอัลไซเมอร์แต่บางชนิด เช่น PNCTR (pyrimidine-rich non-coding transcriptors) ก็มีบทบาทในมะเร็งปอดเนื่องจากบทบาทของ lncRNA ในโรคต่างๆ พวกมันจึงเป็นตัวบ่งชี้ทางชีวภาพ ที่มีศักยภาพ และอาจเป็นเป้าหมายที่มีประโยชน์สำหรับยาหรือการบำบัดด้วยยีนถึงแม้ว่าจะยังไม่มีการอนุมัติยาใดๆ ที่มุ่งเป้าไปที่ lncRNA ก็ตาม จำนวน lncRNA ในจีโนมของมนุษย์ยังไม่ได้รับการกำหนดอย่างชัดเจน แต่มีการประมาณการไว้ว่ามีจำนวนตั้งแต่ 16,000 ถึง 100,000 ยีน lnc [ 5 ]
การควบคุมยีนแบบเอพิเจเนติกส์

เอพิเจเนติกส์หมายถึงการดัดแปลงยีนที่ไม่เปลี่ยนแปลงลำดับ DNA หรือ RNA การดัดแปลงเอพิเจเนติกส์ยังเป็นปัจจัยสำคัญในการมีอิทธิพลต่อการแสดงออกของยีน การดัดแปลง เหล่านี้เกิดขึ้นบน DNA จีโนมและฮิสโตนและการดัดแปลงทางเคมีของพวกมันจะควบคุมการแสดงออกของยีนได้อย่างมีประสิทธิภาพมากขึ้น มีการดัดแปลง DNA หลายอย่าง (โดยปกติคือการเติมหมู่เมทิล ) และการดัดแปลง RNA มากกว่า 100 แบบในเซลล์ของสัตว์เลี้ยงลูกด้วยนม การดัดแปลงเหล่านี้ส่งผลให้การจับกันของโปรตีนกับ DNA เปลี่ยนแปลงไป และความเสถียรของ RNAและประสิทธิภาพการแปล เปลี่ยนแปลง ไป[ 6 ]
กรณีพิเศษในชีววิทยาของมนุษย์และโรคต่างๆ
การควบคุมการถอดรหัสในมะเร็ง
ในสัตว์มีกระดูกสันหลัง โปรโมเตอร์ของยีนส่วนใหญ่มีเกาะ CpG ที่มี ไซต์ CpGจำนวนมาก[ 7 ] เมื่อไซต์ CpG ของโปรโมเตอร์ของยีนจำนวนมากถูกเมทิลเลชันยีนนั้นก็จะถูกปิดการทำงาน[ 8 ] มะเร็งลำไส้ใหญ่และทวารหนักมักมีการกลายพันธุ์ ที่เป็นตัวขับเคลื่อน 3 ถึง 6 ตัว และการกลายพันธุ์ แบบร่วมเดินทางหรือแบบผู้โดยสาร33 ถึง 66 ตัว [ 9 ] อย่างไรก็ตาม การปิดการทำงานของการถอดรหัสอาจมีความสำคัญมากกว่าการกลายพันธุ์ในการทำให้เกิดการลุกลามไปสู่มะเร็ง ตัวอย่างเช่น ในมะเร็งลำไส้ใหญ่และทวารหนัก ยีนประมาณ 600 ถึง 800 ยีนถูกปิดการทำงานของการถอดรหัสโดยการเมทิลเลชันของเกาะ CpG (ดูการควบคุมการถอดรหัสในมะเร็ง ) การยับยั้งการถอดรหัสในมะเร็งยังสามารถเกิดขึ้นได้จาก กลไก ทางพันธุกรรม อื่นๆ เช่น การเปลี่ยนแปลงการแสดงออกของไมโครอาร์เอ็นเอ[ 10 ] ในมะเร็งเต้านม การยับยั้งการถอดรหัสของBRCA1อาจเกิดขึ้นบ่อยกว่าโดยไมโครอาร์เอ็นเอ-182 ที่แสดงออกมากเกินไป มากกว่าโดยไฮเปอร์เมทิลเลชันของโปรโมเตอร์ BRCA1 (ดูการแสดงออกของ BRCA1 ต่ำในมะเร็งเต้านมและมะเร็งรังไข่ )
การควบคุมการถอดรหัสในภาวะเสพติด
หนึ่งในลักษณะสำคัญของการเสพติดคือความคงอยู่ การเปลี่ยนแปลงพฤติกรรมที่คงอยู่ดูเหมือนจะเกิดจากการเปลี่ยนแปลงที่ยั่งยืน ซึ่งเป็นผลมาจาก การเปลี่ยนแปลง ทางพันธุกรรมที่มีผลต่อการแสดงออกของยีนในบริเวณเฉพาะของสมอง[ 11 ] ยาเสพติดก่อให้เกิดการเปลี่ยนแปลงทางพันธุกรรม 3 ประเภทในสมอง ได้แก่ (1) การอะเซทิเลชัน ของฮิสโตนและการเมทิลเลชันของฮิสโตน (2) การเมทิลเลชันของ DNA ที่ไซต์ CpGและ (3) การควบคุมทางพันธุกรรมลดลงหรือเพิ่มขึ้นของไมโครอาร์เอ็นเอ[ 11 ] [ 12 ] (ดูรายละเอียดเพิ่มเติมได้ใน Epigenetics of cocaine addiction )
Chronic nicotine intake in mice alters brain cell epigenetic control of gene expression through acetylation of histones. This increases expression in the brain of the protein FosB, important in addiction.[13] Cigarette addiction was also studied in about 16,000 humans, including never smokers, current smokers, and those who had quit smoking for up to 30 years.[14] In blood cells, more than 18,000 CpG sites (of the roughly 450,000 analyzed CpG sites in the genome) had frequently altered methylation among current smokers. These CpG sites occurred in over 7,000 genes, or roughly a third of known human genes. The majority of the differentially methylated CpG sites returned to the level of never-smokers within five years of smoking cessation. However, 2,568 CpGs among 942 genes remained differentially methylated in former versus never smokers. Such remaining epigenetic changes can be viewed as "molecular scars"[12] that may affect gene expression.
In rodent models, drugs of abuse, including cocaine,[15] methamphetamine,[16][17] alcohol[18] and tobacco smoke products,[19] all cause DNA damage in the brain. During repair of DNA damages some individual repair events can alter the methylation of DNA and/or the acetylations or methylations of histones at the sites of damage, and thus can contribute to leaving an epigenetic scar on chromatin.[20]
Such epigenetic scars likely contribute to the persistent epigenetic changes found in addiction.
Regulation of transcription in learning and memory

In mammals, methylation of cytosine (see Figure) in DNA is a major regulatory mediator. Methylated cytosines primarily occur in dinucleotide sequences where cytosine is followed by a guanine, a CpG site. The total number of CpG sites in the human genome is approximately 28 million.[21] and generally about 70% of all CpG sites have a methylated cytosine.[22]

ในหนู การเรียนรู้ที่เจ็บปวด เช่นการปรับสภาพความกลัว ตามบริบท สามารถส่งผลให้เกิดความทรงจำที่น่ากลัวไปตลอดชีวิตหลังจากการฝึกเพียงครั้งเดียว[ 23 ] การเมทิลเลชั่นของไซโตซีนจะเปลี่ยนแปลงในบริเวณโปรโมเตอร์ของยีนประมาณ 9.17% ของยีนทั้งหมดในดีเอ็นเอของเซลล์ประสาทฮิปโปแคมปัสของหนูที่ได้รับประสบการณ์การปรับสภาพความกลัว ในช่วงสั้นๆ [ 24 ] ฮิปโปแคมปัสเป็นที่ที่ความทรงจำใหม่ๆ ถูกจัดเก็บไว้ในตอนแรก
การเติมหมู่เมทิลให้กับ CpG ในบริเวณโปรโมเตอร์ของยีนจะยับยั้งการถอดรหัส[ 25 ]ในขณะที่การเติมหมู่เมทิลให้กับ CpG ในส่วนของยีนจะเพิ่มการแสดงออก[ 26 ]เอนไซม์ TETมีบทบาทสำคัญในการกำจัดหมู่เมทิลออกจากไซโตซีนที่ถูกเติมหมู่เมทิล การกำจัดหมู่เมทิลออกจาก CpG ในโปรโมเตอร์ของยีนโดย กิจกรรม ของเอนไซม์ TETจะเพิ่มการถอดรหัสของยีน[ 27 ]
เมื่อการปรับสภาพความกลัว ตามบริบท ถูกนำมาใช้กับหนู จะพบว่ามีบริเวณที่มีการเติมหมู่เมทิลแตกต่างกัน มากกว่า 5,000 บริเวณ (DMRs) (แต่ละบริเวณมีนิวคลีโอไทด์ 500 ตัว) เกิดขึ้นใน จีโนมประสาท ของฮิปโปแคมปัส ของหนู ทั้งในเวลาหนึ่งชั่วโมงและ 24 ชั่วโมงหลังจากการปรับสภาพในฮิปโปแคมปัส[ 24 ] ซึ่งทำให้ยีนประมาณ 500 ยีนถูกควบคุมให้ทำงานมากขึ้น (มักเกิดจากการลดหมู่เมทิลของไซต์ CpG ในบริเวณโปรโมเตอร์) และยีนประมาณ 1,000 ยีนถูกควบคุมให้ทำงานน้อยลง (มักเกิดจากการสร้าง 5-เมทิลไซโตซีนขึ้นใหม่ที่ไซต์ CpG ในบริเวณโปรโมเตอร์) รูปแบบของยีนที่ถูกกระตุ้นและยับยั้งภายในเซลล์ประสาทดูเหมือนจะเป็นพื้นฐานทางโมเลกุลสำหรับการสร้างความทรงจำชั่วคราวแรกของเหตุการณ์การฝึกนี้ในฮิปโปแคมปัสของสมองหนู[ 24 ]
การควบคุมหลังการถอดรหัส
หลังจากที่ DNA ถูกถอดรหัสและเกิดเป็น mRNA แล้ว จะต้องมีการควบคุมบางอย่างเกี่ยวกับปริมาณการแปล mRNA เป็นโปรตีน เซลล์จะทำเช่นนี้โดยการปรับเปลี่ยนการเติมหมวก (capping), การตัดต่อ (splicing), การเพิ่มหางโพลีเอ (Poly(A) Tail), อัตราการส่งออกไปยังนิวเคลียสที่จำเพาะต่อลำดับ และในบางบริบท การกักเก็บ RNA ที่ถอดรหัสแล้ว กระบวนการเหล่านี้เกิดขึ้นในยูคาริโอต แต่ไม่เกิดขึ้นในโปรคาริโอต การปรับเปลี่ยนนี้เป็นผลมาจากโปรตีนหรือ RNA ที่ถอดรหัสแล้ว ซึ่งในทางกลับกันก็ได้รับการควบคุมและอาจมีความสัมพันธ์กับลำดับบางอย่าง
สามบริเวณหลักที่ไม่ได้รับการแปลและไมโครอาร์เอ็นเอ
บริเวณ 3'-UTR ของเมสเซนเจอร์อาร์เอ็นเอ (mRNA) มักมีลำดับควบคุมที่ส่งผลต่อการแสดงออกของยีนหลังการถอดรหัส[ 28 ]บริเวณ 3'-UTR ดังกล่าว มักมีทั้งตำแหน่งการจับของไมโครอาร์เอ็นเอ (miRNA) และโปรตีนควบคุม โดยการจับกับตำแหน่งเฉพาะภายใน 3'-UTR miRNA สามารถลดการแสดงออกของยีนของ mRNA ต่างๆ ได้ โดยการยับยั้งการแปลหรือทำให้เกิดการย่อยสลายของทรานสคริปต์โดยตรง นอกจากนี้ 3'-UTR อาจมีบริเวณตัวยับยั้งที่จับกับโปรตีนยับยั้งที่ยับยั้งการแสดงออกของ mRNA
บริเวณ 3'-UTR มักมีองค์ประกอบตอบสนองต่อ miRNA (MREs) MREs คือลำดับเบสที่ miRNA จะจับด้วย ซึ่งเป็นลวดลายที่พบได้ทั่วไปในบริเวณ 3'-UTR ในบรรดาลวดลายควบคุมทั้งหมดภายในบริเวณ 3'-UTR (เช่น รวมถึงบริเวณยับยั้งการแสดงออกของยีน) MREs คิดเป็นประมาณครึ่งหนึ่งของลวดลายทั้งหมด
ณ ปี 2014 เว็บไซต์miRBase [ 29 ]ซึ่งเป็นคลังข้อมูลลำดับและคำอธิบายประกอบของ miRNA ได้แสดงรายการ 28,645 รายการใน 233 สปีชีส์ทางชีวภาพ ในจำนวนนี้ miRNA จำนวน 1,881 รายการอยู่ในตำแหน่ง miRNA ของมนุษย์ที่ได้รับการอธิบายไว้ มีการคาดการณ์ว่า miRNA จะมี mRNA เป้าหมายโดยเฉลี่ยประมาณสี่ร้อยรายการ (ส่งผลต่อการแสดงออกของยีนหลายร้อยยีน) [ 30 ] Freidman et al. [ 30 ]ประมาณการว่าตำแหน่งเป้าหมายของ miRNA มากกว่า 45,000 ตำแหน่งภายใน 3'-UTR ของ mRNA ของมนุษย์ได้รับการอนุรักษ์ไว้เหนือระดับพื้นหลัง และยีนที่เข้ารหัสโปรตีนของมนุษย์มากกว่า 60% อยู่ภายใต้แรงกดดันในการคัดเลือกเพื่อรักษาการจับคู่กับ miRNA
การทดลองโดยตรงแสดงให้เห็นว่า miRNA เพียงตัวเดียวสามารถลดความเสถียรของ mRNA ที่ไม่ซ้ำกันหลายร้อยตัวได้[ 31 ]การทดลองอื่นๆ แสดงให้เห็นว่า miRNA เพียงตัวเดียวอาจยับยั้งการผลิตโปรตีนได้หลายร้อยตัว แต่การยับยั้งนี้มักจะค่อนข้างอ่อน (น้อยกว่า 2 เท่า) [ 32 ] [ 33 ]
ผลกระทบของการควบคุมการแสดงออกของยีนโดย miRNA ที่ผิดปกติดูเหมือนจะมีความสำคัญในมะเร็ง[ 34 ]ตัวอย่างเช่น ในมะเร็งทางเดินอาหาร บทความในปี 2015 ระบุว่า miRNA จำนวน 9 ตัว มีการเปลี่ยนแปลง ทางเอพิเจเนติกส์และมีประสิทธิภาพในการลดการทำงานของเอนไซม์ซ่อมแซม DNA [ 35 ]
ผลกระทบของการควบคุมการแสดงออกของยีนโดย miRNA ที่ผิดปกติดูเหมือนจะมีความสำคัญในความผิดปกติทางจิตเวช เช่นโรคจิตเภทโรคอารมณ์สองขั้ว โรคซึมเศร้า โรค พาร์กินสันโรคอัลไซเมอร์และความผิดปกติ ในกลุ่มอาการ ออทิสติก[ 36 ] [ 37 ] [ 38 ]
ระเบียบการแปล
การแปลรหัส mRNA สามารถควบคุมได้ด้วยกลไกหลายอย่าง โดยส่วนใหญ่จะเกิดขึ้นในระดับการเริ่มต้น การดึงดูดหน่วยย่อยของไรโบโซมขนาดเล็กสามารถถูกปรับเปลี่ยนได้ด้วยโครงสร้างทุติยภูมิของ mRNA การจับกับ RNA แอนติเซนส์ หรือการจับกับโปรตีน ในทั้งโปรคาริโอตและยูคาริโอต มีโปรตีนที่จับกับ RNA อยู่เป็นจำนวนมาก ซึ่งมักจะถูกนำไปยังลำดับเป้าหมายโดยโครงสร้างทุติยภูมิของทรานสคริปต์ ซึ่งอาจเปลี่ยนแปลงได้ขึ้นอยู่กับสภาวะบางอย่าง เช่น อุณหภูมิหรือการมีอยู่ของลิแกนด์ (แอพทาเมอร์) ทรานสคริปต์บางชนิดทำหน้าที่เป็นไรโบไซม์และควบคุมการแสดงออกของตัวเองได้
ตัวอย่างของการควบคุมยีน
- การเหนี่ยวนำเอนไซม์เป็นกระบวนการที่โมเลกุล (เช่น ยา) เหนี่ยวนำ (กล่าวคือ เริ่มต้นหรือเพิ่ม) การแสดงออกของเอนไซม์
- การเหนี่ยวนำโปรตีนช็อกความร้อนในแมลงวันผลไม้Drosophila melanogaster
- โอเปรอน Lacเป็นตัวอย่างที่น่าสนใจของการควบคุมการแสดงออกของยีน
- แม้ว่าไวรัสจะมีเพียงไม่กี่จีน แต่ก็มีกลไกในการควบคุมการแสดงออกของจีน โดยทั่วไปจะแบ่งออกเป็นระยะเริ่มต้นและระยะท้าย โดยใช้ระบบแบบเส้นตรงที่ควบคุมโดยแอนติเทอร์มิเนเตอร์ ( แลมบ์ดาฟาจ ) หรือตัวปรับการต่อเชื่อม ( เอชไอวี )
- Gal4เป็นตัวกระตุ้นการถอดรหัสที่ควบคุมการแสดงออกของ GAL1, GAL7 และ GAL10 (ซึ่งทั้งหมดเข้ารหัสสำหรับการเผาผลาญกาแลคโตสในยีสต์)ระบบ GAL4/UASถูกนำมาใช้ในสิ่งมีชีวิตหลากหลายชนิดในไฟลัมต่างๆ เพื่อศึกษาการแสดงออกของยีน [ 39 ]
ชีววิทยาการพัฒนา
ระบบควบคุมจำนวนมากที่ได้รับการศึกษามาจากชีววิทยาการพัฒนาตัวอย่างเช่น:
- ความสอดคล้องกันของ กลุ่ม ยีน Hoxกับรูปแบบการเรียงตัวแบบซ้อนกันจากด้านหน้าไปด้านหลัง
- การสร้างรูปแบบของมือ (นิ้วมือ - ข้อต่อระหว่างนิ้ว): การไล่ระดับของโซนิคเฮดจ์ฮ็อก (ปัจจัยกระตุ้นที่หลั่งออกมา) จากโซนกิจกรรมโพลาไรซ์ในแขนขา ซึ่งสร้างการไล่ระดับของ Gli3 ที่ทำงานอยู่ ซึ่งจะกระตุ้น Gremlin ซึ่งจะยับยั้ง BMPs ที่หลั่งออกมาในแขนขาเช่นกัน ส่งผลให้เกิดรูปแบบการทำงานสลับกันอันเป็นผลมาจากระบบปฏิกิริยา-การแพร่กระจายนี้
- การสร้างโซไมต์คือการสร้างส่วนต่างๆ (โซไมต์) จากเนื้อเยื่อที่เป็นเนื้อเดียวกัน ( เมโซเดิร์ม ก่อนโซไมต์ ) โดยจะสร้างขึ้นตามลำดับจากด้านหน้าไปด้านหลัง ในสัตว์มีถุงน้ำคร่ำ กระบวนการนี้อาจเกิดขึ้นได้โดยใช้การไล่ระดับสองแบบที่ตรงข้ามกัน คือ กรดเรติโนอิกที่ด้านหน้า (เวฟฟรอนท์) และ Wnt กับ Fgf ที่ด้านหลัง ซึ่งเชื่อมโยงกับรูปแบบการแกว่ง (นาฬิกาการแบ่งส่วน) ที่ประกอบด้วย FGF + Notch และ Wnt ในเฟสตรงข้าม[ 40 ]
- การกำหนดเพศในโซมาของ Drosophila ต้องอาศัยการรับรู้อัตราส่วนของยีนออโตโซมต่อ ยีนที่เข้ารหัสโดย โครโมโซมเพศซึ่งส่งผลให้เกิดปัจจัยการตัดต่อแบบไร้เพศในเพศเมีย ส่งผลให้เกิดไอโซฟอร์มเพศเมียของ doublesex [ 41 ]
วงจร
การเพิ่มขึ้นและการลดลงของการทำงาน
การควบคุมการแสดงออกของยีน (Up-regulation) เป็นกระบวนการที่เกิดขึ้นภายในเซลล์โดยถูกกระตุ้นด้วยสัญญาณ (ทั้งจากภายในหรือภายนอกเซลล์) ซึ่งส่งผลให้มีการแสดงออกของยีนหนึ่งหรือหลายยีนเพิ่มขึ้น และส่งผลให้มีการสร้างโปรตีนจากยีนเหล่านั้นเพิ่มขึ้นด้วย ในทางกลับกัน การควบคุมการแสดงออกของยีน (Down-regulation) เป็นกระบวนการที่ส่งผลให้การแสดงออกของยีนและโปรตีนที่เกี่ยวข้องลดลง
- การควบคุมการแสดงออกของยีนเพิ่มขึ้นเกิดขึ้นเมื่อเซลล์ขาดตัวรับบางชนิด ในกรณีนี้ โปรตีนตัวรับจะถูกสังเคราะห์และลำเลียงไปยังเยื่อหุ้มเซลล์มากขึ้น ส่งผลให้ความไวของเซลล์กลับคืนสู่ระดับปกติและเกิดภาวะสมดุล ขึ้น ใหม่
- การลดระดับการแสดงออกเกิดขึ้นเมื่อเซลล์ถูกกระตุ้นมากเกินไปโดยสารสื่อประสาท ฮอร์โมนหรือยาเป็นเวลานาน และการแสดงออกของโปรตีนตัวรับจะลดลงเพื่อปกป้องเซลล์ (ดูเพิ่มเติมที่ภาวะดื้อยา )
ระบบที่เหนี่ยวนำได้เทียบกับระบบที่ยับยั้งได้

การควบคุมยีนสามารถสรุปได้ด้วยการตอบสนองของระบบนั้นๆ ดังนี้:
- ระบบที่ถูกกระตุ้นได้ - ระบบที่ถูกกระตุ้นได้จะปิดอยู่หากไม่มีโมเลกุลบางชนิด (เรียกว่าตัวกระตุ้น) ที่ทำให้เกิดการแสดงออกของยีน โมเลกุลนั้นเรียกว่า "ตัวกระตุ้นการแสดงออก" วิธีการที่เกิดขึ้นนั้นขึ้นอยู่กับกลไกการควบคุม รวมถึงความแตกต่างระหว่างเซลล์โปรคาริโอติกและเซลล์ยูคาริโอติก
- ระบบที่ถูกยับยั้งได้ - ระบบที่ถูกยับยั้งได้จะทำงานอยู่ ยกเว้นในกรณีที่มีโมเลกุลบางชนิด (เรียกว่า โคเรเพรสเซอร์) ที่ทำหน้าที่ยับยั้งการแสดงออกของยีน โมเลกุลนี้เรียกว่า "ตัวยับยั้งการแสดงออก" วิธีการที่เกิดขึ้นนั้นขึ้นอยู่กับกลไกการควบคุม รวมถึงความแตกต่างระหว่างเซลล์โปรคาริโอติกและยูคาริโอติก
ระบบGAL4/UASเป็นตัวอย่างของทั้งระบบที่เหนี่ยวนำได้และยับยั้งได้Gal4จับกับลำดับการกระตุ้นต้นน้ำ (UAS) เพื่อกระตุ้นการถอดรหัสของคาสเซ็ต GAL1/GAL7/GAL10 ในทางกลับกัน การตอบสนองของ MIG1ต่อการมีอยู่ของกลูโคสสามารถยับยั้ง GAL4 และหยุดการแสดงออกของคาสเซ็ต GAL1/GAL7/GAL10 ได้[ 42 ]
วงจรเชิงทฤษฎี
- ตัวยับยั้ง/ตัวกระตุ้น: การกระตุ้นเซนเซอร์ส่งผลให้เกิดการเปลี่ยนแปลงการแสดงออกของยีน
- กลไกป้อนกลับเชิงลบ: ผลิตภัณฑ์ของยีนจะลดการผลิตของตัวเองลงโดยตรงหรือโดยอ้อม ซึ่งอาจส่งผลให้เกิด...
- รักษาระดับการถอดรหัสให้คงที่/เป็นสัดส่วนกับปัจจัยหนึ่ง
- การยับยั้งปฏิกิริยาที่ควบคุมไม่ได้เมื่อทำงานร่วมกับวงจรป้อนกลับเชิงบวก
- สร้างออสซิลเลเตอร์โดยใช้ประโยชน์จากความล่าช้าของเวลาในการถอดรหัสและการแปล เนื่องจากครึ่งชีวิตของ mRNA และโปรตีนนั้นสั้นกว่า
- ปฏิกิริยาตอบสนองเชิงบวก: ผลิตภัณฑ์ของยีนกระตุ้นการผลิตของตัวเองโดยตรงหรือโดยอ้อม ซึ่งอาจส่งผลให้เกิด...
- การขยายสัญญาณ
- สวิตช์แบบสองสถานะเกิดขึ้นเมื่อยีนสองตัวยับยั้งซึ่งกันและกัน และทั้งสองตัวมีปฏิกิริยาป้อนกลับเชิงบวก
- การสร้างรูปแบบ
วิธีการศึกษา

โดยทั่วไป การทดลองส่วนใหญ่ที่ตรวจสอบการแสดงออกของยีนที่แตกต่างกัน มักใช้สารสกัด RNA จากเซลล์ทั้งหมด ซึ่งเรียกว่าระดับสภาวะคงที่ เพื่อตรวจสอบว่ายีนใดเปลี่ยนแปลงไปและเปลี่ยนแปลงไปมากน้อยเพียงใด อย่างไรก็ตาม วิธีการเหล่านี้ไม่ได้ให้ข้อมูลเกี่ยวกับตำแหน่งที่การควบคุมเกิดขึ้น และอาจปกปิดกระบวนการควบคุมที่ขัดแย้งกัน ( ดูการควบคุมหลังการถอดรหัส ) แต่ก็ยังเป็นวิธีที่วิเคราะห์กันมากที่สุด ( PCR เชิงปริมาณและไมโครอาร์เรย์ดีเอ็นเอ )
ในการศึกษาการแสดงออกของยีน มีหลายวิธีในการตรวจสอบขั้นตอนต่างๆ ในสิ่งมีชีวิตยูคาริโอต วิธีเหล่านี้ได้แก่:
- สภาพแวดล้อมของโครมาตินในบริเวณนั้นสามารถกำหนดได้โดย การวิเคราะห์ ChIP-chipโดยการดึงRNA Polymerase II , การดัดแปลงฮิสโตน 3 , โปรตีนกลุ่ม Trithorax , โปรตีนกลุ่ม Polycombหรือองค์ประกอบจับกับ DNA อื่นๆ ที่มีแอนติบอดีที่ดีพร้อมใช้งาน
- ปฏิสัมพันธ์ แบบเอพิสแตติกสามารถตรวจสอบได้โดยการวิเคราะห์อาร์เรย์ทางพันธุกรรมสังเคราะห์
- เนื่องจากการควบคุมหลังการถอดรหัส อัตราการถอดรหัสและระดับ RNA ทั้งหมดจึงแตกต่างกันอย่างมีนัยสำคัญ การวัดอัตราการถอดรหัส สามารถทำได้โดยใช้การทดสอบ Nuclear Run-onและกำลังมีการพัฒนาวิธีการใหม่ๆ ที่มีประสิทธิภาพสูงขึ้น โดยใช้ การติดฉลาก ไทออลแทนการใช้กัมมันตภาพรังสี[ 43 ]
- มีเพียง 5% ของ RNA ที่ถูกสังเคราะห์ในนิวเคลียสเท่านั้นที่ยังคงอยู่[ 44 ]และไม่เพียงแต่อินทรอนผลิตภัณฑ์ที่ไม่สมบูรณ์และทรานสคริปต์ที่ไม่มีความหมายเท่านั้นที่จะถูกย่อยสลาย ดังนั้น ความแตกต่างในระดับนิวเคลียสและไซโตพลาสซึมสามารถมองเห็นได้โดยการแยกสองส่วนนี้ด้วยการสลายเซลล์อย่างอ่อนโยน[ 45 ]
- สามารถวิเคราะห์การตัดต่อทางเลือกได้โดยใช้ชุดตรวจสอบการตัดต่อหรือชุดตรวจสอบการเรียงตัวของยีน ( ดูที่ไมโครอาร์เรย์ดีเอ็นเอ )
- RNA ทั้งหมดในสิ่งมีชีวิต จะรวมตัวกันเป็น RNPปริมาณของทรานสคริปต์ที่จับกับโปรตีนจำเพาะสามารถวิเคราะห์ได้ด้วยRIP-Chipตัวอย่างเช่นDCP2จะบ่งชี้ถึงโปรตีนที่ถูกกักเก็บไว้ ในขณะที่ การจับกับ ไรโบโซมจะบ่งชี้ถึงทรานสคริปต์ที่ทำงานอยู่ในการถอดรหัส (แม้ว่าวิธีการที่เก่ากว่าที่เรียกว่า การแยกส่วน โพลีโซมยังคงเป็นที่นิยมในบางห้องปฏิบัติการ)
- สามารถวิเคราะห์ระดับโปรตีนได้โดยใช้แมสสเปกโทรเมตรีซึ่งสามารถนำมาเปรียบเทียบกับ ข้อมูล PCR เชิงปริมาณ เท่านั้น เนื่องจาก ข้อมูล ไมโครอาร์เรย์เป็นข้อมูลเชิงสัมพัทธ์ ไม่ใช่ข้อมูลเชิงสัมบูรณ์
- อัตราการสลายตัวของ RNA และโปรตีนจะวัดโดยใช้สารยับยั้งการถอดรหัส ( actinomycin Dหรือα-Amanitin ) หรือสารยับยั้งการแปลรหัส ( Cycloheximide ) ตามลำดับ
ดูเพิ่มเติม
- ปัจจัยถอดรหัสเทียม (โมเลกุลขนาดเล็กที่เลียนแบบโปรตีนปัจจัยถอดรหัส)
- แบบจำลองเซลล์
- ลำดับดีเอ็นเอที่ไม่เข้ารหัสที่ได้รับการอนุรักษ์
- ตัวเร่งปฏิกิริยา (พันธุศาสตร์)
- โครงสร้างยีน
- การแสดงออกของยีนในเชิงพื้นที่และเวลา
- ระบบกลูโคซิลทรานสเฟอเรสของยีนควบคุม (Rgg/SHP)
หมายเหตุและเอกสารอ้างอิง
- ^ "ยีนสามารถเปิดและปิดได้ในเซลล์หรือไม่?" . พันธุศาสตร์อ้างอิง
{{cite web}}: CS1 maint: บริการเก็บถาวรที่เลิกใช้แล้ว ( ลิงก์ ) - ^ Bell JT, Pai AA, Pickrell JK, Gaffney DJ, Pique-Regi R, Degner JF และคณะ (2011). "รูปแบบการเมทิลเลชั่นของ DNA สัมพันธ์กับความแปรผันทางพันธุกรรมและการแสดงออกของยีนในสายเซลล์ HapMap" . Genome Biology . 12 (1) R10. doi : 10.1186/gb-2011-12-1-r10 . PMC 3091299 . PMID 21251332 .
- ^ Vertino PM, Spillare EA, Harris CC, Baylin SB (เมษายน 1993). "รูปแบบการเมทิลเลชั่นของโครโมโซมที่เปลี่ยนแปลงไปเกิดขึ้นพร้อมกับการเปลี่ยนแปลงของเซลล์เยื่อบุผิวหลอดลมของมนุษย์ที่เกิดจากยีนก่อมะเร็ง" (PDF) . การวิจัยมะเร็ง . 53 (7): 1684– 9. PMID 8453642 .
- ^ Austin S, Dixon R (มิถุนายน 1992). "โปรตีน NTRC ที่จับกับตัวเร่งปฏิกิริยาในโปรคาริโอตมีกิจกรรม ATPase ซึ่งขึ้นอยู่กับการฟอสโฟรีเลชันและ DNA"วารสารEMBO 11 ( 6): 2219– 28. doi : 10.1002/j.1460-2075.1992.tb05281.x . PMC 556689 . PMID 1534752 .
- ^ Statello L, Guo CJ, Chen LL, Huarte M (กุมภาพันธ์ 2021). "การควบคุมยีนโดย RNA ที่ไม่เข้ารหัสแบบยาวและหน้าที่ทางชีวภาพ" Nature Reviews. Molecular Cell Biology . 22 (2): 96– 118. doi : 10.1038/ s41580-020-00315-9 . ISSN 1471-0072 . PMC 7754182. PMID 33353982 .
- ^ Kan RL, Chen J, Sallam T (กรกฎาคม 2021). "การสื่อสารระหว่างกลไกเอพิแทรนสคริปโตมิกและเอพิเจเนติกในการควบคุมยีน" . Trends in Genetics . 38 (2): 182– 193. doi : 10.1016/j.tig.2021.06.014 . PMC 9093201 . PMID 34294427 . S2CID 236200223 .
- ^ Saxonov S, Berg P, Brutlag DL (มกราคม 2549). "การวิเคราะห์ไดนิวคลีโอไทด์ CpG ทั่วทั้งจีโนมในจีโนมมนุษย์แยกแยะโปรโมเตอร์ออกเป็นสองกลุ่มที่แตกต่างกัน" Proceedings of the National Academy of Sciences of the United States of America . 103 (5): 1412– 7. Bibcode : 2006PNAS..103.1412S . doi : 10.1073 / pnas.0510310103 . PMC 1345710. PMID 16432200 .
- ^ Bird A (มกราคม 2545). "รูปแบบการเมทิลเลชั่นของ DNA และความทรงจำทางเอพิเจเนติก" Genes & Development . 16 (1): 6– 21. doi : 10.1101/gad.947102 . PMID 11782440 .
- ^ Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA, Kinzler KW (มีนาคม 2013). "ภูมิทัศน์จีโนมมะเร็ง" . Science . 339 (6127): 1546– 58. Bibcode : 2013Sci...339.1546V . doi : 10.1126/science.1235122 . PMC 3749880 . PMID 23539594 .
- ↑เทสซิโตเร เอ, ชิคชาเรลลี จี, เดล เวคคิโอ เอฟ, กัจเจียโน เอ, แวร์เซลลา ดี, ฟิสเคียตติ เอ็ม, และคณะ (2014) "MicroRNAs ในเครือข่ายความเสียหาย/ซ่อมแซม DNA และมะเร็ง " วารสารจีโนมิกส์นานาชาติ . 2557 820248. ดอย : 10.1155/2014/820248 . PMC 3926391 . PMID24616890 .
- ^ a b Nestler EJ (มกราคม 2014). "กลไกเอพิเจเนติกส์ของการติดยาเสพติด" . Neuropharmacology . 76 Pt B: 259– 68. doi : 10.1016/j.neuropharm.2013.04.004 . PMC 3766384 . PMID 23643695 .
- ^ a b Robison AJ, Nestler EJ (ตุลาคม 2011). "กลไกการถอดรหัสและเอพิเจเนติกส์ของการเสพติด" Nature Reviews. Neuroscience . 12 (11): 623– 37. doi : 10.1038/nrn3111 . PMC 3272277 . PMID 21989194 .
- ^ Levine A, Huang Y, Drisaldi B, Griffin EA, Pollak DD, Xu S และคณะ (พฤศจิกายน 2011). "กลไกโมเลกุลของยาเสพติดที่เป็นจุดเริ่มต้น: การเปลี่ยนแปลงทางเอพิเจเนติกส์ที่เริ่มต้นโดยนิโคตินกระตุ้นการแสดงออกของยีนโดยโคเคน" Science Translational Medicine . 3 (107): 107ra109. doi : 10.1126/scitranslmed.3003062 . PMC 4042673 . PMID 22049069 .
- ^ Joehanes R, Just AC, Marioni RE, Pilling LC, Reynolds LM, Mandaviya PR และคณะ (ตุลาคม 2016). "ลายเซ็นเอพิเจเนติกส์ของการสูบบุหรี่" . Circulation: Cardiovascular Genetics . 9 (5): 436– 447. doi : 10.1161/CIRCGENETICS.116.001506 . PMC 5267325 . PMID 27651444 .
- ^ de Souza MF, Gonçales TA, Steinmetz A, Moura DJ, Saffi J, Gomez R, Barros HM (เมษายน 2014). "โคเคนทำให้เกิดความเสียหายต่อ DNA ในบริเวณสมองที่แตกต่างกันของหนูเพศเมียภายใต้สภาวะฮอร์โมนที่แตกต่างกัน" เภสัชวิทยาและสรีรวิทยาทางคลินิกและการทดลอง 41 ( 4): 265– 9. doi : 10.1111/1440-1681.12218 . PMID 24552452 . S2CID 20849951 .
- ^ Johnson Z, Venters J, Guarraci FA, Zewail-Foote M (มิถุนายน 2015). "เมทแอมเฟตามีนทำให้เกิดความเสียหายต่อ DNA ในบริเวณเฉพาะของสมองหนูตัวเมีย" เภสัชวิทยาและสรีรวิทยาทางคลินิกและการทดลอง 42 ( 6): 570– 5. doi : 10.1111/1440-1681.12404 . PMID 25867833 . S2CID 24182756 .
- ^ Tokunaga I, Ishigami A, Kubo S, Gotohda T, Kitamura O (สิงหาคม 2551). "ความเสียหายของดีเอ็นเอจากปฏิกิริยาเปอร์ออกซิเดชันและการเกิดอะพอพโทซิสในสมองหนูที่ได้รับการรักษาด้วยเมทแอมเฟตามีน"วารสารการวิจัยทางการแพทย์ 55 ( 3– 4 ): 241– 5. doi : 10.2152/jmi.55.241 . PMID 18797138 .
- ^ Rulten SL, Hodder E, Ripley TL, Stephens DN, Mayne LV (กรกฎาคม 2551). "แอลกอฮอล์ก่อให้เกิดความเสียหายต่อ DNA และโปรตีน Fanconi anemia D2 ซึ่งบ่งชี้ว่า FANCD2 มีส่วนเกี่ยวข้องในเส้นทางการตอบสนองต่อความเสียหายของ DNA ในสมอง" Alcoholism : Clinical and Experimental Research . 32 (7): 1186– 96. doi : 10.1111/j.1530-0277.2008.00673.x . PMID 18482162 .
- ^ Adhami N, Chen Y, Martins-Green M (ตุลาคม 2017). "สามารถตรวจพบไบโอมาร์กเกอร์ของโรคในหนูได้เร็วที่สุด 4 สัปดาห์หลังจากเริ่มสัมผัสกับควันบุหรี่มือสามในระดับที่เทียบเท่ากับที่พบในบ้านของผู้สูบบุหรี่" . Clinical Science . 131 (19): 2409– 2426. doi : 10.1042/CS20171053 . PMID 28912356 .
- ^ Dabin J, Fortuny A, Polo SE (มิถุนายน 2016). "การบำรุงรักษาเอพิเจโนมเพื่อตอบสนองต่อความเสียหายของ DNA" . Molecular Cell . 62 (5): 712– 27. doi : 10.1016/j.molcel.2016.04.006 . PMC 5476208 . PMID 27259203 .
- ^ Lövkvist C, Dodd IB, Sneppen K, Haerter JO (มิถุนายน 2016). "การเมทิลเลชั่นของ DNA ในเอพิเจโนมของมนุษย์ขึ้นอยู่กับโทโพโลยีเฉพาะที่ของไซต์ CpG" . Nucleic Acids Research . 44 (11): 5123– 32. doi : 10.1093/nar/gkw124 . PMC 4914085 . PMID 26932361 .
- ^ Jabbari K, Bernardi G (พฤษภาคม 2547). "การเมทิลเลชั่นของไซโตซีนและความถี่ของ CpG, TpG (CpA) และ TpA" Gene . 333 : 143– 9. doi : 10.1016/j.gene.2004.02.043 . PMID 15177689 .
- ^ Kim JJ, Jung MW (2006). "วงจรประสาทและกลไกที่เกี่ยวข้องกับการปรับสภาพความกลัวแบบพาฟลอฟ: การทบทวนเชิงวิพากษ์"วารสารประสาทวิทยาและพฤติกรรมชีวภาพ 30 ( 2): 188– 202. doi : 10.1016/j.neubiorev.2005.06.005 . PMC 4342048 . PMID 16120461 .
- ^ a b c Duke CG, Kennedy AJ, Gavin CF, Day JJ, Sweatt JD (กรกฎาคม 2017). "การจัดระเบียบเอพิเจโนมิกส์ที่ขึ้นอยู่กับประสบการณ์ในฮิปโปแคมปัส" . การเรียนรู้และความจำ . 24 (7): 278– 288. doi : 10.1101/lm.045112.117 . PMC 5473107 . PMID 28620075 .
- ^ Weber M, Hellmann I, Stadler MB, Ramos L, Pääbo S, Rebhan M, Schübeler D (เมษายน 2550). "การกระจายตัว ศักยภาพในการปิดกั้น และผลกระทบเชิงวิวัฒนาการของการเมทิลเลชั่นของดีเอ็นเอโปรโมเตอร์ในจีโนมมนุษย์" Nat . Genet . 39 (4): 457–66 . doi : 10.1038/ng1990 . PMID 17334365. S2CID 22446734 .
- ^ Yang X, Han H, De Carvalho DD, Lay FD, Jones PA, Liang G (ตุลาคม 2014). "การเมทิลเลชั่นของยีนสามารถเปลี่ยนแปลงการแสดงออกของยีนและเป็นเป้าหมายในการรักษาโรคมะเร็ง" Cancer Cell . 26 (4): 577– 90. doi : 10.1016/j.ccr.2014.07.028 . PMC 4224113 . PMID 25263941 .
- ^ Maeder ML, Angstman JF, Richardson ME, Linder SJ, Cascio VM, Tsai SQ, Ho QH, Sander JD, Reyon D, Bernstein BE, Costello JF, Wilkinson MF, Joung JK (ธันวาคม 2013). "การกำจัดหมู่เมทิลออกจาก DNA และการกระตุ้นยีนภายในโดยใช้โปรตีนฟิวชั่น TALE-TET1 ที่ตั้งโปรแกรมได้" Nat . Biotechnol . 31 ( 12): 1137– 42. doi : 10.1038/nbt.2726 . PMC 3858462. PMID 24108092 .
- ^ Ogorodnikov A, Kargapolova Y, Danckwardt S (มิถุนายน 2016). "การประมวลผลและการขยายตัวของทรานสคริปโตมที่ปลาย 3' ของ mRNA ในภาวะสุขภาพและโรค: การค้นหาปลายที่ถูกต้อง" . Pflügers Archiv . 468 (6): 993– 1012. doi : 10.1007/s00424-016-1828-3 . PMC 4893057 . PMID 27220521 .
- ^ miRBase.org
- ^ a b Friedman RC, Farh KK, Burge CB, Bartel DP (มกราคม 2552). "mRNA ของสัตว์เลี้ยงลูกด้วยนมส่วนใหญ่เป็นเป้าหมายที่อนุรักษ์ไว้ของไมโครอาร์เอ็นเอ" . Genome Research . 19 (1): 92– 105. doi : 10.1101/gr.082701.108 . PMC 2612969 . PMID 18955434 .
- ^ Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J และคณะ (กุมภาพันธ์ 2548). "การวิเคราะห์ไมโครอาร์เรย์แสดงให้เห็นว่าไมโครอาร์เอ็นเอบางชนิดลดการแสดงออกของเอ็มอาร์เอ็นเอเป้าหมายจำนวนมาก" Nature . 433 (7027): 769– 73. Bibcode : 2005Natur.433..769L . doi : 10.1038/nature03315 . PMID 15685193 . S2CID 4430576 .
- ^ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (กันยายน 2551). "การเปลี่ยนแปลงอย่างกว้างขวางในการสังเคราะห์โปรตีนที่เกิดจากไมโครอาร์เอ็นเอ" Nature . 455 (7209): 58– 63. Bibcode : 2008Natur.455...58S . doi : 10.1038/nature07228 . PMID 18668040 . S2CID 4429008 .
- ^ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP (กันยายน 2551). "ผลกระทบของไมโครอาร์เอ็นเอต่อผลผลิตโปรตีน" Nature . 455 ( 7209): 64– 71. Bibcode : 2008Natur.455...64B . doi : 10.1038/nature07242 . PMC 2745094 . PMID 18668037 .
- ↑พัลเมโร อีไอ, เด กัมโปส เอสจี, กัมโปส เอ็ม, เด ซูซา เอ็นซี, เกร์เรโร ID, คาร์วัลโญ่ อัล, มาร์เกส เอ็มเอ็ม (กรกฎาคม 2554) "กลไกและบทบาทของการลดการควบคุม microRNA ในการโจมตีและการลุกลามของมะเร็ง " พันธุศาสตร์และอณูชีววิทยา . 34 (3): 363– 70. ดอย : 10.1590/S1415-47572011000300001 . PMC 3168173 . PMID21931505 .
- ^ Bernstein C, Bernstein H (พฤษภาคม 2015). "การลดลงของการซ่อมแซม DNA ทางเอพิเจเนติกส์ในความก้าวหน้าของมะเร็งทางเดินอาหาร" . World Journal of Gastrointestinal Oncology . 7 (5): 30– 46. doi : 10.4251/wjgo.v7.i5.30 . PMC 4434036 . PMID 25987950 .
- ^ Maffioletti E, Tardito D, Gennarelli M, Bocchio-Chiavetto L (2014). "สายลับขนาดเล็กจากสมองสู่ส่วนปลาย: เบาะแสใหม่จากการศึกษาเกี่ยวกับไมโครอาร์เอ็นเอในความผิดปกติทางจิตเวช" . Frontiers in Cellular Neuroscience . 8 : 75. doi : 10.3389/fncel.2014.00075 . PMC 3949217 . PMID 24653674 .
- ^ Mellios N, Sur M (2012). "บทบาทที่กำลังเกิดขึ้นของไมโครอาร์เอ็นเอในโรคจิตเภทและกลุ่มอาการออทิสติก" . Frontiers in Psychiatry . 3 : 39. doi : 10.3389/fpsyt.2012.00039 . PMC 3336189 . PMID 22539927 .
- ^ Geaghan M, Cairns MJ (สิงหาคม 2015). "MicroRNA และการควบคุมผิดปกติหลังการถอดรหัสในจิตเวช"จิตเวชศาสตร์ชีวภาพ78 ( 4): 231– 9. doi : 10.1016/j.biopsych.2014.12.009 . hdl : 1959.13/1335073 . PMID 25636176 .
- ^ Barnett JA (กรกฎาคม 2547). "ประวัติการวิจัยเกี่ยวกับยีสต์ 7: การปรับตัวและการควบคุมเอนไซม์" ยีสต์ 21 ( 9 ) : 703– 46. doi : 10.1002/yea.1113 . PMID 15282797 . S2CID 36606279 .
- ^ Dequéant ML, Pourquié O (พฤษภาคม 2551). "รูปแบบการแบ่งส่วนของแกนตัวอ่อนของสัตว์มีกระดูกสันหลัง" Nature Reviews. Genetics . 9 (5): 370– 82. doi : 10.1038/nrg2320 . PMID 18414404 . S2CID 2526914 .
- ^ Gilbert SF (2003). ชีววิทยาพัฒนาการ ฉบับที่ 7 ซันเดอร์แลนด์ แมสซาชูเซตส์: Sinauer Associates, 65–6. ISBN 0-87893-258-5.
- ^ Nehlin JO, Carlberg M, Ronne H (พฤศจิกายน 1991). "การควบคุมยีน GAL ของยีสต์โดยตัวยับยั้ง MIG1: ลำดับการถอดรหัสในการตอบสนองต่อกลูโคส"วารสารEMBO 10 ( 11): 3373– 7. doi : 10.1002/j.1460-2075.1991.tb04901.x . PMC 453065 . PMID 1915298 .
- ^ Cheadle C, Fan J, Cho-Chung YS, Werner T, Ray J, Do L และคณะ (พฤษภาคม 2548). "การควบคุมการแสดงออกของยีนระหว่างการกระตุ้นเซลล์ T: การควบคุมการถอดรหัส mRNA และความเสถียรของ mRNA ที่แตกต่างกัน" . BMC Genomics . 6 75. doi : 10.1186/1471-2164-6-75 . PMC 1156890 . PMID 15907206 .
- ^ Jackson DA, Pombo A, Iborra F (กุมภาพันธ์ 2000). "งบดุลสำหรับการถอดรหัส: การวิเคราะห์การเผาผลาญ RNA ในนิวเคลียสในเซลล์สัตว์เลี้ยงลูกด้วยนม" . FASEB Journal . 14 (2): 242– 54. doi : 10.1096/fasebj.14.2.242 . PMID 10657981 . S2CID 23518786 .
- ^ Schwanekamp JA, Sartor MA, Karyala S, Halbleib D, Medvedovic M, Tomlinson CR (2006). "การวิเคราะห์ทั่วทั้งจีโนมแสดงให้เห็นว่าระดับ RNA ในนิวเคลียสและไซโตพลาสซึมได้รับผลกระทบจากไดออกซินแตกต่างกัน" Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression . 1759 ( 8– 9): 388– 402. doi : 10.1016/j.bbaexp.2006.07.005 . PMID 16962184 .
บรรณานุกรม
- แลทช์แมน, เดวิด เอส. (2005). การควบคุมยีน: มุมมองจากยูคาริโอต . สำนักพิมพ์จิตวิทยา. ISBN 978-0-415-36510-9.
ลิงก์ภายนอก
- ฐานข้อมูลปัจจัยการถอดรหัสทางพันธุกรรมของพืช และแพลตฟอร์มข้อมูลและการวิเคราะห์การควบคุมการถอดรหัสทางพันธุกรรมของพืช
- การควบคุมการแสดงออกของยีน (MeSH) ที่ หัวข้อทางการแพทย์ (MeSH) ของหอสมุดแห่งชาติสหรัฐอเมริกา
- ChIPBaseคือฐานข้อมูลแบบเปิดสำหรับถอดรหัสเครือข่ายควบคุมการถอดรหัสของ RNA ที่ไม่เข้ารหัสและยีนที่เข้ารหัสโปรตีนจากข้อมูล ChIP-seq
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ การควบคุมการแสดงออกของยีน
การควบคุมการแสดงออกของยีนหรือการควบคุมยีนครอบคลุมกลไกที่หลากหลายซึ่งเซลล์ใช้เพื่อเพิ่มหรือลดการผลิตผลิตภัณฑ์ยีน เฉพาะ ( โปรตีนหรือRNA ) โปรแกรมการแสดงออกของยีน ที่ซับซ้อน...
ขั้นตอนที่ถูกควบคุมของการแสดงออกของยีน
ทุกขั้นตอนของการแสดงออกของยีนสามารถถูกควบคุมได้ ตั้งแต่ การส่งสัญญาณ การ ถอดรหัส ไปจนถึง การ ดัดแปลง โปรตีน หลังการสังเคราะห์ ต่อไปนี้คือรายการขั้นตอนที่การแสดงออกของยีนถูกควบคุม โดยจุดที่ถูกนำมาใช้มากที่สุดคือการเริ่มต้นการถอดรหัส...
การดัดแปลงดีเอ็นเอ
ในยูคาริโอต การเข้าถึงบริเวณขนาดใหญ่ของดีเอ็นเออาจขึ้นอยู่กับ โครงสร้าง โครมาติน ซึ่งสามารถเปลี่ยนแปลงได้อันเป็นผลมาจาก การดัดแปลง ฮิสโตน ที่ควบคุมโดย การเมทิลเลชันของดีเอ็นเอ อา ร์ เอ็นเอที่ไม่เข้ารหัส หรือ โปรตีนที่จับกับดีเอ็นเอ...
โครงสร้าง
การถอดรหัส DNA ถูกกำหนดโดยโครงสร้างของมัน โดยทั่วไป ความหนาแน่นของการบรรจุจะบ่งบอกถึงความถี่ของการถอดรหัส โปรตีนเชิงซ้อนแปดหน่วยที่เรียกว่า ฮิสโตน ร่วมกับส่วนของ DNA ที่พันรอบโปรตีนฮิสโตนทั้งแปด (เรียกรวมกันว่านิวคลีโอโซม) มีหน้าที่รับผิดชอบต่อปริมาณ การขดตัว...