อ่าน 11 นาที
แรด52
RAD52 homolog (S. cerevisiae) หรือ ที่รู้จักกันในชื่อ RAD52 เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน RAD52 [ 5 ] [ 6 ]
แรด52
| แรด52 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | RAD52 , โปรตีนซ่อมแซม DNA ที่มีโครงสร้างคล้ายกับ RAD52 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 600392 ; เอ็มจีไอ : 101949 ; โฮโมโลยีน : 31118 ; การ์ดยีน : RAD52 ; OMA : RAD52 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






RAD52 homolog (S. cerevisiae) หรือที่รู้จักกันในชื่อRAD52เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนRAD52 [ 5 ] [ 6 ]
การทำงาน
โปรตีนที่เข้ารหัสโดยยีนนี้มีความคล้ายคลึงกับRad52 ของSaccharomyces cerevisiae ซึ่งเป็น โปรตีนที่สำคัญสำหรับ การซ่อมแซม DNAสายคู่ที่แตกหักและการรวมตัวแบบโฮโมโลกัสผลิตภัณฑ์ของยีนนี้แสดงให้เห็นว่าสามารถจับกับ ปลาย DNA สายเดี่ยวและเป็นตัวกลางในการโต้ตอบ DNA-DNA ที่จำเป็นสำหรับการจับคู่ของ สาย DNA ที่เสริมกันนอกจากนี้ยังพบว่ามีการโต้ตอบกับโปรตีนการรวมตัวของ DNA RAD51ซึ่งชี้ให้เห็นถึงบทบาทของมันในการรวมตัวและการซ่อมแซม DNA ที่เกี่ยวข้องกับ RAD51 [ 6 ]
มีบทบาทในการซ่อมแซมดีเอ็นเอด้วยการรวมตัวใหม่
RAD52 ทำหน้าที่เป็นตัวกลาง ในการทำงาน ของ RAD51ใน การซ่อมแซม แบบโฮโมโลจัสรีคอมบิเนชัน (HRR) ทั้งในยีสต์Saccharomyces cerevisiaeและในเซลล์สัตว์เลี้ยงลูกด้วยนมของหนูและมนุษย์ อย่างไรก็ตาม โปรตีน RAD52 มีหน้าที่ที่แตกต่างกันอย่างชัดเจนใน HRR ของยีสต์และมนุษย์ ในS. cerevisiaeโปรตีน Rad52 ที่ทำงานเพียงอย่างเดียวจะช่วยอำนวยความสะดวกในการโหลดโปรตีน Rad51 ลงบน DNA สายเดี่ยวที่เคลือบไว้ล่วงหน้าด้วยโปรตีนการจำลองแบบ Aใน ระยะ ก่อน ไซแนปส์ ของรีคอมบิเนชัน[ 7 ] [ 8 ]ในยีสต์ หน้าที่ที่ควบคุมโดย ยีน RAD52มีความสำคัญต่อไมโอซิสปกติ[ 9 ] ดูเหมือนว่าหน้าที่ปกติที่ระบุโดยRAD52นั้นไม่จำเป็นสำหรับขั้นตอนเริ่มต้นหรือขั้นตอนสุดท้ายในไมโอซิส แต่จำเป็นสำหรับการเกิดรีคอมบิเนชันที่ ประสบความสำเร็จ [ 9 ]
อย่างไรก็ตาม ในหนูและมนุษย์ BRCA2 ทำหน้าที่เป็นตัวกลางหลักในการประกอบ RAD51 อย่างเป็นระเบียบบน ssDNA ซึ่งเป็นรูปแบบที่ใช้งานได้สำหรับการจับคู่แบบโฮโมล็อกและการบุกรุกสาย[ 10 ] BRCA2 ยังเปลี่ยนทิศทาง RAD51 จาก dsDNA และป้องกันการแยกตัวออกจาก ssDNA [ 10 ] นอกจากนี้พาราล็อก ทั้งสี่ ของRAD51ซึ่งประกอบด้วย RAD51B ( RAD51L1 ), RAD51C (RAD51L2), RAD51D ( RAD51L3 ) และXRCC2ก่อตัวเป็นคอมเพล็กซ์ที่เรียกว่าคอมเพล็กซ์ BCDX2 คอมเพล็กซ์นี้มีส่วนร่วมในการดึงดูดหรือทำให้ RAD51 มีเสถียรภาพที่บริเวณที่เสียหาย[ 11 ] คอมเพล็กซ์ BCDX2 ดูเหมือนจะทำหน้าที่โดยการอำนวยความสะดวกในการประกอบหรือทำให้เส้นใยโปรตีนนิวคลีโอโปรตีน RAD51 มีเสถียรภาพ อย่างไรก็ตาม ในกรณีที่มีการกลายพันธุ์ของ BRCA2 RAD52 ของมนุษย์สามารถเป็นตัวกลางในการประกอบ RAD51 บน ssDNA และทดแทน BRCA2 ในการซ่อมแซม DNA แบบโฮโมโลจัสรีคอมบิ เนชัน [ 12 ]แม้ว่าจะมีประสิทธิภาพต่ำกว่า BRCA2 ก็ตาม
นอกจากนี้ RAD52 ของมนุษย์ร่วมกับERCC1ยังส่งเสริมเส้นทางการซ่อมแซม DNA ที่เป็นโฮโมโลจัสซึ่งมีแนวโน้มที่จะเกิดข้อผิดพลาดของการเชื่อมต่อสายเดี่ยว[ 13 ]แม้ว่าจะมีแนวโน้มที่จะเกิดข้อผิดพลาด แต่เส้นทางการซ่อมแซมนี้อาจจำเป็นต่อการอยู่รอดของเซลล์ที่มีความเสียหายของ DNA ที่ไม่สามารถซ่อมแซมได้ด้วยวิธีอื่น
RAD52 ของมนุษย์ยังมีบทบาทสำคัญในการซ่อมแซมการแตกของสาย DNA สองสายที่ไซต์การถอดรหัสที่ใช้งานอยู่ระหว่างระยะ G0/G1 ของวงจรเซลล์ การซ่อมแซมการแตกของสาย DNA สองสายเหล่านี้ดูเหมือนจะใช้กลไกการรวมตัวใหม่โดยใช้แม่แบบ RNA ที่ขึ้นอยู่กับ RAD52 [ 14 ]โปรตีน Cockayne Syndrome B (CSB) (เข้ารหัสโดยERCC6 ) จะไปอยู่ที่การแตกของสาย DNA สองสายที่ไซต์การถอดรหัสที่ใช้งานอยู่ ตามด้วยRAD51 , RAD51Cและ RAD52 เพื่อดำเนิน การซ่อมแซม การรวมตัวใหม่ที่เหมือนกันโดยใช้ RNA ที่สังเคราะห์ขึ้นใหม่เป็นแม่แบบ[ 14 ]
ไมโครอาร์เอ็นเอและความเสี่ยงต่อมะเร็ง
บริเวณที่ไม่ถูกแปลรหัสสามส่วนหลัก (3'UTRs) ของอาร์เอ็นเอส่งสาร (mRNA) มักมีลำดับควบคุมที่สามารถทำให้เกิดการยับยั้งการแสดงออกของอาร์ เอ็นเอหลังการถอดรหัสได้ 3'-UTRs เหล่านี้มักมีตำแหน่งสำหรับการ จับกับ ไมโครอาร์เอ็นเอ (miRNA) โดยการจับกับตำแหน่งจำเพาะภายใน 3'-UTR miRNA สามารถลดการแสดงออกของยีนใน mRNA ต่างๆ ได้ ทั้งโดยการยับยั้งการแปลรหัสหรือทำให้เกิดการย่อยสลายของสารถอดรหัสโดยตรง
ไมโครอาร์เอ็นเอ (miRNAs) ดูเหมือนจะควบคุมการแสดงออกของ ยีนที่เข้ารหัสโปรตีนมากกว่า 60% ของจีโนมมนุษย์[ 15 ]ไมโครอาร์เอ็นเอตัวหนึ่งคือmiR-210ยับยั้ง RAD52 [ 16 ] ดังที่ Devlin และคณะได้กล่าวไว้ miR-210 มีการเพิ่มขึ้นในเนื้องอกแข็งส่วนใหญ่และส่งผลเสียต่อผลลัพธ์ทางคลินิก[ 17 ]
บริเวณ 3'-UTR ของ RAD52 ยังมีตำแหน่งการจับกับไมโครอาร์เอ็นเอlet-7ด้วย ผู้หญิงที่มีโพลีมอร์ฟิซึมแบบนิวคลีโอไทด์เดี่ยว (SNP) ในตำแหน่งการจับของ let-7 (rs7963551) ซึ่งทำให้การจับของ let-7 ลดลง มีแนวโน้มที่จะมีการแสดงออกของ RAD52 เพิ่มขึ้น (ดังที่แสดงให้เห็นสำหรับ SNP นี้ในตับ[ 18 ] ) ผู้หญิงที่มี SNP นี้ในบริเวณ 3'UTR ของ RAD52 แสดงให้เห็นถึงความเสี่ยงมะเร็งเต้านมที่ลดลง โดยมีอัตราส่วนความเสี่ยง 0.84 ช่วงความเชื่อมั่น 95% อยู่ที่ 0.75-0.95 [ 19 ]
ในประชากรชาวจีนฮั่น SNP เดียวกันกับข้างต้นใน 3'-UTR ของตำแหน่งการจับของ RAD52 สำหรับ let-7 (rs7963551) ลดความเสี่ยงของเนื้องอกสมอง ความเสี่ยงของเนื้องอกสมองที่เกี่ยวข้องกับจีโนไทป์ RAD52 rs7963551 มีอัตราส่วนความเสี่ยง (เมื่อเทียบกับผู้ที่ไม่มี SNP) เท่ากับ 0.44 สำหรับผู้ที่มีอายุมากกว่า 41 ปี และอัตราส่วนความเสี่ยงเท่ากับ 0.58 สำหรับผู้ที่มีอายุ 41 ปีหรือน้อยกว่า[ 20 ]
Li et al. [ 18 ]พบว่าความเสี่ยงต่อมะเร็งเซลล์ตับลดลงอย่างมีนัยสำคัญในบุคคลที่มีจีโนไทป์ RAD52 rs7963551 CC (SNP เดียวกันกับข้างต้น) เมื่อเทียบกับผู้ที่มีจีโนไทป์ AA ในประชากรชาวจีน นอกจากนี้พวกเขายังพบว่าในตัวอย่างเนื้อเยื่อตับของมนุษย์ปกติ 44 ตัวอย่าง การมีอยู่ของ SNP rs7963551 เกี่ยวข้องกับการเพิ่มขึ้นของการแสดงออกของ mRNA RAD52 อย่างมีนัยสำคัญ
ดังนั้น การแสดงออกของ RAD52 ที่เพิ่มขึ้นจึงช่วยป้องกันมะเร็งหลายชนิด
การศึกษาวิจัยอีกชิ้นหนึ่งเกี่ยวกับตำแหน่งการจับไมโครอาร์เอ็นเอที่เปลี่ยนแปลงไปใน RAD52 และผลกระทบต่อความเสี่ยงต่อการเกิดมะเร็งนั้นดำเนินการโดย Naccarati et al. [ 21 ] พวกเขาพบว่ามีตำแหน่งการจับไมโครอาร์เอ็นเอของ RAD52 สองตำแหน่งที่เปลี่ยนแปลงบ่อยครั้งและมีผลต่อความเสี่ยงต่อมะเร็งลำไส้ใหญ่ บุคคลที่มี SNP แบบโฮโมไซกัสหรือเฮเทโรไซกัสใน rs1051669 มีความเสี่ยงต่อมะเร็งลำไส้ใหญ่เพิ่มขึ้น (OR 1.78, 95% CI 1.13–2.80, p = 0.01 สำหรับโฮโมไซกัส และ OR 1.72, 95% CI 1.10–2.692, p = 0.02 สำหรับเฮเทโรไซกัส) ผู้ที่มีอัลเลลต่างชนิดกันของ SNP RAD52 อีกตัวหนึ่ง (rs11571475) มีความเสี่ยงต่อมะเร็งลำไส้ใหญ่ลดลง (OR 0.76, 95% CI 0.58–1.00, p = 0.05) จากยีน 21 ยีนใน วิถีการซ่อมแซม แบบโฮโมโลจัสรีคอมบิเนชันและยีน 7 ยีนใน วิถี การเชื่อมต่อปลายแบบไม่โฮโมโลจัสที่ได้รับการตรวจสอบ พบว่า SNP เพียงสองตัวในบริเวณการจับของไมโครอาร์เอ็นเอที่มีความถี่สูงพอที่จะประเมินได้และส่งผลต่อความเสี่ยงของมะเร็งลำไส้ใหญ่ คือ SNP สองตัวใน RAD52 และหนึ่งตัวใน MRE11A
ความเสียหายของ DNA ดูเหมือนจะเป็นสาเหตุหลักที่อยู่เบื้องหลังโรคมะเร็ง[ 22 ]และความบกพร่องในการซ่อมแซม DNA ดูเหมือนจะเป็นพื้นฐานของมะเร็งหลายชนิด[ 23 ] หากการซ่อมแซม DNA บกพร่อง ความเสียหายของ DNA มีแนวโน้มที่จะสะสมมากขึ้น ความเสียหายของ DNA ที่มากเกินไปอาจเพิ่ม ข้อผิดพลาดใน การ กลายพันธุ์ ระหว่างการจำลอง DNAเนื่องจากการสังเคราะห์แบบทรานส์ เลสที่ผิดพลาด ความเสียหายของ DNA ที่มากเกินไปอาจเพิ่ม การเปลี่ยนแปลง ทางเอพิเจเนติกส์เนื่องจากข้อผิดพลาดระหว่างการซ่อมแซม DNA [ 24 ] [ 25 ]การกลายพันธุ์และการเปลี่ยนแปลงทางเอพิเจเนติกส์ดังกล่าวอาจก่อให้เกิดมะเร็งการเพิ่มขึ้นหรือความบกพร่องของ การซ่อมแซม DNA ที่เกิดจาก RAD52 ที่เกิดจากไมโครอาร์เอ็นเอบ่อยครั้ง เนื่องจากการเปลี่ยนแปลงการจับของไมโครอาร์เอ็นเอมีแนวโน้มที่จะมีส่วนช่วยในการป้องกันหรือการลุกลามของมะเร็งเต้านม สมอง ตับ หรือลำไส้ใหญ่
ปฏิสัมพันธ์
RAD52 ได้รับการแสดงให้เห็นว่าโต้ตอบกับ RAD51 [ 26 ] Rad52 จะช่วยให้การโหลด Rad51 บน ssDNA ง่ายขึ้นโดยการรบกวนโปรตีน RPA
การเสริมยีนภายใน
เมื่อพอลิเปปไทด์หลายชุดที่เข้ารหัสโดยยีนรวมตัวกันเป็นกลุ่ม โปรตีนโครงสร้างนี้เรียกว่ามัลติเมอร์ เมื่อมัลติเมอร์เกิดขึ้นจากพอลิเปปไทด์ที่ผลิตโดย อัลลี ลกลายพันธุ์ สองแบบที่แตกต่างกัน ของยีนเฉพาะ มัลติเมอร์แบบผสมอาจแสดงกิจกรรมการทำงานที่มากกว่ามัลติเมอร์ที่ไม่ผสมซึ่งเกิดจากอัลลีลกลายพันธุ์แต่ละแบบเพียงอย่างเดียว ในกรณีเช่นนี้ ปรากฏการณ์นี้เรียกว่าการเสริมกันภายในยีน อัลลีล กลาย พันธุ์ RAD52 ของSaccharomyces cerevisiaeที่แสดงโปรตีนที่ถูกตัดปลาย C-terminal พบว่าสามารถเสริมกันกับอัลลีลกลายพันธุ์มิสเซนส์ RAD52 อื่นๆ ได้[ 27 ] การค้นพบการเสริมกันภายในยีน นี้ชี้ให้เห็นว่าโปรตีน RAD52 มีโครงสร้างมัลติเมอร์ที่ช่วยให้เกิดปฏิสัมพันธ์แบบร่วมมือกันระหว่างโมโนเมอร์ที่เป็นส่วนประกอบ
อ่านเพิ่มเติม
- Muris DF, Bezzubova O, Buerstedde JM, Vreeken K, Balajee AS, Osgood CJ, Troelstra C, Hoeijmakers JH, Ostermann K, Schmidt H (พ.ย. 1994). "การโคลนยีนของมนุษย์และหนูที่มีความคล้ายคลึงกับ RAD52 ซึ่งเป็นยีนของยีสต์ที่เกี่ยวข้องกับการซ่อมแซมและการรวมตัวใหม่ของ DNA" . Mutation Research . 315 (3): 295– 305. doi : 10.1016/0921-8777(94)90040-x . hdl : 1765/3072 . PMID 7526206 .
- Shen Z, Denison K, Lobb R, Gatewood JM, Chen DJ (ม.ค. 1995). "โฮโมล็อกของมนุษย์และหนูของยีน RAD52 ของยีสต์: การโคลน cDNA การวิเคราะห์ลำดับ การกำหนดตำแหน่งบนโครโมโซมมนุษย์ 12p12.2-p13 และการแสดงออกของ mRNA ในเนื้อเยื่อของหนู" Genomics . 25 ( 1): 199– 206. doi : 10.1016/0888-7543(95)80126-7 . PMID 7774919 .
- Park MS (มิถุนายน 1995). "การแสดงออกของ RAD52 ในมนุษย์ทำให้เซลล์สัตว์เลี้ยงลูกด้วยนมต้านทานต่อรังสีไอออน"วารสารเคมีชีวภาพ 270 ( 26): 15467– 70. doi : 10.1074/jbc.270.26.15467 . PMID 7797537 .
- Shen Z, Pardington-Purtymun PE, Comeaux JC, Moyzis RK, Chen DJ (ก.ย. 1996). "UBL1 โปรตีนคล้ายยูบิควิตินของมนุษย์ที่เชื่อมโยงกับโปรตีน RAD51/RAD52 ของมนุษย์" Genomics 36 ( 2 ): 271– 9. doi : 10.1006/geno.1996.0462 . PMID 8812453 .
- Shen Z, Pardington-Purtymun PE, Comeaux JC, Moyzis RK, Chen DJ (ต.ค. 1996). "การเชื่อมโยงของ UBE2I กับโปรตีน RAD52, UBL1, p53 และ RAD51 ในระบบยีสต์ทูไฮบริด" Genomics 37 ( 2 ): 183– 6. doi : 10.1006/geno.1996.0540 . PMID 8921390 .
- Chen G, Yuan SS, Liu W, Xu Y, Trujillo K, Song B, Cong F, Goff SP, Wu Y, Arlinghaus R, Baltimore D, Gasser PJ, Park MS, Sung P, Lee EY (เมษายน 1999). "การประกอบคอมเพล็กซ์การรวมตัวใหม่ของ Rad51 และ Rad52 ที่เกิดจากการฉายรังสีต้องอาศัย ATM และ c-Abl" (PDF)วารสารเคมีชีวภาพ274 (18): 12748– 52. doi : 10.1074 /jbc.274.18.12748 . PMID 10212258 . S2CID 2587580 .
- Kito K, Wada H, Yeh ET, Kamitani T (ธ.ค. 1999) "การระบุไอโซฟอร์มใหม่ของ RAD52 ของมนุษย์" Biochimica และ Biophysica Acta (BBA) - โครงสร้างและการแสดงออกของยีน . 1489 ( 2– 3): 303– 14. ดอย : 10.1016/s0167-4781(99)00214-6 . PMID 10673031 .
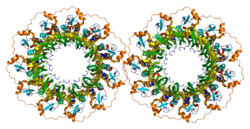
- Stasiak AZ, Larquet E, Stasiak A, Müller S, Engel A, Van Dyck E, West SC, Egelman EH (มีนาคม 2000). "โปรตีน Rad52 ของมนุษย์มีอยู่เป็นวงแหวนเจ็ดหน่วย" Current Biology . 10 (6): 337– 40. Bibcode : 2000CBio...10..337S . doi : 10.1016/S0960-9822(00)00385-7 . PMID 10744977 . S2CID 16087838 .
- Parsons CA, Baumann P, Van Dyck E, West SC (สิงหาคม 2000). "การจับปลาย DNA สายเดี่ยวอย่างแม่นยำโดยโปรตีน RAD52 ของมนุษย์"วารสารEMBO . 19 (15): 4175– 81. doi : 10.1093/emboj/19.15.4175 . PMC 306603 . PMID 10921897 .
- Mer G, Bochkarev A, Gupta R, Bochkareva E, Frappier L, Ingles CJ, Edwards AM, Chazin WJ (ต.ค. 2000). "โครงสร้างพื้นฐานสำหรับการจดจำโปรตีนซ่อมแซม DNA UNG2, XPA และ RAD52 โดยปัจจัยการจำลองแบบ RPA" . Cell . 103 (3): 449– 56. doi : 10.1016/S0092-8674(00)00136-7 . PMID 11081631 . S2CID 16640087 .
- Ranatunga W , Jackson D, Flowers II RA, Borgstahl GE (กรกฎาคม 2544). "โปรตีน RAD52 ของมนุษย์มีเสถียรภาพทางความร้อนสูงมาก". ชีวเคมี40 (29): 8557– 62. CiteSeerX 10.1.1.566.9388 . doi : 10.1021/bi0155089 . PMID 11456495 .
- Van Dyck E, Stasiak AZ, Stasiak A, West SC (ต.ค. 2001). "การแสดงภาพของตัวกลางการรวมตัวใหม่ที่สร้างขึ้นโดยการเชื่อมต่อสายเดี่ยวที่ควบคุมโดย RAD52" EMBO Reports . 2 (10): 905– 9. doi : 10.1093/embo-reports/kve201 . PMC 1084079 . PMID 11571269 .
- Kim PM, Allen C, Wagener BM, Shen Z, Nickoloff JA (พ.ย. 2001). "การแสดงออกมากเกินไปของ RAD51 และ RAD52 ของมนุษย์ช่วยลดการรวมตัวแบบโฮโมโลกัสที่เกิดจากการแตกของสายคู่ในเซลล์สัตว์เลี้ยงลูกด้วยนม" . Nucleic Acids Research . 29 (21): 4352– 60. doi : 10.1093/nar/29.21.4352 . PMC 60192 . PMID 11691922 .
- Yáñez RJ, Porter AC (กุมภาพันธ์ 2545). "ผลกระทบที่แตกต่างกันของการแสดงออกเกินของ Rad52p ต่อการกำหนดเป้าหมายยีนและการรวมตัวของโฮโมโลจัสนอกโครโมโซมในสายเซลล์มนุษย์" . Nucleic Acids Research . 30 (3): 740– 8. doi : 10.1093/nar/30.3.740 . PMC 100286 . PMID 11809887 .
- Jackson D, Dhar K, Wahl JK, Wold MS, Borgstahl GE (สิงหาคม 2545). "การวิเคราะห์โปรตีนการจำลองแบบของมนุษย์ A:คอมเพล็กซ์ Rad52: หลักฐานสำหรับการสื่อสารข้ามระหว่าง RPA32, RPA70, Rad52 และ DNA" วารสารชีววิทยาโมเลกุล 321 ( 1): 133– 48. doi : 10.1016/S0022-2836(02)00541-7 . PMID 12139939 .
- Kagawa W, Kurumizaka H, Ishitani R, Fukai S, Nureki O, Shibata T, Yokoyama S (สิงหาคม 2545). "โครงสร้างผลึกของโดเมนการจับคู่แบบโฮโมโลกัสจากรีคอมบิเนส Rad52 ของมนุษย์ในรูปแบบอันเดคาเมอริก" . Molecular Cell . 10 (2): 359– 71. doi : 10.1016/S1097-2765(02)00587-7 . PMID 12191481 .
- Singleton MR, Wentzell LM, Liu Y, West SC, Wigley DB (ตุลาคม 2545). "โครงสร้างของโดเมนการเชื่อมต่อสายเดี่ยวของโปรตีน RAD52 ของมนุษย์" Proceedings of the National Academy of Sciences of the United States of America . 99 (21): 13492– 7. Bibcode : 2002PNAS...9913492S . doi : 10.1073/pnas.212449899 . PMC 129701. PMID 12370410 .
- Liu J, Meng X, Shen Z (ต.ค. 2545). "การเชื่อมโยงของโปรตีน RAD52 ของมนุษย์กับปัจจัยการถอดรหัส". Biochemical and Biophysical Research Communications . 297 (5): 1191– 6. doi : 10.1016/S0006-291X(02)02353-7 . PMID 12372413 .
- Han J, Hankinson SE, De Vivo I, Colditz GA, Hunter DJ (ต.ค. 2002). "ไม่มีความสัมพันธ์ระหว่างโพลีมอร์ฟิซึมของรหัสหยุดใน RAD52 กับความเสี่ยงมะเร็งเต้านม" Cancer Epidemiology, Biomarkers & Prevention . 11 (10 Pt 1): 1138– 9. PMID 12376524 .
- Kitao H, Yuan ZM (ธันวาคม 2002). "การควบคุมการก่อตัวของจุดโฟกัส Rad52 ในนิวเคลียสที่เกิดจากรังสีไอออนไนซ์โดยการฟอสโฟรีเลชันที่เกิดจาก c-Abl"วารสารเคมีชีวภาพ 277 ( 50): 48944– 8. doi : 10.1074/jbc.M208151200 . PMID 12379650 .
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ แรด52
RAD52 homolog (S. cerevisiae) หรือ ที่รู้จักกันในชื่อ RAD52 เป็น โปรตีน ที่ในมนุษย์ถูกเข้ารหัสโดย ยีน RAD52 [ 5 ] [ 6 ]
การทำงาน
โปรตีนที่เข้ารหัสโดยยีนนี้มีความคล้ายคลึงกับRad52 ของ Saccharomyces cerevisiae ซึ่งเป็น โปรตีน ที่สำคัญสำหรับ การซ่อมแซม DNA สายคู่ที่แตกหักและ การรวมตัวแบบโฮโมโลกัส ผลิตภัณฑ์ของยีนนี้แสดงให้เห็นว่าสามารถจับกับ ปลาย DNA สายเดี่ยว และเป็นตัวกลางในการโต้ตอบ...
มีบทบาทในการซ่อมแซมดีเอ็นเอด้วยการรวมตัวใหม่
RAD52 ทำหน้าที่เป็นตัวกลาง ในการทำงาน ของ RAD51 ใน การซ่อมแซม แบบโฮโมโลจัสรีคอมบิ เนชัน (HRR) ทั้งในยีสต์ Saccharomyces cerevisiae และในเซลล์สัตว์เลี้ยงลูกด้วยนมของหนูและมนุษย์ อย่างไรก็ตาม โปรตีน RAD52 มีหน้าที่ที่แตกต่างกันอย่างชัดเจนใน HRR...
ไมโครอาร์เอ็นเอและความเสี่ยงต่อมะเร็ง
บริเวณที่ไม่ถูกแปลรหัสสามส่วนหลัก (3'UTRs) ของ อาร์เอ็นเอส่งสาร (mRNA) มักมีลำดับควบคุมที่สามารถทำให้เกิด การยับยั้งการแสดงออกของอาร์ เอ็นเอหลังการถอดรหัสได้ 3'-UTRs เหล่านี้มักมีตำแหน่งสำหรับการ จับกับ ไมโครอาร์เอ็นเอ (miRNA) โดยการจับกับตำแหน่งจำเพาะภายใน...