อ่าน 19 นาที
โครงสร้างตติยภูมิของกรดนิวคลีอิก
โครงสร้างตติยภูมิของกรดนิวคลีอิก คือ รูปร่างสามมิติ ของพอลิเมอร์ กรดนิวคลีอิก [ 1 ] โมเลกุล RNA และ DNA มีความสามารถในการทำงานที่หลากหลาย ตั้งแต่ การจดจำโมเลกุล ไปจนถึง...
โครงสร้างตติยภูมิของกรดนิวคลีอิก

โครงสร้างตติยภูมิของกรดนิวคลีอิกคือรูปร่างสามมิติของพอลิเมอร์กรดนิวคลีอิก[ 1 ]โมเลกุลRNAและDNA มีความสามารถในการทำงานที่หลากหลาย ตั้งแต่การจดจำโมเลกุลไปจนถึงการเร่งปฏิกิริยาการทำงานดังกล่าวต้องการโครงสร้างสามมิติที่แม่นยำ แม้ว่าโครงสร้างดังกล่าวจะมีความหลากหลายและดูซับซ้อน แต่ก็ประกอบด้วยรูปแบบโครงสร้าง ตติยภูมิที่เกิดขึ้นซ้ำๆ ซึ่งสามารถจดจำได้ง่าย และทำหน้าที่เป็นหน่วยสร้างโมเลกุล รูปแบบที่พบได้บ่อยที่สุดบางส่วนสำหรับโครงสร้างตติยภูมิของ RNA และ DNA ได้รับการอธิบายไว้ด้านล่าง แต่ข้อมูลนี้อิงจากโครงสร้างที่ได้รับการแก้ไขจำนวนจำกัด รูปแบบโครงสร้างตติยภูมิอีกมากมายจะถูกเปิดเผยเมื่อโมเลกุล RNA และ DNA ใหม่ๆ ได้รับการระบุลักษณะโครงสร้าง
โครงสร้างเกลียว
เกลียวคู่

โครงสร้างเกลียวคู่เป็นโครงสร้างตติยภูมิที่เด่นสำหรับดีเอ็นเอทางชีวภาพ และยังเป็นโครงสร้างที่เป็นไปได้สำหรับอาร์เอ็นเอ เชื่อกันว่ามีโครงสร้างดีเอ็นเอสามแบบที่พบในธรรมชาติ ได้แก่A-DNA , B-DNAและZ-DNAเชื่อกันว่ารูปแบบ "B" ที่อธิบายโดยJames D. WatsonและFrancis Crick เป็นรูปแบบที่เด่นในเซลล์ [ 2 ] James D. WatsonและFrancis Crickอธิบายโครงสร้างนี้ว่าเป็นเกลียวคู่ที่มีรัศมี 10 Åและระยะห่างระหว่างเกลียว 34 Åโดยหมุนครบหนึ่งรอบรอบแกนทุกๆ 10 คู่เบสของลำดับ[ 3 ]เกลียวคู่หมุนครบหนึ่งรอบรอบแกนทุกๆ 10.4–10.5 คู่เบสในสารละลาย ความถี่ของการบิดนี้ (เรียกว่าระยะห่าง ระหว่างเกลียว ) ขึ้นอยู่กับแรงเรียงซ้อนที่แต่ละเบสกระทำต่อเบสข้างเคียงในสายโซ่เป็นส่วนใหญ่ อาร์เอ็นเอแบบเกลียวคู่มีโครงสร้างคล้ายกับโครงสร้างแบบ A
โครงสร้างอื่นๆ ก็เป็นไปได้เช่นกัน อันที่จริง ปัจจุบันมีเพียงตัวอักษร F, Q, U, V และ Y เท่านั้นที่สามารถใช้อธิบายโครงสร้าง DNA ใหม่ใดๆ ที่อาจปรากฏขึ้นในอนาคตได้[ 4 ] [ 5 ]อย่างไรก็ตาม รูปแบบส่วนใหญ่เหล่านี้ถูกสร้างขึ้นโดยสังเคราะห์และไม่พบในระบบชีวภาพที่เกิดขึ้นตามธรรมชาติ
ไตรเพล็กซ์ร่องเมเจอร์และไมเนอร์


โครงสร้างแบบไตร เพล็กซ์ในร่องเล็ก (minor groove triplex) เป็นโครงสร้างโมทีฟของ RNA ที่พบได้ทั่วไป เนื่องจากการโต้ตอบกับร่องเล็กมักเกิดขึ้นโดยผ่านหมู่ 2'-OH ของ น้ำตาล ไรโบส โครงสร้างโมทีฟของ RNA นี้จึงดูแตกต่างจากโครงสร้างโมทีฟ ที่เทียบเท่าใน DNAตัวอย่างที่พบได้บ่อยที่สุดของโครงสร้างแบบไตรเพล็กซ์ในร่องเล็กคือ โมทีฟ A-minor หรือการแทรก เบส อะดีโนซีน เข้าไปในร่องเล็ก (ดูด้านบน) อย่างไรก็ตาม โมทีฟนี้ไม่ได้จำกัดเฉพาะอะดีโนซีนเท่านั้น เนื่องจากมีการสังเกตพบว่า นิวคลีโอเบสอื่นๆก็มีปฏิสัมพันธ์กับร่องเล็กของ RNA ด้วยเช่นกัน
ร่องเล็กเป็นส่วนประกอบที่เกือบสมบูรณ์แบบสำหรับเบสที่แทรกเข้าไป ซึ่งช่วยให้เกิดการสัมผัสแบบแวนเดอร์วาลส์ ที่เหมาะสม พันธะไฮโดรเจนที่กว้างขวางและ การฝังตัวของพื้นผิว ที่ไม่ชอบน้ำและสร้างปฏิสัมพันธ์ที่เอื้อต่อพลังงานสูง[ 8 ] [ 9 ] เนื่องจากสามเบสในร่องเล็กสามารถบรรจุลูปและเกลียวอิสระได้อย่างมั่นคง จึงเป็นองค์ประกอบสำคัญในโครงสร้างของไรโบนิวคลีโอไทด์ ขนาดใหญ่ รวมถึงอินทรอนกลุ่ม I [ 10 ]อินทรอนกลุ่ม II [ 11 ]และไร โบโซม


แม้ว่าร่องหลักของ RNA รูปแบบ A มาตรฐานจะค่อนข้างแคบและจึงมีโอกาสเกิดปฏิกิริยาแบบไตรเพล็กซ์น้อยกว่าร่องรอง แต่ก็สามารถสังเกตปฏิกิริยาแบบไตรเพล็กซ์ในร่องหลักได้ในโครงสร้าง RNA หลายแบบ โครงสร้างเหล่านี้ประกอบด้วยการรวมกันของคู่เบสและปฏิกิริยาแบบฮูกสทีนหลายแบบ ตัวอย่างเช่น ไตรเพล็กซ์ GGC (GGC อะมิโน(N-2)-N-7, อิมิโน-คาร์บอนิล, คาร์บอนิล-อะมิโน(N-4); วัตสัน-คริก) ที่พบในไรโบโซม 50Sประกอบด้วยคู่ GC แบบวัตสัน-คริกและ G ที่เข้ามาซึ่งก่อให้เกิดเครือข่ายแบบซูโด-ฮูกสทีนของปฏิกิริยาพันธะไฮโดรเจนระหว่างเบสทั้งสองที่เกี่ยวข้องกับการจับคู่แบบแคนอนิก[ 12 ]ตัวอย่างที่น่าสนใจอื่นๆ ของไตรเพล็กซ์ร่องหลัก ได้แก่ (i) แกนเร่งปฏิกิริยาของอินทรอนกลุ่ม IIที่แสดงในรูปด้านซ้าย[ 6 ] (ii) เกลียวสามชั้น ที่จำเป็นต่อการเร่งปฏิกิริยา ที่พบในRNA เทโลเมอเรส ของมนุษย์ [ 7 ] (iii) ไรโบสวิตช์ SAM-II [ 14 ]และ (iv) องค์ประกอบสำหรับการแสดงออกในนิวเคลียส (ENE) ซึ่งทำหน้าที่เป็นองค์ประกอบในการทำให้ RNA มีเสถียรภาพผ่านการสร้างเกลียวสามชั้นกับหางโพลี(A) [ 15 ] [ 16 ]
ดีเอ็นเอแบบสามสายยังสามารถเกิดขึ้นได้จากพันธะไฮโดรเจนแบบ Hoogsteen หรือแบบ Hoogsteen กลับด้านในร่องหลักของดีเอ็นเอ แบบ B-form
ควอดรูเพล็กซ์
นอกจากเกลียวคู่และเกลียวสามชั้นที่กล่าวถึงข้างต้นแล้วRNAและDNAยังสามารถสร้างเกลียวสี่ชั้นได้อีกด้วย โครงสร้างของควอดรูเพล็กซ์ของเบส RNA มีความหลากหลาย โมเลกุลกัวนีนสี่โมเลกุล ที่เรียงต่อกัน สามารถสร้างควอดรูเพล็กซ์ใน RNA ได้ด้วย พันธะไฮโดรเจนแบบ Hoogsteenเพื่อสร้าง "วงแหวน Hoogsteen" (ดูรูป) [ 12 ] คู่ GC และ AU ยังสามารถสร้างควอดรูเพล็กซ์ของเบสได้ด้วย การรวมกันของ การจับคู่แบบ Watson-Crickและการจับคู่แบบไม่เป็นไปตามแบบแผนในร่องเล็ก[ 17 ]
แกนกลางของแอปทาเมอร์ สีเขียวมาลาไคต์ ยังเป็นควอดรูเพล็กซ์ฐานชนิดหนึ่งที่มีรูปแบบพันธะไฮโดรเจนที่แตกต่างกัน (ดูรูป) [ 13 ]ควอดรูเพล็กซ์สามารถทำซ้ำได้หลายครั้งติดต่อกัน ทำให้เกิดโครงสร้างที่มีเสถียรภาพอย่างมาก
โครงสร้างเฉพาะของบริเวณควอดรูเพล็กซ์ใน RNA อาจทำหน้าที่ต่างๆ ในระบบชีวภาพได้ สองหน้าที่สำคัญคือ ศักยภาพในการจับกับลิแกนด์หรือโปรตีน และความสามารถในการทำให้โครงสร้างตติยภูมิ ทั้งหมด ของ DNA หรือ RNA มีเสถียรภาพ โครงสร้างที่แข็งแรงสามารถยับยั้งหรือปรับเปลี่ยนการถอดรหัสและการจำลองแบบได้ เช่น ในเทโลเมียร์ของโครโมโซมและUTRของ mRNA [ 18 ] เอกลักษณ์ของเบสมีความสำคัญต่อการจับกับลิแกนด์ โดยทั่วไป G-quartet จะจับกับแคตไอออนที่มีประจุเดียว เช่น โพแทสเซียม ในขณะที่เบสอื่นๆ สามารถจับกับลิแกนด์อื่นๆ ได้มากมาย เช่น ไฮโปแซนทีนในควอดรูเพล็กซ์ UUCU [ 17 ]
นอกเหนือจากหน้าที่เหล่านี้แล้วG-quadruplexใน mRNA รอบบริเวณที่ไรโบโซมจับยังสามารถทำหน้าที่เป็นตัวควบคุมการแสดงออกของยีนในแบคทีเรียได้[ 19 ]อาจมีโครงสร้างและหน้าที่ที่น่าสนใจอีกมากมายที่ยังไม่ถูกค้นพบในร่างกาย
การเรียงซ้อนแบบโคแอกเซียล

การเรียงซ้อนแบบโคแอกเซียล หรือที่รู้จักกันในชื่อการเรียงซ้อนแบบเกลียว เป็นตัวกำหนดโครงสร้างตติยภูมิของ RNA ระดับสูงที่สำคัญ การเรียงซ้อนแบบโคแอกเซียลเกิดขึ้นเมื่อคู่ RNA สองคู่ก่อตัวเป็นเกลียวต่อเนื่อง ซึ่งมีความเสถียรโดยการเรียงซ้อนของเบสที่ส่วนต่อประสานของเกลียวทั้งสอง การเรียงซ้อนแบบโคแอกเซียลถูกสังเกตในโครงสร้างผลึกของ tRNAPhe [ 21 ]เมื่อไม่นานมานี้ การเรียงซ้อนแบบโคแอกเซียลได้รับการสังเกตในโครงสร้างระดับสูงของ ไร โบไซม์หลายชนิด รวมถึง อินทรอน กลุ่ม Iและกลุ่ม IIที่สามารถตัดต่อตัวเองได้หลายรูปแบบ รูปแบบการเรียงซ้อนแบบโคแอกเซียลทั่วไป ได้แก่ ปฏิสัมพันธ์ แบบ kissing loopและpseudoknotความเสถียรของปฏิสัมพันธ์เหล่านี้สามารถทำนายได้โดยการดัดแปลง "กฎของเทอร์เนอร์" [ 22 ] [ 23 ]
ในปี 1994 Walter และ Turner ได้กำหนดส่วนประกอบพลังงานอิสระของปฏิสัมพันธ์การเรียงซ้อนของเพื่อนบ้านที่ใกล้ที่สุดภายในส่วนต่อประสานเฮลิกซ์-เฮลิกซ์โดยใช้ระบบแบบจำลองที่สร้างส่วนต่อประสานเฮลิกซ์-เฮลิกซ์ระหว่างโอลิโกเมอร์ สั้น และส่วนที่ยื่นออกมาสี่ นิว คลีโอไท ด์ ที่ปลายก้านแฮร์พินการทดลองของพวกเขายืนยันว่าส่วนประกอบทางอุณหพลศาสตร์ของการเรียงซ้อนของเบสระหว่างโครงสร้างทุติยภูมิแบบเฮลิกซ์สองโครงสร้างเลียนแบบอุณหพลศาสตร์ของการก่อตัวของดูเพล็กซ์มาตรฐานอย่างใกล้ชิด (ปฏิสัมพันธ์เพื่อนบ้านที่ใกล้ที่สุดทำนายความเสถียรทางอุณหพลศาสตร์ของเฮลิกซ์ที่เกิดขึ้น) ความเสถียรสัมพัทธ์ของปฏิสัมพันธ์เพื่อนบ้านที่ใกล้ที่สุดสามารถใช้ในการทำนายการเรียงซ้อนแบบโคแอกเซียลที่เหมาะสมโดยอิงจากโครงสร้างทุติยภูมิที่ทราบ Walter และ Turner พบว่าโดยเฉลี่ยแล้ว การทำนายโครงสร้าง RNA มีความแม่นยำเพิ่มขึ้นจาก 67% เป็น 74% เมื่อรวมส่วนประกอบการเรียงซ้อนแบบโคแอกเซียลเข้าไปด้วย[ 24 ]
โครงสร้างตติยภูมิของ RNA ที่ได้รับการศึกษาอย่างดีส่วนใหญ่มีตัวอย่างของการเรียงซ้อนแบบแกนร่วม ตัวอย่างที่โดดเด่นบางส่วน ได้แก่ tRNA-Phe, อินทรอนกลุ่ม I, อินทรอนกลุ่ม II และ RNA ไรโบโซม โครงสร้างผลึกของ tRNA เผยให้เห็นการมีอยู่ของเกลียวสองอันที่ยืดออกซึ่งเป็นผลมาจากการเรียงซ้อนแบบแกนร่วมของก้านรับกรดอะมิโนกับแขน T และการเรียงซ้อนของแขน D และแขนแอนติโคดอน ปฏิสัมพันธ์เหล่านี้ภายในtRNAทำให้ก้านแอนติโคดอนตั้งฉากกับก้านกรดอะมิโน นำไปสู่โครงสร้างตติยภูมิรูปตัว L ที่ใช้งานได้[ 21 ]ในอินทรอนกลุ่ม I พบว่าเกลียว P4 และ P6 เรียงซ้อนแบบแกนร่วมโดยใช้การผสมผสานระหว่างวิธีการทางชีวเคมี[ 25 ]และวิธีการทางผลึกศาสตร์ โครงสร้างผลึก P456 ให้มุมมองโดยละเอียดว่าการเรียงซ้อนแบบแกนร่วมทำให้การบรรจุเกลียว RNA เข้าสู่โครงสร้างตติยภูมิมีเสถียรภาพได้อย่างไร[ 26 ]ในอินทรอนกลุ่ม II ที่ตัดต่อตัวเองจาก Oceanobacillus iheyensis ลำต้น IA และ IB เรียงซ้อนกันแบบแกนร่วมและมีส่วนช่วยในการวางแนวสัมพัทธ์ของเกลียวที่เป็นส่วนประกอบของจุดเชื่อมต่อห้าทาง[ 6 ]การวางแนวนี้ช่วยให้เกิดการพับที่เหมาะสมของบริเวณที่ใช้งานของไรโบไซม์ที่ทำงานได้ ไรโบโซมมีตัวอย่างมากมายของการเรียงซ้อนกันแบบแกนร่วม รวมถึงส่วนที่เรียงซ้อนกันยาวถึง 70 bp [ 27 ]
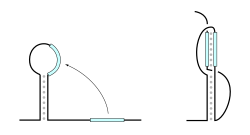
รูปแบบทั่วไปสองแบบที่เกี่ยวข้องกับการเรียงซ้อนแบบแกนร่วมคือ ลูปจูบและปมเทียม ในปฏิกิริยาลูปจูบ บริเวณลูปสายเดี่ยวของแฮร์พินสองตัวจะโต้ตอบกันผ่านการจับคู่เบส ก่อให้เกิดเกลียวแบบเรียงซ้อนแกนร่วมที่ซับซ้อน ที่น่าสังเกตคือ โครงสร้างนี้ทำให้นิวคลีโอไทด์ทั้งหมดในแต่ละลูปมีส่วนร่วมในการจับคู่เบสและปฏิกิริยาการเรียงซ้อน รูปแบบนี้ได้รับการมองเห็นและศึกษาโดยใช้การวิเคราะห์ NMR โดย Lee และ Crothers [ 28 ]รูปแบบปมเทียมเกิดขึ้นเมื่อบริเวณสายเดี่ยวของลูปแฮร์พินจับคู่เบสกับลำดับต้นน้ำหรือปลายน้ำภายในสาย RNA เดียวกัน บริเวณคู่สองบริเวณที่เกิดขึ้นมักจะเรียงซ้อนกัน ก่อให้เกิดเกลียวแบบเรียงซ้อนแกนร่วมที่เสถียร ตัวอย่างหนึ่งของรูปแบบปมเทียมคือ ไรโบไซม์ ของไวรัสตับอักเสบเดลต้าที่ มีความเสถียรสูง ซึ่งโครงสร้างหลักแสดงให้เห็นถึงโทโพโลยีปมเทียมคู่โดยรวม[ 29 ]
ได้มีการสังเกตพบผลที่คล้ายกับการเรียงซ้อนแบบโคแอกเซียลในโครงสร้าง DNA ที่ออกแบบอย่างมีเหตุผล โครงสร้าง DNA origamiประกอบด้วยเกลียวคู่จำนวนมากที่มีปลายทู่ที่เปิดเผย โครงสร้างเหล่านี้ถูกสังเกตว่าเกาะติดกันตามขอบที่มีปลายทู่ที่เปิดเผยเหล่านี้ เนื่องมาจากปฏิกิริยาการเรียงซ้อนแบบไฮโดรโฟบิก[ 30 ]ด้วยการรวมโครงสร้างนาโน DNA ที่ออกแบบอย่างมีเหตุผลเหล่านี้เข้ากับการถ่ายภาพความละเอียดสูง DNA-PAINT นักวิจัยสามารถแยกแยะความแข็งแรงของพลังงานการเรียงซ้อนระหว่างไดนิวคลีโอไทด์ที่เป็นไปได้ทั้งหมดได้[ 31 ]
การวัดการเรียงซ้อนแบบแกนร่วมในกรดนิวคลีอิก
การวัดการเรียงซ้อนแบบโคแอกเซียลในระยะแรกนั้นดำเนินการโดยใช้การทดสอบทางชีวเคมีที่ศึกษาการเคลื่อนที่สัมพัทธ์ของโมเลกุลกรดนิวคลีอิกที่แตกต่างกันโดยพิจารณาจากโครงสร้างและชนิดของปฏิกิริยาที่มีอยู่ โมเลกุล DNA สั้นๆ ที่มีรอยบากซึ่งยังคงสามารถเรียงซ้อนแบบโคแอกเซียลได้นั้นเคลื่อนที่เร็วกว่าโมเลกุล DNA ที่มีช่องว่างและไม่มีการเรียงซ้อนแบบโคแอกเซียล ซึ่งสามารถอธิบายได้ด้วยคุณสมบัติพอลิเมอร์ของ DNA ที่โมเลกุลที่มีลักษณะเป็นแท่งแข็งกว่าจะเคลื่อนที่ได้เร็วกว่าตามความลาดชันทางไฟฟ้าในเมทริกซ์เมื่อเทียบกับโมเลกุลที่มีความยืดหยุ่นมากกว่า[ 32 ]การพัฒนาเทคนิคใหม่ๆ เช่น แหนบแสงและความสามารถในการพับโครงสร้างนาโนของ DNA นำไปสู่การวัดมัด DNA และความสามารถในการเรียงซ้อนซึ่งกันและกัน จากนั้นจึงสามารถวิเคราะห์แรงที่จำเป็นในการดึงมัดเหล่านี้ออกจากกันโดยใช้แหนบแสงเพื่อวัดพลังงานการเรียงซ้อนของคู่เบส[ 33 ]การวัดเหล่านี้ส่วนใหญ่ดำเนินการภายใต้สภาวะที่ไม่สมดุล และมีการประมาณค่าต่างๆ เพื่อให้ได้ค่าที่แน่นอนของการเรียงซ้อนแบบโคแอกเซียลระหว่างเบส การศึกษาโมเลกุลเดี่ยวล่าสุดโดยใช้โครงสร้างนาโน DNA และกล้องจุลทรรศน์ความละเอียดสูง DNA-PAINT ทำให้สามารถวัดปฏิสัมพันธ์ระหว่างไดนิวคลีโอไทด์เหล่านี้ได้โดยใช้การวิเคราะห์จลนศาสตร์เชิงลึกของเวลาการจับของโมเลกุล DNA สั้นกับลำดับที่เสริมกันในกรณีที่มีหรือไม่มีปฏิสัมพันธ์การเรียงซ้อนของ DNA [ 31 ]
ลวดลายอื่นๆ
ปฏิสัมพันธ์ระหว่าง Tetraloop กับตัวรับ

ปฏิสัมพันธ์ระหว่าง Tetraloop และตัวรับจะรวมการจับคู่เบสและปฏิสัมพันธ์การเรียงซ้อนระหว่างนิวคลีโอไทด์ของลูปใน โมทีฟ Tetraloopและโมทีฟตัวรับที่อยู่ใน RNA duplex ทำให้เกิดการสัมผัสระดับตติยภูมิที่ช่วยให้โครงสร้างระดับตติยภูมิโดยรวมของ โมเลกุล RNA มีเสถียรภาพ Tetraloop ยังเป็นโครงสร้างที่เป็นไปได้ใน DNA duplex อีกด้วย[ 35 ]
ก้านห่วงสามารถแตกต่างกันอย่างมากในขนาดและลำดับ แต่ก้านห่วงที่มี นิว คลีโอไทด์ สี่ตัว นั้นพบได้ทั่วไปและมักจะอยู่ในหนึ่งในสามประเภทตามลำดับ[ 36 ] สามตระกูลนี้ได้แก่ ก้านห่วงCUYG, UNCG และ GNRA (ดูรูปทางด้านขวา) [ 37 ]ในแต่ละตระกูลก้านห่วงเหล่านี้ นิวคลีโอไทด์ตัวที่สองและตัวที่สามจะก่อให้เกิดการโค้งงอในสาย RNA และคู่เบสระหว่างนิวคลีโอไทด์ตัวแรกและตัวที่สี่จะทำให้โครงสร้างก้านห่วงมีความเสถียร โดยทั่วไปแล้ว ความเสถียรของก้านห่วงขึ้นอยู่กับองค์ประกอบของเบสภายในห่วงและองค์ประกอบของ "คู่เบสปิด" นี้[ 38 ]ตระกูลก้านห่วง GNRA เป็นตระกูลที่พบได้บ่อยที่สุดในปฏิสัมพันธ์ระหว่างก้านห่วงกับตัวรับ นอกจากนี้ ก้านห่วง UMAC ยังเป็นที่รู้จักกันว่าเป็นเวอร์ชันทางเลือกของก้านห่วง GNRA โดยทั้งสองมีโครงสร้างกระดูกสันหลังที่คล้ายกัน แม้จะมีความคล้ายคลึงกัน แต่ก็มีความแตกต่างกันในปฏิสัมพันธ์ระยะไกลที่เป็นไปได้ที่พวกมันสามารถทำได้[ 39 ]

"โมทีฟตัวรับเททราลูป" คือปฏิสัมพันธ์ตติยภูมิระยะไกล[ 40 ] ซึ่งประกอบด้วยพันธะไฮโดรเจนระหว่างเบสในเททราลูปกับลำดับสเต็มลูปในส่วนปลายของโครงสร้าง RNA ทุติยภูมิ[ 41 ] นอกเหนือจากพันธะไฮโดรเจนแล้ว ปฏิสัมพันธ์แบบเรียงซ้อนยังเป็นองค์ประกอบสำคัญของปฏิสัมพันธ์ตติยภูมิเหล่านี้ ตัวอย่างเช่น ในปฏิสัมพันธ์ GNRA-เททราลูป นิวคลีโอไทด์ตัวที่สองของเททราลูปจะเรียงซ้อนโดยตรงกับโมทีฟแพลตฟอร์ม A (ดูด้านบน) ภายในตัวรับ[ 26 ] ลำดับของเททราลูปและตัวรับมักจะแปรผันร่วมกันเพื่อให้สามารถสร้างการสัมผัสตติยภูมิประเภทเดียวกันได้กับไอโซฟอร์มต่างๆ ของเททราลูปและตัวรับที่เกี่ยวข้อง[ 42 ]
ตัวอย่างเช่น อินทรอนกลุ่ม I ที่ตัดต่อตัวเองนั้นอาศัยโมทีฟตัวรับแบบเททราลูปสำหรับโครงสร้างและการทำงาน[ 26 ] [ 41 ] โดยเฉพาะอย่างยิ่ง หมู่แอดินีนสามหมู่ของโมทีฟ GAAA แบบดั้งเดิมจะเรียงซ้อนกันบนเกลียวตัวรับและสร้างพันธะไฮโดรเจนที่ทำให้เสถียรหลายพันธะกับตัวรับ แอดินีนตัวแรกของลำดับ GAAA สร้างคู่เบสสามตัวกับเบส AU ของตัวรับ แอดินีนตัวที่สองมีความเสถียรโดยพันธะไฮโดรเจนกับยูริดีนตัว เดียวกัน รวมถึงผ่าน 2'-OH กับตัวรับและผ่านปฏิกิริยากับกัวนีนของเททราลูป GAAA แอดินีนตัวที่สามสร้างคู่เบสสามตัว
ลวดลายเอไมเนอร์


โครงสร้างโมทีฟ A-minor เป็น โครงสร้างโมที ฟระดับตติยภูมิ ของ RNA ที่พบได้ทั่วไป เกิด จากการแทรกตัวของนิวคลีโอไซด์ ที่ไม่มีคู่ เข้าไปในร่องเล็กของ RNA แบบคู่ ดังนั้นจึงเป็นตัวอย่างของโครงสร้างแบบสามตัวในร่องเล็กแม้ว่ากัวโนซีน ไซโตซีน และยูริดีนก็สามารถสร้างโครงสร้างแบบสามตัวในร่องเล็กได้เช่นกัน แต่การสร้างโครงสร้างในร่องเล็กโดยอะดีนีนนั้นพบได้บ่อยมาก ในกรณีของอะดีนีน ขอบ N1-C2-N3 ของเบสที่แทรกเข้าไปจะสร้างพันธะไฮโดรเจนกับ 2'-OH หนึ่งหรือทั้งสองข้างของ RNA แบบคู่ รวมถึงเบสของ RNA แบบคู่ด้วย (ดูรูป: โครงสร้างโมทีฟ A-minor) RNA แบบคู่ที่เป็นตัวรับมักจะเป็นเบสคู่ GC
รูปแบบ A-minor ได้รับการแยกออกเป็นสี่ประเภท[ 8 ] [ 9 ]ประเภท 0 ถึง III โดยพิจารณาจากตำแหน่งของเบสที่แทรกเข้าไปเมื่อเทียบกับ 2'-OH สองตัวของคู่เบส Watson-Crick ในรูปแบบ A-minor ประเภท I และ II นั้น N3 ของอะดีนีนจะแทรกเข้าไปลึกในร่องเล็กของดูเพล็กซ์ (ดูรูป: ปฏิสัมพันธ์ A-minor - ปฏิสัมพันธ์ประเภท II) และมีความเข้ากันได้ของรูปร่างที่ดีกับคู่เบส แตกต่างจากประเภท 0 และ III ปฏิสัมพันธ์ประเภท I และ II มีความจำเพาะต่ออะดีนีนเนื่องจากปฏิสัมพันธ์พันธะไฮโดรเจน ในปฏิสัมพันธ์ประเภท III ทั้ง O2' และ N3 ของเบสที่แทรกเข้าไปจะเชื่อมโยงกับร่องเล็กของดูเพล็กซ์น้อยกว่า รูปแบบประเภท 0 และ III นั้นอ่อนแอกว่าและไม่จำเพาะเจาะจงเนื่องจากเกิดจากการมีปฏิสัมพันธ์กับ 2'-OH เพียงตัวเดียว (ดูรูป: ปฏิสัมพันธ์ A-minor - ปฏิสัมพันธ์ประเภท 0 และประเภท III)
ลวดลาย A-minor เป็นหนึ่งในลวดลายโครงสร้าง RNA ที่พบได้บ่อยที่สุดในไรโบโซม ซึ่งมีส่วนช่วยในการจับ tRNA กับซับยูนิต 23S [ 44 ]โดยส่วนใหญ่มักจะทำให้ปฏิสัมพันธ์ของ RNA duplex มีเสถียรภาพในลูปและเกลียว เช่น ในแกนกลางของอินทรอนกลุ่ม II [ 6 ]
ตัวอย่างที่น่าสนใจของ A-minor คือบทบาทของมันใน การจดจำ แอนติโคดอน ไรโบโซมต้องแยกแยะระหว่างคู่โคดอน-แอนติโคดอนที่ถูกต้องและไม่ถูกต้อง โดยส่วนหนึ่งทำได้โดยการแทรกเบสอะดีนีนเข้าไปในร่องเล็ก คู่โคดอน-แอนติโคดอนที่ไม่ถูกต้องจะมีรูปทรงเกลียวที่บิดเบี้ยว ซึ่งจะป้องกันไม่ให้ปฏิกิริยาของ A-minor ทำให้การจับกันมีเสถียรภาพ และเพิ่มอัตราการแยกตัวของ tRNA ที่ไม่ถูกต้อง[ 45 ]
การวิเคราะห์โมทีฟ A-minor ในRNA ไรโบโซม 23Sได้เปิดเผยเครือข่ายลำดับชั้นของการพึ่งพาโครงสร้าง ซึ่งคาดว่าเกี่ยวข้องกับวิวัฒนาการของไรโบโซมและลำดับเหตุการณ์ที่นำไปสู่การพัฒนาหน่วยย่อยขนาดใหญ่ของแบคทีเรียสมัยใหม่[ 46 ]
มีรายงานว่าโมทีฟ A-minor และคลาสย่อยใหม่ของมัน ปฏิสัมพันธ์ WC/H A-minor ช่วยเสริมความแข็งแกร่งให้กับโครงสร้างตติยภูมิของ RNA อื่นๆ เช่น เกลียวสามชั้นร่องหลักที่ระบุในองค์ประกอบการทำให้เสถียรของ RNA [ 16 ] [ 15 ]
ซิปริโบส

ริโบสซิปเปอร์เป็น องค์ประกอบโครงสร้างตติยภูมิ ของ RNAซึ่งสาย RNA สองสายถูกยึดเข้าด้วยกันโดย ปฏิกิริยา พันธะไฮโดรเจนที่เกี่ยวข้องกับ 2'OH ของ น้ำตาล ริโบสบนสายที่แตกต่างกัน 2'OH สามารถทำหน้าที่เป็นทั้งผู้ให้และผู้รับพันธะไฮโดรเจน ซึ่งช่วยให้เกิดพันธะไฮโดรเจนแบบแยกสาขากับ 2'OH อีกตัวหนึ่งได้[ 47 ] [ 48 ]
มีการรายงานรูปแบบต่างๆ ของริโบสซิปเปอร์จำนวนมาก แต่รูปแบบทั่วไปเกี่ยวข้องกับพันธะไฮโดรเจนสี่พันธะระหว่างกลุ่ม 2'-OH ของน้ำตาลสองโมเลกุลที่อยู่ติดกัน ริโบสซิปเปอร์มักเกิดขึ้นในอาร์เรย์ที่ทำให้ปฏิสัมพันธ์ระหว่างสาย RNA ที่แยกจากกัน มีเสถียรภาพ [ 49 ] ริโบสซิปเปอร์มักพบเห็นเป็น ปฏิสัมพันธ์ แบบก้านและห่วงที่มีความจำเพาะของลำดับต่ำมาก อย่างไรก็ตาม ใน หน่วยย่อย ของไรโบโซม ขนาดเล็กและขนาดใหญ่ มีแนวโน้มที่จะเกิดริโบสซิปเปอร์ของลำดับ CC/AA ซึ่งก็คือไซโตซีน สองตัวบนสายแรกจับคู่กับอะ ดีนีนสอง ตัว บนสายที่สอง
บทบาทของไอออนโลหะ


RNAที่ทำหน้าที่มักเป็นโมเลกุลที่พับและเสถียร มีรูปร่างสามมิติ แทนที่จะเป็นสายเชิงเส้นที่ยืดหยุ่น[ 51 ]แคตไอออนมีความสำคัญต่อการรักษาเสถียรภาพทางอุณหพลศาสตร์ของโครงสร้างตติยภูมิของ RNA แคตไอออนโลหะที่จับกับ RNA อาจเป็นโมโนวาเลนต์ ไดวาเลนต์ หรือไตรวาเลนต์ โพแทสเซียม (K + ) เป็นไอออนโมโนวาเลนต์ทั่วไปที่จับกับ RNA ไอออนไดวาเลนต์ทั่วไปที่จับกับ RNA คือแมกนีเซียม (Mg2 + ) ไอออนอื่นๆ รวมถึงโซเดียม (Na + ) แคลเซียม (Ca2 + ) และแมงกานีส (Mn2 + ) พบว่าจับกับ RNA ในร่างกายและในหลอดทดลอง แคตไอออน อินทรีย์หลายวาเลนต์ เช่นสเปอร์มิดีนหรือสเปอร์มีนก็พบในเซลล์และมีส่วนสำคัญต่อการพับของ RNA ไอออนไตรวาเลนต์ เช่น โคบอลต์เฮกซามีน หรือไอออนแลนทานัม เช่นเทอร์เบียม (Tb3 + ) เป็นเครื่องมือทดลองที่มีประโยชน์สำหรับการศึกษาการจับของโลหะกับ RNA [ 52 ] [ 53 ]
ไอออนโลหะสามารถทำปฏิกิริยากับ RNA ได้หลายวิธี ไอออนสามารถจับกับโครงสร้างหลักของ RNA ได้อย่างไม่เป็นระเบียบ ช่วยป้องกันปฏิกิริยาทางไฟฟ้าสถิต ที่ไม่พึงประสงค์ การป้องกันประจุนี้มักเกิดขึ้นโดยไอออนโมโนวาเลนต์ ไอออนที่จับกับตำแหน่งเฉพาะจะช่วยทำให้โครงสร้างตติยภูมิของ RNA มีความเสถียรมากขึ้น ปฏิกิริยาที่จับกับตำแหน่งเฉพาะสามารถแบ่งย่อยออกเป็นสองประเภท ขึ้นอยู่กับว่าน้ำเป็นตัวกลางในการจับกับโลหะหรือไม่ ปฏิกิริยาแบบ "วงนอก" เกิดขึ้นโดยโมเลกุลของน้ำที่ล้อมรอบไอออนโลหะ ตัวอย่างเช่น แมกนีเซียมเฮกซาไฮเดรตทำปฏิกิริยาและทำให้โครงสร้างตติยภูมิของ RNA มีความเสถียรมากขึ้นผ่านปฏิกิริยากับกัวโนซีนในร่องหลัก ในทางกลับกัน ปฏิกิริยาแบบ "วงใน" เกิดขึ้นโดยตรงจากไอออนโลหะ RNA มักพับตัวในหลายขั้นตอน และขั้นตอนเหล่านี้สามารถทำให้เสถียรได้ด้วยแคตไอออนชนิดต่างๆ ในขั้นตอนแรกๆ RNA จะสร้างโครงสร้างทุติยภูมิที่มีความเสถียรผ่านการจับกับแคตไอออนโมโนวาเลนต์ แคตไอออนไดวาเลนต์ และอะมีนโพลีแอนไอออนิก เพื่อทำให้โครงสร้างหลักโพลีแอนไอออนิกเป็นกลาง ขั้นตอนต่อมาของกระบวนการนี้เกี่ยวข้องกับการก่อตัวของโครงสร้างตติยภูมิของ RNA ซึ่งจะมีความเสถียรเกือบทั้งหมดผ่านการจับตัวของไอออนประจุคู่ เช่น แมกนีเซียม โดยอาจมีการมีส่วนร่วมจากการจับตัวของโพแทสเซียมด้วย
Metal-binding sites are often localized in the deep and narrow major groove of the RNA duplex, coordinating to the Hoogsteen edges of purines. In particular, metal cations stabilize sites of backbone twisting where tight packing of phosphates results in a region of dense negative charge. There are several metal ion-binding motifs in RNA duplexes that have been identified in crystal structures. For instance, in the P4-P6 domain of the Tetrahymena thermophilagroup I intron, several ion-binding sites consist of tandem G-U wobble pairs and tandem G-A mismatches, in which divalent cations interact with the Hoogsteen edge of guanosine via O6 and N7.[54][55][56] Another ion-binding motif in the Tetrahymena group I intron is the A-A platform motif, in which consecutive adenosines in the same strand of RNA form a non-canonical pseudobase pair.[57] Unlike the tandem G-U motif, the A-A platform motif binds preferentially to monovalent cations. In many of these motifs, absence of the monovalent or divalent cations results in either greater flexibility or loss of tertiary structure.
Divalent metal ions, especially magnesium, have been found to be important for the structure of DNA junctions such as the Holliday junction intermediate in genetic recombination. The magnesium ion shields the negatively charged phosphate groups in the junction and allows them to be positioned closer together, allowing a stacked conformation rather than an unstacked conformation.[58] Magnesium is vital in stabilizing these kinds of junctions in artificially designed structures used in DNA nanotechnology, such as the double crossover motif.[59]
History
The earliest work in RNA structural biology coincided, more or less, with the work being done on DNA in the early 1950s. In their seminal 1953 paper, Watson and Crick suggested that van der Waals crowding by the 2`OH group of ribose would preclude RNA from adopting a double helical structure identical to the model they proposed - what we now know as B-form DNA.[60] This provoked questions about the three dimensional structure of RNA: could this molecule form some type of helical structure, and if so, how?
ในช่วงกลางทศวรรษ 1960 บทบาทของ tRNA ในการสังเคราะห์โปรตีนได้รับการศึกษาอย่างเข้มข้น ในปี 1965 Holley และคณะได้ทำการแยกและจัดลำดับโมเลกุล tRNA ตัวแรก โดยเสนอในเบื้องต้นว่ามันมีโครงสร้างแบบใบโคลเวอร์ โดยอาศัยความสามารถของบางส่วนของโมเลกุลในการสร้างโครงสร้างแบบก้านและห่วงเป็นหลัก[ 61 ]การแยก tRNA พิสูจน์แล้วว่าเป็นความสำเร็จครั้งสำคัญครั้งแรกในชีววิทยาโครงสร้างของ RNA ในปี 1971 Kim และคณะประสบความสำเร็จอีกครั้ง โดยสร้างผลึกของยีสต์ tRNA PHE ที่สามารถเลี้ยวเบนได้ถึงความละเอียด 2–3 อังสตรอม โดยใช้สเปอร์มีน ซึ่ง เป็นโพลีอะมีนที่เกิดขึ้นตามธรรมชาติซึ่งจับกับและทำให้ tRNA มีเสถียรภาพ[ 62 ]
หลังจากโครงสร้าง tRNA แรกๆ ออกมาได้ไม่นาน สาขาโครงสร้าง RNA ก็ไม่ได้ก้าวหน้าไปมากนัก ความสามารถในการศึกษาโครงสร้าง RNA ขึ้นอยู่กับศักยภาพในการแยกเป้าหมาย RNA ซึ่งพิสูจน์แล้วว่าเป็นข้อจำกัดของสาขานี้มาหลายปี ส่วนหนึ่งเป็นเพราะเป้าหมายอื่นๆ ที่รู้จักกัน เช่น ไรโบโซมนั้นยากต่อการแยกและตกผลึกมากกว่ามาก ดังนั้น ในช่วงประมาณยี่สิบปีหลังจากที่มีการตีพิมพ์โครงสร้าง tRNA PHE ครั้งแรก โครงสร้างของเป้าหมาย RNA อื่นๆ เพียงไม่กี่ตัวเท่านั้นที่ได้รับการแก้ไข โดยเกือบทั้งหมดอยู่ในตระกูล transfer RNA [ 63 ]
ข้อจำกัดที่น่าเสียดายนี้ในที่สุดก็จะได้รับการแก้ไขส่วนใหญ่เนื่องจากความก้าวหน้าครั้งสำคัญสองประการในการวิจัยกรดนิวคลีอิก ได้แก่ การระบุไรโบไซม์และความสามารถในการผลิตไรโบไซม์ผ่าน การถอดรหัส ในหลอดทดลองหลังจากการตีพิมพ์ของ Tom Cech ที่ระบุว่า อินทรอนกลุ่ม I ของ Tetrahymenaเป็นไรโบไซม์แบบเร่งปฏิกิริยา ด้วยตนเอง [ 64 ]และรายงานของ Sidney Altman เกี่ยวกับการเร่งปฏิกิริยาโดยไรโบนิวคลีเอส P RNA [ 65 ] RNA ที่เร่งปฏิกิริยาอื่นๆ อีกหลายชนิดได้รับการระบุในช่วงปลายทศวรรษ 1980 [ 66 ]รวมถึงไรโบไซม์หัวค้อน ในปี 1994 McKay และคณะได้ตีพิมพ์โครงสร้างของ 'คอมเพล็กซ์ไรโบไซม์-สารยับยั้ง RNA-DNA หัวค้อน' ที่ความละเอียด 2.6 อังสตรอม ซึ่งกิจกรรมเร่งปฏิกิริยาด้วยตนเองของไรโบไซม์ถูกขัดขวางโดยการจับกับสารตั้งต้น DNA [ 67 ] นอกเหนือจากความก้าวหน้าในการกำหนดโครงสร้างทั่วโลกผ่านทางผลึกศาสตร์แล้ว ช่วงต้นทศวรรษ 1990 ยังมีการนำ NMR มาใช้ซึ่งเป็นเทคนิคที่มีประสิทธิภาพในชีววิทยาโครงสร้าง RNA การวิจัยเช่นนี้ทำให้สามารถระบุลักษณะเฉพาะของการจับคู่เบสและปฏิสัมพันธ์การเรียงซ้อนของเบสได้อย่างแม่นยำยิ่งขึ้น ซึ่งทำให้โครงสร้างโดยรวมของโมเลกุล RNA ขนาดใหญ่มีความเสถียร
การฟื้นตัวของชีววิทยาโครงสร้าง RNA ในช่วงกลางทศวรรษ 1990 ทำให้เกิดการระเบิดอย่างแท้จริงในสาขาการวิจัยโครงสร้างกรดนิวคลีอิก นับตั้งแต่การตีพิมพ์โครงสร้างแฮมเมอร์เฮดและ P 4-6 ได้มีการสร้างผล งานสำคัญมากมายในสาขานี้ ตัวอย่างที่น่าสนใจที่สุดบางส่วน ได้แก่ โครงสร้างของ อินทรอน กลุ่ม Iและกลุ่ม II [ 6 ]และ ไร โบโซม[ 43 ]โครงสร้างสามแบบแรกสร้างขึ้นโดยใช้ การถอดรหัส ในหลอดทดลองและ NMR มีบทบาทในการตรวจสอบส่วนประกอบย่อยของโครงสร้างทั้งสี่ ซึ่งเป็นเครื่องพิสูจน์ถึงความจำเป็นของทั้งสองเทคนิคสำหรับการวิจัย RNA รางวัลโนเบลสาขาเคมีประจำปี 2009 มอบให้แก่Ada Yonath , Venkatraman RamakrishnanและThomas Steitz สำหรับผลงานโครงสร้าง ไรโบโซมของพวกเขาซึ่งแสดงให้เห็นถึงบทบาทที่โดดเด่นของชีววิทยาโครงสร้าง RNA ในชีววิทยาโมเลกุลสมัยใหม่
ดูเพิ่มเติม
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ โครงสร้างตติยภูมิของกรดนิวคลีอิก
โครงสร้างตติยภูมิของกรดนิวคลีอิก คือ รูปร่างสามมิติ ของพอลิเมอร์ กรดนิวคลีอิก [ 1 ] โมเลกุล RNA และ DNA มีความสามารถในการทำงานที่หลากหลาย ตั้งแต่ การจดจำโมเลกุล ไปจนถึง...
เกลียวคู่
โครงสร้างเกลียวคู่เป็นโครงสร้างตติยภูมิที่เด่นสำหรับดีเอ็นเอทางชีวภาพ และยังเป็นโครงสร้างที่เป็นไปได้สำหรับอาร์เอ็นเอ เชื่อกันว่ามีโครงสร้างดีเอ็นเอสามแบบที่พบในธรรมชาติ ได้แก่ A-DNA , B-DNA และ Z-DNA เชื่อกันว่ารูปแบบ "B" ที่อธิบายโดย James D.
ไตรเพล็กซ์ร่องเมเจอร์และไมเนอร์
โครงสร้างแบบไตร เพล็กซ์ในร่องเล็ก (minor groove triplex) เป็น โครงสร้างโมทีฟ ของ RNA ที่พบได้ทั่วไป เนื่องจากการโต้ตอบกับ ร่องเล็ก มักเกิดขึ้นโดยผ่านหมู่ 2'-OH ของ น้ำตาล ไรโบ ส โครงสร้างโมทีฟของ RNA นี้จึงดูแตกต่างจากโครงสร้างโมทีฟ ที่เทียบเท่าใน DNA...
ควอดรูเพล็กซ์
นอกจาก เกลียวคู่ และเกลียวสามชั้นที่กล่าวถึงข้างต้นแล้ว RNA และ DNA ยังสามารถสร้างเกลียวสี่ชั้นได้อีกด้วย โครงสร้างของควอดรูเพล็กซ์ของเบส RNA มีความหลากหลาย โมเลกุลกัวนีน สี่โมเลกุล ที่เรียงต่อกัน สามารถสร้างควอดรูเพล็กซ์ใน RNA ได้ด้วย พันธะไฮโดรเจนแบบ...