อ่าน 14 นาที
สติม2
โมเลกุลปฏิสัมพันธ์ของสโตรมัล 2 (STIM2)เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนSTIM2
สติม2
| สติม2 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| ตัวระบุ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| ชื่อเรียกอื่น | STIM2 , โมเลกุลปฏิสัมพันธ์ระหว่างสโตรมา 2 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| รหัสภายนอก | โอมิม : 610841 ; เอ็มจีไอ : 2151156 ; โฮโมโลยีน : 32490 ; GeneCards : STIM2 ; OMA : STIM2 - ออโธโลจี | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| วิกิดาต้า | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




โมเลกุลปฏิสัมพันธ์ของสโตรมัล 2 (STIM2)เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนSTIM2 [ 5 ] [ 6 ]
ยีนนี้เป็นสมาชิกของตระกูลโมเลกุลปฏิสัมพันธ์ระหว่างสโตรมา (STIM) ซึ่งประกอบด้วยสมาชิกเพียงสองตัวร่วมกับโฮโมล็อกSTIM1และน่าจะสืบเชื้อสายมาจากยีนบรรพบุรุษร่วมกัน ยีนเหล่านี้เข้ารหัสโปรตีนทรานส์เมมเบรนชนิดที่ 1ซึ่งตั้งอยู่ในซาร์โค/เอนโดพลาสมิกเรติคูลัม ( SR / ER ) ภายในเซลล์ การเริ่มต้นการแปลรหัสแบบทางเลือกจากไซต์เริ่มต้น AUG และไซต์เริ่มต้นที่ไม่ใช่ AUG (UUG) ส่งผลให้เกิดไอโซฟอร์ม STIM2 ที่แตกต่างกันสอง แบบ
สมาชิกทั้งสองของตระกูล STIM ถูกระบุในปี 2005 ว่าเป็น เซนเซอร์ แคลเซียม อิสระ (Ca 2+ ) ซึ่งมีส่วนร่วมในกลไกการเข้าสู่เซลล์ของ Ca 2+ที่เรียกว่า การเข้าสู่เซลล์โดยอาศัยแหล่งเก็บ ( store-operated Ca 2+ entry หรือ SOCE)กระบวนการต่างๆ ในเซลล์และเส้นทางการส่งสัญญาณจำนวนมากเริ่มต้นด้วยการปลดปล่อย Ca 2+ที่เก็บไว้ในออร์แกเนลล์ภายในเซลล์ซึ่งจำเป็นต้องมีการเติมเต็มอย่างต่อเนื่องSOCEถือเป็นกลไกการเติมเต็มแหล่งเก็บและเป็นกลไกสำคัญของการส่งสัญญาณ Ca 2+ในเซลล์ที่ไม่สามารถกระตุ้นด้วยไฟฟ้าได้ ในขณะที่ STIM1 กระตุ้นSOCEการวิจัยเกี่ยวกับหน้าที่ของ STIM2 ชี้ให้เห็นถึงบทบาทสำคัญในฐานะตัวควบคุมป้อนกลับที่ช่วยรักษา ระดับความเข้มข้นของ Ca 2+ ในไซ โตพลาสมิกและ S/ER ให้คงที่ [Ca 2+ ] STIM2 ตรวจจับการลดลงเล็กน้อยของปริมาณ Ca 2+ที่เก็บไว้ใน S/ER เปลี่ยนไปอยู่ในสถานะที่เปิดใช้งาน และมีปฏิสัมพันธ์กับ ช่อง Ca 2+ที่ควบคุมโดยแหล่งเก็บ(SOC)ที่อยู่บนเยื่อหุ้มเซลล์เช่น ช่อง OraiหรือTRPCทำให้เกิดSOCEขึ้น แม้ว่าบทบาทเชิงหน้าที่ของ STIM2 จะยังไม่เป็นที่แน่ชัดมาหลายปี แต่การศึกษาที่ดำเนินการในปี 2009-2010 ในแบบจำลองหนูชี้ให้เห็นว่า STIM2 มีส่วนร่วมในกระบวนการพัฒนาและการทำงานของเซลล์หลายชนิด รวมถึงไมโอบลาสต์ ของกล้ามเนื้อเรียบ เซลล์ของระบบภูมิคุ้มกันและเซลล์ประสาทและมีส่วนเกี่ยวข้องกับ การเกิด เนื้องอกการพัฒนาของโรคภูมิต้านตนเองและกลไกความเสียหายของเซลล์ประสาทหลังจากภาวะ ขาดเลือด ชั่วคราว
ยีน
ในปี พ.ศ. 2544 STIM2ได้รับการระบุว่าเป็นโฮโมล็อกของมนุษย์ตัวใหม่ของ ยีน STIM1ซึ่งเป็นสมาชิกตัวที่สองของกลุ่มยีนสองตัวในสัตว์มีกระดูกสันหลัง[ 5 ]ยีนSTIM2 ประกอบด้วย เอ็กซอน 12 ตัว และอินทรอน 11 ตัวซึ่งตั้งอยู่บนโครโมโซม 4p15.1 ของมนุษย์ และบนแขนยาวของโครโมโซม 5 ของหนู ใกล้กับเซนโทรเมียร์สมาชิกใน กลุ่มยีน STIMน่าจะวิวัฒนาการมาจากยีนเดียวในยูคาริโอต หลายเซลล์ระดับต่ำ ไปเป็นยีนที่เกี่ยวข้องสองตัวในสัตว์มีกระดูกสันหลัง เนื่องจากSTIM1และSTIM2 ของมนุษย์ รวมถึงStim ของDrosophila melanogaster ( D-Stim ) มีโครงสร้างจีโนมที่อนุรักษ์ไว้ โปรตีน D-STIM ที่มีกรดอะมิโน 570 ตัว แสดงความคล้ายคลึงกันเท่ากันทั้งกับ STIM1 (เหมือนกัน 33%; ลำดับกรด อะมิโนอนุรักษ์ไว้ 50%) และ STIM2 (เหมือนกัน 31%; ลำดับกรดอะมิโนอนุรักษ์ไว้ 46%) ยูคาริโอตเซลล์เดียวเช่นMonosiga brevicollis ซึ่งเป็น โคอาโนแฟลเจลเลตเซลล์เดียวได้รับการรายงานว่ามียีนที่คล้ายกับ STIM [ 7 ]อย่างไรก็ตาม ยังไม่มี การระบุยีนที่คล้ายกับ STIMในโปรคาริโอตและยังไม่มีการระบุโปรตีนที่คล้ายกับ STIM เพิ่มเติมในสัตว์มีกระดูกสันหลังจนถึงปัจจุบัน[ 5 ]
โครงสร้างโปรตีน
โปรตีน STIM2 เป็นโปรตีนทรานส์เมมเบรนชนิดที่ 1 ที่อยู่ใน S/ER โปรตีน STIM2 ของมนุษย์ประกอบด้วยกรดอะมิโน 833 โมเลกุล( 105-115 kDa) (รูปที่ 1) ซึ่งมากกว่า STIM1 ของมนุษย์ 148 โมเลกุล บริเวณ ปลายด้าน N ของทั้งสองโปรตีน มีความคล้ายคลึงกัน 66% ในส่วนของกรดอะมิโน 577 โมเลกุล (85% ของลำดับกรดอะมิโนของ STIM1) มีเพียง บริเวณ ปลายด้าน C เท่านั้น ที่แสดงความแตกต่างของลำดับอย่างมีนัยสำคัญ โครงสร้างโดเมนของทั้งสองไอโซฟอร์มมีการอนุรักษ์ไว้อย่างสูงในสัตว์มีกระดูกสันหลัง (รูปที่ 1) STIM2 ของหนูมีความเหมือนกับ STIM2 ของมนุษย์ 92% ในลำดับกรดอะมิโนตามการจัดเรียงแบบคู่ที่สร้างโดยBLAST โครงสร้าง โดเมนของ ทั้งสองโปรตีน ก็มีการอนุรักษ์ไว้อย่างสูงเช่นกัน (รูปที่ 1) STIM2 ของมนุษย์ได้รับการดัดแปลงหลังการแปลในร่างกายเช่น การเจริญเติบโตโดยการตัดเปปไทด์ส่งสัญญาณ S/ER ที่ปลาย N (14 กรดอะมิโน) การเติมหมู่ไกลโคซิลและการเติมหมู่ฟอสเฟต ในระดับต่างๆ แต่ตำแหน่งที่มีการเติมหมู่ฟอสเฟตยังไม่เป็นที่ทราบแน่ชัด (รูปที่ 1) [ 5 ]
สถาปัตยกรรมโดเมน
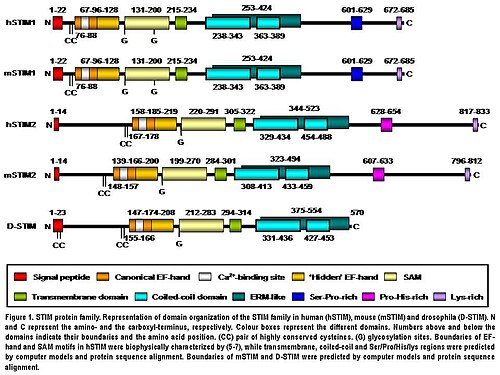
บริเวณปลาย N ของ STIM2 ตั้งอยู่ในลูเมน S/ER และประกอบด้วยโมทีฟการจับCa2 + แบบ EF-hand มาตรฐาน โมทีฟการจับ Ca2 +แบบ EF-hand ที่ "ซ่อนอยู่" ซึ่งเพิ่งค้นพบเมื่อเร็ว ๆ นี้ และโดเมน sterile α-motif (SAM) ซึ่งเป็นโมทีฟการโต้ตอบระหว่างโปรตีนที่รู้จักกันดี (รูปที่ 1) [ 8 ] [ 9 ] [ 10 ] ส่วนปลาย N ถูกแยกออกจากบริเวณปลาย C ด้วยโมทีฟทรานส์เมมเบรนแบบ ผ่านครั้งเดียวซึ่งมีการอนุรักษ์ไว้อย่างสูงในโปรตีน STIM ทั้งหมด บริเวณปลาย C ประกอบด้วยโครงสร้าง α-helixจำนวนมากสัดส่วนขนาดใหญ่ที่อยู่ใกล้กับโดเมนทรานส์เมมเบรนประกอบด้วยบริเวณที่คล้ายกับโดเมน ezrin/radixin/moesin ( ERM ) ซึ่งมี โดเมน coiled-coil สอง โดเมน[ 11 ]โดเมน coiled-coil ทำหน้าที่เป็นตัวกลางในการโต้ตอบระหว่างโปรตีน STIM ทำให้พวกมันสามารถจับกันและสร้าง homodimer และ heterodimer ได้ (รูปที่ 1) [ 12 ] [ 13 ] [ 14 ]สุดท้ายนี้ ทางด้านปลาย C-terminus โปรตีน STIM2 จะมีโมทีฟที่อุดมไปด้วยโพรลีน/ฮิสติดีนและหางที่อุดมไปด้วยไลซีนจำนวน 17 กรดอะมิโน (รูปที่ 1) [ 5 ]
ภูมิภาค EF-hand-SAM
เนื่องจากโดเมน EF-hand และ SAM (EF-SAM) มีความสำคัญต่อการทำงานของ STIM และการควบคุม SOCE จึงจะมีการอธิบายรายละเอียดต่อไป โดเมน EF-hand เป็นเซนเซอร์ Ca2 +ที่โปรตีน STIM ใช้ในการตรวจจับการเปลี่ยนแปลงความเข้มข้นของ Ca2 +ภายใน S/ER ไอโซฟอร์มของ STIM จะถูกกระตุ้นเมื่อ Ca2 +ที่จับกับโมทีฟ EF-hand ถูกปล่อยออกมาอันเป็นผลมาจากการลดลงของระดับ Ca2 +ภายในแหล่งเก็บ S/ER หลังจาก การลดลงของ ตัวรับIP3มีรายงานว่า STIM EF-hand กลายพันธุ์ที่ไม่สามารถจับกับ Ca2 + ได้ นั้นยังคงทำงานอย่างต่อเนื่องและกระตุ้น SOCE อย่างต่อเนื่องโดยไม่ขึ้นอยู่กับ [Ca2 + ] ของ S/ER ทั้ง ในหลอดทดลอง [ 15 ]และในร่างกาย [ 16 ] [ 17 ] [ 18 ] โดเมน SAM มีความสำคัญต่อการเกิดโอลิโกเมอไรเซชันของ STIM เนื่องจากตัวกลายพันธุ์ในโดเมนนี้ขาดความสามารถในการสร้างจุดเหนี่ยวนำได้[ 19 ]การทดลองการจับ Ca 2+ ในหลอดทดลองโดยใช้ชิ้นส่วน STIM1 EF–SAM ของมนุษย์ (ส่วนประกอบ 58–201) หรือ STIM2 EF–SAM (ส่วนประกอบ 149–292) แสดงให้เห็นว่าทั้งสองไอโซฟอร์มจับ Ca 2+ด้วยความสัมพันธ์ที่คล้ายคลึงกัน (STIM2 Kd~0.5 mM; STIM1 Kd~0.2–0.6 mM) [ 20 ] [ 21 ]ซึ่งอยู่ในช่วงค่าที่รายงานสำหรับ [Ca 2+ ] ของ S/ER [ 22 ] [ 23 ]อย่างไรก็ตาม STIM2 แตกต่างจาก STIM1 ตรงที่มันทำงานบางส่วนแล้วที่ [Ca 2+ ] พื้นฐานของ S/ER และจะทำงานเต็มที่เร็วกว่าในระหว่างการพร่องของแหล่งเก็บ S/ER แม้ว่าชิ้นส่วน STIM EF-SAM จะแสดงความสัมพันธ์กับ Ca 2+ เหมือนกัน แต่โปรตีน STIM2 แบบเต็มกลับแสดงความไวต่อ [Ca 2+ ] ที่ต่ำกว่า STIM1 ในเซลล์ที่ถ่ายยีน ในหลอดทดลอง [ 24 ] ความแตกต่างนี้บ่งชี้ว่าบริเวณโปรตีนอื่นๆ ยังมีส่วนร่วมในความไวต่อ [Ca 2+ ] หรือเกณฑ์การกระตุ้นที่แตกต่างกันของทั้งสองไอโซฟอร์ม โดเมน EF-hand ที่ "ซ่อนอยู่" ไม่จับกับ Ca 2+แต่มีความสำคัญต่อการเชื่อมโยงภายในโมเลกุล การพับ และความเสถียรของโดเมน EF-hand และ SAM เมื่อไม่นานมานี้ มีรายงานว่าการกลายพันธุ์ที่สำคัญทางโครงสร้างใน EF-hand แบบดั้งเดิม EF-hand ที่ "ซ่อนอยู่" หรือโดเมน SAM ทำให้ความไวต่อ Ca 2+ ลดลง เนื่องจากการทำให้บริเวณ EF-SAM ทั้งหมดไม่เสถียร[ 25 ]
บริเวณปลายซี
นอกจากปลาย N แล้ว บริเวณปลาย C ยังเป็นส่วนสำคัญของโปรตีน STIM ด้วย โดยแสดงความแตกต่างของลำดับอย่างมีนัยสำคัญระหว่างไอโซฟอร์มทั้งสอง และใน STIM1 บริเวณปลาย C มีความสำคัญต่อการโต้ตอบกับช่องSOC [ 26 ] STIM2 ของมนุษย์มี โมทีฟที่อุดมไปด้วย โพรลีนและฮิสติดีน (PHAPHPSHPRHPHHPQHTPHSLPSPDP) ในตำแหน่งที่คล้ายกับ บริเวณที่อุดมไป ด้วยซีรีนและโพรลีน (SPSAPPGGSPHLDSSRSHSPSSPDPDTPSP) ใน STIM1 ความแตกต่างอย่างมีนัยสำคัญในบริเวณเหล่านี้อาจบ่งชี้ถึงความแตกต่างในหน้าที่ของไอโซฟอร์ม STIM ต่างจาก STIM1 STIM2 มี สัญญาณการกักเก็บ ER แบบไดไลซีน (K(X)KXX) ที่ปลาย C สุดขั้วซึ่งกักเก็บโปรตีนไว้ในERในขณะที่ STIM1 สามารถเคลื่อนที่ไปยังพื้นผิวเซลล์ได้[ 27 ]สุดท้ายนี้ หางที่อุดมด้วย ไลซีน ที่คล้ายกัน จำนวน 14 และ 17 หน่วยใน STIM1 และ STIM2 ตามลำดับนั้นตั้งอยู่ที่ปลายสุดของบริเวณปลาย C เปปไทด์ เชิงเส้น จากบริเวณโพลีเบสิกปลาย C ของ STIM1 ของมนุษย์ (หน่วย 667-685) และ STIM2 (หน่วย 730-746) จับกับแคลโมดูลินด้วยความสัมพันธ์สูงหรือต่ำในกรณีที่มีหรือไม่มี Ca 2+ตามลำดับ[ 28 ]การศึกษาส่วนใหญ่เกี่ยวกับการโต้ตอบของบริเวณปลาย C ได้ดำเนินการกับไอโซฟอร์ม STIM1 การเติมแทปซิกาจิน ( สารยับยั้งปั๊ม SERCAที่กระตุ้น SOCE โดยการลดปริมาณ Ca 2+ ภายในเซลล์แบบพาสซีฟ) ลงในเซลล์ต่อมน้ำลายของมนุษย์ เช่นเดียวกับเซลล์ต่อม ใต้คางของหนูที่กระจายตัวจะเพิ่มการตกตะกอนร่วมของ TRPC1 และ Orai1 กับ STIM1 [ 29 ] จาก การแสดงออกร่วมกัน ในหลอดทดลองของ STIM1 กลายพันธุ์ของมนุษย์ที่แตกต่างกันซึ่งขาดบริเวณปลาย C ที่แตกต่างกันในเซลล์ HEK293 งานวิจัยล่าสุด 3 ชิ้นรายงานว่าโดเมน ERM ในปลาย C (กรดอะมิโน 251-535, รูปที่ 1) ซึ่งประกอบด้วย โดเมน coiled-coilทำหน้าที่เป็นตัวกลางในการจับกันของ STIM1 กับ TRPC(1, 2, 4 และ 5) และการเคลื่อนย้ายของ STIM1 ไปยังเยื่อหุ้มพลาสมา นอกจากนี้ บริเวณที่อุดมไปด้วยไลซีนที่มีประจุบวกยังมีความสำคัญต่อการควบคุม TRPC1 [ 14 ] [ 26 ] [ 30 ] Li และคณะได้ระบุบริเวณเหล่านี้ (กรดอะมิโน 425-672) เพิ่มเติมว่าเป็นไซต์ปฏิสัมพันธ์ที่เป็นไปได้ระหว่าง STIM1 และ Orai1 [ 14 ] การทดลอง coimmunoprecipitation ในหลอดทดลอง หลังจากการแสดงออกร่วมกันชั่วคราวของโปรตีน STIM2 และ Orai1 ในHEK293เซลล์เผยให้เห็นว่า STIM2 สามารถโต้ตอบทางกายภาพกับ Orai1 ได้เช่นกัน โดยอาจผ่านทางบริเวณปลาย C ของ STIM2 [ 31 ]
การแสดงออกและการกระจายตัวในเนื้อเยื่อ
mRNAของ STIM2 ถูกแสดงออกโดยเนื้อเยื่อของมนุษย์ส่วนใหญ่ โปรตีน STIM2 ถูกแสดงออกโดยเซลล์ไลน์ ของมนุษย์หลายเซลล์ ร่วมกับ STIM1 ซึ่งบ่งชี้ว่าไอโซฟอร์มของ STIM ถูกแสดงออกร่วมกันในเซลล์เดียวกัน อย่างน้อยก็ในเซลล์ไลน์ที่จัดตั้งขึ้น[ 5 ]โปรตีน STIM2 ถูกแสดงออกอย่างกว้างขวางในเนื้อเยื่อ โดยปกติจะมีระดับต่ำกว่า STIM1 ยกเว้นในสมองหรือตับ ซึ่ง STIM2 ดูเหมือนจะเป็นไอโซฟอร์มที่เด่นกว่า[ 5 ] [ 32 ]การถอดรหัสของ Stim2 ยังถูกควบคุมแบบไดนามิก ตัวอย่างเช่น มีการเพิ่มขึ้นเมื่อเซลล์ T ที่ยังไม่เจริญเต็มที่เกิดการแยกตัว เป็นลิมโฟไซต์Th 1หรือTh 2 [ 33 ]
การทำงาน
หน้าที่ของ STIM2 เป็นที่ถกเถียงกัน การศึกษาเบื้องต้นพบว่าการลดระดับ STIM1 ด้วย siRNA แต่ไม่ใช่ STIM2 ช่วยลด SOCE ในเซลล์สัตว์เลี้ยงลูกด้วยนมได้อย่างมาก[ 15 ] [ 24 ] [ 34 ] [ 35 ] [ 36 ] Liou et al . รายงานว่าการลดระดับ STIM2 ในเซลล์ HeLa ช่วยลด SOCE ลงเล็กน้อยเช่นกัน[ 15 ] Soboloff et al . แนะนำว่า STIM2 ยับยั้ง SOCE เมื่อแสดงออกเพียงอย่างเดียว[ 13 ]แต่เมื่อแสดงออกร่วมกับ Orai1 จะทำให้เกิด SOCE ที่เกิดขึ้นอย่างต่อเนื่องจำนวนมาก[ 37 ]ในทางตรงกันข้าม Brandman et al . แนะนำว่า STIM2 อาจทำหน้าที่เป็นตัวควบคุมที่ทำให้ระดับ Ca 2+ ในไซ โตพลาสมิกและ ER มีเสถียรภาพ [ 24 ] Parvez และคณะใช้ การแสดงออกร่วมกันชั่วคราว ในหลอดทดลองของ STIM2 ของมนุษย์และ ช่อง SOC ที่แตกต่างกัน ใน เซลล์ HEK293รายงานว่า STIM2 เป็นตัวกลางของ SOCE ผ่านโหมดที่ขึ้นอยู่กับแหล่งเก็บและไม่ขึ้นอยู่กับแหล่งเก็บสองโหมด[ 31 ]เมื่อพิจารณาร่วมกัน ผลลัพธ์เหล่านี้บ่งชี้ถึงปฏิสัมพันธ์ที่ซับซ้อนซึ่งถูกควบคุมอย่างละเอียดโดยอัตราส่วนเซลล์ STIM1: STIM2: Orai และระดับภายในของพวกมัน
การศึกษาที่ดำเนินการในปี 2009-2010 โดยใช้ แบบ จำลองในหลอดทดลอง ของมนุษย์ หรือ แบบจำลอง ในร่างกายของ หนู ยืนยันผลลัพธ์ของ Brandman et al . และชี้ให้เห็นว่า STIM2 มีส่วนร่วมในกระบวนการพัฒนาและการทำงานของเซลล์หลายประเภท รวมถึงไมโอบลาสต์ ของกล้ามเนื้อเรียบ เซลล์ของระบบภูมิคุ้มกัน และเซลล์ประสาท นอกจากนี้ยังเกี่ยวข้องกับการเกิดเนื้องอก การพัฒนาของโรคภูมิต้านตนเอง และกลไกของความเสียหายของเซลล์ประสาทหลังจากสภาวะขาดเลือดชั่วคราว ในสภาวะพัก เซลล์ HEK293 ที่เพาะเลี้ยงซึ่งมีการแสดงออกของ STIM2 มากเกินไปหรือเซลล์ประสาทคอร์เทกซ์ที่ขาด STIM2 จะมีระดับ Ca2 + ภายในเซลล์ที่เพิ่มขึ้นหรือลดลง ตามลำดับ[ 31 ] [ 32 ]ซึ่งสนับสนุนแนวคิดที่ว่า STIM2 มีความสำคัญต่อการควบคุมระดับ Ca2 + ภายในเซลล์ อย่างไรก็ตาม เซลล์มีการทำงานอย่างมากในร่างกายและระดับ Ca2 + ภายใน เซลล์จะผันผวนอย่างต่อเนื่อง การพัฒนาวิธีการใหม่เพื่อศึกษาบทบาทของ STIM2 ในระดับ Ca2 + ภายใน เซลล์ ในร่างกาย จึงเป็นสิ่งจำเป็น ในไมโอบลาสต์ของมนุษย์ที่เพาะเลี้ยง STIM2 มีส่วนร่วมในการแยกเซลล์ไปเป็นไมโอทิวบ์[ 38 ]ในระบบภูมิคุ้มกัน STIM2 มีส่วนร่วมในการกระตุ้นการทำงานของเซลล์ T ในการผลิต อินเตอร์ลิวคิน 2 (IL-2)และอินเตอร์เฟรอนแกมมา (IFN γ )ซึ่งอาจเกิดจากการทำให้NFATคงอยู่ในนิวเคลียสตลอดจนการแยกเซลล์ T ที่ยังไม่เจริญ เต็มที่ไปเป็น ลิมโฟไซต์ Th 17ซึ่งคาดว่ามีความสำคัญในระยะเริ่มต้นของโรคภูมิต้านตนเอง[ 33 ] [ 39 ]ในความเป็นจริง หนูที่ขาด STIM2 แสดงอาการเล็กน้อยในระยะเริ่มต้นของโรคภูมิต้านตนเอง[ 39 ]ในเนื้อเยื่อประสาท STIM2 มีบทบาทสำคัญในความเสียหายของเซลล์ประสาทที่เกิดจากภาวะขาดเลือดและการไม่มี STIM2 ในหนูที่น็อกเอาต์ช่วยลดความเสียหายของเซลล์ประสาทที่เกิดจากภาวะขาดเลือดหลังจากการหยุดไหลเวียนของเลือดในสมองชั่วคราว[ 32 ]ผลการป้องกันระบบประสาทของการขาด STIM2 หลังเกิดภาวะขาดเลือดบ่งชี้ว่าสารยับยั้งการทำงานของ STIM2 อาจมีคุณค่าในการรักษาในฐานะตัวแทนป้องกันระบบประสาทเพื่อรักษาอาการบาดเจ็บจากภาวะขาดเลือด และ โรคความเสื่อมของระบบประสาทอื่นๆ ที่เกี่ยวข้องกับการเปลี่ยนแปลง สมดุลของ Ca 2+นอกจากนี้ การศึกษาทางวิทยาศาสตร์เดียวกันยังชี้ให้เห็นถึงบทบาทสำคัญของ STIM2 ในหน่วยความจำเชิงพื้นที่ที่ขึ้นอยู่กับฮิปโปแคมปัสการ ส่ง สัญญาณไซแนปส์และความยืดหยุ่น[ 32 ]
สุดท้ายนี้ ได้มีการพิสูจน์แล้วว่า STIM2 มีหน้าที่ก่อให้เกิดมะเร็งร่วมกับ STIM1 ในglioblastoma multiformeซึ่งโปรตีนทั้งสองชนิดมีการแสดงออกเพิ่มขึ้นและ/หรือมีจำนวนสำเนาเพิ่มขึ้น[ 40 ] [ 41 ]นอกจากนี้ STIM2 ยังตั้งอยู่ในโครโมโซม 4p15.1 ซึ่งเป็นบริเวณที่เกี่ยวข้องกับมะเร็งชนิดรุกรานของปอด เต้านม คอ และศีรษะ[ 42 ] [ 43 ] [ 44 ]
ปฏิสัมพันธ์
ดังที่กล่าวไว้ก่อนหน้านี้ STIM2 ได้รับการแสดงให้เห็นว่ามีปฏิสัมพันธ์กับSTIM1 [ 12 ] [ 13 ] [ 14 ] ช่อง SOCเช่นOrai (ICRACM) หรือTRPC [ 31 ]แคลโมดูลิน (CaM) [ 28 ] [ 31 ]และฟอสโฟอิโนซิไทด์ของเยื่อหุ้มพลาสมา[ 45 ] การแสดงออกของ STIM2 ได้รับการแสดงให้เห็นว่าได้รับอิทธิพลหรือถูกควบคุมโดยพรีเซนิลิ นในไฟโบรบลาสต์ตัวอ่อนของหนูและลิมโฟไซต์ B ของมนุษย์[ 46 ]
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ สติม2
โมเลกุลปฏิสัมพันธ์ของสโตรมัล 2 (STIM2)เป็นโปรตีนที่ในมนุษย์ถูกเข้ารหัสโดยยีนSTIM2
ยีน
ในปี พ.ศ. 2544 STIM2 ได้รับการระบุว่าเป็นโฮโมล็อกของมนุษย์ตัวใหม่ของ ยีน STIM1 ซึ่งเป็นสมาชิกตัวที่สองของกลุ่มยีนสองตัวในสัตว์มีกระดูกสันหลัง [ 5 ] ยีน STIM2 ประกอบด้วย เอ็กซอน 12 ตัว และ อินทรอน 11 ตัว ซึ่งตั้งอยู่บนโครโมโซม 4p15.
โครงสร้างโปรตีน
โปรตีน STIM2 เป็นโปรตีนทรานส์เมมเบรนชนิดที่ 1 ที่อยู่ใน S/ER โปรตีน STIM2 ของมนุษย์ประกอบด้วยกรดอะมิโน 833 โมเลกุล ( 105-115 kDa) (รูปที่ 1) ซึ่งมากกว่า STIM1 ของมนุษย์ 148 โมเลกุล บริเวณ ปลายด้าน N ของทั้งสองโปรตีน มีความคล้ายคลึงกัน 66% ในส่วนของกรดอะมิโน...
สถาปัตยกรรมโดเมน
บริเวณปลาย N ของ STIM2 ตั้งอยู่ใน ลูเมน S/ER และประกอบด้วย โมทีฟ การจับCa2 + แบบ EF-hand มาตรฐาน โมทีฟการจับ Ca2 + แบบ EF-hand ที่ "ซ่อนอยู่" ซึ่งเพิ่งค้นพบเมื่อเร็ว ๆ นี้ และโดเมน sterile α-motif (SAM) ซึ่งเป็นโมทีฟการโต้ตอบระหว่างโปรตีนที่รู้จักกันดี...