อ่าน 13 นาที
เอทีพี ซินเทส
เอนไซม์ ATP synthase เป็น เอนไซม์ ที่เร่งปฏิกิริยาการสร้างโมเลกุลเก็บพลังงาน อะดีโนซีนไตรฟอสเฟต (ATP) โดยใช้ อะดีโนซีนไดฟอสเฟต (ADP) และ ฟอสเฟต อนินทรีย์ (Pi ) ATP synthase เป็น...
เอทีพี ซินเทส
| เอทีพี ซินเทส | |||||||||
|---|---|---|---|---|---|---|---|---|---|
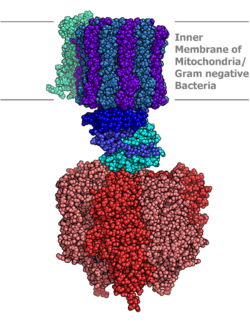 แบบจำลองโมเลกุลของเอนไซม์ ATP synthase ที่ได้จากการวิเคราะห์ด้วยรังสีเอกซ์ ส่วนสเตเตอร์ไม่ได้แสดงในภาพนี้ | |||||||||
| ตัวระบุ | |||||||||
| หมายเลข EC | 7.1.2.2 | ||||||||
| หมายเลข CAS | 9000-83-3 | ||||||||
| ฐานข้อมูล | |||||||||
| อินท์เอ็นซ์ | มุมมองของ IntEnz | ||||||||
| เบรนด้า | เบรนด้าเข้าร่วม | ||||||||
| เอ็กซ์แพซี่ | มุมมองของ NiceZyme | ||||||||
| เคกก์ | รายการ KEGG | ||||||||
| เมตาไซค์ | วิถีการเผาผลาญ | ||||||||
| ไพรแอม | ประวัติโดยย่อ | ||||||||
| โครงสร้างPDB | RCSB PDB PDBe PDBsum | ||||||||
| ออนโทโลยีของยีน | อามิโก้ / ควิกโก้ | ||||||||
| |||||||||
เอนไซม์ ATP synthaseเป็นเอนไซม์ที่เร่งปฏิกิริยาการสร้างโมเลกุลเก็บพลังงานอะดีโนซีนไตรฟอสเฟต (ATP) โดยใช้อะดีโนซีนไดฟอสเฟต (ADP) และฟอสเฟต อนินทรีย์ (Pi ) ATP synthase เป็นเครื่องจักรระดับโมเลกุลปฏิกิริยารวมที่เร่งโดย ATP synthase คือ:
- ADP + Pi + 2H + ออก ⇌ ATP + H2O + 2H + เข้า
เอนไซม์ ATP synthase อยู่บริเวณเยื่อหุ้มเซลล์และสร้างช่องเปิดที่โปรตอนสามารถเคลื่อนที่ผ่านจากบริเวณที่มีความเข้มข้นสูงไปยังบริเวณที่มีความเข้มข้นต่ำ ทำให้เกิดพลังงานสำหรับการสังเคราะห์ ATP ความแตกต่างของศักย์ไฟฟ้า เคมีนี้ เกิดขึ้นจากห่วงโซ่การขนส่งอิเล็กตรอนและช่วยให้เซลล์สามารถเก็บพลังงานในรูป ATP ไว้ใช้ในภายหลัง ใน เซลล์โปรคาริโอต เอนไซม์ ATP synthase อยู่บริเวณเยื่อหุ้มพลาสมาในขณะที่ใน เซลล์ยูคาริโอตจะอยู่บริเวณเยื่อหุ้มไมโทคอนเดรีย ชั้นใน สิ่งมีชีวิตที่สามารถสังเคราะห์แสงได้ยังมีเอนไซม์ ATP synthase อยู่บริเวณเยื่อไทลาคอยด์ซึ่งในพืชจะอยู่ในคลอโรพลาสต์และในไซยาโนแบคทีเรียจะอยู่ในไซโตพลาสซึม
ATP synthase ของยูคาริโอตเป็นF-ATPases (ซึ่งโดยปกติจะทำงานเป็น ATP synthase แทนที่จะเป็นATPaseในสภาพแวดล้อมของเซลล์) และทำงาน "ในทิศทางตรงกันข้าม" กับ ATPase (ATPases เร่งปฏิกิริยาการสลายตัวของATPเป็นADPและไอออนฟอสเฟต อิสระ ) บทความนี้กล่าวถึงประเภทนี้เป็นหลัก F-ATPase ประกอบด้วยหน่วยย่อยหลักสองหน่วยคือ F Oและ F 1ซึ่งมีกลไกมอเตอร์แบบหมุนที่ช่วยให้สามารถผลิต ATP ได้[ 1 ] [ 2 ]
การตั้งชื่อ
เศษส่วน F 1ได้รับชื่อมาจากคำว่า "เศษส่วน 1" และ F O (เขียนเป็นตัวห้อย "o" ไม่ใช่ "ศูนย์") ได้รับชื่อมาจากการเป็นเศษส่วนที่จับกับโอลิโกไมซินซึ่งเป็นยาปฏิชีวนะที่ได้จากธรรมชาติชนิดหนึ่งที่สามารถยับยั้งหน่วย F Oของ ATP synthase ได้ [ 3 ] [ 4 ]บริเวณการทำงานเหล่านี้ประกอบด้วยหน่วยย่อยโปรตีนที่แตกต่างกัน — โปรดดูตาราง เอนไซม์นี้ใช้ในการสังเคราะห์ ATP ผ่านการหายใจแบบใช้ออกซิเจน
โครงสร้างและหน้าที่

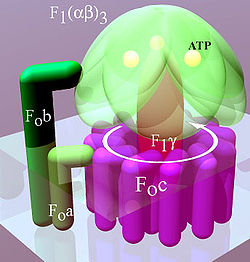

ATP synthase ซึ่งอยู่ภายในเยื่อไทลาคอยด์และเยื่อไมโทคอนเดรียชั้นในประกอบด้วยสองส่วนคือ F Oและ F 1 F Oทำให้เกิดการหมุนของ F 1และประกอบด้วยวงแหวน c และหน่วยย่อย a, b สองหน่วย และ F6 F 1ประกอบด้วยหน่วยย่อย α, β, γ และ δ F 1มีส่วนที่ละลายน้ำได้ซึ่งสามารถไฮโดรไลซ์ ATP ได้ ในทางกลับกัน F Oส่วนใหญ่มีบริเวณที่ไม่ชอบน้ำ F Oและ F 1สร้างทางเดินสำหรับการเคลื่อนที่ของโปรตอนข้ามเยื่อหุ้มเซลล์[ 7 ]
ภูมิภาคF 1
ส่วน F1 ของเอนไซม์ ATP synthase มี คุณสมบัติ ชอบน้ำและทำหน้าที่ไฮโดรไลซ์ ATP หน่วย F1 ยื่นเข้าไปในไซโตพลาสซึม ( เมทริกซ์และสโตรมาในไมโทคอนเดรียและคลอโรพลาสต์ตามลำดับ) หน่วยย่อย α และ β รวมกันเป็นเฮกซาเมอร์ที่มีตำแหน่งจับ 6 ตำแหน่ง โดยสามตำแหน่งนั้นไม่มีฤทธิ์เร่งปฏิกิริยาและจับกับ ADP
หน่วยย่อยอีกสามหน่วยทำหน้าที่เร่งปฏิกิริยาการสังเคราะห์ ATP หน่วยย่อย F 1 อื่นๆ γ, δ และ ε เป็นส่วนหนึ่งของกลไกมอเตอร์หมุน (โรเตอร์/แกน) หน่วยย่อย γ ช่วยให้ β เปลี่ยนแปลงโครงสร้าง (เช่น สถานะปิด สถานะเปิดครึ่งหนึ่ง และสถานะเปิด) ซึ่งช่วยให้ ATP สามารถจับและปล่อยออกมาได้เมื่อสังเคราะห์แล้ว อนุภาค F 1มีขนาดใหญ่และสามารถมองเห็นได้ในกล้องจุลทรรศน์อิเล็กตรอนแบบส่งผ่านโดยการย้อมสีแบบลบ[ 8 ]อนุภาคเหล่านี้มีเส้นผ่านศูนย์กลาง 9 นาโนเมตรที่กระจายอยู่ทั่วเยื่อหุ้มไมโทคอนเดรียชั้นใน
| หน่วยย่อย | ยีนมนุษย์ | บันทึก |
|---|---|---|
| อัลฟ่า | เอทีพี5เอฟ1เอ ,เอทีพีเอเอฟ2 | |
| เบต้า | เอทีพี5เอฟ1บี ,เอทีพีเอเอฟ1 | |
| แกมมา | เอทีพี5เอฟ1ซี | |
| เดลต้า | เอทีพี5เอฟ1ดี | "เดลต้า" ของไมโทคอนเดรีย คือ "เอปซิลอน" ของแบคทีเรีย/คลอโรพลาสต์ |
| เอปซิลอน | เอทีพี5เอฟ1อี | ลักษณะเฉพาะของไมโทคอนเดรีย |
| โอเอสซีพี | เอทีพี5พีโอ | เรียกว่า "เดลต้า" ในรูปแบบแบคทีเรียและคลอโรพลาสต์ |
ภูมิภาคF O

F Oเป็น โปรตีน ที่ไม่ละลาย น้ำ มีซับยูนิต 8 หน่วยและวงแหวนทรานส์เมมเบรน วงแหวนมีรูปร่างแบบเตตระเมอร์ ที่มีโปรตีน แบบเฮลิกซ์-ลูป-เฮลิกซ์ซึ่งจะมีการเปลี่ยนแปลงโครงสร้างเมื่อถูกโปรตอนและดีโปรตอนทำให้ซับยูนิตข้างเคียงหมุน ส่งผลให้ F O หมุน ซึ่งส่งผลต่อโครงสร้างของ F 1ด้วยส่งผลให้สถานะของซับยูนิตอัลฟาและเบตาเปลี่ยนไป บริเวณ F Oของ ATP synthase เป็นรูพรุนโปรตอนที่ฝังอยู่ในเยื่อหุ้มไมโทคอนเดรีย ประกอบด้วยซับยูนิตหลัก 3 หน่วย คือ a, b และ c ซับยูนิต c 6 หน่วยประกอบเป็นวงแหวนโรเตอร์ และซับยูนิต b ประกอบเป็นก้านที่เชื่อมต่อกับ F 1 OSCP ซึ่งป้องกันไม่ให้เฮกซาเมอร์ αβ หมุน ซับยูนิต a เชื่อมต่อ b กับวงแหวน c [ 11 ]มนุษย์มีซับยูนิตเพิ่มเติมอีก 6 หน่วย คือd , e , f , g , F6และ8 (หรือ A6L) ส่วนนี้ของเอนไซม์ตั้งอยู่ในเยื่อหุ้มชั้นในของไมโทคอนเดรีย และเชื่อมโยงการเคลื่อนย้ายโปรตอนเข้ากับการหมุนซึ่งก่อให้เกิดการสังเคราะห์ ATP ในบริเวณ F1
ในยูคาริโอต F O ของไมโทคอนเดรีย จะก่อตัวเป็นไดเมอร์ที่โค้งงอเยื่อหุ้มเซลล์ ไดเมอร์เหล่านี้จะจัดเรียงตัวเองเป็นแถวยาวที่ปลายคริสเตซึ่งอาจเป็นขั้นตอนแรกของการก่อตัวของคริสเต[ 12 ] แบบจำลองอะตอมสำหรับบริเวณ F Oไดเมอร์ของยีสต์ถูกกำหนดโดย cryo-EM ที่ความละเอียดโดยรวม 3.6 Å [ 13 ]
| หน่วยย่อย | ยีนมนุษย์ |
|---|---|
| เอ | เอ็มที-เอทีพี6 |
| ข | เอทีพี5พีบี |
| ค | ATP5MC1 , ATP5MC2 , ATP5MC3 |
แบบจำลองการผูกมัด


ในช่วงทศวรรษ 1960 ถึง 1970 พอล บอยเออร์ศาสตราจารย์จาก UCLAได้พัฒนาทฤษฎีกลไกการเปลี่ยนแปลงการจับตัว หรือกลไกพลิกกลับ (flip-flop) ซึ่งตั้งสมมติฐานว่าการสังเคราะห์ ATP ขึ้นอยู่กับการเปลี่ยนแปลงโครงสร้างของ ATP synthase ที่เกิดจากการหมุนของหน่วยย่อยแกมมา กลุ่มวิจัยของจอห์น อี. วอล์คเกอร์ซึ่งขณะนั้นอยู่ที่ห้องปฏิบัติการชีววิทยาโมเลกุล MRCในเคมบริดจ์ได้ตกผลึกโดเมนเร่งปฏิกิริยา F1 ของ ATP synthase โครงสร้างดังกล่าว ซึ่งในขณะนั้นเป็นโครงสร้างโปรตีนแบบไม่สมมาตรที่ใหญ่ที่สุดเท่าที่รู้จัก บ่งชี้ว่าแบบจำลองการเร่งปฏิกิริยาแบบหมุนของบอยเออร์นั้นถูกต้องโดยพื้นฐานแล้ว สำหรับการไขปริศนานี้ บอยเออร์และวอล์คเกอร์จึงได้รับรางวัลโนเบลสาขาเคมีร่วมกันใน ปี 1997
โครงสร้างผลึกของ F1 แสดง ให้เห็น ซับยูนิตอัลฟาและเบตาที่สลับกัน(อย่างละ 3 หน่วย) เรียงตัวคล้ายกลีบส้มรอบซับยูนิตแกมมาที่ไม่สมมาตรซึ่งหมุนอยู่ ตามแบบจำลองปัจจุบันของการสังเคราะห์ ATP (ที่รู้จักกันในชื่อแบบจำลองเร่งปฏิกิริยาแบบสลับ) ศักย์ไฟฟ้าข้ามเยื่อหุ้มเซลล์ที่สร้างขึ้นโดยไอออนโปรตอน (H+) ที่ส่งมาจากห่วงโซ่การขนส่งอิเล็กตรอน จะผลักดันไอออนโปรตอน (H+) จากช่องว่างระหว่างเยื่อหุ้มเซลล์ผ่านเยื่อหุ้มเซลล์โดยผ่านบริเวณ F1O ของ ATP synthase ส่วนหนึ่งของ F1O (วงแหวนของซับยูนิต c ) จะหมุนเมื่อโปรตอนผ่านเยื่อหุ้มเซลล์วงแหวน cยึดติดแน่นกับก้านกลางที่ไม่สมมาตร (ซึ่งประกอบด้วยซับยูนิตแกมมาเป็นหลัก) ทำให้มันหมุนภายในอัลฟา3เบตา3ของ F1 ส่งผลให้ตำแหน่งการจับนิวคลีโอไทด์เร่งปฏิกิริยา 3 ตำแหน่งเกิดการเปลี่ยนแปลงโครงสร้างหลายขั้นตอนซึ่งนำไปสู่การสังเคราะห์ ATP หน่วยย่อย หลัก F1 ถูกขัดขวางไม่ให้หมุนไปพร้อมกับแกนหมุนกลางโดยแกนรอบนอกที่เชื่อมต่ออัลฟา3เบตา3กับส่วนที่ไม่หมุนของ F2O โครงสร้างของเอนไซม์ ATP synthase ที่สมบูรณ์ในปัจจุบันเป็นที่ทราบกันในระดับความละเอียดต่ำจาก การศึกษา ด้วยกล้องจุลทรรศน์อิเล็กตรอนแบบแช่แข็ง (cryo-EM) แบบจำลอง cryo-EM ของ ATP synthase ชี้ให้เห็นว่าแกนรอบนอกเป็นโครงสร้างที่ยืดหยุ่นซึ่งพันรอบคอมเพล็กซ์ขณะที่เชื่อมต่อ F1 กับ F2O ภายใต้สภาวะที่เหมาะสม ปฏิกิริยาของเอนไซม์ยังสามารถเกิดขึ้นในทางกลับกันได้ โดยการไฮโดรไลซิสของATPจะขับเคลื่อนการปั๊มโปรตอนข้ามเยื่อหุ้มเซลล์
กลไกการเปลี่ยนแปลงการจับยึดเกี่ยวข้องกับการหมุนเวียนของไซต์ที่ใช้งานอยู่ของซับยูนิต β ระหว่างสามสถานะ[ 14 ]ในสถานะ "หลวม" ADP และฟอสเฟตจะเข้าสู่ไซต์ที่ใช้งานอยู่ ในแผนภาพด้านข้างนี้แสดงด้วยสีชมพู จากนั้นเอนไซม์จะมีการเปลี่ยนแปลงรูปร่างและบังคับให้โมเลกุลเหล่านี้มารวมกัน โดยไซต์ที่ใช้งานอยู่ในสถานะ "แน่น" ที่เกิดขึ้น (แสดงด้วยสีแดง) จะจับกับโมเลกุล ATP ที่ผลิตขึ้นใหม่ด้วยความสัมพันธ์ ที่สูงมาก สุดท้าย ไซต์ที่ใช้งานอยู่จะหมุนเวียนกลับไปสู่สถานะเปิด (สีส้ม) ปล่อย ATP และจับกับ ADP และฟอสเฟตเพิ่มเติม พร้อมสำหรับรอบการผลิต ATP ครั้งต่อไป[ 15 ]
บทบาททางสรีรวิทยา
เช่นเดียวกับเอนไซม์อื่นๆ กิจกรรมของ F1FO ATP synthase สามารถย้อนกลับได้ หาก มีATP ในปริมาณมากพอ จะทำให้เกิดการไล่ระดับโปรตอน ข้ามเยื่อหุ้มเซลล์ ซึ่งแบคทีเรีย ที่หมักอาหารจะใช้ประโยชน์จากสิ่งนี้ โดยแบคทีเรียเหล่านี้ไม่มีห่วงโซ่การขนส่งอิเล็กตรอน แต่จะไฮโดรไลซ์ ATP เพื่อสร้างการไล่ระดับโปรตอน ซึ่งใช้ในการขับเคลื่อนแฟลเจลลาและการขนส่งสารอาหารเข้าสู่เซลล์
ในแบคทีเรีย ที่หายใจ ภายใต้สภาวะทางสรีรวิทยา เอนไซม์ ATP synthase โดยทั่วไปจะทำงานในทิศทางตรงกันข้าม โดยสร้าง ATP โดยใช้แรงขับเคลื่อนโปรตอนที่สร้างขึ้นโดย ห่วงโซ่ การขนส่งอิเล็กตรอนเป็นแหล่งพลังงาน กระบวนการโดยรวมของการสร้างพลังงานในลักษณะนี้เรียกว่าการฟอสฟอริเลชันแบบออกซิเดชันกระบวนการเดียวกันนี้เกิดขึ้นในไมโทคอนเดรียซึ่ง ATP synthase ตั้งอยู่ในเยื่อหุ้มไมโทคอนเดรียชั้นใน และส่วน F1 ยื่นเข้าไปในเมทริกซ์ของไมโทคอนเดรียโดยการปั๊มไอออนโปรตอนเข้าไปในเมทริกซ์ ATP-synthase จะเปลี่ยน ADP ให้เป็น ATP
วิวัฒนาการ
เชื่อกันว่า วิวัฒนาการของ ATP synthase เป็นแบบโมดูลาร์ โดยที่หน่วยย่อยสองหน่วยที่ทำงานแยกจากกันได้มารวมกันและได้รับฟังก์ชันการทำงานใหม่[ 16 ] [ 17 ] การรวมตัวนี้ดูเหมือนจะเกิดขึ้นในช่วงต้นของประวัติศาสตร์วิวัฒนาการ เนื่องจากโครงสร้างและกิจกรรมของเอนไซม์ ATP synthase ที่เหมือนกันโดยพื้นฐานนั้นมีอยู่ในสิ่งมีชีวิตทุกอาณาจักร[ 16 ] F-ATP synthase แสดงความคล้ายคลึงกันในเชิงฟังก์ชันและกลไกกับV-ATPaseอย่าง มาก [ 18 ]อย่างไรก็ตาม ในขณะที่ F-ATP synthase สร้าง ATP โดยใช้การไล่ระดับโปรตอน V-ATPase สร้างการไล่ระดับโปรตอนโดยใช้ ATP ทำให้ค่า pH ต่ำถึง 1 [ 19 ]
บริเวณ F1 ยังแสดงความคล้ายคลึงอย่างมีนัยสำคัญกับดีเอ็นเอเฮลิเคส แบบเฮกซาเมอร์ (โดยเฉพาะปัจจัย Rho ) และบริเวณเอนไซม์ทั้งหมดแสดงความคล้ายคลึงกับH ใน ระดับหนึ่ง+-ขับเคลื่อนT3SSหรือคอมเพล็กซ์มอเตอร์แฟลเจลลา[ 18 ] [ 20 ] [ 21 ] เฮกซาเมอร์ α 3 β 3ของบริเวณ F 1แสดงความคล้ายคลึงทางโครงสร้างอย่างมีนัยสำคัญกับเฮลิเคส DNA แบบเฮกซาเมอร์ ทั้งสองก่อตัวเป็นวงแหวนที่มีสมมาตรการหมุน 3 เท่าโดยมีรูพรุนตรงกลาง ทั้งสองมีบทบาทที่ขึ้นอยู่กับการหมุนสัมพัทธ์ของโมเลกุลขนาดใหญ่ภายในรูพรุน เฮลิเคส DNA ใช้รูปร่างเกลียวของ DNA เพื่อขับเคลื่อนการเคลื่อนที่ไปตามโมเลกุล DNA และตรวจจับการขดตัว ในขณะที่เฮกซาเมอร์ α 3 β 3ใช้การเปลี่ยนแปลงโครงสร้างผ่านการหมุนของซับยูนิต γ เพื่อขับเคลื่อนปฏิกิริยาเอนไซม์[ 22 ]
เอช+มอเตอร์ของอนุภาค F Oแสดงให้เห็นถึงความคล้ายคลึงกันในเชิงฟังก์ชันอย่างมากกับH+มอเตอร์ที่ขับเคลื่อนแฟลเจลลา[ 18 ]ทั้งสองมีวงแหวนของโปรตีนอัลฟาเฮลิกซ์ขนาดเล็กจำนวนมากที่หมุนสัมพันธ์กับโปรตีนที่อยู่กับที่ใกล้เคียง โดยใช้H+ความชันศักยภาพเป็นแหล่งพลังงาน อย่างไรก็ตาม การเชื่อมโยงนี้ค่อนข้างอ่อนแอ เนื่องจากโครงสร้างโดยรวมของมอเตอร์แฟลเจลลัมมีความซับซ้อนมากกว่าอนุภาค F O มาก และวงแหวนที่มีโปรตีนหมุนประมาณ 30 ตัวนั้นมีขนาดใหญ่กว่าโปรตีนเกลียว 10, 11 หรือ 14 ตัวในคอมเพล็กซ์ F O มาก อย่างไรก็ตาม ข้อมูลโครงสร้างล่าสุดแสดงให้เห็นว่าวงแหวนและก้านมีโครงสร้างคล้ายกับ อนุภาคF 1 [ 21 ]
ทฤษฎีวิวัฒนาการแบบโมดูลาร์สำหรับการกำเนิดของ ATP synthase ชี้ให้เห็นว่ามีหน่วยย่อยสองหน่วยที่มีหน้าที่อิสระต่อกัน ได้แก่ DNA helicase ที่มีกิจกรรม ATPase และH+มอเตอร์สามารถจับกันได้ และการหมุนของมอเตอร์จะขับเคลื่อนกิจกรรม ATPase ของเฮลิเคสในทิศทางตรงกันข้าม[ 16 ] [ 22 ]จากนั้นคอมเพล็กซ์นี้ก็พัฒนาให้มีประสิทธิภาพมากขึ้นและในที่สุดก็พัฒนาเป็น ATP synthase ที่ซับซ้อนในปัจจุบัน หรืออีกทางหนึ่ง DNA helicase/ H+คอมเพล็กซ์มอเตอร์อาจมีH+กิจกรรมของปั๊มร่วมกับกิจกรรม ATPase ของเฮลิเคสที่ขับเคลื่อนH+มอเตอร์กลับทิศทาง[ 16 ]สิ่งนี้อาจวิวัฒนาการเพื่อดำเนินการปฏิกิริยาย้อนกลับและทำหน้าที่เป็น ATP synthase [ 17 ] [ 23 ] [ 24 ]
สารยับยั้ง
มีการค้นพบสารยับยั้ง ATP synthase ทั้งจากธรรมชาติและสังเคราะห์หลายชนิด[ 25 ]สารเหล่านี้ถูกนำมาใช้เพื่อตรวจสอบโครงสร้างและกลไกของ ATP synthase บางชนิดอาจมีประโยชน์ในการรักษา มีสารยับยั้ง ATP synthase หลายประเภท ได้แก่ สารยับยั้งเปปไทด์ สารเคมีจากพืชกลุ่ม โพลีฟี นอล โพลีคี ไทด์ สารประกอบออร์กา โนทิน อนุพันธ์โพลีเอนิก α-ไพโรน สารยับยั้งประจุบวก สารอะนาล็อกของสารตั้งต้น สารปรับเปลี่ยนกรดอะมิโน และสารเคมีอื่นๆ[ 25 ]สารยับยั้ง ATP synthase ที่ใช้กันทั่วไปบางชนิด ได้แก่โอลิโกไมซิน และDCCD
ในสิ่งมีชีวิตต่าง ๆ
แบคทีเรีย
E. coli ATP synthase เป็น ATP synthase รูปแบบที่ง่ายที่สุดที่รู้จัก โดยมีหน่วยย่อยที่แตกต่างกัน 8 ชนิด [ 11 ]
F-ATPases ของแบคทีเรียบางครั้งสามารถทำงานย้อนกลับได้ ทำให้กลายเป็น ATPase [ 26 ]แบคทีเรียบางชนิดไม่มี F-ATPase แต่ใช้ ATPase ชนิด A/V ที่ทำงานแบบสองทิศทาง[ 9 ]
ยีสต์
ยีสต์ ATP synthase เป็นหนึ่งใน ATP synthase ของยูคาริโอตที่ได้รับการศึกษามากที่สุด และมีการระบุซับยูนิต F 1 จำนวน 5 ซับยูนิต F O จำนวน 8 ซับยูนิต และโปรตีนที่เกี่ยวข้องอีก 7 ซับยูนิต [ 7 ]โปรตีนเหล่านี้ส่วนใหญ่มีโฮโมล็อกในยูคาริโอตอื่นๆ[ 27 ] [ 28 ] [ 29 ] [ 30 ]
ปลูก
ในพืช เอนไซม์ ATP synthase ยังพบได้ในคลอโรพลาสต์ (CF 1 F O -ATP synthase) เอนไซม์นี้ถูกรวมเข้ากับ เยื่อ ไทลาคอยด์ส่วนCF 1 จะยึดติดอยู่ใน สโตรมาซึ่งเป็นที่เกิดปฏิกิริยามืดของการสังเคราะห์แสง (เรียกอีกอย่างว่าปฏิกิริยาที่ไม่ขึ้นกับแสงหรือวัฏจักรแคลวิน ) และการสังเคราะห์ ATP โครงสร้างโดยรวมและกลไกการเร่งปฏิกิริยาของ ATP synthase ในคลอโรพลาสต์นั้นเกือบจะเหมือนกับเอนไซม์ในแบคทีเรีย อย่างไรก็ตาม ในคลอโรพลาสต์แรงขับเคลื่อนโปรตอนไม่ได้เกิดจากห่วงโซ่การขนส่งอิเล็กตรอนของการหายใจ แต่เกิดจากโปรตีนสังเคราะห์แสงขั้นต้นซินเทสมีส่วนแทรก 40 กรดอะมิโนในหน่วยย่อยแกมมาเพื่อยับยั้งกิจกรรมที่สิ้นเปลืองเมื่ออยู่ในที่มืด[ 31 ]
สัตว์เลี้ยงลูกด้วยนม
เอนไซม์ ATP synthase ที่แยกได้จากไมโทคอนเดรียหัวใจวัว ( Bos taurus ) ถือเป็น ATP synthase ที่ได้รับการศึกษาลักษณะเฉพาะมากที่สุดในแง่ของชีวเคมีและโครงสร้าง หัวใจวัวถูกใช้เป็นแหล่งของเอนไซม์เนื่องจากมีไมโทคอนเดรียในกล้ามเนื้อ หัวใจในปริมาณมาก ยีนของ เอนไซม์เหล่านี้มีความคล้ายคลึงกับ ATP synthase ของมนุษย์[ 32 ] [ 33 ] [ 34 ]
ยีนของมนุษย์ที่เข้ารหัสส่วนประกอบของเอนไซม์ ATP synthase:
- เอทีพี5เอฟ1เอ
- เอทีพี5เอฟ1บี
- ATP5F1C , ATP5F1D , ATP5F1E , ATP5F1 , ATP5MC1 , ATP5MC2 , ATP5MC3 , ATP5PD , ATP5ME , ATP5PF , ATP5MF , ATP5MG , ATP5PO
- MT-ATP6 , MT-ATP8
ยูคาริโอตอื่นๆ
ยูคาริ โอตที่อยู่ในสายพันธุ์ที่แตกต่างกันบางสายพันธุ์มีโครงสร้างพิเศษของ ATP synthase ATP synthase ของยูเกลโนซัวก่อตัวเป็นไดเมอร์ที่มีหัว F 1 รูปทรงบูมเมอแรง เช่นเดียวกับ ATP synthase ไมโทคอนเดรียอื่นๆ แต่ซับคอมเพล็กซ์ F Oมีซับยูนิตเฉพาะจำนวนมาก มันใช้คาร์ดิโอลิปิน นอกจากนี้ IF 1 ที่ยับยั้ง ยังจับตัวในลักษณะที่แตกต่างกัน ซึ่งเป็นลักษณะที่พบได้ในไทรพาโนโซมาติดา[ 35 ]
อะพิคอมเพล็กซานเช่นพลาสโมเดียมใช้โครงสร้าง ATP ซินเทสแบบเฮกซาเมอร์ิกซึ่งมีอยู่เป็นไตรเมอร์ของไดเมอร์ ATP ซินเทส โครงสร้างเฮกซาเมอร์ิกเหล่านี้จัดเรียงตัวเป็นพีระมิดห้าเหลี่ยมซึ่งทำให้เกิดรูปร่างของเยื่อหุ้มคริสตาที่มีลักษณะเป็นกระเปาะที่เป็นเอกลักษณ์ โครงสร้างนี้ได้รับการสนับสนุนโดยทั้งซับยูนิตเฉพาะของอะพิคอมเพล็กซานและคาร์ดิโอลิปิน[ 36 ]
อาร์เคีย
โดยทั่วไปอาร์เคียไม่มี F-ATPase แต่พวกมันสังเคราะห์ ATP โดยใช้ A-ATPase/synthase ซึ่งเป็นเครื่องจักรหมุนที่มีโครงสร้างคล้ายกับ V-ATPase แต่ส่วนใหญ่ทำหน้าที่เป็น ATP synthase [ 26 ]เช่นเดียวกับ F-ATPase ของแบคทีเรีย เชื่อกันว่ามันยังทำหน้าที่เป็น ATPase ด้วย[ 9 ]
LUCA และรุ่นก่อนหน้า
การเชื่อมโยงยีน F-ATPase และลำดับยีนได้รับการอนุรักษ์อย่างกว้างขวางในสายพันธุ์โปรคาริโอตโบราณ ซึ่งหมายความว่าระบบนี้มีอยู่แล้วตั้งแต่ก่อนบรรพบุรุษร่วมสากลสุดท้าย LUCA [ 37 ]
ดูเพิ่มเติม
- โปรตีน ATP10จำเป็นสำหรับการประกอบส่วน F Oของคอมเพล็กซ์ ATPase ในไมโทคอนเดรีย
- คลอโรพลาสต์
- ห่วงโซ่การถ่ายโอนอิเล็กตรอน
- ฟลาโวโปรตีน
- ไมโตคอนเดรีย
- การฟอสโฟรีเลชันแบบออกซิเดทีฟ
- พี-เอทีพีเอส
- ปั๊มโปรตอน
- การเคลื่อนที่แบบหมุนในสิ่งมีชีวิต
- ทรานส์เมมเบรน เอทีพีเอส
- วี-เอทีพีเอส
อ่านเพิ่มเติม
- Nick Lane: คำถามสำคัญ: พลังงาน วิวัฒนาการ และต้นกำเนิดของสิ่งมีชีวิตที่ซับซ้อน , Ww Norton, 2015-07-20, ISBN 978-0393088816(ลิงก์ไปยังรูปที่ 10 ซึ่งแสดงแบบจำลองของเอนไซม์ ATP synthase)
ลิงก์ภายนอก
- Boris A. Feniouk: "เอนไซม์ ATP synthase — เครื่องจักรโมเลกุลอันน่าทึ่ง"
- วิดีโอบรรยายเรื่อง ATP synthase ที่มีภาพประกอบอย่างดีถูกเก็บไว้ในWayback Machine เมื่อวันที่ 2 ธันวาคม 2008 โดย Antony Crofts จากมหาวิทยาลัยอิลลินอยส์ เออร์บานา-แชมเปญ
- โปรตอนและโซเดียมทรานสโลเคติง ATPase ชนิด F, V และ A ในฐานข้อมูล OPM
- รางวัลโนเบลสาขาเคมีประจำปี 1997มอบให้แก่ พอล ดี. โบเยอร์ และ จอห์น อี. วอล์กเกอร์ สำหรับกลไกทางเอนไซม์ของการสังเคราะห์ ATP และมอบให้แก่ เยนส์ ซี. สโกว์ สำหรับการค้นพบเอนไซม์ขนส่งไอออนNa++, เค+-เอทีพีเอส
- เว็บไซต์ผลิตสื่อมัลติมีเดียของมหาวิทยาลัยฮาร์วาร์ด — วิดีโอ – แอนิเมชั่นการสังเคราะห์ ATP
- เดวิด กู๊ดเซลล์: "เอนไซม์ ATP Synthase - โมเลกุลประจำเดือน" เก็บถาวรเมื่อ 5 กันยายน 2015 ที่Wayback Machine
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เอทีพี ซินเทส
เอนไซม์ ATP synthase เป็น เอนไซม์ ที่เร่งปฏิกิริยาการสร้างโมเลกุลเก็บพลังงาน อะดีโนซีนไตรฟอสเฟต (ATP) โดยใช้ อะดีโนซีนไดฟอสเฟต (ADP) และ ฟอสเฟต อนินทรีย์ (Pi ) ATP synthase เป็น...
การตั้งชื่อ
เศษส่วน F 1 ได้รับชื่อมาจากคำว่า "เศษส่วน 1" และ F O (เขียนเป็นตัวห้อย "o" ไม่ใช่ "ศูนย์") ได้รับชื่อมาจากการเป็นเศษส่วนที่จับกับ โอลิโกไมซิน ซึ่งเป็นยาปฏิชีวนะที่ได้จากธรรมชาติชนิดหนึ่งที่สามารถยับยั้งหน่วย F O ของ ATP synthase ได้ [ 3 ] [ 4 ]...
โครงสร้างและหน้าที่
ATP synthase ซึ่งอยู่ภายใน เยื่อไทลาคอยด์ และ เยื่อไมโทคอนเดรียชั้นใน ประกอบด้วยสองส่วนคือ F O และ F 1 F O ทำให้เกิดการหมุนของ F 1 และประกอบด้วยวงแหวน c และหน่วยย่อย a, b สองหน่วย และ F6 F 1 ประกอบด้วยหน่วยย่อย α, β, γ และ δ F 1...
ภูมิภาคF 1
ส่วน F1 ของเอนไซม์ ATP synthase มี คุณสมบัติ ชอบน้ำ และทำหน้าที่ไฮโดรไลซ์ ATP หน่วย F1 ยื่น เข้าไปในไซโตพลาสซึม ( เมทริกซ์ และ สโตรมา ใน ไมโทคอนเดรียและคลอโรพลาสต์ตามลำดับ) หน่วยย่อย α และ β รวมกันเป็นเฮกซาเมอร์ที่มีตำแหน่งจับ 6 ตำแหน่ง...