อ่าน 59 นาที
คลอโรพลาสต์
คลอ โรพลาสต์ ( / ˈ k l ɔːr ə ˌ p l æ s t , - p l ɑː s t / KLOR -ə-plast, -plahst ) [ 1 ] [ 2 ] เป็น ออร์แกเนลล์ ชนิดหนึ่งที่เรียกว่า พลาสติด ซึ่งทำหน้าที่ สังเคราะห์แสง...
คลอโรพลาสต์


คลอโรพลาสต์ ( / ˈ k l ɔːr ə ˌ p l æ s t , - p l ɑː s t / KLOR -ə-plast, -plahst ) [ 1 ] [ 2 ] เป็น ออร์แกเนลล์ชนิดหนึ่งที่เรียกว่าพลาสติดซึ่งทำหน้าที่สังเคราะห์แสงส่วนใหญ่ใน เซลล์ พืชและสาหร่ายคลอโรพลาสต์มีความเข้มข้นของ รงควัตถุ คลอโรฟิลล์ สูง ซึ่งดักจับพลังงานจากแสงแดดเปลี่ยนเป็นพลังงานเคมีและสลายน้ำเพื่อปล่อยออกซิเจนพลังงานเคมีที่เกิดขึ้นจะถูกนำไปใช้ในการสร้างน้ำตาลและโมเลกุลอินทรีย์อื่นๆ จากคาร์บอนไดออกไซด์ในวัฏจักรแคลวินคลอโรพลาสต์ยังทำหน้าที่อื่นๆ อีกหลายอย่าง รวมถึงการสังเคราะห์กรดไขมันการ สังเคราะห์ กรดอะมิโนและการตอบสนองทางภูมิคุ้มกันในพืช จำนวนคลอโรพลาสต์ต่อเซลล์แตกต่างกันไป ตั้งแต่หนึ่งอันในสาหร่ายเซลล์เดียวบางชนิด ไปจนถึง 100 อันในพืช เช่นอาราบิโดปซิสและข้าวสาลี
คลอโรพลาสต์มีความเคลื่อนไหวสูงมาก - พวกมันหมุนเวียนและเคลื่อนที่ไปมาภายในเซลล์ พฤติกรรมของพวกมันได้รับอิทธิพลอย่างมากจากปัจจัยด้านสิ่งแวดล้อม เช่น สีและความเข้มของแสง เซลล์พืชไม่สามารถสร้างคลอโรพลาสต์ขึ้นใหม่ได้ และต้องได้รับการถ่ายทอดไปยังเซลล์ลูกแต่ละเซลล์ในระหว่างการแบ่งเซลล์ ซึ่งเชื่อกันว่าได้รับการถ่ายทอดมาจากบรรพบุรุษของพวกมัน คือไซยาโนแบคทีเรีย สังเคราะห์แสงโบราณ ที่ถูกกลืนกิน โดย เซลล์ยูคาริโอตยุคแรก[ 3 ]
เนื่องจากมีต้นกำเนิดจากการอยู่ร่วมกันแบบพึ่งพาอาศัยกันภายในเซลล์ คลอโร พลาสต์จึงมี ดีเอ็นเอของตัวเองแยกต่างหากจากนิวเคลียสของเซลล์ เช่นเดียวกับไม โทคอนเดรียยกเว้นเพียงกรณีเดียว ( อะมีบาPaulinella chromatophora ) คลอโรพลาสต์ทั้งหมดสามารถสืบย้อนกลับไปถึง เหตุการณ์การอยู่ร่วมกันแบบพึ่งพาอาศัยกันภายในเซลล์เพียงครั้งเดียวได้อย่างไรก็ตาม คลอโรพลาสต์สามารถพบได้ในสิ่งมีชีวิตที่หลากหลายอย่างยิ่งซึ่งไม่ได้มีความสัมพันธ์โดยตรงต่อกัน—ซึ่งเป็นผลมาจากเหตุการณ์การอยู่ร่วมกันแบบพึ่งพาอาศัยกันภายในเซลล์ หลายครั้ง ทั้งในระดับ ทุติยภูมิและตติย ภูมิ
การค้นพบและนิรุกติศาสตร์
คำอธิบายที่ชัดเจนครั้งแรกของคลอโรพลาสต์ ( Chlorophyllkörnen , "เมล็ดคลอโรฟิลล์") ได้รับการกล่าวถึงโดยHugo von Mohlในปี 1837 ว่าเป็นส่วนประกอบที่แยกจากกันภายในเซลล์พืชสีเขียว[ 4 ]ในปี 1883 Andreas Franz Wilhelm Schimperได้ตั้งชื่อส่วนประกอบเหล่านี้ว่า "คลอโรพลาสติด" ( Chloroplastida ) [ 5 ]ในปี 1884 Eduard Strasburgerได้นำคำว่า "คลอโรพลาสต์" ( Chloroplasten ) มาใช้ [ 6 ] [ 7 ] [ 8 ]
คำว่าคลอโรพลาสต์มาจากคำภาษากรีกchloros (χλωρός) ซึ่งหมายถึงสีเขียว และplastes (πλάστης) ซึ่งหมายถึง "ผู้สร้าง" [ 9 ]
กำเนิดคลอโรพลาสต์แบบเอนโดซิมไบโอซิส
คลอโรพลาสต์เป็นออร์แกเนลล์ชนิดหนึ่งในเซลล์ยูคาริโอตที่สังเคราะห์แสงได้ พวกมันวิวัฒนาการมาจากไซยาโนแบคทีเรียผ่านกระบวนการที่เรียกว่าออร์แกเนลล์เจเนซิส [ 10 ] ไซ ยาโนแบคทีเรียเป็น ไฟลัมที่หลากหลายของแบคทีเรียแกรมลบ ที่สามารถสังเคราะห์แสงแบบใช้ออกซิเจน ได้ เช่นเดียวกับคลอโรพลาสต์ พวกมันมีไทลาคอยด์ [ 11 ] เยื่อไทลาคอยด์ประกอบด้วยรงควัตถุสังเคราะห์แสงรวมถึงคลอโรฟิลล์เอ[ 12 ] [ 13 ]ต้นกำเนิดของคลอโรพลาสต์นี้ได้รับการเสนอครั้งแรกโดยนักชีววิทยาชาวรัสเซียKonstantin Mereschkowskiในปี 1905 [ 14 ]หลังจากที่Andreas Franz Wilhelm Schimperสังเกตในปี 1883 ว่าคลอโรพลาสต์มีลักษณะคล้ายกับไซยาโนแบคทีเรียอย่าง มาก [ 5 ]คลอโรพลาสต์พบได้เฉพาะในพืชสาหร่าย[ 15 ]และบางชนิดของ อะมีบ อยด์Paulinellaเท่านั้น[ 16 ]
เชื่อกันว่า ไมโตคอนเดรียมีต้นกำเนิดมาจาก เหตุการณ์ เอนโดซิมไบโอซิส ที่คล้ายคลึงกัน ซึ่งโปรคาริโอตแบบใช้ออกซิเจน ถูกกลืนเข้าไป[ 17 ]
เอนโดซิมไบโอซิสขั้นต้น
![เอนโดซิมไบโอซิสขั้นต้น ยูคาริโอตที่มีไมโตคอนเดรียกลืนกินไซยาโนแบคทีเรียในเหตุการณ์เอนโดซิมไบโอซิสขั้นต้นแบบอนุกรม ทำให้เกิดสายพันธุ์ของเซลล์ที่มีออร์แกเนลล์ทั้งสองชนิด[17]](https://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Chloroplast_endosymbiosis_simple.svg/500px-Chloroplast_endosymbiosis_simple.svg.png)
เมื่อประมาณสอง พันล้านปีก่อน[ 18 ] [ 19 ] [ 20 ]ไซยาโนแบคทีเรียที่ดำรงชีวิตอิสระได้เข้าไปใน เซลล์ ยูคาริโอต ยุคแรก ไม่ว่าจะเป็นอาหาร หรือปรสิตภายใน[ 17 ]แต่สามารถหลบหนีออกจากแวคิวโอลฟาโกไซติกที่มันบรรจุอยู่และคงอยู่ภายในเซลล์ได้[ 12 ]เหตุการณ์นี้เรียกว่าเอนโดซิมไบโอซิสหรือ "เซลล์ที่อาศัยอยู่ภายในเซลล์อื่นโดยต่างฝ่ายต่างได้ประโยชน์ร่วมกัน" เซลล์ภายนอกมักเรียกว่าโฮสต์ในขณะที่เซลล์ภายในเรียกว่าเอนโดซิมไบโอนต์ [ 17 ] ไซยาโนแบคทีเรียที่ถูกกลืนกินเข้าไปนั้นให้ประโยชน์แก่โฮสต์โดยการให้น้ำตาลจากการสังเคราะห์แสง[ 17 ]เมื่อเวลาผ่านไป ไซยาโนแบคทีเรียจะถูกดูดซึม และยีนจำนวนมากของมันสูญหายไปหรือถูกถ่ายโอนไปยังนิวเคลียสของโฮสต์[ 21 ]จากนั้นโปรตีนไซยาโนแบคทีเรียบางส่วนจะถูกสังเคราะห์โดยเซลล์เจ้าบ้านและนำกลับเข้าไปในคลอโรพลาสต์ (เดิมคือไซยาโนแบคทีเรีย) ทำให้เจ้าบ้านสามารถควบคุมคลอโรพลาสต์ได้[ 21 ] [ 22 ]
คลอโรพลาสต์ที่สามารถสืบย้อนกลับไปถึงบรรพบุรุษของไซยาโนแบคทีเรียโดยตรง (เช่น โดยไม่มีเหตุการณ์เอนโดซิมไบโอติกเกิดขึ้นภายหลัง) เรียกว่าพลาสติดปฐมภูมิ (" พลาสติด " ในบริบทนี้มีความหมายเกือบเหมือนกับคลอโรพลาสต์[ 17 ] ) [ 23 ]คลอโรพลาสต์ที่สามารถสืบย้อนกลับไปถึงเอนโดซิมไบโอติกยูคาริโอตสังเคราะห์แสงอื่น ๆ เรียกว่าพลาสติดทุติยภูมิหรือพลาสติดตติยภูมิ (จะกล่าวถึงต่อไป)
เป็นที่ถกเถียงกันมานานแล้วว่าคลอโรพลาสต์ปฐมภูมิเกิดขึ้นจากเหตุการณ์เอนโดซิมไบโอติกเพียงครั้งเดียวหรือจากการกลืนกินอย่างอิสระหลายครั้งในสายพันธุ์ยูคาริโอตต่างๆ ปัจจุบันเป็นที่ยอมรับกันโดยทั่วไปว่า ยกเว้นเพียงกรณีเดียว (Paulinella chromatophora ที่เป็นอะมีบา ) คลอโรพลาสต์เกิดขึ้นจากเหตุการณ์เอนโดซิมไบโอติกเพียงครั้งเดียวเมื่อประมาณสองพัน ล้านปีก่อน และคลอโรพลาสต์เหล่านี้ทั้งหมดมีบรรพบุรุษ ร่วมกัน [ 19 ]มีการเสนอว่าญาติที่ใกล้ชิดที่สุดของไซยาโนแบคทีเรียบรรพบุรุษที่ถูกกลืนกินคือGloeomargarita lithophora [ 24 ] [ 25 ] [ 26 ] นอกจากนี้ เมื่อประมาณ 90–140 ล้านปีก่อน กระบวนการนี้เกิดขึ้นอีกครั้งในPaulinellaที่เป็นอะมีบา โดยมีไซยาโนแบคทีเรียในสกุลProchlorococcus คลอโรพลา สต์ที่วิวัฒนาการอย่างอิสระนี้มักเรียกว่าโครมาโทฟอร์แทนที่จะเป็นคลอโรพลาสต์[ 27 ] [หมายเหตุ 1 ]
เชื่อกันว่าคลอโรพลาสต์เกิดขึ้นหลังจากไมโทคอนเดรียเนื่องจากยูคาริโอต ทั้งหมด มีไมโทคอนเดรีย แต่ไม่ใช่ทั้งหมดที่มีคลอโรพลาสต์[ 17 ] [ 28 ]นี่เรียกว่าเอนโดซิมไบโอซิสแบบอนุกรมซึ่งยูคาริโอตในยุคแรกกลืนกิน บรรพบุรุษของ ไมโทคอนเดรียจากนั้นลูกหลานของมันก็กลืนกินบรรพบุรุษของคลอโรพลาสต์ ทำให้เกิดเซลล์ที่มีทั้งคลอโรพลาสต์และไมโทคอนเดรีย[ 17 ]
ภาวะพึ่งพาอาศัยกันภายในเซลล์ระดับทุติยภูมิและตติยภูมิ

สิ่งมีชีวิตอื่นๆ อีกมากมายได้รับคลอโรพลาสต์จากสายพันธุ์คลอโรพลาสต์หลักผ่านเอนโดซิมไบโอซิสรอง—โดยการกลืนสาหร่ายสีแดงหรือสีเขียวที่มีคลอโรพลาสต์หลัก คลอโรพลาสต์เหล่านี้เรียกว่า พลา สติดรอง[ 23 ]
ผลจากการเกิดเอนโดซิมไบโอซิสครั้งที่สอง คลอโรพลาสต์ทุติยภูมิจะมีเยื่อหุ้มเพิ่มเติมนอกเหนือจากสองชั้นดั้งเดิมในคลอโรพลาสต์ปฐมภูมิ[ 29 ]ในพลาสติดทุติยภูมิ โดยทั่วไปจะมีเพียงคลอโรพลาสต์ และบางครั้งเยื่อหุ้มเซลล์และนิวเคลียส เท่านั้น ที่ยังคงอยู่ ทำให้เกิดคลอโรพลาสต์ที่มีเยื่อหุ้มสามหรือสี่ชั้น[ 30 ] — เยื่อหุ้มไซยาโนแบคทีเรียสองชั้น บางครั้งเยื่อหุ้มเซลล์ของสาหร่ายที่ถูกกิน และแวคิวโอลฟาโกโซมจากเยื่อหุ้มเซลล์ของโฮสต์[ 29 ]
ยีนในนิวเคลียสของยูคาริโอตที่ถูกฟาโกไซโทซิสมักจะถูกถ่ายโอนไปยังนิวเคลียสของโฮสต์รอง[ 29 ]คริปโตโมนาสและคลอราแรคนิโอไฟต์จะเก็บนิวเคลียสของยูคาริโอตที่ถูกฟาโกไซโทซิสไว้ ซึ่งเป็นวัตถุที่เรียกว่านิวคลีโอเมอร์ฟ[ 29 ] ซึ่งอยู่ระหว่างเยื่อหุ้มชั้นที่สองและชั้นที่สามของคลอโรพลาสต์[ 12 ] [ 22 ]
คลอโรพลาสต์ทุติยภูมิทั้งหมดมาจากสาหร่ายสีเขียวและ สีแดง ไม่มีการสังเกตคลอโรพลาสต์ทุติยภูมิจากกลอโคไฟต์อาจเป็นเพราะกลอโคไฟต์ค่อนข้างหายากในธรรมชาติ ทำให้มีโอกาสน้อยที่จะถูกยูคาริโอตอื่นนำไปใช้[ 29 ]
สิ่งมีชีวิตอื่นๆ รวมถึงไดโนแฟลเจลเลตKarlodiniumและKareniaได้รับคลอโรพลาสต์โดยการกลืนกินสิ่งมีชีวิตที่มีพลาสติดรอง สิ่งเหล่านี้เรียกว่า พลา สติดตติยภูมิ[ 23 ]

ก.ขณะนี้เป็นที่ยืนยันแล้วว่า Chromalveolataเป็นกลุ่มพาราไฟเลติกต่อ Rhizaria [ 32 ]
สายพันธุ์คลอโรพลาสต์หลัก
คลอโรพลาสต์หลักทั้งหมดเป็นของสาย พันธุ์คลอโรพลาสต์หนึ่งในสี่สายพันธุ์ ได้แก่สายพันธุ์คลอโรพลาสต์กลอโคไฟต์ สายพันธุ์คลอโรพลาสต์โรโดไฟต์ ("สีแดง") สายพันธุ์คลอโรพลา สติดา ("สีเขียว") และสายพันธุ์ อะมีบอยด์ Paulinella chromatophora [ 33 ] สายพันธุ์กลอโคไฟต์ โรโดไฟต์ และคลอโรพลาสติดา ล้วนสืบเชื้อสายมาจากเหตุการณ์เอนโดซิ มไบโอซิสบรรพบุรุษเดียวกัน และทั้งหมดอยู่ในกลุ่มArchaeplastida [ 29 ]
คลอโรพลาสต์ของกลอโคไฟต์

กลุ่มคลอโรพลาสต์ของกลอโคไฟต์เป็นกลุ่มที่เล็กที่สุดในบรรดาสายพันธุ์คลอโรพลาสต์หลักทั้งสามสายพันธุ์ เนื่องจากมีกลอโคไฟต์เพียง 25 ชนิดที่ได้รับการอธิบายไว้[ 34 ]กลอโคไฟต์แยกตัวออกมาก่อนที่สายพันธุ์คลอโรพลาสต์สีแดงและสีเขียวจะแยกตัวออกมา[ 35 ]ด้วยเหตุนี้ บางครั้งจึงถือว่ากลอโคไฟต์เป็นตัวกลางระหว่างไซยาโนแบคทีเรียและคลอโรพลาสต์สีแดงและสีเขียว[ 36 ]การแยกตัวออกมาในช่วงต้นนี้ได้รับการสนับสนุนจากทั้ง การศึกษา ทางวิวัฒนาการและลักษณะทางกายภาพที่มีอยู่ในคลอโรพลาสต์ของกลอโคไฟต์และไซยาโนแบคทีเรีย แต่ไม่พบในคลอโรพลาสต์สีแดงและสีเขียว ประการแรก คลอโรพลาสต์ของกลอโคไฟต์มี ผนังเซลล์แบบ เพปติโดไกลแคน ซึ่งเป็นผนังเซลล์ชนิดหนึ่งที่พบเฉพาะในแบคทีเรีย (รวมถึงไซยาโนแบคทีเรีย) [หมายเหตุ 2 ]ประการที่สอง คลอโรพลาสต์ของกลอโคไฟต์ประกอบด้วยไทลาคอยด์แบบวงกลมที่ไม่เรียงซ้อน กัน ซึ่งล้อม รอบคาร์ บอกซีโซม ซึ่งเป็นโครงสร้าง ทรงยี่สิบ หน้าที่ มีเอนไซม์RuBisCOที่รับผิดชอบในการตรึงคาร์บอนประการที่สาม แป้งที่สร้างโดยคลอโรพลาสต์จะถูกรวบรวมไว้ภายนอกคลอโรพลาสต์[ 37 ]นอกจากนี้ เช่นเดียวกับไซยาโนแบคทีเรีย ทั้งไทลาคอยด์ของกลอโคไฟต์และโรโดไฟต์ต่างก็มีโครงสร้างรวบรวมแสงที่เรียกว่าไฟโคบิลิโซม
โรโดไฟตา (คลอโรพลาสต์สีแดง)
- ความหลากหลายของสาหร่ายสีแดง คลอโรพลาสต์ของสาหร่ายสีแดงมีลักษณะเด่นคือมี รงควัตถุ ไฟโคบิลินซึ่งมักทำให้สาหร่ายมีสีแดง




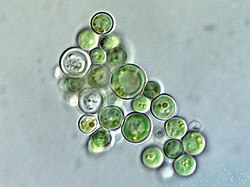





กลุ่ม โรโดไฟต์ หรือสาหร่ายสีแดงเป็นกลุ่มสายพันธุ์ขนาดใหญ่และหลากหลาย[ 29 ]คลอโรพลาสต์ของโรโดไฟต์เรียกอีกอย่างว่าโรโดพลาสต์ [ 23 ]ซึ่งแปลตรงตัวว่า "คลอโรพลาสต์สีแดง" [ 38 ] โรโดพลาสต์มีเยื่อหุ้มสอง ชั้นที่มีช่องว่างระหว่างเยื่อหุ้มและรงควัตถุไฟโคบิลิลินที่จัดเรียงเป็นไฟโค บิลิโซม บนเยื่อไทลาคอยด์ ป้องกันไม่ให้ไทลาคอยด์ซ้อนกัน[ 12 ]บางชนิดมี ไพรี นอยด์[ 23 ]โรโดพลาสต์มีคลอโรฟิลล์เอและไฟโคบิลิลิน[ 32 ]เป็นรงควัตถุสังเคราะห์แสง ไฟโคเอริทริน ซึ่งเป็นไฟโคบิลิลิน มีส่วนทำให้สาหร่ายสีแดงหลายชนิด มี สีแดง ที่โดดเด่น [ 39 ]อย่างไรก็ตาม เนื่องจากพวกมันยังมีคลอโรฟิลล์เอ สีเขียวอมฟ้า และรงควัตถุอื่นๆ อีกด้วย หลายชนิดจึงมีสีแดงถึงม่วงจากการผสมผสานกัน[ 23 ]รงควัตถุไฟโคเอริทรินสีแดงเป็นการปรับตัวเพื่อช่วยให้สาหร่ายสีแดงจับแสงแดดได้มากขึ้นในน้ำลึก[ 23 ]ดังนั้น สาหร่ายสีแดงบางชนิดที่อาศัยอยู่ในน้ำตื้นจึงมีไฟโคเอริทรินในโรโดพลาสต์น้อยลง และอาจมีสีเขียวมากขึ้น[ 39 ]โรโดพลาสต์สังเคราะห์แป้งชนิดหนึ่งที่เรียกว่าแป้งฟลอริเดียน[ 23 ]ซึ่งรวมตัวกันเป็นเม็ดเล็กๆ นอกโรโดพลาสต์ ในไซโตพลาสซึมของสาหร่ายสีแดง[ 12 ]
คลอโรพลาสติดา (คลอโรพลาสต์สีเขียว)
- ความหลากหลายของสาหร่ายสีเขียว คลอโรพลาสต์ของสาหร่ายสีเขียวมีลักษณะเฉพาะคือมีรงควัตถุคลอโรฟิลล์เอและคลอโรฟิลล์บีซึ่งทำให้สาหร่ายมีสีเขียว


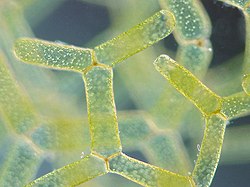







กลุ่มคลอโรพลาสติดาเป็นสายพันธุ์ขนาดใหญ่ที่มีความหลากหลายสูงอีกสายพันธุ์หนึ่ง ซึ่งรวมถึงทั้งสาหร่ายสีเขียวและพืชบก [ 40 ] กลุ่มนี้ยังเรียกว่าViridiplantaeซึ่งประกอบด้วยกลุ่มหลักสองกลุ่มได้แก่ ChlorophytaและStreptophyta
คลอโรพลาสต์สีเขียวส่วนใหญ่มี สี เขียวแม้ว่าบางส่วนจะไม่มีสีเขียวเนื่องจากรงควัตถุเสริมที่บดบังสีเขียวจากคลอโรฟิลล์ เช่น ในเซลล์พักตัวของHaematococcus pluvialisคลอโรพลาสต์สีเขียวแตกต่างจากคลอโรพลาสต์ของกลอโคไฟต์และสาหร่ายสีแดงตรงที่พวกมันสูญเสีย ไฟโคบิ ลิโซมไปและมีคลอโรฟิลล์บี[ 12 ] พวกมันยังสูญเสีย ผนัง เพปติโดไกลแคนระหว่างเยื่อหุ้มสองชั้น ทำให้มีช่องว่างระหว่างเยื่อหุ้ม[ 12 ]พืชบางชนิดยังคงยีน บางส่วนที่จำเป็นสำหรับการ สังเคราะห์เพปติโดไกลแคน แต่ได้นำยีนเหล่านั้นมาใช้ในการแบ่งคลอโรพลาสต์แทน[ 41 ]สายพันธุ์คลอโรพลาสติดายังคงเก็บแป้ง ไว้ ภายในคลอโรพลาสต์[ 12 ] [ 32 ] [ 40 ]ในพืชและสาหร่ายบางชนิด ไทลาคอยด์ของคลอโรพลาสต์จะเรียงตัวเป็นกองกรานา คลอโรพลาสต์ของสาหร่ายสีเขียวบางชนิด รวมถึงคลอโรพลาสต์ของฮอร์นเวิร์ตมีโครงสร้างที่เรียกว่าไพรีนอยด์[ 12 ]ซึ่งรวมRuBisCOและ CO 2 ไว้ ในคลอโรพลาสต์ ทำหน้าที่คล้ายกับคาร์บอกซีโซมของ กลอโคไฟต์ [ 42 ] [ 43 ]
มีสายพันธุ์ของสาหร่ายสีเขียวปรสิตที่ไม่สังเคราะห์แสงบางสายพันธุ์ที่สูญเสียคลอโรพลาสต์ไปทั้งหมด เช่นPrototheca [ 32 ] หรือไม่มีคลอโรพลาสต์แต่ยังคงมีจีโนมคลอโรพลาสต์แยกต่างหาก เช่นในHelicosporidium [ 44 ]ความคล้ายคลึงกันทางสัณฐานวิทยาและสรีรวิทยา รวมถึงวิวัฒนาการทางสายพันธุ์ยืนยันว่าสายพันธุ์เหล่านี้เคยมีคลอโรพลาสต์ในบรรพบุรุษ แต่ได้สูญเสียไปแล้ว[ 44 ] [ 45 ]
พอลิเนลลา โครมาโทโฟรา

อะมีบอยด์สังเคราะห์แสงในสกุลPaulinella —P. chromatophora, P. microporaและP. longichromatophora ที่อาศัยอยู่ในทะเล — มีคลอโรพลาสต์ที่วิวัฒนาการขึ้นเองโดยอิสระเพียงชนิดเดียว ซึ่งมักเรียกว่าโครมาโทฟอร์ [ หมายเหตุ 1 ]ในขณะที่คลอโรพลาสต์อื่นๆ ทั้งหมดมีต้นกำเนิดมาจากเหตุการณ์เอนโดซิมไบโอติกโบราณเพียงครั้งเดียวPaulinellaได้รับไซยาโนแบคทีเรียเอนโดซิมไบโอติกจากสกุลSynechococcus โดยอิสระ เมื่อประมาณ 90 – 140 ล้านปีก่อน[ 27 ] [ 29 ] เซลล์ Paulinellaแต่ละเซลล์มีคลอโรพลาสต์รูปทรงไส้กรอกหนึ่งหรือสองอัน[ 21 ] [ 46 ]ซึ่งได้รับการอธิบายครั้งแรกในปี 1894 โดยนักชีววิทยาชาวเยอรมัน Robert Lauterborn [ 47 ]
โครมาโทฟอร์มีขนาดเล็กมากเมื่อเทียบกับญาติไซยาโนแบคทีเรียที่ดำรงชีวิตอิสระ และมีฟังก์ชันจำกัด ตัวอย่างเช่น มีจีโนมประมาณ 1 ล้านเบสแพร์ ซึ่ง มีขนาดเพียงหนึ่งในสามของ จีโนม Synechococcusและเข้ารหัสโปรตีนเพียงประมาณ 850 ชนิด[ 21 ]อย่างไรก็ตาม ขนาดนี้ก็ยังใหญ่กว่าจีโนมคลอโรพลาสต์อื่นๆ ซึ่งโดยทั่วไปจะมีขนาดประมาณ 150,000 เบสแพร์ โครมาโทฟอร์ยังถ่ายโอน DNA ไปยังนิวเคลียสของโฮสต์น้อยกว่ามาก ประมาณ 0.3–0.8% ของ DNA ในนิวเคลียสของPaulinellaมาจากโครมาโทฟอร์ เมื่อเทียบกับ 11–14% จากคลอโรพลาสต์ในพืช[ 46 ]เช่นเดียวกับคลอโรพลาสต์อื่นๆPaulinellaให้โปรตีนเฉพาะแก่โครมาโทฟอร์โดยใช้ลำดับเป้าหมายเฉพาะ[ 48 ]เนื่องจากโครมาโทฟอร์มีอายุน้อยกว่าคลอโรพลาสต์แบบดั้งเดิมมาก จึงมีการศึกษา Paulinella chromatophoraเพื่อทำความเข้าใจว่าคลอโรพลาสต์ในยุคแรกเริ่มมีวิวัฒนาการอย่างไร[ 21 ]
สายพันธุ์คลอโรพลาสต์ขั้นที่สองและขั้นที่สาม
คลอโรพลาสต์ที่ได้จากสาหร่ายสีเขียว
สาหร่ายสีเขียวถูกนำไปใช้โดยหลายกลุ่มในเหตุการณ์แยกกันสามหรือสี่ครั้ง[ 49 ]โดยหลักแล้ว คลอโรพลาสต์รองที่ได้มาจากสาหร่ายสีเขียวจะอยู่ในยูเกลนอยด์และคลอราแรค นิโอไฟต์ นอกจากนี้ยังพบในสายพันธุ์หนึ่งของไดโนแฟลเจลเลต[ 32 ]และอาจเป็นบรรพบุรุษของสายพันธุ์ CASH ( คริปโตโมนาดอัลวีโอเลต สตราเมโนไพล์และแฮปโทไฟต์ ) [ 50 ]คลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียวจำนวนมากมีไพรีนอยด์แต่ต่างจากคลอโรพลาสต์ในบรรพบุรุษของสาหร่ายสีเขียวตรงที่ ผลิตภัณฑ์ที่เก็บสะสมจะรวมตัวกันเป็นเม็ดอยู่นอกคลอโรพลาสต์[ 12 ]
ยูเกลโนไฟต์
ยูเกลโนไฟต์เป็นกลุ่มของโปรติสต์ที่มีแฟลเจลลา ทั่วไปซึ่งมีคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียว [ 29 ]ยูเกลโนไฟต์เป็นกลุ่มเดียวนอกเหนือจากไดอะโฟเรติเคสที่มีคลอโรพลาสต์โดยไม่ต้องทำการเคลปโทพลาสตี[ 51 ] [ 52 ]คลอโรพลาสต์ของยูเกลโนไฟต์มีเยื่อหุ้มสามชั้น เชื่อกันว่าเยื่อหุ้มของโฮสต์เอนโดซิมไบออนต์หลักสูญหายไป (เช่น เยื่อหุ้มของสาหร่ายสีเขียว) เหลือเพียงเยื่อหุ้มไซยาโนแบคทีเรียสองชั้นและเยื่อหุ้มฟาโกโซมของโฮสต์รอง[ 29 ]คลอโรพลาสต์ของยูเกลโนไฟต์มีไพรีนอยด์และไทลาคอยด์เรียงซ้อนกันเป็นกลุ่มละสามอัน คาร์บอนที่ถูกตรึงผ่านการสังเคราะห์แสงจะถูกเก็บไว้ในรูปของพาราไมลอนซึ่งบรรจุอยู่ในเม็ดที่ล้อมรอบด้วยเยื่อหุ้มในไซโตพลาสซึมของยูเกลโนไฟต์[ 12 ] [ 32 ]
คลอราแรคนิโอไฟต์

คลอราแรคนิโอไฟต์เป็นกลุ่มสิ่งมีชีวิตที่หายากซึ่งมีคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียวเช่นกัน[ 29 ]แม้ว่าเรื่องราวของพวกมันจะซับซ้อนกว่ายูเกลโนไฟต์ก็ตาม บรรพบุรุษของคลอราแรคนิโอไฟต์เชื่อกันว่าเป็นยูคาริโอตที่มี คลอโรพลาสต์ที่ได้มาจากสาหร่าย สีแดงจากนั้นเชื่อกันว่ามันสูญเสียคลอโรพลาสต์สาหร่ายสีแดงอันแรกไป และต่อมาได้กลืนกินสาหร่ายสีเขียว ทำให้ได้รับคลอโรพลาสต์อันที่สองซึ่งได้มาจากสาหร่ายสีเขียว[ 32 ]
คลอโรพลาสต์ของคลอราแรคนิโอไฟต์ถูกล้อมรอบด้วยเยื่อหุ้มสี่ชั้น ยกเว้นบริเวณใกล้เยื่อหุ้มเซลล์ ซึ่งเยื่อหุ้มคลอโรพลาสต์จะรวมกันเป็นเยื่อหุ้มสองชั้น[ 12 ]ไทลาคอยด์ของพวกมันเรียงตัวเป็นกองหลวมๆ สามกอง[ 12 ]คลอราแรคนิโอไฟต์มีพอลิแซ็กคาไรด์ชนิดหนึ่งที่เรียกว่าคริโซลามินารินซึ่งพวกมันเก็บไว้ในไซโตพลาสซึม[ 32 ]มักจะสะสมอยู่รอบๆไพรีนอยด์ ของคลอโรพลา สต์ ซึ่งยื่นออกมาในไซโตพลาสซึม[ 12 ]
คลอโรพลาสต์ของคลอราราคนิโอไฟต์มีความโดดเด่นเนื่องจากสาหร่ายสีเขียวที่พวกมันได้รับมานั้นไม่ได้ถูกทำลายอย่างสมบูรณ์—นิวเคลียสของมันยังคงอยู่เป็นนิวคลีโอมอร์ฟ[ 29 ]ซึ่งพบอยู่ระหว่างเยื่อหุ้มคลอโรพลาสต์ที่สองและสาม[ 12 ] — ช่องว่างเพริพลาสมิกซึ่งสอดคล้องกับไซโตพลาสซึมของสาหร่ายสีเขียว[ 32 ]
คลอโรพลาสต์ที่ได้จากพราซิโนไฟต์

ไดโนแฟลเจลเลตในสกุลLepidodiniumได้สูญเสียคลอโรพลาสต์เพอริดินินดั้งเดิมไปและแทนที่ด้วยคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียว (โดยเฉพาะอย่างยิ่งพราซิโนไฟต์ ) [ 12 ] [ 53 ] Lepidodiniumเป็นไดโนแฟลเจลเลตเพียงชนิดเดียวที่มีคลอโรพลาสต์ที่ไม่ได้มาจาก สายพันธุ์ โรโดพลาสต์คลอโรพลาสต์นี้ถูกล้อมรอบด้วยเยื่อหุ้มสองชั้นและไม่มีนิวคลีโอเมอร์ฟ—ยีนนิวคลีโอเมอร์ฟทั้งหมดถูกถ่ายโอนไปยังนิวเคลียสของไดโนไฟต์[ 53 ]เหตุการณ์เอนโดซิมไบโอซิสที่นำไปสู่คลอโรพลาสต์นี้คือเอนโดซิมไบโอซิสทุติยภูมิแบบอนุกรมมากกว่าเอนโดซิมไบโอซิสตติยภูมิ—เอนโดซิมไบออนต์คือสาหร่ายสีเขียวที่มีคลอโรพลาสต์ปฐมภูมิ (ทำให้เกิดคลอโรพลาสต์ทุติยภูมิ) [ 32 ]
ภาวะพึ่งพาอาศัยกันแบบสามฝ่าย

ซิลิเอตPseudoblepharisma tenueมีแบคทีเรียที่เป็นซิมไบโอต 2 ชนิด ชนิดหนึ่งสีชมพู อีกชนิดหนึ่งสีเขียว ในปี 2021 ได้มีการยืนยันว่าซิมไบโอตทั้งสองชนิดสามารถสังเคราะห์แสงได้ ได้แก่ Ca. Thiodictyon intracellulare ( Chromatiaceae ) ซึ่งเป็นแบคทีเรียกำมะถันสีม่วงที่มีจีโนมขนาดเพียงครึ่งหนึ่งของญาติที่ใกล้เคียงที่สุดที่รู้จัก และChlorella sp. K10 ซึ่งเป็นสาหร่ายสีเขียว[ 54 ]นอกจากนี้ยังมีPseudoblepharisma tenue สายพันธุ์หนึ่ง ที่มีเฉพาะคลอโรพลาสต์จากสาหร่ายสีเขียวและไม่มีแบคทีเรียสีม่วงที่เป็นเอนโดซิมไบโอติก[ 55 ]
คลอโรพลาสต์ที่ได้จากสาหร่ายแดง
คลอโรพลาสต์ทุติยภูมิที่ได้มาจากสาหร่ายแดงดูเหมือนจะถูกรับมาเพียงครั้งเดียวเท่านั้น จากนั้นจึงแตกแขนงออกเป็นกลุ่มใหญ่ที่เรียกว่าโครมิสต์หรือโครมัลวีโอเลต ปัจจุบันพบได้ในแฮปโทไฟต์คริปโตโมนาด เฮเท อ โรค อนต์ ไดโน แฟล เจลเลตและอะพิคอมเพล็กซาน (สายพันธุ์ CASH) [ 32 ]คลอโรพลาสต์ทุติยภูมิของสาหร่ายแดงมักมีคลอโรฟิลล์ซีและล้อมรอบด้วยเยื่อหุ้มสี่ชั้น[ 12 ]
อย่างไรก็ตามทฤษฎีโมโนฟิลี ของโครมิสต์ ถูกปฏิเสธ และถือว่ามีความเป็นไปได้มากกว่าที่โครมิสต์บางชนิดได้รับพลาสติดโดยการรวมโครมิสต์อื่นเข้ามาแทนที่จะสืบทอดมาจากบรรพบุรุษร่วมกันคริปโตไฟต์ดูเหมือนจะได้รับพลาสติดจากสาหร่ายสีแดง ซึ่งต่อมาถูกส่งต่อจากพวกมันไปยังทั้งเฮเทอโรคอนโตไฟต์และแฮปโตไฟต์และจากกลุ่มหลังนี้ไปยังไมโซซัว[ 56 ]
คริปโตไฟต์

คริปโตไฟต์หรือคริปโตโมนาด เป็นกลุ่มสาหร่ายที่มีคลอโรพลาสต์ที่ได้มาจากสาหร่ายแดง คลอโรพลาสต์ของคริปโตไฟต์มี นิ วคลีโอเมอร์ฟที่มีลักษณะคล้ายคลึงกับของคลอราแรคนิโอไฟต์ [ 29 ] คลอโรพลาสต์ของคริปโตไฟต์มีเยื่อหุ้มสี่ชั้น เยื่อหุ้มชั้นนอกสุดต่อเนื่องกับเอนโดพลาสมิกเรติคูลัมแบบหยาบพวกมันสังเคราะห์แป้ง ธรรมดา ซึ่งถูกเก็บไว้ในเม็ดที่พบในช่องว่างรอบพลาสติด—นอกเยื่อหุ้มสองชั้นดั้งเดิม ในตำแหน่งที่สอดคล้องกับไซโตพลาสซึมของสาหร่ายแดงบรรพบุรุษ ภายในคลอโรพลาสต์ของคริปโตไฟต์มีไพรีนอยด์และไทลาคอยด์เรียงซ้อนกันเป็นชั้นละสองอัน[ 12 ]คลอโรพลาสต์ของคริปโตไฟต์ไม่มีไฟโคบิลิโซม[ 12 ]แต่มีรงควัตถุไฟโคบิลิลินซึ่งเก็บไว้ในช่องว่างไทลาคอยด์แทนที่จะยึดอยู่ด้านนอกของเยื่อไทลาคอยด์[ 12 ] [ 29 ]
คริปโตไฟต์อาจมีบทบาทสำคัญในการแพร่กระจายคลอโรพลาสต์จากสาหร่ายสีแดง[ 57 ] [ 58 ]
แฮปโทไฟต์

แฮปโทไฟต์มีความคล้ายคลึงและมีความสัมพันธ์ใกล้ชิดกับคริปโทไฟต์หรือเฮเทอโรคอนโทไฟต์[ 32 ]คลอโรพลาสต์ของพวกมันไม่มีนิวคลีโอเมอร์ฟ[ 12 ] [ 29 ]ไทลาคอยด์ของพวกมันเรียงซ้อนกันเป็นชั้นละสาม และพวกมันสังเคราะห์ น้ำตาล ไครโซลามินารินซึ่งถูกเก็บไว้ในเม็ดเล็กๆ ที่อยู่นอกคลอโรพลาสต์โดยสมบูรณ์ ในไซโทพลาสซึมของแฮปโทไฟต์[ 12 ]
Stramenopiles (เฮเทอโรคอนโตไฟต์)

Stramenopiles หรือที่เรียกว่าเฮเทอโรคอนโตไฟ ต์เป็นกลุ่มยูคาริโอตที่มีขนาดใหญ่มากและหลากหลาย ประกอบด้วยOchrophytaซึ่งประกอบด้วยไดอะตอมสาหร่ายสีน้ำตาล (สาหร่ายทะเล) และสาหร่ายสีทอง (chrysophytes) [ 39 ]และXanthophyceae (เรียกอีกอย่างว่าสาหร่ายสีเหลืองแกมเขียว) [ 32 ]
คลอโรพลาสต์ของเฮเทอโรคอนต์มีความคล้ายคลึงกับคลอโรพลาสต์ของแฮปโทไฟต์มาก พวกมันมีไพรีนอยด์ไทลาคอยด์สามอัน และโดยส่วนใหญ่[ 12 ] มี เยื่อหุ้มพลาสติดสี่ชั้น โดยเยื่อหุ้มชั้นนอกสุดเชื่อมต่อกับเอนโดพลาสมิกเรติคูลัม เช่นเดียวกับแฮปโทไฟต์ สตราเมโนไพล์เก็บน้ำตาลไว้ในเม็ดไครโซลามินาริน ในไซโตพลาสซึม [ 12 ] คลอโรพลาส ต์ของสตราเมโนไพล์มีคลอโรฟิลล์เอและโดยส่วนใหญ่[ 12 ] มี คลอโรฟิลล์ซี[ 29 ]พวกมันยังมีแคโรทีนอยด์ซึ่งทำให้พวกมันมีสีสันมากมาย[ 39 ]
อะพิคอมเพล็กซาน โครเมอริด และไดโนไฟต์
อัลวีโอเลตเป็นกลุ่มหลักของยูคาริโอตเซลล์เดียวที่มีทั้ง สมาชิก ที่สร้างอาหารเองได้และ สมาชิก ที่ต้องพึ่งพาอาหารจากภายนอกสมาชิกหลายกลุ่มมีพลาสติดที่ได้มาจากสาหร่ายสีแดง ลักษณะเด่นอย่างหนึ่งของกลุ่มที่หลากหลายนี้คือการสูญเสียการสังเคราะห์แสงบ่อยครั้ง อย่างไรก็ตาม เฮเทอโรโทรฟส่วนใหญ่ยังคงประมวลผลพลาสติดที่ไม่สังเคราะห์แสงได้[ 59 ]
อะพิคอมเพล็กซาน

อะพิคอมเพล็กซานเป็นกลุ่มของอัลวีโอเลต เช่นเดียวกับเฮลิโคสปรอยเดียพวกมันเป็นปรสิตและมีคลอโรพลาสต์ที่ไม่สามารถสังเคราะห์แสงได้[ 32 ]ครั้งหนึ่งเคยคิดว่าพวกมันมีความเกี่ยวข้องกับเฮลิโคสปรอยเดีย แต่ปัจจุบันเป็นที่ทราบกันแล้วว่าเฮลิโคสปรอยเดียเป็นสาหร่ายสีเขียวมากกว่าที่จะเป็นส่วนหนึ่งของสายพันธุ์ CASH [ 32 ]อะพิคอมเพล็กซานรวมถึงพลาสโมเดีย ม ซึ่งเป็นปรสิต มาลาเรียอะพิ คอมเพ ล็กซานจำนวนมากยังคงมีคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีแดงที่เหลืออยู่[ 60 ] [ 32 ]เรียกว่าอะพิโคพลาสต์ซึ่งพวกมันได้รับสืบทอดมาจากบรรพบุรุษ อะพิโคพลาสต์สูญเสียหน้าที่การสังเคราะห์แสงทั้งหมด และไม่มีรงควัตถุสังเคราะห์แสงหรือไทลาคอยด์ที่แท้จริง พวกมันถูกล้อมรอบด้วยเยื่อหุ้มสี่ชั้น แต่เยื่อหุ้มเหล่านี้ไม่ได้เชื่อมต่อกับเอนโดพลาสมิกเรติคูลัม[ 12 ]อะพิคอมเพล็กซานอื่นๆ เช่นคริปโตสปอริเดียมสูญเสียคลอโรพลาสต์ไปอย่างสมบูรณ์[ 60 ]อะพิคอมเพล็กซานเก็บพลังงานไว้ใน เม็ด อะไมโลเพคตินที่อยู่ในไซโตพลาสซึม แม้ว่าพวกมันจะไม่สามารถสังเคราะห์แสงได้ก็ตาม[ 12 ]
ข้อเท็จจริงที่ว่าอะพิคอมเพล็กซานยังคงมีคลอโรพลาสต์ที่ไม่สังเคราะห์แสงอยู่ แสดงให้เห็นว่าคลอโรพลาสต์ทำหน้าที่สำคัญอื่นๆ นอกเหนือจากการสังเคราะห์แสง คลอโรพลา ส ต์ ของพืชให้สิ่งสำคัญหลายอย่างแก่เซลล์พืชนอกเหนือจากน้ำตาล และอะพิโคพลาสต์ก็ไม่แตกต่างกัน พวกมันสังเคราะห์กรดไขมันไอโซเพนเทนิลไพโรฟอสเฟตกลุ่มเหล็ก-กำมะถันและทำหน้าที่ส่วนหนึ่งของวิถีฮีม[ 60 ]หน้าที่ที่สำคัญที่สุดของอะพิโคพลาสต์คือ การสังเคราะห์ ไอโซเพนเทนิลไพโรฟอสเฟตอันที่จริง อะพิคอมเพล็กซานจะตายเมื่อมีสิ่งใดมารบกวนการทำงานของอะพิโคพลาสต์ และเมื่ออะพิคอมเพล็กซานเติบโตในอาหารเลี้ยงเชื้อที่อุดมไปด้วยไอโซเพนเทนิลไพโรฟอสเฟต พวกมันจะทิ้งออร์แกเนลล์นี้ไป[ 60 ]
โครเมอริดส์

โครเมอริดเป็นกลุ่มสาหร่ายที่พบในปะการังของออสเตรเลีย ซึ่งประกอบด้วยญาติใกล้ชิดในการสังเคราะห์แสงของอะพิคอมเพล็กซาน สมาชิกตัวแรกคือChromera veliaถูกค้นพบและแยกออกมาเป็นครั้งแรกในปี 2001 การค้นพบChromera velia ที่มีโครงสร้างคล้ายกับอะพิคอมเพล็กซาน ถือเป็นจุดเชื่อมโยงที่สำคัญในประวัติศาสตร์วิวัฒนาการของอะพิคอมเพล็กซานและไดโนไฟต์ พลาสติดของพวกมันมีเยื่อหุ้มสี่ชั้น ขาดคลอโรฟิลล์ซี และใช้ RuBisCOรูปแบบประเภท II ที่ได้มาจากการถ่ายโอนในแนวนอน[ 61 ]
ไดโนไฟต์

ไดโนแฟลเจลเลตเป็นอีกกลุ่มหนึ่งที่มีขนาดใหญ่และหลากหลายมาก โดยประมาณครึ่งหนึ่งสามารถสังเคราะห์แสงได้บางส่วน (เช่น มิกโซโทรฟิก ) [ 39 ] [ 53 ]คลอโรพลาสต์ของไดโนแฟลเจลเลตมีประวัติที่ค่อนข้างซับซ้อน คลอโรพลาสต์ของไดโนแฟลเจลเลตส่วนใหญ่เป็น คลอโรพลาสต์ที่ได้มาจาก สาหร่ายสีแดงทุติยภูมิ ไดโนแฟลเจลเลตหลายชนิดสูญเสียคลอโรพลาสต์ไป (กลายเป็นไม่สังเคราะห์แสง) บางชนิดได้ทดแทนด้วยเอนโดซิมไบโอซิสตติยภูมิ[ 63 ]บางชนิดได้ทดแทนคลอโรพลาสต์ดั้งเดิมด้วยคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียว[ 29 ] [ 32 ] [ 53 ]คลอโรพลาสต์เพอริดินินถือเป็นคลอโรพลาสต์ "ดั้งเดิม" ของไดโนไฟต์[ 53 ]ซึ่งสูญหาย ลดลง ถูกแทนที่ หรือมีร่วมกันในสายพันธุ์ไดโนไฟต์อื่นๆ อีกหลายสายพันธุ์[ 32 ]
คลอโรพลาสต์ไดโนไฟต์ที่พบได้บ่อยที่สุดคือ คลอโรพลา ส ต์ ชนิดเพอริดินิน ซึ่งมีลักษณะเฉพาะคือมี รงควัตถุแคโรทีนอยด์เพอริดินิน อยู่ ในคลอโรพลาสต์ พร้อมกับคลอโรฟิลล์เอและคลอโรฟิลล์ซี2 [ 29 ] [ 53 ]ไม่พบเพอริดินินในคลอโรพลาสต์กลุ่มอื่น[ 53 ]คลอโรพลาสต์เพอริดินินมีเยื่อหุ้ม 3 ชั้น (บางครั้ง 2 ชั้น) [ 12 ]โดยสูญเสียเยื่อหุ้มเซลล์ดั้งเดิมของเอนโดซิมไบออนต์สาหร่ายสีแดงไป[ 29 ] [ 32 ]เยื่อหุ้มชั้นนอกสุดไม่ได้เชื่อมต่อกับเอนโดพลาสมิกเรติคูลัม[ 12 ] [ 53 ] คลอโรพลาสต์ ชนิดนี้มีไพรีนอยด์และมีไทลาคอยด์เรียงซ้อนกัน 3 ชั้น พบแป้งอยู่นอกคลอโรพลาสต์[ 12 ]คลอโรพลาสต์เพอริดินินยังมี DNA ที่ลดลง อย่างมาก และแตกเป็นวงกลมเล็กๆ จำนวนมาก[ 53 ]จีโนมส่วนใหญ่ได้ย้ายไปยังนิวเคลียส และมีเพียงยีนที่สำคัญที่เกี่ยวข้องกับการสังเคราะห์แสงเท่านั้นที่ยังคงอยู่ในคลอโรพลาสต์
คลอโรพลาสต์ของไดโนไฟต์ส่วนใหญ่มี RuBisCO รูปแบบ II อย่างน้อยก็มีรงควัตถุสังเคราะห์แสงคลอโรฟิลล์เอคลอโรฟิลล์ซี2เบต้าแคโรทีน และแซนโทฟิลล์เฉพาะของไดโน ไฟต์อย่างน้อยหนึ่งชนิด( เพอริดินินไดนอกแซนทินหรือไดอะไดนอกแซนทิน ) ทำให้หลายชนิดมีสีน้ำตาลทอง[ 59 ] [ 53 ]ไดโนไฟต์ทั้งหมดเก็บแป้งไว้ในไซโตพลาสซึม และส่วนใหญ่มีคลอโรพลาสต์ที่มีไทลาคอยด์เรียงซ้อนกันสามชั้น[ 12 ]
คลอโรพลาสต์ที่ได้จากแฮปโทไฟต์

สาย พันธุ์ไดโนไฟต์ ฟูโคแซนทิน (รวมถึงKarlodiniumและKarenia ) [ 32 ]สูญเสียคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีแดงดั้งเดิม และแทนที่ด้วยคลอโรพลาสต์ใหม่ที่ได้มาจาก เอนโดซิมไบออนต์ ของแฮปโทไฟต์ทำให้เกิดพลาสติดระดับตติยภูมิเหล่านี้KarlodiniumและKareniaอาจรับเอนโดซิมไบออนต์ที่แตกต่างกัน[ 32 ]เนื่องจากคลอโรพลาสต์ของแฮปโทไฟต์มีเยื่อหุ้มสี่ชั้น จึงคาดว่าเอนโดซิมไบโอซิสระดับตติยภูมิจะสร้างคลอโรพลาสต์ที่มีเยื่อหุ้มหกชั้น โดยเพิ่มเยื่อหุ้มเซลล์ของแฮปโทไฟต์และแวคิวโอลฟาโกโซมของไดโนไฟต์[ 64 ]อย่างไรก็ตาม แฮปโทไฟต์ถูกลดจำนวนลงอย่างมาก ถูกกำจัดเยื่อหุ้มบางส่วนและนิวเคลียส เหลือเพียงคลอโรพลาสต์ (ที่มีเยื่อหุ้มสองชั้นดั้งเดิม) และอาจมีเยื่อหุ้มเพิ่มเติมอีกหนึ่งหรือสองชั้นรอบๆ[ 32 ] [ 64 ]
คลอโรพลาสต์ที่มีฟูโคแซนทินมีลักษณะเฉพาะคือมีรงควัตถุฟูโคแซนทิน (จริงๆ แล้วคือ19′-เฮกซาโนอิลออกซี-ฟูโคแซนทินและ/หรือ19′-บิวทาโนอิลออกซี-ฟูโคแซนทิน ) และไม่มีเพอริดินิน ฟูโคแซนทินยังพบในคลอโรพลาสต์ของแฮปโทไฟต์ ซึ่งเป็นหลักฐานของบรรพบุรุษ[ 53 ]
คลอโรพลาสต์ที่ได้จากไดอะตอม

ไดโนไฟต์บางชนิด เช่นKryptoperidiniumและDurinskia [ 32 ]มี คลอโรพลาสต์ที่ได้มาจาก ไดอะตอม ( เฮเทอโรคอนโทไฟต์ ) [ 29 ]คลอโรพลาสต์เหล่านี้ถูกล้อมรอบด้วยเยื่อหุ้มมากถึงห้าชั้น[ 29 ] (ขึ้นอยู่กับว่าไดอะตอมเอนโดซิมไบออนต์ทั้งหมดถูกนับรวมเป็นคลอโรพลาสต์ หรือเฉพาะคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีแดงที่อยู่ภายใน) ไดอะตอมเอนโดซิมไบออนต์ลดลงค่อนข้างน้อย—มันยังคงรักษาไมโทคอนเดรีย ดั้งเดิมไว้ [ 32 ]และมีเอนโดพลาสมิกเรติคูลัม ไรโบโซมนิวเคลียสและแน่นอนคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีแดง—แทบจะเป็นเซลล์ที่สมบูรณ์[ 66 ] ทั้งหมดอยู่ภายใน ลูเมน ของเอนโดพลาสมิกเรติคูลัมของ โฮสต์ [ 32 ]อย่างไรก็ตาม เอนโดซิมไบออนต์ของไดอะตอมไม่สามารถเก็บสะสมอาหารของตัวเองได้—พอลิแซ็กคาไรด์ที่เก็บสะสมไว้จะพบอยู่ในเม็ดเล็กๆ ในไซโตพลาสซึมของโฮสต์ไดโนไฟต์แทน[ 12 ] [ 66 ]นิวเคลียสของเอนโดซิมไบออนต์ของไดอะตอมมีอยู่ แต่คงไม่สามารถเรียกว่านิวคลีโอเมอร์ฟได้เพราะไม่แสดงสัญญาณของการลดขนาดจีโนมและอาจขยายใหญ่ขึ้น ด้วย ซ้ำ[ 32 ]ไดอะตอมถูกไดโนแฟลเจลเลตกลืนกินอย่างน้อยสามครั้ง[ 32 ]
เอนโดซิมไบออนต์ของไดอะตอมถูกล้อมรอบด้วยเยื่อหุ้มเซลล์ชั้นเดียว[ 53 ]ภายในมีคลอโรพลาสต์ที่มีเยื่อหุ้มเซลล์สี่ชั้น เช่นเดียวกับบรรพบุรุษของเอนโดซิมไบออนต์ของไดอะตอม คลอโรพลาสต์มีไทลาคอยด์และไพรีนอยด์ สามชั้น [ 66 ]
ในบางสกุล เหล่านี้ คลอโรพลาสต์ของไดอะตอมที่เป็นเอนโดซิมไบออนต์ไม่ใช่คลอโรพลาสต์เพียงชนิดเดียวในไดโนไฟต์ คลอโรพลาสต์เพอริดินินที่มีเยื่อหุ้มสามชั้นดั้งเดิมยังคงอยู่ โดยถูกแปลงเป็นจุดตา[ 29 ] [ 32 ]
การผ่าตัดตกแต่งเปลือกตา
ใน โปรติสต์แบบมิกโซโทรฟิก บางกลุ่มเช่นไดโนแฟลเจลเลต บางชนิด (เช่นไดโนฟิซิส ) คลอโรพลาสต์จะถูกแยกออกจากสาหร่ายที่ถูกจับและนำมาใช้ชั่วคราวคลอโรพลาสต์ที่ขโมยมา นี้ อาจมีอายุเพียงไม่กี่วันแล้วก็จะถูกแทนที่[ 67 ] [ 68 ]
คลอโรพลาสต์ไดโนไฟต์ที่ได้จากคริปโตไฟต์

สมาชิกของสกุลDinophysisมี คลอโรพลาสต์ ที่มีไฟโคบิลิน[ 64 ]ซึ่งได้มาจากคริปโทไฟต์ [ 29 ] อย่างไรก็ตามคริปโทไฟต์ไม่ใช่เอนโดซิมไบออนต์ ดูเหมือนว่าจะมีเพียงคลอโรพลาสต์เท่านั้นที่ถูกนำมาใช้ และคลอโรพลาสต์นั้นถูกแยกนิวคลีโอเมอร์ฟและเยื่อหุ้มสองชั้นนอกสุดออกไป เหลือเพียงคลอโรพลาสต์ที่มีเยื่อหุ้มสองชั้น คลอโรพลาสต์ของคริปโทไฟต์ต้องการนิวคลีโอเมอร์ฟเพื่อดำรงอยู่ และ สายพันธุ์ Dinophysisที่ปลูกในวัฒนธรรมเซลล์เพียงอย่างเดียวไม่สามารถอยู่รอดได้ ดังนั้นจึงเป็นไปได้ (แต่ยังไม่ได้รับการยืนยัน) ว่า คลอโรพลาสต์ ของ Dinophysisเป็นเคลปโทพลาสต์ —หากเป็นเช่นนั้น คลอโรพลาสต์ ของ Dinophysis จะ เสื่อมสภาพ และ สายพันธุ์ Dinophysisต้องกลืนกินคริปโทไฟต์อย่างต่อเนื่องเพื่อรับคลอโรพลาสต์ใหม่มาแทนที่อันเก่า[ 53 ]
ดีเอ็นเอของคลอโรพลาสต์
คลอโรพลาสต์ เช่นเดียวกับออร์แกเนลล์เอนโดซิมไบโอติกอื่นๆ มีจีโนม แยกต่างหากจากจีโนมใน นิวเคลียสของเซลล์การมีอยู่ของดีเอ็นเอคลอโรพลาสต์ (cpDNA) ได้รับการระบุทางชีวเคมีในปี 1959 [ 69 ]และได้รับการยืนยันโดยกล้องจุลทรรศน์อิเล็กตรอนในปี 1962 [ 70 ]การค้นพบว่าคลอโรพลาสต์มีไรโบโซม[ 71 ]และทำการสังเคราะห์โปรตีน[ 72 ]เผยให้เห็นว่าคลอโรพลาสต์มีความเป็นอิสระทางพันธุกรรมบางส่วน ดีเอ็นเอคลอโรพลาสต์ได้รับการจัดลำดับครั้งแรกในปี 1986 [ 73 ] ตั้งแต่นั้นมา มี การจัดลำดับจีโนมคลอโรพลาสต์หลายร้อยจีโนมจากหลากหลายสายพันธุ์แต่ส่วนใหญ่เป็นพืชบกและสาหร่ายสีเขียว — กลอโคไฟต์สาหร่ายสีแดงและกลุ่มสาหร่ายอื่นๆ มีจำนวนน้อยมาก ซึ่งอาจทำให้เกิดอคติในมุมมองเกี่ยวกับโครงสร้างและเนื้อหาของดีเอ็นเอคลอโรพลาสต์ "ทั่วไป" [ 74 ]
โครงสร้างโมเลกุล
โดยส่วนใหญ่แล้ว คลอโรพลาสต์จะมีจีโนมทั้งหมดรวมกันเป็นโมเลกุล DNA วงกลมขนาดใหญ่เพียงโมเลกุลเดียว[ 74 ] ซึ่งโดยทั่วไป มีความยาว120,000–170,000 คู่เบส[ 75 ] [ 76 ] [ 77 ] [ 18 ]และมีมวลประมาณ 80–130 ล้านดาลตัน [ 78 ] แม้ว่าจีโนมของคลอโรพลาสต์จะสามารถประกอบเป็นแผนที่วงกลมได้เกือบตลอดเวลา แต่โมเลกุล DNA ทางกายภาพภายในเซลล์จะมีรูปแบบเชิงเส้นและแตกแขนงที่หลากหลาย[ 74 ] [ 79 ]คลอโรพลาสต์ใหม่ๆ อาจมีสำเนาจีโนมได้มากถึง 100 ชุด[ 75 ]แม้ว่าจำนวนสำเนาจะลดลงเหลือประมาณ 15–20 ชุดเมื่อคลอโรพลาสต์มีอายุมากขึ้น[ 80 ]
ดีเอ็นเอของคลอโรพลาสต์มักจะถูกอัดแน่นเป็นนิวคลีออยด์ซึ่งสามารถบรรจุสำเนาจีโนมของคลอโรพลาสต์ได้หลายชุด นิวคลีออยด์จำนวนมากสามารถพบได้ในคลอโรพลาสต์แต่ละอัน[ 78 ]ในสาหร่ายแดง ดั้งเดิม นิวคลีออยด์ดีเอ็นเอของคลอโรพลาสต์จะรวมกลุ่มกันอยู่ที่ใจกลางของคลอโรพลาสต์ ในขณะที่ในพืชสีเขียวและสาหร่ายสีเขียว นิ วคลี ออยด์จะกระจายอยู่ทั่วสโตรมา [ 81 ] ดีเอ็นเอของคลอโรพลาสต์ไม่ได้เกี่ยวข้องกับฮิสโตน ที่แท้จริง ซึ่งเป็นโปรตีนที่ใช้ในการบรรจุโมเลกุลดีเอ็นเอให้แน่นในนิวเคลียสของยูคาริโอต[ 17 ]แม้ว่าในสาหร่ายแดงโปรตีนที่คล้ายกันจะบรรจุวงแหวนดีเอ็นเอของคลอโรพลาสต์แต่ละวงในนิวคลีออยด์ให้ แน่น [ 81 ]
จีโนมคลอโรพลาสต์จำนวนมากมีลำดับซ้ำแบบกลับหัว สองชุด ซึ่งคั่นระหว่างส่วนสำเนาเดี่ยวที่ยาว (LSC) กับส่วนสำเนาเดี่ยวที่สั้น (SSC) [ 77 ]ลำดับซ้ำแบบกลับหัวแต่ละคู่แทบจะไม่เหมือนกัน แต่มักจะคล้ายคลึงกันมาก ซึ่งเห็นได้ชัดว่าเป็นผลมาจากการวิวัฒนาการร่วมกัน [ 74 ] ลำดับซ้ำแบบกลับหัวมีความยาวแตกต่างกันอย่างมาก โดยมีความยาวตั้งแต่ 4,000 ถึง 25,000 คู่เบสและมีจำนวนยีนน้อยที่สุดสี่หรือมากที่สุดมากกว่า 150 ยีน[ 74 ]บริเวณลำดับซ้ำแบบกลับหัวได้รับการอนุรักษ์ไว้ อย่างสูง ในพืชบก และมีการกลายพันธุ์สะสมน้อย[ 77 ] [ 82 ]
ลำดับซ้ำแบบกลับหัวที่คล้ายกันนี้มีอยู่ในจีโนมของไซยาโนแบคทีเรียและคลอโรพลาสต์อีกสองสายพันธุ์ ( กลอโคไฟตาและโรโดไฟซี ) ซึ่งบ่งชี้ว่าพวกมันมีมาก่อนคลอโรพลาสต์[ 74 ]จีโนมของคลอโรพลาสต์บางส่วนได้สูญเสีย[ 82 ] [ 83 ]หรือพลิกกลับลำดับซ้ำแบบกลับหัว (ทำให้กลายเป็นลำดับซ้ำแบบตรง ) [ 74 ]เป็นไปได้ว่าลำดับซ้ำแบบกลับหัวช่วยทำให้จีโนมของคลอโรพลาสต์ส่วนที่เหลือมีเสถียรภาพมากขึ้น เนื่องจากจีโนมของคลอโรพลาสต์ที่สูญเสียส่วนของลำดับซ้ำแบบกลับหัวบางส่วนมีแนวโน้มที่จะถูกจัดเรียงใหม่มากขึ้น[ 83 ]
การซ่อมแซมและการจำลองดีเอ็นเอ
ในคลอโรพลาสต์ของมอสPhyscomitrella patensโปรตีนซ่อมแซม DNA ที่ไม่ตรงกัน Msh1 จะทำปฏิกิริยากับโปรตีนซ่อมแซมการรวมตัวใหม่RecAและ RecG เพื่อรักษาเสถียรภาพ ของ จีโนม คลอโรพลาสต์ [ 84 ]ในคลอโรพลาสต์ของพืชArabidopsis thalianaโปรตีนRecAจะรักษาความสมบูรณ์ของ DNA ของคลอโรพลาสต์โดยกระบวนการที่น่าจะเกี่ยวข้องกับการซ่อมแซมความเสียหายของ DNA โดยการรวมตัว ใหม่[ 85 ]

กลไกการจำลองดีเอ็นเอของคลอโรพลาสต์ (cpDNA) ยังไม่ได้รับการกำหนดอย่างแน่ชัด แต่มีการเสนอแบบจำลองหลักสองแบบ นักวิทยาศาสตร์พยายามสังเกตการจำลองคลอโรพลาสต์ผ่านกล้องจุลทรรศน์อิเล็กตรอนตั้งแต่ทศวรรษ 1970 [ 86 ] [ 87 ]ผลการทดลองด้วยกล้องจุลทรรศน์นำไปสู่แนวคิดที่ว่าดีเอ็นเอของคลอโรพลาสต์จำลองโดยใช้ลูปการเคลื่อนที่แบบคู่ (D-loop) เมื่อ D-loop เคลื่อนที่ผ่านดีเอ็นเอแบบวงกลม มันจะใช้รูปแบบตัวกลางเธต้า หรือที่รู้จักกันในชื่อตัวกลางการจำลองของ Cairns และเสร็จสิ้นการจำลองด้วยกลไกวงกลมกลิ้ง[ 86 ] [ 88 ]การถอดรหัสเริ่มต้นที่จุดกำเนิดเฉพาะ ส้อมการจำลองหลายอันเปิดออก ทำให้เครื่องจักรการจำลองสามารถถอดรหัสดีเอ็นเอได้ เมื่อการจำลองดำเนินต่อไป ส้อมจะเติบโตและในที่สุดก็มาบรรจบกัน โครงสร้าง cpDNA ใหม่จะแยกออกจากกัน สร้างโครโมโซม cpDNA ลูกสาว
นอกจากการทดลองด้วยกล้องจุลทรรศน์ในช่วงแรกแล้ว แบบจำลองนี้ยังได้รับการสนับสนุนจากปริมาณการดีอะมิเนชันที่พบใน cpDNA อีกด้วย [ 86 ]การดีอะมิเนชันเกิดขึ้นเมื่อหมู่เอมีนหายไป และเป็นการกลายพันธุ์ที่มักส่งผลให้เกิดการเปลี่ยนแปลงของเบส เมื่ออะดีนีนถูกดีอะมิเนชัน มันจะกลายเป็นไฮโป แซนทีน ไฮโปแซนทีนสามารถจับกับไซโตซีนได้ และเมื่อคู่เบส XC ถูกจำลอง มันจะกลายเป็น GC (ดังนั้น การเปลี่ยนแปลงเบส A → G) [ 89 ]

ใน cpDNA มีการไล่ระดับการดีอะมิเนชัน A → G หลายระดับ DNA จะไวต่อเหตุการณ์ดีอะมิเนชันเมื่อเป็นสายเดี่ยว เมื่อเกิดฟอร์กการจำลองแบบ สายที่ไม่ถูกคัดลอกจะเป็นสายเดี่ยว และมีความเสี่ยงต่อการดีอะมิเนชัน A → G ดังนั้น การไล่ระดับในการดีอะมิเนชันจึงบ่งชี้ว่าฟอร์กการจำลองแบบน่าจะมีอยู่ และทิศทางที่ฟอร์กเปิดขึ้นในตอนแรก (การไล่ระดับสูงสุดน่าจะอยู่ใกล้กับไซต์เริ่มต้นมากที่สุด เพราะเป็นสายเดี่ยวเป็นเวลานานที่สุด) [ 86 ]กลไกนี้ยังคงเป็นทฤษฎีชั้นนำในปัจจุบัน อย่างไรก็ตาม ทฤษฎีที่สองแนะนำว่า cpDNA ส่วนใหญ่เป็นเส้นตรงและจำลองแบบผ่านการรวมตัวแบบโฮโมโลจัส นอกจากนี้ยังกล่าวอีกว่ามีเพียงส่วนน้อยของวัสดุพันธุกรรมเท่านั้นที่เก็บไว้ในโครโมโซมวงกลม ในขณะที่ส่วนที่เหลืออยู่ในโครงสร้างแบบแตกแขนง เส้นตรง หรือโครงสร้างที่ซับซ้อนอื่นๆ[ 86 ] [ 88 ]
หนึ่งในแบบจำลองการแข่งขันสำหรับการจำลองแบบ cpDNA ยืนยันว่า cpDNA ส่วนใหญ่เป็นเส้นตรงและมีส่วนร่วมในการรวมตัวแบบโฮโมโลจัสและโครงสร้างการจำลองแบบที่คล้ายกับโครงสร้าง DNA แบบเส้นตรงและวงกลมของแบคทีริโอเฟจ T4 [ 88 ] [ 90 ] เป็นที่ทราบกันดีว่าพืชบางชนิดมี cpDNA แบบเส้นตรง เช่น ข้าวโพด และยังมีอีกหลายสายพันธุ์ที่มีโครงสร้างที่ซับซ้อนซึ่งนักวิทยาศาสตร์ยังไม่เข้าใจ[ 88 ]เมื่อทำการทดลอง cpDNA ครั้งแรก นักวิทยาศาสตร์สังเกตเห็นโครงสร้างแบบเส้นตรง อย่างไรก็ตาม พวกเขาระบุว่ารูปแบบเส้นตรงเหล่านี้เกิดจากวงกลมที่แตกหัก[ 88 ]หากโครงสร้างแบบแตกแขนงและซับซ้อนที่เห็นในการทดลอง cpDNA เป็นของจริงและไม่ใช่สิ่งประดิษฐ์ของ DNA วงกลมที่ต่อกันหรือวงกลมที่แตกหัก กลไกการจำลองแบบ D-loop ก็ไม่เพียงพอที่จะอธิบายว่าโครงสร้างเหล่านั้นจะจำลองแบบได้อย่างไร[ 88 ]ในขณะเดียวกัน การรวมตัวแบบโฮโมโลจัสไม่ได้ขยายการไล่ระดับ A --> G หลายรายการที่เห็นในพลาสโตม[ 86 ]เนื่องจากความล้มเหลวในการอธิบายการไล่ระดับการกำจัดอะมิโน รวมถึงพืชหลายชนิดที่แสดงให้เห็นว่ามี cpDNA แบบวงกลม ทฤษฎีหลักจึงยังคงถือว่า cpDNA ส่วนใหญ่เป็นแบบวงกลมและน่าจะจำลองแบบผ่านกลไก D loop
เนื้อหายีนและการสังเคราะห์โปรตีน
ไซยาโนแบคทีเรียบรรพบุรุษที่นำไปสู่คลอโรพลาสต์น่าจะมีจีโนมที่มีมากกว่า 3,000 ยีน แต่ในจีโนมคลอโรพลาสต์ในปัจจุบันเหลือเพียงประมาณ 100 ยีนเท่านั้น[ 18 ] [ 22 ] [ 76 ]ยีนเหล่านี้เข้ารหัสสำหรับสิ่งต่างๆ มากมาย ส่วนใหญ่เกี่ยวข้องกับกระบวนการสร้างโปรตีนและการสังเคราะห์แสงเช่นเดียวกับในโปรคาริ โอต ยีนในดีเอ็นเอของคลอโรพลาสต์จะถูกจัดเรียงเป็นโอเปรอน[ 22 ]ซึ่งแตกต่างจากโมเลกุลดีเอ็นเอของโปรคาริ โอต โมเลกุลดีเอ็นเอของคลอโรพลาสต์มี อินทรอน ( ดีเอ็นเอไมโทคอนเด รียของพืช ก็มีเช่นกัน แต่ดีเอ็นเอไมโทคอนเดรียของมนุษย์ไม่มี) [ 91 ]
ในบรรดาพืชบก เนื้อหาของจีโนมคลอโรพลาสต์ค่อนข้างคล้ายคลึงกัน[ 77 ]
การลดขนาดจีโนมคลอโรพลาสต์และการถ่ายโอนยีน
เมื่อเวลาผ่านไป จีโนมของคลอโรพลาสต์หลายส่วนถูกถ่ายโอนไปยังจีโนมนิวเคลียร์ของโฮสต์[ 75 ] [ 76 ] [ 92 ]ซึ่งเป็นกระบวนการที่เรียกว่าการถ่ายโอนยีนแบบเอนโดซิมไบโอติก ส่งผลให้จีโนมของคลอโรพลาสต์ลดลง อย่างมาก เมื่อเทียบกับไซยาโนแบคทีเรียที่ดำรงชีวิตอิสระ คลอโรพลาสต์อาจมี 60–100 ยีน ในขณะที่ไซยาโนแบคทีเรียมักมีมากกว่า 1500 ยีนในจีโนม[ 93 ]เมื่อเร็วๆ นี้ มีการค้นพบพลาสติดที่ไม่มีจีโนม ซึ่งแสดงให้เห็นว่าคลอโรพลาสต์สามารถสูญเสียจีโนมได้ในระหว่างกระบวนการถ่ายโอนยีนแบบเอนโดซิมไบโอติก[ 94 ]
การถ่ายทอดยีนแบบเอนโดซิมไบโอติกเป็นวิธีที่เราทราบเกี่ยวกับคลอโรพลาสต์ที่หายไปในสายพันธุ์ CASH หลายสายพันธุ์ แม้ว่าคลอโรพลาสต์จะหายไปในที่สุด แต่ยีนที่มันบริจาคให้กับนิวเคลียสของโฮสต์เดิมยังคงอยู่ ซึ่งเป็นหลักฐานยืนยันการมีอยู่ของคลอโรพลาสต์ที่หายไป ตัวอย่างเช่น ในขณะที่ไดอะตอม (เฮเทอโรคอนโทไฟต์ ) ในปัจจุบันมีคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีแดงการมีอยู่ของ ยีน สาหร่ายสีเขียว จำนวนมาก ในนิวเคลียสของไดอะตอมเป็นหลักฐานว่าบรรพบุรุษของไดอะตอมเคยมีคลอโรพลาสต์ที่ได้มาจากสาหร่ายสีเขียวในช่วงเวลาหนึ่ง ซึ่งต่อมาถูกแทนที่ด้วยคลอโรพลาสต์สีแดง[ 50 ]
ในพืชบก ดีเอ็นเอในนิวเคลียสประมาณ 11–14% สามารถสืบย้อนกลับไปถึงคลอโรพลาสต์ได้[ 46 ]สูงถึง 18% ในArabidopsisซึ่งสอดคล้องกับยีนที่เข้ารหัสโปรตีนประมาณ 4,500 ยีน[ 95 ]มีการถ่ายโอนยีนจากดีเอ็นเอของคลอโรพลาสต์ไปยังจีโนมของนิวเคลียสในพืชบกเมื่อไม่นานมานี้[ 76 ]
จากโปรตีนประมาณ 3,000 ชนิดที่พบในคลอโรพลาสต์ ประมาณ 95% ของโปรตีนเหล่านั้นถูกเข้ารหัสโดยยีนในนิวเคลียส โปรตีนคอมเพล็กซ์จำนวนมากในคลอโรพลาสต์ประกอบด้วยซับยูนิตจากทั้งจีโนมของคลอโรพลาสต์และจีโนมของนิวเคลียสของโฮสต์ ดังนั้นการสังเคราะห์โปรตีนจึงต้องมีการประสานงานระหว่างคลอโรพลาสต์และนิวเคลียส คลอโรพลาสต์ส่วนใหญ่อยู่ภายใต้การควบคุมของนิวเคลียส แม้ว่าคลอโรพลาสต์จะสามารถส่งสัญญาณควบคุมการแสดงออกของยีนในนิวเคลียสได้เช่นกันซึ่งเรียกว่าการส่งสัญญาณย้อนกลับ [ 96 ] งานวิจัยล่าสุดระบุว่าส่วนหนึ่งของเครือข่ายการส่งสัญญาณย้อนกลับที่เคยถือว่าเป็นลักษณะเฉพาะของพืชบกได้ปรากฏขึ้นแล้วในบรรพบุรุษของสาหร่าย[ 97 ] [ 98 ] [ 99 ]โดยรวมเข้ากับกลุ่มยีนที่แสดงออกร่วมกันในญาติสาหร่ายที่ใกล้เคียงที่สุดของพืชบก[ 100 ]
การสังเคราะห์โปรตีน
การสังเคราะห์โปรตีนภายในคลอโรพลาสต์อาศัยRNA โพลีเมอเร ส 2 ชนิด ชนิดหนึ่งถูกเข้ารหัสโดย DNA ของคลอโรพลาสต์ อีกชนิดหนึ่งมี ต้น กำเนิด จากนิวเคลียส RNA โพลีเมอเรสทั้งสองชนิดอาจจดจำและจับกับ โปรโมเตอร์ชนิดต่างๆภายในจีโนมของคลอโรพลาสต์ได้[ 101 ]ไรโบโซมในคลอโรพลาสต์มีลักษณะคล้ายกับไรโบโซมของแบคทีเรีย[ 102 ]
การกำหนดเป้าหมายและการนำเข้าโปรตีน
เนื่องจากยีนคลอโรพลาสต์จำนวนมากถูกย้ายไปยังนิวเคลียสโปรตีน หลายชนิด ที่เดิมทีจะถูกแปลในคลอโรพลาสต์จึงถูกสังเคราะห์ในไซโตพลาสซึมของเซลล์พืช โปรตีนเหล่านี้จะต้องถูกส่งกลับไปยังคลอโรพลาสต์ และนำเข้าผ่านเยื่อหุ้มคลอโรพลาสต์อย่างน้อยสองชั้น[ 103 ]
ที่น่าสนใจคือ ผลิตภัณฑ์โปรตีนประมาณครึ่งหนึ่งของยีนที่ถ่ายโอนไม่ได้ถูกส่งกลับไปยังคลอโรพลาสต์ด้วยซ้ำ หลายตัวกลายเป็นexaptationซึ่งทำหน้าที่ใหม่ เช่น การมีส่วนร่วมในการแบ่งเซลล์การกำหนดเส้นทางโปรตีนและแม้แต่การต้านทานโรคยีนคลอโรพลาสต์บางส่วนพบที่อยู่ใหม่ในจีโนมไมโทคอนเดรีย ซึ่งส่วนใหญ่กลายเป็น ยีนเทียมที่ไม่ทำงาน แม้ว่ายีน tRNAบางส่วนยังคงทำงานในไมโทคอนเดรีย [ 93 ] ผลิตภัณฑ์โปรตีน DNA คลอโรพลาสต์ที่ถ่ายโอนบางส่วนถูกส่งไปยังเส้นทางการหลั่ง [ 93 ]แม้ว่าพลาสติดรอง จำนวนมากจะถูกล้อมรอบด้วยเยื่อหุ้มชั้นนอกสุดที่ได้มาจาก เยื่อหุ้มเซลล์ของโฮสต์และดังนั้นจึงอยู่นอกเซลล์ในเชิงโครงสร้าง เนื่องจากในการเข้าถึงคลอโรพลาสต์จาก ไซโตซอลจะต้องข้ามเยื่อหุ้มเซลล์ซึ่ง หมายถึงการเข้าสู่ พื้นที่นอกเซลล์ในกรณีเหล่านั้น โปรตีนที่มุ่งเป้าไปที่คลอโรพลาสต์จะเดินทางไปตามเส้นทางการหลั่งในตอนแรก[ 32 ]
เนื่องจากเซลล์ที่ได้รับคลอโรพลาสต์มีไมโตคอนเดรียอยู่แล้ว (และเพอร์ออกซิโซมและเยื่อหุ้มเซลล์สำหรับการหลั่ง) โฮสต์คลอโรพลาสต์ใหม่จึงต้องพัฒนาระบบกำหนดเป้าหมายโปรตีน เฉพาะ เพื่อหลีกเลี่ยงไม่ให้โปรตีนคลอโรพลาสต์ถูกส่งไปยังออร์แกเนลล์ที่ ไม่ถูกต้อง [ 103 ]
![ปลายทั้งสองข้างของพอลิเปปไทด์เรียกว่าปลาย N หรือปลายอะมิโน และปลาย C หรือปลายคาร์บอกซิล[104] พอลิเปปไทด์นี้มีกรดอะมิโนสี่ตัวที่เชื่อมต่อกัน ด้านซ้ายคือปลาย N ซึ่งมีหมู่อะมิโน (H2N) สีเขียว ปลาย C สีน้ำเงินซึ่งมีหมู่คาร์บอกซิล (CO2H) อยู่ทางด้านขวา](https://upload.wikimedia.org/wikipedia/commons/thumb/c/c0/Tetrapeptide_structural_formulae.svg/500px-Tetrapeptide_structural_formulae.svg.png)
ในกรณีส่วนใหญ่ แต่ไม่ใช่ทุกกรณี โปรตีนคลอโรพลาสต์ที่เข้ารหัสโดยนิวเคลียสจะถูกแปลด้วยเปปไทด์นำส่งที่สามารถแยกออกได้ซึ่งถูกเพิ่มเข้าไปที่ปลาย N ของโปรตีนต้นแบบ บางครั้งลำดับนำส่งจะอยู่ที่ปลาย C ของโปรตีน[ 105 ]หรือภายในส่วนที่ทำหน้าที่ของโปรตีน[ 103 ]
โปรตีนขนส่งและทรานสโลคอนของเยื่อหุ้มเซลล์
หลังจากที่โพลีเปปไทด์ ของคลอโรพลาสต์ ถูกสังเคราะห์บนไรโบโซมในไซโตซอลเอนไซม์เฉพาะสำหรับโปรตีนของคลอโรพลาสต์[ 106 ]จะทำการฟอส โฟรีเลต หรือเพิ่มหมู่ฟอสเฟตให้กับโปรตีนเหล่านั้นจำนวนมาก (แต่ไม่ใช่ทั้งหมด) ในลำดับการส่งผ่าน[ 103 ] การฟอสโฟรีเลตช่วยให้โปรตีนจำนวนมากจับกับโพ ลีเปปไทด์ ป้องกันไม่ให้โพลีเปปไทด์ พับตัวก่อนกำหนด[ 103 ]ซึ่งเป็นสิ่งสำคัญเพราะมันป้องกันไม่ให้โปรตีนของคลอโรพลาสต์อยู่ในรูปแบบที่ใช้งานได้และทำหน้าที่ของคลอโรพลาสต์ในที่ที่ไม่ถูกต้อง นั่นคือไซโตซอล [ 107 ] [ 108 ] ในขณะเดียวกัน พวกมันต้องรักษารูปร่างให้พอดีเพื่อให้คลอโรพลาสต์สามารถจดจำได้[ 107 ]โปรตีนเหล่านี้ยังช่วยให้โพลีเปปไทด์ถูกนำเข้าสู่คลอโรพลาสต์[ 103 ]
จากตรงนี้ โปรตีนคลอโรพลาสต์ที่มุ่งไปยังสโตรมาจะต้องผ่านคอมเพล็กซ์โปรตีนสองชุด ได้แก่คอมเพล็กซ์ TOCหรือทรานสโลคอนบนเยื่อ หุ้ม คลอโรพลาสต์ด้านนอกและท รานส โลคอน TICหรือทรานสโลคอนบน ท รานสโลคอน ของเยื่อหุ้ม คลอ โรพลาสต์ ด้าน ใน [ 103 ]สายโซ่โพลีเปปไทด์ของคลอโรพลาสต์มักจะเดินทางผ่านคอมเพล็กซ์ทั้งสองพร้อมกัน แต่คอมเพล็กซ์ TIC ยังสามารถดึงโปรตีนก่อนการสร้างที่สูญหายไปในช่องว่างระหว่างเยื่อหุ้มกลับคืนมาได้[ 103 ]
โครงสร้าง

ในพืชบกคลอโรพลาสต์โดยทั่วไปมีรูปร่างคล้ายเลนส์ มีเส้นผ่านศูนย์กลาง 3–10 ไมโครเมตร และหนา 1–3 ไมโครเมตร[ 109 ] [ 18 ]คลอโรพลาสต์ของต้นกล้าข้าวโพดมีปริมาตร ประมาณ 20 ไมโครเมตร ³ [ 18 ]ความหลากหลายของรูปร่างคลอโรพลาสต์มีมากขึ้นในสาหร่ายซึ่งมักจะมีคลอโรพลาสต์เพียงอันเดียว[ 12 ]ที่สามารถมีรูปร่างคล้ายตาข่าย (เช่นOedogonium ) [ 110 ]รูปถ้วย (เช่นChlamydomonas ) [ 111 ]รูปทรงเกลียวคล้ายริบบิ้นรอบขอบเซลล์ (เช่นSpirogyra ) [ 112 ]หรือแถบที่บิดเล็กน้อยที่ขอบเซลล์ (เช่นSirogonium ) [ 113 ]สาหร่ายบางชนิดมีคลอโรพลาสต์สองอันในแต่ละเซลล์ ใน Zygnemaคลอ โรพลาส ต์จะมีรูปร่างคล้ายดาว[ 114 ]หรืออาจมีรูปร่างคล้ายครึ่งเซลล์ในอันดับDesmidiales [ 115 ]ในสาหร่ายบางชนิด คลอโรพลาสต์จะกินพื้นที่ส่วนใหญ่ของเซลล์ โดยมีช่องสำหรับนิวเคลียสและออร์แกเนลล์อื่นๆ[ 12 ]ตัวอย่างเช่นChlorella บางชนิด มีคลอโรพลาสต์รูปถ้วยที่กินพื้นที่ส่วนใหญ่ของเซลล์[ 116 ]
คลอโรพลาสต์ทั้งหมดมีระบบเยื่อหุ้มอย่างน้อยสามระบบ ได้แก่ เยื่อหุ้มคลอโรพลาสต์ชั้นนอก เยื่อหุ้มคลอโรพลาสต์ชั้นใน และ ระบบ ไทลาคอยด์เยื่อหุ้มลิปิดสองชั้นในสุด[ 117 ]ที่ล้อมรอบคลอโรพลาสต์ทั้งหมดนั้นสอดคล้องกับเยื่อหุ้ม ชั้นนอกและชั้นในของ ผนังเซลล์แกรมลบของไซยาโนแบคทีเรียบรรพบุรุษ[ 29 ] [ 118 ] [ 119 ]และไม่ใช่ เยื่อ หุ้มฟาโกโซมจากโฮสต์ ซึ่งอาจสูญหายไปแล้ว[ 29 ]คลอโรพลาสต์ที่เป็นผลผลิตจากเอนโดซิมไบโอซิสทุติยภูมิอาจมีเยื่อหุ้มเพิ่มเติมล้อมรอบทั้งสามชั้นนี้[ 30 ]ภายในเยื่อหุ้มคลอโรพลาสต์ชั้นนอกและชั้นในคือสโตรมา ของคลอโรพลาสต์ ซึ่ง เป็นของเหลวกึ่งเจล[ 23 ]ที่ประกอบขึ้นเป็นปริมาตรส่วนใหญ่ของคลอโรพลาสต์ และมีระบบไทลาคอยด์ลอยอยู่ภายใน

มีความเข้าใจผิดทั่วไปบางประการเกี่ยวกับเยื่อหุ้มคลอโรพลาสต์ชั้นนอกและชั้นใน ข้อเท็จจริงที่ว่าคลอโรพลาสต์ถูกล้อมรอบด้วยเยื่อหุ้มสองชั้นมักถูกอ้างถึงว่าเป็นหลักฐานว่าพวกมันเป็นลูกหลานของไซยาโนแบคทีเรีย แบบเอนโดซิมไบโอติก ซึ่งมักถูกตีความว่าหมายความว่าเยื่อหุ้มคลอโรพลาสต์ชั้นนอกเป็นผลมาจากการพับตัวของ เยื่อหุ้มเซลล์ของโฮสต์เพื่อสร้างถุงล้อมรอบไซยาโนแบคทีเรีย ดั้งเดิม ซึ่งไม่เป็นความจริง เยื่อหุ้มคลอโรพลาสต์ทั้งสองชั้นมีความคล้ายคลึงกับเยื่อหุ้มสองชั้นดั้งเดิมของไซยาโนแบคทีเรีย[ 29 ]
เยื่อหุ้มสองชั้นของคลอโรพลาสต์มักถูกนำไปเปรียบเทียบกับ เยื่อหุ้มสองชั้น ของไมโทคอนเดรียแต่การเปรียบเทียบนี้ไม่ถูกต้อง เพราะเยื่อหุ้มชั้นในของไมโทคอนเดรียใช้ในการขับเคลื่อนปั๊มโปรตอนและทำการฟอสโฟรีเลชันแบบออกซิเดชันเพื่อสร้าง พลังงาน ATPโครงสร้างเดียวของคลอโรพลาสต์ที่สามารถถือได้ว่าคล้ายคลึงกันคือระบบไทลาคอยด์ภายใน ถึงกระนั้น ในแง่ของ "เข้า-ออก" ทิศทางการไหลของไอออนH + ในคลอโร พลาสต์จะอยู่ในทิศทางตรงกันข้ามกับการฟอสโฟรีเลชันแบบออกซิเดชันในไมโทคอนเดรีย[ 23 ] [ 120 ]นอกจากนี้ ในแง่ของหน้าที่ เยื่อหุ้มชั้นในของคลอโรพลาสต์ซึ่งควบคุมการผ่านของเมตาบอไลต์และสังเคราะห์วัสดุบางอย่าง ไม่มีคู่เทียบในไมโทคอนเดรีย[ 23 ]
เยื่อหุ้มคลอโรพลาสต์ชั้นนอก
เยื่อหุ้มคลอโรพลาสต์ชั้นนอกเป็นเยื่อกึ่งพรุนที่โมเลกุลขนาดเล็กและไอออนสามารถแพร่ผ่านได้ง่าย[ 121 ] อย่างไรก็ตาม โปรตีนขนาดใหญ่ไม่สามารถผ่านได้ดังนั้นโพลีเปปไทด์ ของคลอโรพลาสต์ ที่ถูกสังเคราะห์ในไซโตพลาสซึม ของเซลล์จะต้องถูกขนส่งผ่านเยื่อหุ้ม คลอ โรพลาสต์ ชั้นนอกโดยคอมเพล็กซ์ TOCหรือทรานสโลคอนบนเยื่อหุ้มคลอโรพลาสต์ ชั้น นอก[ 103 ]
บางครั้งเยื่อหุ้มคลอโรพลาสต์จะยื่นออกมาในไซโตพลาสซึม ก่อตัวเป็นสตรอมูลหรือท่อที่มีสตรอ มูล สตรอมูลพบได้น้อยมากในคลอโรพลาสต์ และพบได้บ่อยกว่ามากในพลาสติดชนิด อื่น เช่นโครโมพลา สต์ และอะไมโลพลาสต์ในกลีบดอกและรากตามลำดับ[ 122 ] [ 123 ]สตรอมูลอาจมีอยู่เพื่อเพิ่มพื้นที่ผิว ของคลอโรพลาสต์ สำหรับการขนส่งข้ามเยื่อหุ้มเซลล์ เนื่องจากมักแตกแขนงและพันกันกับเอนโดพลาสมิกเรติคูลัม [ 124 ] เมื่อมีการสังเกตพบครั้งแรกในปี 1962 นักชีววิทยาพืชบางคนปฏิเสธโครงสร้างเหล่านี้ว่าเป็นสิ่งประดิษฐ์ โดยอ้างว่าสตรอมูลเป็นเพียงคลอโรพลาสต์ที่มีรูปร่างแปลก ๆ มีบริเวณที่หดตัวหรือ คลอโรพลาส ต์ที่กำลังแบ่งตัว[ 125 ]อย่างไรก็ตาม มีหลักฐานเพิ่มมากขึ้นเรื่อยๆ ว่าสตรอมูลเป็นส่วนประกอบที่สำคัญและทำหน้าที่ของพลาสติดในเซลล์พืช ไม่ใช่เพียงแค่สิ่งประดิษฐ์[ 126 ]
ช่องว่างระหว่างเยื่อหุ้มเซลล์และผนังเพปติโดไกลแคน

โดยปกติจะมีช่องว่างระหว่างเยื่อหุ้มเซลล์บางๆ ที่มีความหนาประมาณ 10–20 นาโนเมตรอยู่ระหว่างเยื่อหุ้มคลอโรพลาสต์ชั้นนอกและชั้นใน[ 127 ]
คลอโรพลาสต์ของสาหร่าย กลอโคไฟต์มี ชั้น เพปติโดไกลแคนอยู่ระหว่างเยื่อหุ้มคลอโรพลาสต์ ซึ่งสอดคล้องกับผนังเซลล์เพปติโดไกลแคนของ บรรพบุรุษ ไซยาโนแบคทีเรียซึ่งอยู่ระหว่างเยื่อหุ้มเซลล์ทั้งสองชั้น คลอโรพลาสต์เหล่านี้เรียกว่ามูโรพลาสต์ (จากภาษาละติน"mura"ซึ่งหมายถึง "ผนัง") คลอโรพลาสต์อื่นๆ สันนิษฐานว่าสูญเสียผนังไซยาโนแบคทีเรียไป ทำให้เกิดช่องว่างระหว่างเยื่อหุ้มคลอโรพลาสต์ทั้งสองชั้น[ 23 ]แต่ต่อมาพบว่ามีอยู่ในมอส ไลโคไฟต์ และเฟิร์นด้วย[ 128 ]
เยื่อหุ้มคลอโรพลาสต์ชั้นใน
เยื่อหุ้มคลอโรพลาสต์ชั้นในติดกับสโตรมาและควบคุมการผ่านเข้าออกของสารต่างๆ ในคลอโรพลาสต์ หลังจากผ่านคอมเพล็กซ์ TOCในเยื่อหุ้มคลอโรพลาสต์ชั้นนอกแล้ว โพ ลีเปปไทด์จะต้องผ่านคอมเพล็กซ์ TIC ( ทรานสโลคอนบนเยื่อหุ้มคลอ โรพลาสต์ชั้น ใน )ซึ่งตั้งอยู่ในเยื่อหุ้มคลอโรพลาสต์ชั้นใน[ 103 ]
นอกจากการควบคุมการผ่านของสารต่างๆ แล้ว เยื่อหุ้มคลอโรพลาสต์ชั้นในยังเป็นแหล่งสังเคราะห์กรดไขมันลิปิดและแคโรทีนอยด์ อีกด้วย [ 23 ]
เรติคูลัมส่วนปลาย
คลอโรพลาสต์บางชนิดมีโครงสร้างที่เรียกว่าคลอโรพลาสต์ เพอริฟิเชียล เรติคูลัม[ 127 ]มักพบในคลอโรพลาสต์ของพืช C4แม้ว่าจะพบในพืชดอก C3 บางชนิด[ 23 ] และแม้แต่พืชเมล็ดเปลือยบาง ชนิด [ 129 ]คลอโรพลาสต์ เพอริฟิเชียล เรติคูลัมประกอบด้วยท่อและถุงเมมเบรนที่ซับซ้อนต่อเนื่องกับเยื่อหุ้มคลอโรพลาสต์ชั้นในที่ยื่นเข้าไปใน ของเหลว สโตรมา ภายใน ของคลอโรพลาสต์ เชื่อกันว่าจุดประสงค์ของมันคือการเพิ่มพื้นที่ผิว ของคลอโรพลาสต์ สำหรับการขนส่งข้ามเยื่อหุ้มระหว่างสโตรมาและไซโตพลาสซึม ของเซลล์ ถุงขนาดเล็กที่บางครั้งพบเห็นอาจทำหน้าที่เป็นถุงขนส่งเพื่อลำเลียงสารระหว่างไทลาคอยด์และช่องว่างระหว่างเยื่อหุ้ม[ 130 ]
สโตรมา
ของเหลว ที่มี โปรตีนสูง[ 23 ] เป็นด่าง[ 120 ] ใน น้ำภายในเยื่อหุ้มคลอโรพลาสต์ชั้นในและภายนอกช่องว่างไทลาคอยด์เรียกว่าสโตรมา[ 23 ]ซึ่งสอดคล้องกับไซโตซอล ของไซยา โนแบคทีเรียดั้งเดิมนิว คลีออยด์ ของดีเอ็นเอคลอโร พลา สต์ ไรโบโซมคลอโรพลาส ต์ ระบบไทลาคอยด์ที่มีพลาสโตโกลบู ลี เม็ด แป้งและโปรตีน จำนวนมาก สามารถพบได้ลอยอยู่ภายในนั้นวัฏจักรแคลวินซึ่งตรึงCO 2ให้เป็นG3Pเกิดขึ้นในสโตรมา
ไรโบโซมของคลอโรพลาสต์

คลอโรพลาสต์มีไรโบโซมของตัวเอง ซึ่งใช้ในการสังเคราะห์โปรตีนเพียงส่วนน้อย ไรโบโซมของคลอโรพลาสต์มีขนาดประมาณสองในสามของไรโบโซมในไซโตพลาสซึม (ประมาณ 17 นาโนเมตรเทียบกับ 25 นาโนเมตร ) [ 127 ]พวกมันรับmRNAที่ถอดรหัสจากDNA ของคลอโรพลาสต์และแปลเป็นโปรตีน แม้ว่าจะคล้ายกับไรโบโซมของแบคทีเรีย [ 17 ] การ แปลของคลอโรพลาสต์มีความซับซ้อนมากกว่าในแบคทีเรีย ดังนั้นไรโบโซมของคลอโรพลาสต์จึงมีคุณสมบัติเฉพาะของคลอโรพลาสต์บางประการ[ 131 ] [ 132 ]
RNA ไรโบโซมหน่วยย่อยขนาดเล็กในคลอโรพลาสต์ของ Chlorophyta และ euglenid หลายชนิดขาดโมทีฟสำหรับการจดจำลำดับ Shine-Dalgarno [ 133 ] ซึ่งถือว่าจำเป็นสำหรับการเริ่มต้นการแปลในคลอโรพลาสต์และโปรคาริโอตส่วนใหญ่ [ 134 ] [ 135 ] การสูญเสียดังกล่าวพบได้น้อยมากในพลาสติดและโปรคาริโอตอื่นๆ[ 133 ] [ 136 ]พบrRNA 4.5S เพิ่มเติมที่มีความคล้ายคลึงกับหาง 3' ของ 23S ในพืช "ชั้นสูง" [ 132 ]
พลาสโตโกลบูลี
พลาสโตโกล บูลี ( เอกพจน์ คือ พลาสโตโกลบูลัสบางครั้งสะกดว่า พลาสโตโกลบูล ) เป็นฟองทรงกลมของไขมันและโปรตีน[ 23 ]มีขนาดประมาณ 45–60 นาโนเมตร[ 137 ]ล้อมรอบด้วยชั้นไขมันชั้นเดียว [ 137 ] พบพลาสโตโกลบูลีในคลอโรพลาสต์ทั้งหมด[ 127 ]แต่จะพบได้บ่อยขึ้นเมื่อคลอโรพลาสต์อยู่ภายใต้ความเครียดจากออกซิเดชัน [ 137 ]หรือเมื่อมันแก่และเปลี่ยนไปเป็นเจอรอนโทพลาสต์ [ 23 ] พลาสโตโกลบูลียังแสดงความแปรผันของขนาดที่มากขึ้นภายใต้สภาวะเหล่านี้[ 137 ]นอกจากนี้ยังพบได้ทั่วไปใน เอทิโอพ ลาสต์แต่จำนวนจะลดลงเมื่อเอทิโอพลาสต์เจริญเติบโตเป็นคลอโรพลาสต์[ 137 ]
พลาสโตโกลบูลีประกอบด้วยโปรตีนโครงสร้างและเอนไซม์ที่เกี่ยวข้องกับ การสังเคราะห์ และการเผาผลาญไขมัน พวกมันประกอบด้วย ไขมันหลายชนิดรวมถึงพลาสโตควิโนนวิตามินอีแคโรทีนอยด์และคลอโรฟิลล์[ 137 ]
เดิมทีเชื่อกันว่าพลาสโตโกลบูลีลอยตัวอิสระอยู่ในสโตรมาแต่ปัจจุบันเชื่อกันว่าพลาสโตโกลบูลีจะยึดติดอยู่กับไทลาคอยด์ อย่างถาวร หรือยึดติดอยู่กับพลาสโตโกลบูลีอื่นที่ยึดติดอยู่กับไทลาคอยด์ ซึ่งเป็นโครงสร้างที่ทำให้พลาสโตโกลบูลีสามารถแลกเปลี่ยนเนื้อหาภายในกับเครือข่ายไทลาคอยด์ได้[ 137 ]ในคลอโรพลาสต์สีเขียวปกติ พลาสโตโกลบูลีส่วนใหญ่จะอยู่เดี่ยวๆ โดยยึดติดอยู่กับไทลาคอยด์แม่โดยตรง ในคลอโรพลาสต์ที่เก่าหรืออยู่ในสภาวะเครียด พลาสโตโกลบูลีมักจะอยู่รวมกันเป็นกลุ่มหรือเป็นโซ่ โดยยังคงยึดติดอยู่กับไทลาคอยด์เสมอ[ 137 ]
พลาสโตโกลบูลีเกิดขึ้นเมื่อมีฟองอากาศปรากฏขึ้นระหว่างชั้นของเยื่อไขมันไบเลเยอร์ของเยื่อไทลาคอยด์ หรือแตกหน่อจากพลาสโตโกลบูลีที่มีอยู่แล้ว—แม้ว่าจะไม่หลุดออกและลอยไปในสโตรมาก็ตาม[ 137 ]ในทางปฏิบัติ พลาสโตโกลบูลีทั้งหมดจะก่อตัวขึ้นบนหรือใกล้ขอบที่โค้งมากของ แผ่นหรือจาน ไทลาคอยด์ นอกจากนี้ยังพบได้บ่อยกว่าบนไทลาคอยด์ในสโตรมามากกว่าบนไทลาคอยด์ในกรานัล[ 137 ]

เม็ดแป้ง
เม็ดแป้งพบได้ทั่วไปในคลอโรพลาสต์ โดยทั่วไปจะกินพื้นที่ 15% ของปริมาตรออร์แกเนลล์[ 138 ]แม้ว่าในพลาสติดชนิดอื่น เช่นอะไมโลพลาสต์ เม็ดแป้ง อาจมีขนาดใหญ่พอที่จะทำให้รูปร่างของออร์แกเนลล์ผิดเพี้ยนไปได้[ 127 ]เม็ดแป้งเป็นเพียงการสะสมของแป้งในสโตรมา และไม่มีเยื่อหุ้มล้อมรอบ[ 127 ]
เม็ดแป้งปรากฏและเติบโตตลอดทั้งวัน เนื่องจากคลอโรพลาสต์สังเคราะห์น้ำตาลและถูกบริโภคในเวลากลางคืนเพื่อเป็นเชื้อเพลิงในการหายใจและส่งออกน้ำตาลไปยังโฟลเอ็มอย่าง ต่อเนื่อง [ 139 ]แม้ว่าในคลอโรพลาสต์ที่เจริญเต็มที่แล้ว เม็ดแป้งจะไม่ถูกบริโภคจนหมดหรือมีการสะสมเม็ดใหม่เกิดขึ้นได้ยาก[ 138 ]
เม็ดแป้งมีองค์ประกอบและตำแหน่งที่แตกต่างกันไปในคลอโรพลาสต์สายพันธุ์ต่างๆ ในสาหร่ายสีแดงเม็ดแป้งจะพบในไซโตพลาสซึมมากกว่าในคลอโรพลาสต์[ 140 ]ในพืช C4 คลอโรพลาสต์ ของมีโซฟิ ลล์ ซึ่งไม่สังเคราะห์น้ำตาลจะไม่มีเม็ดแป้ง[ 23 ]
รูบิสโก้

สโตรมาของคลอโรพลาสต์ประกอบด้วยโปรตีนหลายชนิด แม้ว่า โปรตีนที่พบได้ทั่วไปและสำคัญที่สุดคือRuBisCOซึ่งอาจเป็นโปรตีนที่มีปริมาณมากที่สุดในโลกด้วย[ 120 ] RuBisCOเป็นเอนไซม์ที่ตรึงCO2ให้เป็นโมเลกุลน้ำตาล ในพืชC3 นั้น RuBisCO มีปริมาณมากในคลอโรพลาสต์ทั้งหมด แต่ในพืชC4 นั้น RuBisCO จะจำกัดอยู่เฉพาะใน คลอโรพลาสต์ ของปลอกมัดซึ่งเป็นบริเวณที่วัฏจักรแคลวินเกิดขึ้นในพืชC4 [ 141 ]
ไพรีนอยด์
คลอโรพลาสต์ของฮอร์นเวิร์ต บางชนิด [ 142 ]และสาหร่ายมีโครงสร้างที่เรียกว่าไพรีนอยด์ซึ่งไม่พบในพืชชั้นสูง[ 143 ]ไพรีนอยด์มีรูปร่างคล้ายทรงกลมและมีการหักเหของแสงสูง ซึ่งเป็นแหล่งสะสมแป้งในพืชที่มีไพรีนอยด์ ไพรีนอยด์ประกอบด้วยเมทริกซ์ทึบแสงต่ออิเล็กตรอน ล้อมรอบด้วยแผ่นแป้งรูปครึ่งวงกลมสองแผ่น แป้งจะสะสมเมื่อไพรีนอยด์เจริญเติบโต เต็มที่ [ 144 ]ในสาหร่ายที่มีกลไกการรวมคาร์บอนเอนไซม์RuBisCOพบได้ในไพรีนอยด์ แป้งยังสามารถสะสมรอบๆ ไพรีนอยด์ได้เมื่อ CO 2ขาดแคลน[ 143 ]ไพรีนอยด์สามารถแบ่งตัวเพื่อสร้างไพรีนอยด์ใหม่ หรือสร้างขึ้นใหม่ได้[ 144 ] [ 145 ]
ระบบไทลาคอยด์

ไทลาคอยด์ (บางครั้งสะกดว่าthylakoïds ) [ 147 ]เป็นถุงเล็กๆ ที่เชื่อมต่อกันซึ่งมีเยื่อหุ้มที่ปฏิกิริยาแสงของการสังเคราะห์แสงเกิดขึ้น คำว่าไทลาคอยด์มาจากคำภาษากรีกว่าthylakosซึ่งหมายถึง "ถุง" [ 148 ]
ภายในสโตรมาของคลอโรพลาสต์จะมี ระบบ ไทลาคอยด์ซึ่งเป็นกลุ่มของถุงเยื่อบางๆ ที่เรียกว่าไทลาคอยด์ ที่มีพลวัตสูง เป็นที่ที่พบคลอโรฟิลล์ และ เกิดปฏิกิริยาแสงของการสังเคราะห์ แสง [ 11 ] ใน คลอโรพลาสต์ ของพืช มีท่อลำเลียงส่วนใหญ่ ไทลาคอยด์จะเรียงซ้อนกันเป็นชั้นๆ เรียกว่ากรานา[ 149 ]แม้ว่าในคลอโรพลาสต์ของพืชC4 บาง ชนิด [ 141 ]และ คลอโรพลาสต์ ของสาหร่าย บางชนิด ไทลาคอยด์จะลอยอยู่ได้อย่างอิสระ[ 12 ]
โครงสร้างไทลาคอยด์

การใช้กล้องจุลทรรศน์แบบใช้แสงทำให้สามารถมองเห็นเม็ดสีเขียวเล็กๆ ได้เพียงเล็กน้อย ซึ่งเรียกว่ากรานา [ 127 ] ด้วยกล้องจุลทรรศน์อิเล็กตรอนทำให้สามารถมองเห็นระบบไทลาคอยด์ได้อย่างละเอียดมากขึ้น เผยให้เห็นว่าประกอบด้วยไทลาคอยด์ แบนเรียง ซ้อนกันเป็นกรานา และไทลาคอยด์สโตรมัลที่เชื่อมต่อกันเป็นแนวยาวซึ่งเชื่อมโยงกรานาต่างๆ เข้าด้วยกัน[ 127 ] ในกล้องจุลทรรศน์อิเล็กตรอนแบบส่งผ่าน เยื่อไทลาคอยด์จะปรากฏเป็นแถบสีอ่อนและสีเข้มสลับกัน หนา 8.5 นาโนเมตร[ 127 ]
โครงสร้างสามมิติของระบบเยื่อไทลาคอยด์เป็นที่ถกเถียงกัน มีการเสนอแบบจำลองหลายแบบ โดยแบบที่แพร่หลายที่สุดคือ แบบ จำลองเกลียวซึ่งกลุ่มไทลาคอยด์ของกรานัมถูกห่อหุ้มด้วยไทลาคอยด์สโตรมัลแบบเกลียว[ 150 ]แบบจำลองอีกแบบหนึ่งที่เรียกว่า 'แบบจำลองการแตกแขนง' ซึ่งอิงจากการศึกษาเอกซเรย์คอมพิวเตอร์อิเล็กตรอนครั้งแรกของเยื่อไทลาคอยด์ของพืช แสดงให้เห็นเยื่อสโตรมัลเป็นแผ่นลามิลลาร์กว้างที่ตั้งฉากกับคอลัมน์กรานัมซึ่งแตกแขนงออกเป็นแผ่นดิสก์ขนานหลายแผ่นก่อตัวเป็นชุดกรานัม-สโตรมา[ 151 ]แบบจำลองเกลียวได้รับการสนับสนุนจากงานเพิ่มเติมอีกหลายชิ้น[ 149 ] [ 152 ]แต่ในที่สุดก็มีการกำหนดในปี 2019 ว่าคุณสมบัติจากทั้งแบบจำลองเกลียวและแบบจำลองการแตกแขนงนั้นรวมกันโดยจุดเชื่อมต่อเยื่อแบบเกลียวซ้ายที่เพิ่งค้นพบใหม่[ 146 ]เพื่อความสะดวก ระบบไทลาคอยด์จึงมักถูกแสดงด้วยแบบจำลอง "ฮับและซี่ล้อ" รุ่นเก่า โดยที่กรานาจะเชื่อมต่อกันด้วยท่อของไทลาคอยด์ในสโตรมา[ 153 ]
กรานาประกอบด้วยไทลาคอยด์กรานาลรูปวงกลมแบนเรียงซ้อนกันคล้ายแพนเค้ก แต่ละกรานาลสามารถมีไทลาคอยด์ได้ตั้งแต่สองถึงหนึ่งร้อยอัน[ 127 ]แม้ว่ากรานาลที่มีไทลาคอยด์ 10–20 อันจะพบได้บ่อยที่สุด[ 149 ]รอบๆ กรานาลจะมีไทลาคอยด์สโตรมัลแบบเกลียวขวาขนานกันหลายอัน หรือที่เรียกว่าเฟร็ตหรือไทลาคอยด์แบบแผ่น เกลียวเหล่านี้ทำมุมประมาณ 20° เชื่อมต่อกับไทลาคอยด์กรานาลแต่ละอันที่จุดเชื่อมต่อแบบช่องแคบคล้ายสะพาน[ 149 ] [ 152 ] [ 146 ]
แผ่นสโตรมาลามิลลาแผ่ขยายออกเป็นแผ่นขนาดใหญ่ตั้งฉากกับคอลัมน์กรานา แผ่นเหล่านี้เชื่อมต่อกับเกลียวขวาโดยตรงหรือผ่านการแตกแขนงที่ก่อให้เกิดพื้นผิวเมมเบรนเกลียวซ้าย[ 146 ]พื้นผิวเกลียวซ้ายมีมุมเอียงคล้ายกับเกลียวขวา (~20°) แต่มีระยะห่างเพียง ¼ โดยประมาณจะมีจุดเชื่อมต่อเกลียวซ้าย 4 จุดต่อกรานัม ส่งผลให้เกิดอาร์เรย์ของพื้นผิวเมมเบรนเกลียวขวาและซ้ายที่มีรัศมีและระยะห่างต่างกันซึ่งสมดุลระยะห่าง ทำให้เครือข่ายมีความแข็งแกร่งด้วยพลังงานพื้นผิวและการดัดงอน้อยที่สุด[ 146 ]แม้ว่าส่วนต่างๆ ของระบบไทลาคอยด์จะมีโปรตีนเมมเบรนที่แตกต่างกัน แต่เมมเบรนไทลาคอยด์นั้นต่อเนื่องกัน และช่องว่างไทลาคอยด์ที่ล้อมรอบนั้นก่อให้เกิดเขาวงกตต่อเนื่องเพียงแห่งเดียว[ 149 ]
องค์ประกอบของไทลาคอยด์
ภายในเยื่อไทลาคอยด์มีโปรตีนคอมเพล็กซ์ ที่สำคัญ ซึ่งทำหน้าที่เกี่ยวกับปฏิกิริยาแสงของการสังเคราะห์ แสง โฟโตซิสเต็ม IIและโฟโตซิสเต็ม Iประกอบด้วยคอมเพล็กซ์เก็บเกี่ยวแสงที่มีคลอโรฟิลล์และแคโรทีน อยด์ ซึ่งดูดซับพลังงานแสงและใช้พลังงานนั้นในการเพิ่มพลังงานให้กับอิเล็กตรอน โมเลกุลในเยื่อไทลาคอยด์ใช้อิเล็กตรอนที่มีพลังงานเพื่อปั๊มไอออนไฮโดรเจน เข้าไปในช่องว่างไทลาคอยด์ ทำให้ ค่า pHลดลงและกลายเป็นกรดเอนไซม์ ATP synthaseเป็นโปรตีนคอมเพล็กซ์ขนาดใหญ่ที่ใช้ประโยชน์จากความเข้มข้นของไอออนไฮโดรเจนในช่องว่างไทลาคอยด์เพื่อสร้าง พลังงาน ATPเมื่อไอออนไฮโดรเจนไหลกลับออกไปยังสโตรมา คล้ายกับกังหันน้ำ[ 120 ]
ไทลาคอยด์มีสองประเภท ได้แก่ ไทลาคอยด์กรานัล ซึ่งเรียงตัวกันเป็นกรานา และไทลาคอยด์สโตรมัล ซึ่งสัมผัสกับสโตรมาไทลาคอยด์กรานัลมีลักษณะเป็นแผ่นกลมแบนคล้ายแพนเค้ก มีเส้นผ่านศูนย์กลางประมาณ 300–600 นาโนเมตร ส่วนไทลาคอยด์สโตรมัล มีลักษณะ เป็นแผ่นเกลียว ที่พันรอบกรานา [ 149 ]ส่วนบนและล่างที่แบนราบของไทลาคอยด์กรานัลประกอบด้วย โปรตีนคอมเพล็กซ์ โฟโตซิสเต็ม II ที่ค่อนข้างแบนราบเท่านั้น ทำให้สามารถเรียงซ้อนกันได้อย่างแน่นหนา เกิดเป็นกรานาที่มีเยื่อหุ้มหลายชั้นที่แนบชิดกันอย่างแน่นหนา เรียกว่าเยื่อหุ้มกรานัล ซึ่งช่วยเพิ่มความเสถียรและพื้นที่ผิวสำหรับการจับแสง[ 149 ]
ในทางตรงกันข้ามโฟโตซิสเต็ม IและATP ซินเทสเป็นโปรตีนเชิงซ้อนขนาดใหญ่ที่ยื่นออกมาในสโตรมา พวกมันไม่สามารถเข้าไปอยู่ในเยื่อกรานัลที่แนบชิดกันได้ ดังนั้นจึงพบอยู่ในเยื่อไทลาคอยด์ของสโตรมา—ขอบของแผ่นไทลาคอยด์กรานัลและไทลาคอยด์ของสโตรมา โปรตีนเชิงซ้อนขนาดใหญ่เหล่านี้อาจทำหน้าที่เป็นตัวคั่นระหว่างแผ่นไทลาคอยด์ของสโตรมา[ 149 ]
จำนวนไทลาคอยด์และพื้นที่ไทลาคอยด์ทั้งหมดของคลอโรพลาสต์ได้รับอิทธิพลจากการได้รับแสง คลอโรพลาสต์ที่อยู่ในที่ร่มจะมีกรานา ขนาดใหญ่และจำนวนก รานามากกว่า และมีพื้นที่เยื่อไทลาคอยด์มากกว่าคลอโรพลาสต์ที่ได้รับแสงสว่าง ซึ่งจะมีกรานาขนาดเล็กและจำนวนกรานาน้อยกว่า และมีพื้นที่ไทลาคอยด์น้อยกว่า ขอบเขตของไทลาคอยด์สามารถเปลี่ยนแปลงได้ภายในไม่กี่นาทีหลังจากได้รับแสงหรือนำแสงออก[ 130 ]
เม็ดสีและสีของคลอโรพลาสต์
ภายในระบบสังเคราะห์แสงที่ฝังอยู่ในเยื่อไทลาคอยด์ของคลอโรพลาสต์ มีรงควัตถุสังเคราะห์ แสงหลายชนิด ซึ่งทำหน้าที่ดูดซับและถ่ายโอนพลังงานแสงชนิดของรงควัตถุที่พบนั้นแตกต่างกันในคลอโรพลาสต์แต่ละกลุ่ม และเป็นสาเหตุของสีสันที่หลากหลายในคลอโรพลาสต์ พลาสติดชนิดอื่นๆ เช่นลิวโคพลาสต์และโครโมพลาสต์มีคลอโรฟิลล์น้อยและไม่สามารถสังเคราะห์แสงได้

คลอโรฟิลล์
คลอโรฟิลล์เอพบได้ในคลอโรพลาสต์ทั้งหมด รวมถึงบรรพบุรุษ ของ ไซยาโนแบคทีเรีย ด้วย คลอโรฟิลล์ เอเป็นรงควัตถุสีเขียวอมฟ้า[ 154 ] ซึ่งมีส่วนทำให้ไซยาโนแบคทีเรียและคลอโรพลาสต์ส่วนใหญ่มีสี นอกจาก นี้ยังมีคลอโรฟิลล์ในรูปแบบอื่นๆ เช่น รงควัตถุเสริมคลอโรฟิลล์บีคลอโรฟิลล์ซีคลอโรฟิลล์ดี [ 12 ]และคลอโรฟิลล์เอฟ
คลอโรฟิลล์บีเป็น รงควัตถุ สีเขียวมะกอกที่พบเฉพาะในคลอโรพลาสต์ของพืชสาหร่ายสีเขียวคลอโรพลาสต์รองใดๆ ที่ได้มาจากการเกิดเอนโดซิมไบโอซิสรอง ของสาหร่ายสีเขียว และ ไซยาโนแบคทีเรียบางชนิด[ 12 ]คลอโรฟิลล์เอและบีรวมกันทำให้คลอโรพลาสต์ของพืชและสาหร่ายสีเขียวส่วนใหญ่มีสีเขียว[ 154 ]
คลอโรฟิลล์ซีส่วนใหญ่พบในคลอโรพลาสต์เอนโดซิมไบโอติกทุติยภูมิที่มาจากสาหร่ายแดงแม้ว่าจะไม่พบในคลอโรพลาสต์ของสาหร่ายแดงเองก็ตาม คลอโรฟิลล์ซียังพบในสาหร่ายสีเขียวและไซยาโนแบคทีเรีย บางชนิด ด้วย[ 12 ]
คลอโรฟิลล์dและfเป็นรงควัตถุที่พบเฉพาะในไซยาโนแบคทีเรียบางชนิดเท่านั้น[ 12 ] [ 155 ]
แคโรทีนอยด์
![Delesseria sanguinea ซึ่งเป็นสาหร่ายสีแดง มีคลอโรพลาสต์ที่มีเม็ดสีสีแดง เช่น ไฟโคเอริเทอริน ซึ่งปกปิดคลอโรฟิลล์เอสีเขียวอมฟ้า[39]](https://upload.wikimedia.org/wikipedia/commons/thumb/1/19/Delesseria_sanguinea_Helgoland.JPG/250px-Delesseria_sanguinea_Helgoland.JPG)
นอกจากคลอโรฟิลล์แล้ว ยังมี เม็ดสีสีเหลืองส้ม อีกกลุ่มหนึ่ง [ 154 ] ที่เรียกว่า แคโรทีนอยด์อยู่ในระบบสังเคราะห์แสงด้วย มีแคโรทีนอยด์ที่เกี่ยวข้องกับการสังเคราะห์แสงประมาณสามสิบชนิด[ 156 ]พวกมันช่วยในการถ่ายโอนและกระจายพลังงานส่วนเกิน[ 12 ]และสีที่สดใสของพวกมันบางครั้งก็กลบสีเขียวของคลอโรฟิลล์ เช่น ในช่วงฤดูใบไม้ร่วงเมื่อใบของพืชบกบางชนิดเปลี่ยนสี[ 157 ]เบต้าแคโรทีนเป็นแคโรทีนอยด์สีแดงส้มสดใสที่พบในคลอโรพลาสต์เกือบทั้งหมด เช่นเดียวกับคลอโรฟิลล์เอ[ 12 ] แซ นโทฟิลล์ โดยเฉพาะ ซีแซนทีนสีส้มแดงก็พบได้ทั่วไปเช่นกัน[ 156 ] นอกจาก นี้ ยัง มีแคโรทีนอยด์ในรูปแบบอื่นๆ อีกมากมายที่พบเฉพาะในคลอโรพลาสต์บางกลุ่มเท่านั้น[ 12 ]
ไฟโคบิลินส์
ไฟโคบิลินเป็นเม็ดสีกลุ่มที่สามที่พบในไซยาโนแบคทีเรีย กล อโคไฟต์สาหร่ายแดงและคลอโรพลาสต์ของ คริ ปโทไฟต์[ 12 ] [ 158 ]ไฟโคบิลินมีทุกสี แม้ว่าไฟโคเอริเทอรินจะเป็นหนึ่งในเม็ดสีที่ทำให้สาหร่ายแดงหลายชนิดมีสีแดง[ 159 ]ไฟโคบิลินมักจะรวมตัวกันเป็นโปรตีนเชิงซ้อนขนาดค่อนข้างใหญ่ประมาณ 40 นาโนเมตร เรียกว่าไฟ โค บิ ลิโซม [ 12 ]เช่นเดียวกับโฟโตซิสเต็ม IและATP ซินเทสไฟโคบิลิโซมจะยื่นเข้าไปในสโตรมา ป้องกันการเรียงซ้อนของไทลาคอยด์ในคลอโรพลาสต์ของสาหร่ายแดง[ 12 ] คลอโรพลาสต์ ของคริปโท ไฟต์ และไซยาโนแบคทีเรียบางชนิดไม่มีเม็ดสีไฟโคบิลินรวมตัวกันเป็นไฟโคบิลิโซม แต่เก็บไว้ในช่องว่างไทลาคอยด์แทน[ 12 ]
| รงควัตถุสังเคราะห์แสง การมีอยู่ของรงควัตถุในกลุ่มคลอโรพลาสต์และไซยาโนแบคทีเรีย เซลล์ที่มีสีแสดงถึงการมีอยู่ของเม็ดสี Chl = คลอโรฟิลล์[ 12 ] [ 156 ] [ 158 ] | |||||||||
| คลอโรฟิลล์ เอ | คลอร์ บี | คลอร์ ซี | คลอโร ฟิลล์ ดีและเอฟ | แซนโทฟิลล์ | อัลฟา-แคโรทีน | เบต้าแคโรทีน | ไฟโคบิลินส์ | ||
| พืชบก | |||||||||
| สาหร่ายสีเขียว | |||||||||
| ยูเกลโนไฟต์และคลอราแรคนิโอไฟต์ | |||||||||
| สาหร่ายแดงหลายเซลล์ | |||||||||
| สาหร่ายแดงเซลล์เดียว | |||||||||
| แฮปโทไฟต์และไดโนไฟต์ | |||||||||
| คริปโตไฟต์ | |||||||||
| กลอโคไฟต์ | |||||||||
| ไซยาโนแบคทีเรีย | |||||||||
คลอโรพลาสต์ชนิดพิเศษในพืชC4
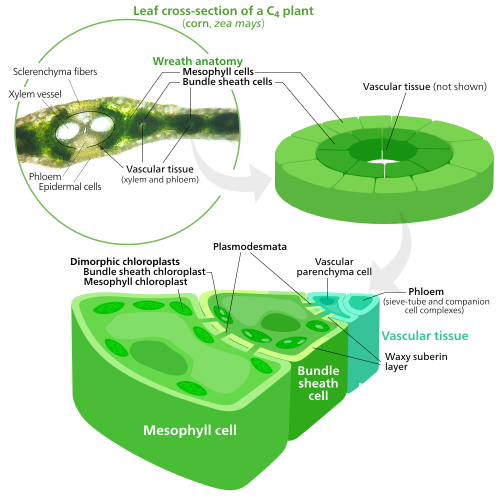
เพื่อตรึงคาร์บอนไดออกไซด์ให้กลายเป็นโมเลกุลน้ำตาลในกระบวนการสังเคราะห์แสงคลอโรพลาสต์ใช้เอนไซม์ที่เรียกว่าRuBisCO RuBisCO มีปัญหาในการแยกแยะระหว่างคาร์บอนไดออกไซด์และออกซิเจนดังนั้นที่ความเข้มข้นของออกซิเจนสูง RuBisCO จะเริ่มเติมออกซิเจนลงในสารตั้งต้นของน้ำตาลโดยไม่ได้ตั้งใจ ซึ่งส่งผลให้ พลังงาน ATPสูญเปล่าและ CO2 ถูกปล่อยออกมา โดยไม่มีการผลิตน้ำตาล นี่เป็นปัญหาใหญ่ เนื่องจาก O2 ถูกผลิตขึ้นจากปฏิกิริยาแสง เริ่มต้น ของการสังเคราะห์แสง ทำให้เกิดปัญหาในวัฏจักรแคลวินซึ่งใช้ RuBisCO [ 160 ]
พืชC4พัฒนาวิธีการแก้ปัญหานี้โดยการแยกปฏิกิริยาแสงและวัฏจักรแคลวินออกจากกันในเชิงพื้นที่ ปฏิกิริยาแสงซึ่งเก็บพลังงานแสงไว้ในATPและNADPHเกิดขึ้นใน เซลล์ มีโซฟิลล์ของใบพืช C4 ส่วนวัฏจักรแคลวินซึ่งใช้พลังงานที่เก็บไว้เพื่อสร้างน้ำตาลโดยใช้ RuBisCO เกิดขึ้นในเซลล์หุ้มมัดซึ่งเป็นชั้นของเซลล์ที่ล้อมรอบเส้นใบในใบ[ 160 ]
ด้วยเหตุนี้ คลอโรพลาสต์ในเซลล์มีโซฟิลล์ C4 และ เซลล์ปลอกมัด จึงมีความเชี่ยวชาญเฉพาะด้านสำหรับแต่ละขั้นตอนของการสังเคราะห์แสง ในเซลล์มีโซฟิลล์ คลอโรพลาสต์มีความเชี่ยวชาญเฉพาะด้านปฏิกิริยาแสง ดังนั้นจึงไม่มีRuBisCOและมีกรานาและไทลาคอยด์ ปกติ [ 141 ]ซึ่งใช้ในการสร้าง ATP และ NADPH รวมถึงออกซิเจน พวกมันเก็บ CO2 ไว้ใน สารประกอบ สี่คาร์บอน ซึ่งเป็นเหตุผลว่าทำไมกระบวนการนี้จึงเรียกว่าการสังเคราะห์แสงC4สารประกอบสี่คาร์บอนจะถูกขนส่งไปยังคลอโรพลาสต์ปลอกมัด ซึ่งจะปล่อย CO2 ออกมาและกลับไปยังมีโซฟิลล์ คลอโรพลาสต์ปลอกมัดไม่ดำเนินการปฏิกิริยาแสง ป้องกันไม่ให้ออกซิเจนสะสมในนั้นและรบกวนกิจกรรมของ RuBisCO [ 160 ]ด้วยเหตุนี้ พวกมันจึงขาดไทลาคอยด์ที่จัดเรียงเป็น กลุ่ม กรานา —ถึงแม้ว่าคลอโรพลาสต์ปลอกมัดจะยังมีไทลาคอยด์ลอยตัวอิสระอยู่ในสโตรมา ซึ่งพวกมันยังคงดำเนินการไหลเวียนของอิเล็กตรอนแบบวงจร ซึ่งเป็นวิธีการสังเคราะห์ ATPที่ขับเคลื่อนด้วยแสงเพื่อขับเคลื่อนวัฏจักรแคลวินโดยไม่สร้างออกซิเจน พวกมันขาดโฟโตซิสเต็ม IIและมีเพียงโฟโตซิสเต็ม I เท่านั้น —ซึ่งเป็นโปรตีนเชิงซ้อนเพียงชนิดเดียวที่จำเป็นสำหรับการไหลเวียนของอิเล็กตรอนแบบวงจร[ 141 ] [ 160 ]เนื่องจากหน้าที่ของคลอโรพลาสต์ปลอกมัดคือการดำเนินวัฏจักรแคลวินและสร้างน้ำตาล พวกมันจึงมักมีเม็ดแป้ง ขนาดใหญ่ [ 141 ]
คลอโรพลาสต์ทั้งสองชนิดมีเรติคูลัมรอบนอกของคลอโรพลาสต์จำนวนมาก [ 141 ] ซึ่งใช้เพื่อเพิ่มพื้นที่ผิวในการขนส่งสารเข้าและออกจาก คลอโรพลาสต์ [ 129 ] [ 130 ]คลอโรพลาสต์ในเนื้อเยื่อมีโซฟิลล์มีเรติคูลัมรอบนอกมากกว่าคลอโรพลาสต์ในปลอกมัดเล็กน้อย[ 161 ]
หน้าที่และเคมี
คลอโรพลาสต์ของเซลล์ยาม
ต่างจากเซลล์ผิวหนังส่วนใหญ่เซลล์ยามของปากใบ พืช มีคลอโรพลาสต์ที่พัฒนาค่อนข้างดี[ 162 ]อย่างไรก็ตาม หน้าที่ที่แท้จริงของพวกมันยังเป็นที่ถกเถียงกันอยู่[ 163 ]
ภูมิคุ้มกันโดยธรรมชาติของพืช
พืชขาดเซลล์ภูมิคุ้มกัน เฉพาะ —เซลล์พืชทั้งหมดมีส่วนร่วมในการตอบสนองภูมิคุ้มกัน ของพืช คลอโรพลาสต์ ร่วมกับนิวเคลียสเยื่อหุ้มเซลล์และเอนโดพลาสมิกเรติคูลัม [ 164 ] เป็นผู้เล่นหลักใน การป้องกัน เชื้อโรคเนื่องจากบทบาทของมันในการตอบสนองภูมิคุ้มกันของเซลล์พืช เชื้อโรคจึงมักโจมตีคลอโรพลาสต์[ 164 ]
พืชมีการตอบสนองทางภูมิคุ้มกันหลักสองแบบ ได้แก่การตอบสนองแบบไวเกิน (hypersensitive response ) ซึ่งเซลล์ที่ติดเชื้อจะปิดตัวเองและเกิดการตายของเซลล์ตามโปรแกรมและการต้านทานที่ได้รับมาแบบเป็นระบบ (systemic acquired resistance ) ซึ่งเซลล์ที่ติดเชื้อจะปล่อยสัญญาณเตือนส่วนที่เหลือของพืชถึงการมีอยู่ของเชื้อโรค คลอโรพลาสต์กระตุ้นการตอบสนองทั้งสองแบบโดยการทำลายระบบสังเคราะห์แสงโดยเจตนา ทำให้เกิดอนุมูลอิสระออกซิเจน (reactive oxygen species) ระดับอนุมูลอิสระออกซิเจนที่สูงจะทำให้เกิด การตอบสนองแบบไว เกิน อนุมูลอิสระออกซิเจนยังฆ่าเชื้อโรคภายในเซลล์โดยตรงด้วย ระดับอนุมูลอิสระออกซิเจนที่ต่ำลงจะเริ่มต้นการต้านทานที่ได้รับมาแบบเป็นระบบกระตุ้นการผลิตโมเลกุลป้องกันในส่วนที่เหลือของพืช[ 164 ]
ในพืชบางชนิด คลอโรพลาสต์จะเคลื่อนที่เข้าใกล้บริเวณที่เกิดการติดเชื้อและนิวเคลียส มากขึ้น ในระหว่างการติดเชื้อ[ 164 ]
คลอโรพลาสต์สามารถทำหน้าที่เป็นเซนเซอร์ของเซลล์ได้ หลังจากตรวจพบความเครียดในเซลล์ ซึ่งอาจเกิดจากเชื้อโรค คลอโรพลาสต์จะเริ่มผลิตโมเลกุลต่างๆ เช่นกรดซาลิไซลิกกรดจัสมอนิก ไนตริกออกไซด์และสารออกซิเจนที่ว่องไวซึ่งสามารถทำหน้าที่เป็นสัญญาณป้องกันได้ เนื่องจากสารออกซิเจนที่ว่องไวเป็นสัญญาณของเซลล์ จึงเป็นโมเลกุลที่ไม่เสถียร ดังนั้นจึงไม่น่าจะออกจากคลอโรพลาสต์ แต่จะส่งสัญญาณต่อไปยังโมเลกุลตัวกลางที่ไม่ทราบชนิดแทน โมเลกุลเหล่านี้ทั้งหมดจะเริ่มต้นการส่งสัญญาณย้อนกลับซึ่งเป็นสัญญาณจากคลอโรพลาสต์ที่ควบคุมการแสดงออกของยีนในนิวเคลียส[ 164 ]
นอกจากการส่งสัญญาณป้องกันแล้ว คลอโรพลาสต์ยังช่วยสังเคราะห์โมเลกุลป้องกันที่สำคัญอย่างจัสมอเนตโดย อาศัย เพอร์ออกซิโซม[ 165 ]คลอโรพลาสต์สังเคราะห์กรดไขมัน ทั้งหมด ในเซลล์พืช[ 164 ] [ 166 ] — กรดลิโนเลอิกซึ่งเป็นกรดไขมันชนิดหนึ่ง เป็นสารตั้งต้นของจัสมอเนต[ 164 ]
การสังเคราะห์แสง
หน้าที่หลักอย่างหนึ่งของคลอโรพลาสต์คือบทบาทในการสังเคราะห์แสง ซึ่งเป็นกระบวนการที่แสงถูกเปลี่ยนเป็นพลังงานเคมี เพื่อผลิตอาหารในรูปของน้ำตาล ต่อ ไปน้ำ (H₂O )และคาร์บอนไดออกไซด์ (CO₂ )ถูกนำมาใช้ในการสังเคราะห์แสง และน้ำตาลและออกซิเจน (O₂ )ถูกสร้างขึ้นโดยใช้พลังงานแสงการสังเคราะห์แสงแบ่งออกเป็นสองขั้นตอน คือปฏิกิริยาแสงซึ่งน้ำถูกแยกออกเพื่อผลิตออกซิเจน และปฏิกิริยามืดหรือวัฏจักรแคลวินซึ่งสร้างโมเลกุลน้ำตาลจากคาร์บอนไดออกไซด์ ทั้งสองขั้นตอนเชื่อมโยงกันด้วยตัวนำพลังงานอะดีโนซีนไตรฟอสเฟต (ATP) และนิโคตินาไมด์อะดีนีนไดนิวคลีโอไทด์ฟอสเฟต ( NADP⁺ ) [ 167 ] [ 168 ]
ปฏิกิริยาแสง

ปฏิกิริยาแสงเกิดขึ้นบนเยื่อไทลาคอยด์ โดยจะดึงพลังงานแสงมาเก็บไว้ในรูป ของ NADPH , NADP + รูปแบบหนึ่ง และATPเพื่อใช้เป็นเชื้อเพลิงสำหรับปฏิกิริยา มืด
ตัวนำพลังงาน
ATP คือรูปแบบฟอสฟอริเลตของอะดีโนซีนไดฟอสเฟต (ADP) ซึ่งเก็บสะสมพลังงานในเซลล์และขับเคลื่อนกิจกรรมส่วนใหญ่ของเซลล์ ATP เป็นรูปแบบที่มีพลังงาน ในขณะที่ ADP เป็นรูปแบบที่ (บางส่วน) ถูกทำให้หมดพลังงานไปแล้ว NADP +คือตัวนำอิเล็กตรอนที่ลำเลียงอิเล็กตรอนพลังงานสูง ในปฏิกิริยาแสง มันจะถูกรีดิวซ์ ซึ่งหมายความว่ามันจะรับอิเล็กตรอนและกลายเป็นNADPH
การสังเคราะห์ด้วยแสง
เช่นเดียวกับไมโตคอนเดรีย คลอโรพลาสต์ใช้พลังงานศักยภาพที่เก็บไว้ในเก รเดียนต์ H + หรือไอออนไฮโดรเจนเพื่อสร้างพลังงาน ATP โฟโตซิสเต็มทั้งสองจับพลังงานแสงเพื่อเพิ่มพลังงานให้กับอิเล็กตรอน ที่ นำมาจากน้ำและปล่อยอิเล็กตรอนเหล่านั้นลงไปตามห่วงโซ่การขนส่งอิเล็กตรอนโมเลกุลระหว่างโฟโตซิสเต็มจะควบคุมพลังงานของอิเล็กตรอนเพื่อปั๊มไอออนไฮโดรเจนเข้าไปในช่องว่างไทลาคอยด์ ทำให้เกิดเกรเดียนต์ความเข้มข้นโดยมีไอออนไฮโดรเจนมากกว่า (มากถึงหนึ่งพันเท่า) [ 120 ]ภายในระบบไทลาคอยด์มากกว่าในสโตรมา จากนั้นไอออนไฮโดรเจนในช่องว่างไทลาคอยด์จะแพร่ กลับลงมาตาม เกรเดียนต์ความเข้มข้น ไหลกลับออกไปสู่สโตรมาผ่าน ATP synthase ATP synthase ใช้พลังงานจากไอออนไฮโดรเจนที่ไหลเพื่อฟอสโฟรีเลตอะดีโนซีนไดฟอสเฟตเป็นอะดีโนซีนไตรฟอสเฟตหรือ ATP [ 120 ] [ 169 ]เนื่องจากคลอโรพลาสต์ ATP ซินเทสยื่นออกไปในสโตรมา ATP จึงถูกสังเคราะห์ที่นั่น ในตำแหน่งที่พร้อมใช้งานในปฏิกิริยามืด[ 170 ]
NADP +การลด
อิเล็กตรอนมักจะถูกนำออกจากห่วงโซ่การขนส่งอิเล็กตรอนเพื่อชาร์จNADP +ด้วยอิเล็กตรอนลด NADP+ ให้เป็นNADPHเช่นเดียวกับ ATP synthase เฟอร์เรดอกซิน-NADP +รีดักเทส ซึ่งเป็น เอนไซม์ที่ลด NADP +จะปล่อย NADPH ที่สร้างขึ้นเข้าสู่สโตรมา ซึ่งเป็นตำแหน่งที่จำเป็นสำหรับปฏิกิริยามืด[ 170 ]
เนื่องจากการลด NADP + จะกำจัดอิเล็กตรอนออกจากห่วงโซ่การขนส่งอิเล็กตรอน จึงต้องมีการแทนที่ อิเล็กตรอนเหล่านั้น ซึ่งเป็นหน้าที่ของโฟโตซิสเต็ม IIที่แยกโมเลกุลน้ำ ( H2O ) เพื่อรับอิเล็กตรอนจากอะตอมไฮโดรเจน [ 120 ] [ 167 ]
การสังเคราะห์ฟอสเฟตด้วยแสงแบบวัฏจักร
ในขณะที่โฟโตซิสเต็ม II สลายน้ำด้วยแสงเพื่อรับและเพิ่มพลังงานให้กับอิเล็กตรอนใหม่โฟโตซิสเต็ม Iเพียงแค่เพิ่มพลังงานให้กับอิเล็กตรอนที่หมดไปที่ปลายสุดของห่วงโซ่การขนส่งอิเล็กตรอน โดยปกติแล้ว อิเล็กตรอนที่ได้รับพลังงานใหม่จะถูกนำโดย NADP +แต่บางครั้งพวกมันอาจไหลกลับลงมาตามห่วงโซ่การขนส่งอิเล็กตรอนที่ปั๊ม H + มากขึ้นเพื่อ ขนส่ง ไอออนไฮโดรเจนเพิ่มเติมเข้าไปในช่องว่างไทลาคอยด์เพื่อสร้าง ATP เพิ่มขึ้น กระบวนการนี้เรียกว่าการสังเคราะห์ฟอสเฟตแบบวงจรเนื่องจากอิเล็กตรอนถูกนำกลับมาใช้ใหม่ การสังเคราะห์ฟอสเฟตแบบวงจรพบได้ทั่วไปในพืชC4ซึ่งต้องการATPมากกว่าNADPH [ 160 ]
ปฏิกิริยาด้านมืด

วัฏจักรแคลวินหรือที่รู้จักกันในชื่อปฏิกิริยามืดเป็นชุดของปฏิกิริยาชีวเคมีที่ตรึงCO2 ลงใน โมเลกุลน้ำตาล G3Pและใช้พลังงานและอิเล็กตรอนจากATPและNADPHที่สร้างขึ้นในปฏิกิริยาแสง วัฏจักรแคลวินเกิดขึ้นในสโตรมาของคลอโรพลาสต์[ 160 ]
แม้ว่าจะเรียกว่า"ปฏิกิริยามืด"แต่ในพืชส่วนใหญ่ ปฏิกิริยาเหล่านี้เกิดขึ้นในที่สว่าง เนื่องจากปฏิกิริยามืดขึ้นอยู่กับผลิตภัณฑ์ของปฏิกิริยาสว่าง[ 11 ]
การตรึงคาร์บอนและการสังเคราะห์ G3P
วัฏจักรแคลวินเริ่มต้นด้วยการใช้เอนไซม์RuBisCOเพื่อตรึง CO2 ให้ เป็นโมเลกุล Ribulose bisphosphate (RuBP) ที่มีคาร์บอน 5 อะตอม ผลลัพธ์ที่ได้คือโมเลกุลที่มีคาร์บอน 6 อะตอมที่ไม่เสถียร ซึ่งจะสลายตัวทันทีเป็นโมเลกุลที่มีคาร์บอน 3 อะตอม เรียกว่า 3-phosphoglyceric acidหรือ 3-PGA ATPและNADPHที่สร้างขึ้นในปฏิกิริยาแสงจะถูกนำไปใช้ในการเปลี่ยน 3-PGA ให้เป็น โมเลกุลน้ำตาล glyceraldehyde-3-phosphateหรือ G3P โมเลกุล G3P ส่วนใหญ่จะถูกนำกลับมาใช้ใหม่เป็น RuBP โดยใช้พลังงานจาก ATP เพิ่มเติม แต่หนึ่งในหกของโมเลกุลที่ผลิตได้จะออกจากวัฏจักร ซึ่งเป็นผลิตภัณฑ์สุดท้ายของปฏิกิริยามืด[ 160 ]
น้ำตาลและแป้ง
กลีเซอรัลดีไฮด์-3-ฟอสเฟตสามารถรวมตัวกันเพื่อสร้างโมเลกุลน้ำตาลขนาดใหญ่ขึ้น เช่นกลูโคสและฟรุกโตส โมเลกุลเหล่านี้จะถูกประมวลผล และจากนั้น จะได้ ซูโครสซึ่งเป็นไดแซ็กคาไรด์ที่รู้จักกันทั่วไปในชื่อน้ำตาลทราย แม้ว่ากระบวนการนี้จะเกิดขึ้นนอกคลอโรพลาสต์ในไซโตพลาสซึมก็ตาม[ 171 ]
อีกทางเลือก หนึ่ง โมโนเมอร์ของกลูโคสในคลอโรพลาสต์สามารถเชื่อมต่อกันเพื่อสร้างแป้งซึ่งจะสะสมเป็นเม็ดแป้งที่พบในคลอโรพลาสต์[ 171 ] ภายใต้สภาวะเช่นความเข้มข้นของ CO2 ในบรรยากาศสูงเม็ดแป้งเหล่านี้อาจเติบโตจนมีขนาดใหญ่มาก ทำให้กรานาและไทลาคอยด์ผิดรูป เม็ดแป้งจะเข้ามาแทนที่ไทลาคอยด์ แต่ยังคงรักษาสภาพของไทลาคอยด์ไว้[ 172 ]ราก ที่แช่น้ำยังสามารถทำให้เกิด การสะสม ของแป้งในคลอโรพลาสต์ ซึ่งอาจเป็นเพราะซูโครสถูกส่งออกจากคลอโรพลาสต์ (หรือที่แม่นยำกว่านั้นคือเซลล์พืช ) น้อยลง ซึ่งจะทำให้ปริมาณ ฟอสเฟตอิสระของพืชลดลงซึ่งกระตุ้นการสังเคราะห์แป้งในคลอโรพลาสต์ทางอ้อม[ 172 ] แม้ว่าจะเชื่อมโยงกับอัตราการสังเคราะห์แสงที่ต่ำ แต่เม็ดแป้งเองอาจไม่จำเป็นต้องรบกวนประสิทธิภาพของการสังเคราะห์แสงอย่างมีนัยสำคัญ[ 173 ]และอาจเป็นเพียงผลข้างเคียงของปัจจัยอื่นที่ทำให้การสังเคราะห์แสงลดลง[ 172 ]
การหายใจระดับแสง
การหายใจระดับแสงสามารถเกิดขึ้นได้เมื่อความเข้มข้นของออกซิเจนสูงเกินไป RuBisCO ไม่สามารถแยกแยะระหว่างออกซิเจนและคาร์บอนไดออกไซด์ได้ดีนัก ดังนั้นจึงอาจเติม O2 แทนที่จะเป็น CO2 ลงใน RuBP โดยไม่ได้ตั้งใจกระบวนการนี้ลดประสิทธิภาพของการสังเคราะห์แสง—มันใช้ ATP และออกซิเจน ปล่อย CO2 ออกมาและไม่สร้างน้ำตาล มันอาจทำให้คาร์บอนที่ถูกตรึงโดยวัฏจักรแคลวินสูญเปล่าได้ถึงครึ่งหนึ่ง[ 167 ]กลไกหลายอย่างได้วิวัฒนาการในสายพันธุ์ต่างๆ ที่เพิ่มความเข้มข้นของคาร์บอนไดออกไซด์เมื่อเทียบกับออกซิเจนภายในคลอโรพลาสต์ ซึ่งเพิ่มประสิทธิภาพของการสังเคราะห์แสง กลไกเหล่านี้เรียกว่ากลไกการเพิ่มความเข้มข้นของคาร์บอนไดออกไซด์หรือ CCMs ซึ่งรวมถึงการเผาผลาญกรด Crassulacean การตรึงคาร์บอนC4 [ 167 ]และไพรีนอย ด์ คลอโรพลาส ต์ในพืช C4 มี ลักษณะเด่นคือแสดงให้เห็นถึง ภาวะสองรูปแบบของคลอโรพลาสต์ที่แตก ต่างกัน
ค่า pH
เนื่องจาก ความแตกต่างของความเข้มข้น ของ H +ข้ามเยื่อไทลาคอยด์ ภายในไทลาคอยด์จึงมีสภาพเป็นกรดโดยมีค่า pHประมาณ 4 [ 174 ]ในขณะที่สโตรมามีสภาพเป็นเบสเล็กน้อย โดยมีค่า pH ประมาณ 8 [ 175 ] ค่า pH ของสโตรมาที่เหมาะสมที่สุดสำหรับวัฏจักรแคลวินคือ 8.1 โดยปฏิกิริยาจะหยุดลงเกือบหมดเมื่อค่า pH ลดลงต่ำกว่า 7.3 [ 176 ]
CO2ในน้ำสามารถก่อตัวเป็นกรดคาร์บอนิก ซึ่งอาจรบกวนค่า pH ของคลอโรพลาสต์ที่แยกออกมา ทำให้การสังเคราะห์แสงถูก รบกวนแม้ว่า CO2 จะถูกใช้ในการสังเคราะห์แสงก็ตาม อย่างไรก็ตาม คลอโรพลาสต์ในเซลล์พืช ที่มีชีวิต จะไม่ได้รับผลกระทบจากสิ่งนี้มากนัก[ 175 ]
คลอโรพลาสต์สามารถสูบ ไอออน K +และ H +เข้าและออกจากตัวเองโดยใช้ระบบขนส่งที่ขับเคลื่อนด้วยแสงซึ่งยังไม่เป็นที่เข้าใจดีนัก[ 175 ]
เมื่อมีแสง ค่า pH ของลูเมนไทลาคอยด์อาจลดลงได้ถึง 1.5 หน่วย pH ในขณะที่ค่า pH ของสโตรมาอาจเพิ่มขึ้นได้เกือบ 1 หน่วย pH [ 176 ]
การสังเคราะห์กรดอะมิโน
คลอโรพลาสต์เพียงอย่างเดียวสร้าง กรดอะมิโนเกือบทั้งหมดของเซลล์พืชในสโตรมา[ 177 ]ยกเว้นกรดอะมิโนที่มีกำมะถันเช่นซิสเทอีนและเมไทโอนีน [ 178 ] [ 179 ] ซิสเทอีนถูกสร้างขึ้นในคลอโรพลาสต์ ( รวมถึง โปรพลาสติดด้วย) แต่ก็มีการสังเคราะห์ในไซโตซอลและไมโทคอนเดรีย ด้วย อาจเป็นเพราะมันมีปัญหาในการข้ามเยื่อหุ้มเซลล์ไปยังตำแหน่งที่ต้องการ[ 179 ]เป็นที่ทราบกันว่าคลอโรพลาสต์สร้างสารตั้งต้นของเมไทโอนีน แต่ยังไม่ชัดเจนว่าออร์แกเนลล์นี้ดำเนินการขั้นตอนสุดท้ายของกระบวนการหรือเกิดขึ้นในไซโตซอล[ 180 ]
สารประกอบไนโตรเจนอื่นๆ
คลอโรพลาสต์สร้าง พิวรีนและไพริมิดีนทั้งหมดของเซลล์ซึ่งเป็นเบสไนโตรเจน ที่ พบในDNAและRNA [ 177 ]นอกจากนี้ยังเปลี่ยนไนไตรต์ (NO 2 − ) ให้เป็นแอมโมเนีย (NH 3 ) ซึ่งให้ ไนโตรเจนแก่พืชเพื่อสร้างกรดอะมิโนและนิวคลีโอไทด์[ 177 ]
ผลิตภัณฑ์เคมีอื่นๆ
พลาสติดเป็นแหล่ง สังเคราะห์ ไขมัน ที่หลากหลายและซับซ้อน ในพืช[ 181 ] [ 182 ]คาร์บอนที่ใช้ในการสร้างไขมันส่วนใหญ่มาจากอะเซทิล-โคเอ ซึ่งเป็นผลิตภัณฑ์จากการดีคาร์บอกซิเล ชันของไพรูเวต[ 181 ]ไพรูเวตอาจเข้าสู่พลาสติดจากไซโตซอลโดยการแพร่แบบพาสซีฟผ่านเยื่อหุ้มเซลล์หลังจากการผลิตในกระบวนการไกลโคไลซิส [ 183 ] ไพรูเวตยังถูกสร้างขึ้นในพลาสติดจากฟอสโฟอีโนลไพรูเวต ซึ่งเป็นเมตาบอไลต์ที่สร้างขึ้นในไซโตซอลจากไพรูเวตหรือPGA [ 181 ] อะซิเตตในไซโตซอลไม่สามารถนำมาใช้ในการสังเคราะห์ไขมันในพลาสติดได้[ 184 ]ความยาวทั่วไปของกรดไขมันที่ผลิตในพลาสติดคือ 16 หรือ 18 คาร์บอน โดยมีพันธะคู่ซิส 0-3 พันธะ [ 185 ]
การสังเคราะห์กรดไขมันจากอะเซทิล-โคเอส่วนใหญ่ต้องอาศัยเอนไซม์สองชนิด อะเซทิล-โคเอ คาร์บอกซิเลสสร้างมาโลนิล-โคเอ ซึ่งใช้ทั้งในขั้นตอนแรกและขั้นตอนการขยายของการสังเคราะห์ แฟตตี้แอซิดซินเทส (FAS) เป็นเอนไซม์และโคแฟคเตอร์ที่ซับซ้อนขนาดใหญ่ รวมถึงโปรตีนตัวพาอะซิล (ACP) ซึ่งยึดโซ่อะซิลไว้ในขณะที่สังเคราะห์ การเริ่มต้นของการสังเคราะห์เริ่มจากการควบแน่นของมาโลนิล-ACP กับอะเซทิล-โคเอเพื่อสร้างคีโตบิวทิริล-ACP การรีดิวซ์ 2 ครั้งโดยใช้NADPHและการกำจัดน้ำ 1 ครั้งจะสร้างบิวทิริล-ACP การขยายกรดไขมันมาจากการควบแน่น การรีดิวซ์ และการกำจัดน้ำของมาโลนิล-ACP ซ้ำๆ[ 181 ]
ลิปิดอื่นๆ ได้มาจากวิถีเมทิล-อีริทริทอลฟอสเฟต (MEP)และประกอบด้วยจิบเบอเรลินสเตอรอล กรดแอ็บซิสิก ไฟทอลและเมตาโบไลต์รอง จำนวนนับ ไม่ ถ้วน [ 181 ]
ที่ตั้ง

การกระจายตัวภายในพืช
ไม่ใช่ทุกเซลล์ในพืชหลายเซลล์จะมีคลอโรพลาสต์ ส่วนสีเขียวทั้งหมดของพืชจะมีคลอโรพลาสต์เนื่องจากสีเขียวมาจากคลอโรฟิลล์ [ 11 ] เซลล์พืชที่มีคลอโรพลาสต์มักจะเป็น เซลล์ พาเรนไคมาแม้ว่าคลอโรพลาสต์จะพบได้ในเนื้อเยื่อคอลเลนไคมา ด้วยก็ตาม [ 186 ]เซลล์พืชที่มีคลอโรพลาสต์เรียกว่า เซลล์ คลอเรนไคมา เซลล์ คลอเรนไคมาทั่วไปของพืชบกจะมีคลอโรพลาสต์ประมาณ 10 ถึง 100 เซลล์
ในพืชบางชนิด เช่นกระบองเพชรคลอโรพลาสต์จะพบได้ในลำต้น [ 187 ]แม้ว่าในพืชส่วนใหญ่ คลอโรพลาสต์จะกระจุกตัวอยู่ในใบเนื้อเยื่อใบหนึ่งตารางมิลลิเมตร สามารถมีคลอโรพลาสต์ได้ถึงครึ่งล้าน [ 11 ]ภายในใบ คลอโรพลาสต์ส่วนใหญ่พบใน ชั้น มีโซฟิลล์ของใบและเซลล์ยามของปากใบเซลล์พาลิเซดมีโซฟิลล์สามารถมีคลอโรพลาสต์ได้ 30–70 เซลล์ต่อเซลล์ ในขณะที่เซลล์ยามของปากใบมีเพียงประมาณ 8–15 เซลล์ต่อเซลล์ และมีคลอโรฟิลล์น้อยกว่ามากคลอโรพลาสต์ยังสามารถพบได้ใน เซลล์ หุ้มมัดท่อลำเลียงของใบ โดยเฉพาะอย่างยิ่งในพืชC4ซึ่งดำเนินวัฏจักรแคลวินในเซลล์หุ้มมัดท่อลำเลียง มักจะไม่มีอยู่ในชั้นเอพิเดอร์มิสของใบ[ 162 ]
ตำแหน่งเซลลูลาร์

การเคลื่อนที่ของคลอโรพลาสต์
คลอโรพลาสต์ของเซลล์พืชและสาหร่ายสามารถปรับทิศทางตัวเองให้เหมาะสมกับแสงที่มีอยู่ได้ดีที่สุด ในสภาวะแสงน้อย พวกมันจะแผ่ออกเป็นแผ่นเพื่อเพิ่มพื้นที่ผิวในการดูดซับแสงให้มากที่สุด ภายใต้แสงที่เข้มข้น พวกมันจะหาที่กำบังโดยการเรียงตัวเป็นแนวตั้งตามผนังเซลล์ ของเซลล์พืช หรือหันข้างเพื่อให้แสงตกกระทบด้านข้าง ซึ่งจะช่วยลดการสัมผัสและปกป้องพวกมันจากความเสียหายจากปฏิกิริยาออกซิเดชันจาก แสง [ 188 ]ความสามารถในการกระจายคลอโรพลาสต์เพื่อให้สามารถหลบอยู่ด้านหลังกันหรือแผ่ออกอาจเป็นเหตุผลที่พืชบกวิวัฒนาการให้มีคลอโรพลาสต์ขนาดเล็กจำนวนมากแทนที่จะมีคลอโรพลาสต์ขนาดใหญ่เพียงไม่กี่อัน[ 189 ] การเคลื่อนที่ของคลอโรพลาสต์ถือเป็นหนึ่งในระบบการตอบสนองต่อสิ่งเร้าที่มีการควบคุมอย่างใกล้ชิดที่สุดที่พบได้ในพืช[ 190 ] นอกจากนี้ยังพบว่า ไมโทคอนเดรียเคลื่อนที่ตามคลอโรพลาสต์ด้วย[ 191 ]
ในพืชชั้นสูง การเคลื่อนที่ของคลอโรพลาสต์ถูกควบคุมโดยโฟโตโทรปิน ซึ่ง เป็นตัวรับแสงสีฟ้า ที่รับผิดชอบต่อการตอบสนอง ต่อแสงของพืชในสาหร่ายบางชนิดมอสเฟิร์นและพืชดอกการเคลื่อนที่ของคลอโรพลาสต์ได้รับอิทธิพลจากแสงสีแดงนอกเหนือจากแสงสีฟ้า[ 188 ]แม้ว่าความยาวคลื่นสีแดงที่ยาวมากจะยับยั้งการเคลื่อนที่แทนที่จะเร่งให้เร็วขึ้น โดยทั่วไปแล้วแสงสีฟ้าจะทำให้คลอโรพลาสต์หาที่หลบภัย ในขณะที่แสงสีแดงจะดึงพวกมันออกมาเพื่อเพิ่มการดูดซับแสงให้สูงสุด[ 191 ]
การศึกษาเกี่ยวกับVallisneria giganteaซึ่งเป็นพืชน้ำที่มีดอกแสดงให้เห็นว่าคลอโรพลาสต์สามารถเคลื่อนที่ได้ภายในห้านาทีหลังจากได้รับแสง แม้ว่าในตอนแรกจะยังไม่แสดงทิศทางที่ชัดเจนก็ตาม พวกมันอาจเคลื่อนที่ไปตาม เส้นทางของ ไมโครฟิลาเมนต์และข้อเท็จจริงที่ว่าตาข่ายไมโครฟิลาเมนต์เปลี่ยนรูปร่างเป็นโครงสร้างรังผึ้งล้อมรอบคลอโรพลาสต์หลังจากที่พวกมันเคลื่อนที่แล้ว แสดงให้เห็นว่าไมโครฟิลาเมนต์อาจช่วยยึดคลอโรพลาสต์ให้อยู่กับที่[ 190 ] [ 191 ]
การจำแนกความแตกต่าง การจำลองแบบ และการถ่ายทอดทางพันธุกรรม

คลอโรพลาสต์เป็นออร์แกเนลล์ชนิดพิเศษของเซลล์พืชที่เรียกว่าพลาสติดแม้ว่าบางครั้งจะใช้สองคำนี้แทนกันได้ก็ตาม มีพลาสติดประเภทอื่นๆ อีกมากมายซึ่งทำหน้าที่ต่างๆ กัน คลอโรพลาสต์ทั้งหมดในพืชสืบเชื้อสายมาจากโปรพลาสติดที่ยังไม่แยกชนิดซึ่งพบในไซโกต [ 177 ] หรือไข่ ที่ ได้รับการปฏิสนธิ โปรพลาสติดมักพบใน เนื้อเยื่อเจริญปลายยอดของพืชที่โตเต็มวัย โดยปกติแล้วคลอโรพลาสต์จะไม่พัฒนามาจากโปรพลาสติดในเนื้อเยื่อเจริญปลายราก[ 192 ]แต่การก่อตัวของอะไมโลพลาสต์ ที่เก็บสะสมแป้งนั้น พบได้บ่อยกว่า[ 177 ]
ในยอดพืชโปรพลาสติดจากเนื้อเยื่อเจริญปลายยอดสามารถพัฒนาไปเป็นคลอโรพลาสต์ใน เนื้อเยื่อใบ ที่สังเคราะห์ แสงได้ เมื่อใบเจริญเติบโตเต็มที่ หากได้รับแสงที่เหมาะสม[ 15 ]กระบวนการนี้เกี่ยวข้องกับการยุบตัวของเยื่อหุ้มพลาสติดชั้นใน ทำให้เกิดแผ่นเยื่อหุ้มที่ยื่นเข้าไปในสโตรมา ภายใน จากนั้นแผ่นเยื่อหุ้มเหล่านี้จะพับตัวเพื่อสร้างไทลาคอยด์และกรานา[ 193 ]
หาก หน่อของพืช ดอกไม่ได้รับแสงที่จำเป็นสำหรับการสร้างคลอโรพลาสต์ โปรพลาสติดอาจพัฒนาไปเป็น ระยะ เอทิโอพลาสต์ก่อนที่จะกลายเป็นคลอโรพลาสต์ เอทิโอพลาสต์เป็นพลาสติดที่ไม่มี คลอ โรฟิลล์ และมีเยื่อหุ้มชั้นในที่เว้าเข้าไปด้านในซึ่งก่อตัวเป็นโครงตาข่ายของท่อในสโตรมา เรียกว่าโพรลาเมลลาร์ บอดี้แม้ว่าเอทิโอพลาสต์จะไม่มีคลอโรฟิลล์ แต่ก็มีสารตั้งต้น ของคลอโรฟิลล์สีเหลือง สะสม อยู่ [ 15 ]ภายในไม่กี่นาทีของการได้รับแสง โพรลาเมลลาร์ บอดี้จะเริ่มจัดระเบียบใหม่เป็นกองของไทลาคอยด์ และเริ่มผลิตคลอโรฟิลล์ กระบวนการนี้ซึ่งเอทิโอพลาสต์กลายเป็นคลอโรพลาสต์ใช้เวลาหลายชั่วโมง[ 193 ]พืชเมล็ดเปลือยไม่ต้องการแสงในการสร้างคลอโรพลาสต์[ 193 ]
อย่างไรก็ตาม แสงไม่ได้รับประกันว่าโปรพลาสติดจะพัฒนาเป็นคลอโรพลาสต์ การที่โปรพลาสติดจะพัฒนาเป็นคลอโรพลาสต์หรือพลาสติดชนิดอื่นนั้น ส่วนใหญ่จะถูกควบคุมโดยนิวเคลียส[ 15 ]และได้รับอิทธิพลอย่างมากจากชนิดของเซลล์ที่มันอาศัยอยู่[ 177 ]

การเปลี่ยนรูปพลาสติด
การเปลี่ยนแปลงของพลาสติดไม่ใช่สภาวะถาวร ในความเป็นจริงแล้ว การเปลี่ยนแปลงระหว่างพลาสติดหลายชนิดเป็นไปได้ คลอโรพลาสต์อาจถูกเปลี่ยนเป็นโครโมพลาสต์ซึ่งเป็น พลาสติดที่เต็มไปด้วยเม็ด สีที่ทำให้เกิดสีสันสดใสในดอกไม้และผลไม้ สุก อะไมโลพ ลาสต์ที่เก็บสะสมแป้งก็สามารถเปลี่ยนเป็นโครโมพลาสต์ได้เช่นกัน และโปรพลาสติดก็สามารถพัฒนาไปเป็นโครโมพลาสต์ได้โดยตรง โครโมพลาสต์และอะไมโลพลาสต์ยังสามารถกลายเป็นคลอโรพลาสต์ได้ เช่นเดียวกับที่เกิดขึ้นเมื่อแครอทหรือมันฝรั่งได้รับแสง หากพืชได้รับบาดเจ็บ หรือมีสิ่งอื่นใดทำให้เซลล์พืชกลับไปสู่ สภาวะ เมริสเตมคลอโรพลาสต์และพลาสติดอื่นๆ ก็สามารถเปลี่ยนกลับไปเป็นโปรพลาสติดได้ คลอโรพลาสต์ อะไมโลพลาสต์ โครโมพลาสต์ และโปรพลาสติด ไม่ใช่สภาวะที่แน่นอน สภาวะต่างๆ มักพบได้ทั่วไป[ 177 ]
แผนก
คลอโรพลาสต์ส่วนใหญ่ในเซลล์สังเคราะห์แสงไม่ได้พัฒนาโดยตรงจากโพรพลาสติดหรือเอทิโอพลาสต์ อันที่จริง เซลล์เนื้อเยื่อ เจริญปลายยอดของพืชโดยทั่วไปจะมีโพรพลาสติดเพียง 7–20 โพรพลาสติด เท่านั้น โพรพลาสติดเหล่านี้จะแตกต่างไปเป็นคลอโรพลาสต์ ซึ่งจะแบ่งตัวเพื่อสร้างคลอโรพลาสต์ 30–70 คลอโรพลาสต์ที่พบในเซลล์สังเคราะห์แสงของพืชที่เจริญเต็มที่ หากเซลล์แบ่งตัวการแบ่งตัวของคลอโรพลาสต์จะให้คลอโรพลาสต์เพิ่มเติมเพื่อแบ่งปันระหว่างเซลล์ลูกทั้งสองเซลล์[ 194 ]
ใน สาหร่ายเซลล์เดียวการแบ่งคลอโรพลาสต์เป็นวิธีเดียวในการสร้างคลอโรพลาสต์ใหม่ ไม่มีการแบ่งตัวของโปรพลาสติด—เมื่อเซลล์สาหร่ายแบ่งตัว คลอโรพลาสต์ก็จะแบ่งตัวไปพร้อมกัน และเซลล์ลูก แต่ละเซลล์ จะได้รับคลอโรพลาสต์ที่เจริญเต็มที่[ 193 ]
คลอโรพลาสต์เกือบทั้งหมดในเซลล์จะแบ่งตัว แทนที่จะเป็นกลุ่มเล็กๆ ของคลอโรพลาสต์ที่แบ่งตัวอย่างรวดเร็ว[ 195 ]คลอโรพลาสต์ไม่มีระยะ S ที่แน่นอน —การจำลองดีเอ็นเอของพวกมันไม่ได้ซิงโครไนซ์หรือจำกัดเฉพาะเซลล์เจ้าบ้าน[ 196 ] ความรู้ส่วนใหญ่ที่เรามีเกี่ยวกับการแบ่งตัวของคลอโรพลาสต์มาจากการศึกษาสิ่งมีชีวิตเช่นอาราบิโดปซิสและสาหร่ายสีแดงไซยานิดิโอสคีซอน เมโรเล[ 189 ]
![คลอโรพลาสต์ส่วนใหญ่ในเซลล์พืช และคลอโรพลาสต์ทั้งหมดในสาหร่าย เกิดจากการแบ่งตัวของคลอโรพลาสต์[193] อ้างอิงรูปภาพ[189][197]](https://upload.wikimedia.org/wikipedia/commons/thumb/d/df/Chloroplast_division.svg/960px-Chloroplast_division.svg.png)
{kind=link}
กระบวนการแบ่งตัวเริ่มต้นเมื่อโปรตีนFtsZ1และFtsZ2รวมตัวกันเป็นเส้นใย และด้วยความช่วยเหลือของโปรตีนARC6จะสร้างโครงสร้างที่เรียกว่าวงแหวน Z ภายในสโตรมาของคลอโรพลาสต์[ 189 ] [ 197 ]ระบบMinทำหน้าที่จัดการตำแหน่งของวงแหวน Z เพื่อให้แน่ใจว่าคลอโรพลาสต์จะถูกแบ่งออกอย่างสม่ำเสมอ โปรตีนMinDป้องกันไม่ให้ FtsZ เชื่อมต่อกันและก่อตัวเป็นเส้นใย โปรตีนARC3 อีกตัวหนึ่ง อาจมีส่วนเกี่ยวข้องด้วย แต่ยังไม่เป็นที่เข้าใจดีนัก โปรตีนเหล่านี้ทำงานที่ขั้วของคลอโรพลาสต์ ป้องกันการก่อตัวของวงแหวน Z ที่นั่น แต่ใกล้กับศูนย์กลางของคลอโรพลาสต์MinEจะยับยั้งพวกมัน ทำให้วงแหวน Z สามารถก่อตัวได้[ 189 ]
ต่อไปวงแหวนแบ่งพลาสติด สองวง หรือวงแหวน PD จะก่อตัวขึ้น วงแหวนแบ่งพลาสติดด้านในตั้งอยู่ด้านในของเยื่อหุ้มชั้นในของคลอโรพลาสต์ และก่อตัวขึ้นก่อน[ 189 ]วงแหวนแบ่งพลาสติดด้านนอกพบว่าพันรอบเยื่อหุ้มคลอโรพลาสต์ชั้นนอก ประกอบด้วยเส้นใยที่มีขนาดประมาณ 5 นาโนเมตร[ 189 ]เรียงเป็นแถวห่างกัน 6.4 นาโนเมตร และหดตัวเพื่อบีบคลอโรพลาสต์ นี่คือจุดเริ่มต้นของการหดตัวของคลอโรพลาสต์[ 197 ] ในบางสปีชีส์ เช่นCyanidioschyzon merolæคลอโรพลาสต์จะมีวงแหวนแบ่งพลาสติดวงที่สามตั้งอยู่ในช่องว่างระหว่างเยื่อหุ้มของคลอโรพลาสต์[ 189 ] [ 197 ]
ในช่วงปลายของระยะการหด ตัว โปรตีน ไดนามินจะประกอบกันรอบวงแหวนแบ่งพลาสติดด้านนอก[ 197 ]ช่วยให้เกิดแรงบีบคลอโรพลาสต์[ 189 ]ในขณะเดียวกัน วงแหวน Z และวงแหวนแบ่งพลาสติดด้านในจะสลายตัว[ 197 ]ในระหว่างขั้นตอนนี้ พลาสมิด DNA ของคลอโรพลาสต์จำนวนมากที่ลอยอยู่รอบๆ ในสโตรมาจะถูกแบ่งและกระจายไปยังคลอโรพลาสต์ลูกสาวสองตัวที่กำลังก่อตัว[ 198 ]
ต่อมา ไดนามินจะเคลื่อนที่ไปใต้วงแหวนแบ่งพลาสติดชั้นนอก เข้าไปสัมผัสโดยตรงกับเยื่อหุ้มชั้นนอกของคลอโรพลาสต์[ 197 ]เพื่อแยกคลอโรพลาสต์ออกเป็นคลอโรพลาสต์ลูกสาว 2 อัน[ 189 ]
เศษของวงแหวนแบ่งพลาสติดด้านนอกยังคงลอยอยู่ระหว่างคลอโรพลาสต์ลูกสาวทั้งสอง และเศษของวงแหวนไดนามินยังคงติดอยู่กับคลอโรพลาสต์ลูกสาวตัวหนึ่ง[ 197 ]
จากวงแหวนห้าหรือหกวงที่เกี่ยวข้องกับการแบ่งคลอโรพลาสต์ มีเพียงวงแหวนแบ่งพลาสติดด้านนอกเท่านั้นที่มีอยู่ตลอดระยะการหดตัวและการแบ่ง—ในขณะที่วงแหวน Z ก่อตัวขึ้นก่อน การหดตัวจะไม่เริ่มต้นจนกว่าวงแหวนแบ่งพลาสติดด้านนอกจะก่อตัวขึ้น[ 197 ]

ระเบียบข้อบังคับ
ใน สาหร่ายชนิดที่มีคลอโรพลาสต์เพียงอันเดียว การควบคุมการแบ่งตัวของคลอโรพลาสต์มีความสำคัญอย่างยิ่งเพื่อให้แน่ใจว่าเซลล์ลูกแต่ละเซลล์จะได้รับคลอโรพลาสต์—คลอโรพลาสต์ไม่สามารถสร้างขึ้นมาใหม่ได้[ 91 ] [ 189 ]ในสิ่งมีชีวิตเช่นพืช ซึ่งเซลล์มีคลอโรพลาสต์หลายอัน การประสานงานจะหลวมกว่าและมีความสำคัญน้อยกว่า เป็นไปได้ว่าการแบ่งตัวของคลอโรพลาสต์และเซลล์มีการประสานกันบ้าง แม้ว่ากลไกส่วนใหญ่ยังไม่เป็นที่รู้จัก[ 189 ]
แสงได้รับการพิสูจน์แล้วว่าเป็นสิ่งจำเป็นสำหรับการแบ่งตัวของคลอโรพลาสต์ คลอโรพลาสต์สามารถเจริญเติบโตและก้าวหน้าผ่านขั้นตอนการหดตัวบางส่วนได้ภายใต้แสงสีเขียวคุณภาพต่ำแต่จะแบ่งตัวได้ช้า—จำเป็นต้องได้รับแสงสีขาวที่สว่างจ้าจึงจะแบ่งตัวได้ ใบผักโขมที่ปลูกภายใต้แสงสีเขียวพบว่ามีคลอโรพลาสต์รูปทรงดัมเบลขนาดใหญ่จำนวนมาก การได้รับแสงสีขาวสามารถกระตุ้นให้คลอโรพลาสต์เหล่านี้แบ่งตัวและลดจำนวนคลอโรพลาสต์รูปทรงดัมเบลได้[ 195 ] [ 198 ]
การถ่ายทอดทางพันธุกรรมของคลอโรพลาสต์
เช่นเดียวกับไมโตคอนเดรีย คลอโรพลาสต์มักจะได้รับการถ่ายทอดมาจากพ่อแม่เพียงฝ่ายเดียว การถ่ายทอดคลอโรพลาสต์จากพ่อแม่ทั้งสองฝ่าย—ซึ่งยีนของพลาสติดได้รับการถ่ายทอดมาจากพืชพ่อแม่ทั้งสอง—เกิดขึ้นในระดับที่ต่ำมากในพืชดอกบางชนิด[ 199 ]
กลไกหลายอย่างป้องกันการถ่ายทอดดีเอ็นเอคลอโรพลาสต์จากพ่อแม่ทั้งสองฝ่าย รวมถึงการทำลายคลอโรพลาสต์หรือยีนของคลอโรพลาสต์ภายในแกมีตหรือไซโกต แบบเลือกสรร และการกีดกันคลอโรพลาสต์จากพ่อแม่ฝ่ายใดฝ่ายหนึ่งออกจากเอ็มบริโอ คลอโรพลาสต์ของพ่อแม่สามารถถูกคัดแยกเพื่อให้มีเพียงชนิดเดียวในลูกหลานแต่ละตัว[ 200 ]
พืชเมล็ด เปลือย เช่นต้นสนส่วนใหญ่จะถ่ายทอดคลอโรพลาสต์จากพ่อ[ 201 ]ในขณะที่พืชดอกมักจะได้รับคลอโรพลาสต์จากแม่[ 202 ] [ 203 ]เดิมทีเชื่อกันว่าพืชดอกได้รับคลอโรพลาสต์จากแม่เท่านั้น แต่ปัจจุบันมีหลักฐานมากมายที่แสดงให้เห็นว่าพืชดอกได้รับคลอโรพลาสต์จากพ่อ[ 199 ]
พืชดอกซึ่งส่งต่อคลอโรพลาสต์จากแม่สู่ลูก มีหลายวิธีในการป้องกันการถ่ายทอดทางพ่อ ส่วนใหญ่สร้างเซลล์สเปิร์มที่ไม่มีพลาสติด นอกจากนี้ยังมีกลไกอื่นๆ ที่ได้รับการบันทึกไว้มากมายที่ป้องกันการถ่ายทอดทางพ่อในพืชดอกเหล่านี้ เช่น อัตราการจำลองคลอโรพลาสต์ที่แตกต่างกันภายในเอ็มบริโอ[ 199 ]
ในกลุ่มพืชดอก การถ่ายทอดคลอโรพลาสต์จากพ่อมักพบในลูกผสม มากกว่า ในลูกหลานจากพ่อแม่ที่เป็นสายพันธุ์เดียวกัน ซึ่งแสดงให้เห็นว่ายีนลูกผสมที่ไม่เข้ากันอาจรบกวนกลไกที่ป้องกันการถ่ายทอดจากพ่อ[ 199 ]
พืชทรานส์พลาสโตมิก
เมื่อไม่นานมานี้ คลอโรพลาสต์ได้รับความสนใจจากนักพัฒนาพืชดัดแปลงพันธุกรรมเนื่องจากในพืชดอกส่วนใหญ่ คลอโรพลาสต์ไม่ได้ถูกถ่ายทอดมาจากพ่อพันธุ์ ดังนั้นยีนที่ถูกถ่ายทอดในพลาสติดเหล่านี้จึงไม่สามารถแพร่กระจายผ่านละอองเรณูได้ นี่ทำให้การเปลี่ยนแปลงพลาสติดเป็นเครื่องมือที่มีค่าสำหรับการสร้างและการเพาะปลูกพืชดัดแปลงพันธุกรรมที่ถูกควบคุมทางชีวภาพ ซึ่งก่อให้เกิดความเสี่ยงต่อสิ่งแวดล้อมน้อยลงอย่างมาก กลยุทธ์ การควบคุมทางชีวภาพ นี้ จึงเหมาะสมสำหรับการสร้างการอยู่ร่วมกันของเกษตรกรรมแบบดั้งเดิมและเกษตรอินทรีย์แม้ว่าความน่าเชื่อถือของกลไกนี้ยังไม่ได้มีการศึกษาในพืชทุกชนิดที่เกี่ยวข้อง แต่ผลลัพธ์ล่าสุดในต้นยาสูบนั้นมีแนวโน้มที่ดี โดยแสดงให้เห็นอัตราความล้มเหลวในการควบคุมของพืชที่มีพลาสติดที่ถูกถ่ายทอดที่ 3 ใน 1,000,000 [ 203 ]
เชิงอรรถ
- ^ a bอย่าสับสนกับโครมาโทฟอร์ซึ่งเป็นเซลล์ที่มีเม็ดสีในสัตว์บางชนิด หรือโครมาโทฟอร์ ซึ่งเป็นถุงที่เกี่ยวข้องกับเยื่อหุ้มเซลล์ในแบคทีเรียบางชนิด
- ด้วยเหตุนี้ คลอโรพลาสต์ของกลอโคไฟต์จึงถูกเรียกว่า 'มูโรพลาสต์' ซึ่งมาจากภาษาละติน muroที่แปลว่ากำแพง
ลิงก์ภายนอก
- คลอโรพลาสต์ – ฐานข้อมูลที่เน้นเซลล์เป็นศูนย์กลาง
- Clegg MT, Gaut BS, Learn GH, Morton BR (กรกฎาคม 1994). "อัตราและรูปแบบของการวิวัฒนาการของดีเอ็นเอคลอโรพลาสต์" . Proceedings of the National Academy of Sciences of the United States of America . 91 (15): 6795– 801. Bibcode : 1994PNAS...91.6795C . doi : 10.1073/pnas.91.15.6795 . PMC 44285 . PMID 8041699 .
- งานวิจัยร่วมเพิ่มเติมเกี่ยวกับการเปลี่ยนแปลงของคลอโรพลาสต์
- จีโนมคลอโรพลาสต์ฉบับสมบูรณ์ของ NCBI
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ คลอโรพลาสต์
คลอ โรพลาสต์ ( / ˈ k l ɔːr ə ˌ p l æ s t , - p l ɑː s t / KLOR -ə-plast, -plahst ) [ 1 ] [ 2 ] เป็น ออร์แกเนลล์ ชนิดหนึ่งที่เรียกว่า พลาสติด ซึ่งทำหน้าที่ สังเคราะห์แสง...
การค้นพบและนิรุกติศาสตร์
คำอธิบายที่ชัดเจนครั้งแรกของคลอโรพลาสต์ ( Chlorophyllkörnen , "เมล็ดคลอโรฟิลล์") ได้รับการกล่าวถึงโดย Hugo von Mohl ในปี 1837 ว่าเป็นส่วนประกอบที่แยกจากกันภายในเซลล์พืชสีเขียว [ 4 ] ในปี 1883 Andreas Franz Wilhelm Schimper ได้ตั้งชื่อส่วนประกอบเหล่านี้ว่า...
กำเนิดคลอโรพลาสต์แบบเอนโดซิมไบโอซิส
คลอโรพลาสต์เป็นออร์แกเนลล์ชนิดหนึ่งในเซลล์ยูคาริโอตที่สังเคราะห์แสงได้ พวกมันวิวัฒนาการมาจากไซ ยาโนแบคทีเรีย ผ่านกระบวนการที่เรียกว่า ออร์แกเนลล์เจเนซิส [ 10 ] ไซ ยาโนแบคทีเรียเป็น ไฟลัม ที่หลากหลายของ แบคทีเรียแก รมลบ ที่สามารถสังเคราะห์ แสงแบบใช้ออกซิเจน...
เอนโดซิมไบโอซิสขั้นต้น
เมื่อประมาณสอง พันล้านปีก่อน [ 18 ] [ 19 ] [ 20 ] ไซยาโนแบคทีเรียที่ ดำรงชีวิตอิสระได้เข้าไปใน เซลล์ ยูคาริโอต ยุคแรก ไม่ว่าจะเป็นอาหาร หรือ ปรสิต ภายใน [ 17 ] แต่สามารถหลบหนีออกจากแว คิวโอลฟาโกไซติกที่ มันบรรจุอยู่และคงอยู่ภายในเซลล์ได้ [ 12 ]...