อ่าน 12 นาที
เนื้อเยื่อเจริญ
ในชีววิทยาของเซลล์เนื้อเยื่อเจริญเป็นโครงสร้างที่ประกอบด้วยเนื้อเยื่อ เฉพาะ ที่พบในพืช...
เนื้อเยื่อเจริญ
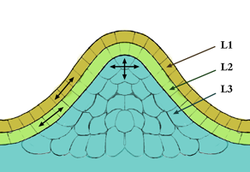
ในชีววิทยาของเซลล์เนื้อเยื่อเจริญเป็นโครงสร้างที่ประกอบด้วยเนื้อเยื่อ เฉพาะ ที่พบในพืช ซึ่งประกอบด้วยเซลล์ต้นกำเนิดที่เรียกว่าเซลล์เจริญซึ่งเป็นเซลล์ที่ยังไม่แตกต่างและสามารถแบ่งตัว ได้อย่างต่อเนื่อง เซลล์เจริญเหล่านี้มีบทบาทสำคัญใน การเจริญ เติบโตการสร้างใหม่และการปรับตัวของพืชเนื่องจากเป็นแหล่งกำเนิดของเนื้อเยื่อและอวัยวะที่แตกต่างกัน ทั้งหมดของพืช พวกมันมีส่วนช่วยในการสร้างโครงสร้างต่างๆ เช่น ผลไม้ ใบ และเมล็ด รวมถึงเนื้อเยื่อค้ำจุน เช่น ลำต้นและราก[ 1 ]
เซลล์เมริสเต็มเป็นเซลล์ที่มีศักยภาพในการเปลี่ยนแปลงไปเป็นเซลล์พืชชนิดใดก็ได้ หมายความว่าพวกมันมีความสามารถในการพัฒนาไปเป็นเซลล์พืชชนิดใดก็ได้ เมื่อพวกมันแบ่งตัว พวกมันจะสร้างเซลล์ใหม่ ซึ่งบางส่วนยังคงเป็นเซลล์เมริสเต็ม ในขณะที่บางส่วนจะพัฒนาไปเป็นเซลล์เฉพาะทาง ซึ่งโดยทั่วไปแล้วจะสูญเสียความสามารถในการแบ่งตัวหรือสร้างเซลล์ชนิดใหม่ เนื่องจากการแบ่งตัวอย่างต่อเนื่องและลักษณะที่ไม่จำเพาะเจาะจง เซลล์เมริสเต็มจึงเป็นรากฐานสำหรับการสร้างอวัยวะพืชใหม่และการขยายตัวอย่างต่อเนื่องของลำต้นพืชตลอดวงจรชีวิตของพืช
เซลล์เมริสเตมเป็นเซลล์ขนาดเล็ก มีผนังเซลล์ปฐมภูมิ ที่บาง และ มี แวคิวโอล ขนาดเล็กหรือไม่มีเลย โปรโตพลาซึมของเซลล์เหล่านี้มีความหนาแน่นสูง เติมเต็มเซลล์ทั้งหมด และไม่มีช่องว่างระหว่างเซลล์ แทนที่จะมีพลาสติดที่เจริญ เต็มที่ เช่นคลอโรพลาสต์หรือโค ร โมพลาสต์ เซลล์ เหล่านี้จะมีโปรพลาสติดซึ่งจะพัฒนาไปเป็นพลาสติดที่ทำงานได้อย่างสมบูรณ์ในภายหลัง
เนื้อเยื่อเจริญแบ่งออกเป็นสามประเภทหลักตามตำแหน่งและหน้าที่ ได้แก่ เนื้อเยื่อเจริญ ปลายยอดซึ่งพบที่ปลายรากและยอด เนื้อเยื่อ เจริญระหว่างกลางหรือฐานซึ่งอยู่บริเวณกลางลำต้นหรือใบ ทำหน้าที่ช่วยในการเจริญเติบโตใหม่และ เนื้อเยื่อเจริญ ด้านข้างหรือแคมเบียมซึ่งรับผิดชอบการเจริญเติบโตทุติยภูมิในพืชเนื้อไม้ที่ส่วนยอดของเนื้อเยื่อเจริญจะมีกลุ่มเซลล์ขนาดเล็กที่แบ่งตัวช้าๆ เรียกว่าโซนกลาง ทำหน้าที่เป็นแหล่งสะสมของเซลล์ต้นกำเนิด ซึ่งจำเป็นต่อการรักษาการทำงานของเนื้อเยื่อเจริญ อัตราการเจริญเติบโตและการเพิ่มจำนวนของเซลล์ภายในเนื้อเยื่อเจริญจะแตกต่างกัน โดยมีการทำงานที่บริเวณรอบนอกสูงกว่าบริเวณกลาง
คำว่าเมริสเต็มถูกใช้ครั้งแรกในปี พ.ศ. 2391 โดยนักพฤกษศาสตร์ชาวสวิสCarl Wilhelm von Nägeli (พ.ศ. 2360–2334) ในหนังสือBeiträge zur Wissenschaftlichen Botanik ("Contributions to Scientific Botany") [ 2 ]มาจากภาษากรีก μερίζειν (merizein) ' แบ่ง'เพื่อเป็นการยอมรับหน้าที่ที่แท้จริงของมัน
เนื้อเยื่อเจริญขั้นต้น
เนื้อเยื่อเจริญปลายยอด หรือที่เรียกว่าเนื้อเยื่อเจริญปฐมภูมิ ก่อให้เกิดลำต้นหลักของพืชและมีหน้าที่ในการเจริญเติบโตขั้นต้นหรือการเพิ่มความยาวหรือความสูง[ 3 ] [ 4 ]เนื้อเยื่อเจริญปลายยอดอาจแบ่งตัวออกเป็นเนื้อเยื่อเจริญปฐมภูมิได้ 3 ชนิด:
- โปรโตเดิร์ม: อยู่รอบนอกของลำต้นและพัฒนาไปเป็นชั้นหนังกำพร้า
- โปรแคมเบียม: อยู่ด้านในของโปรโตเดอร์มและพัฒนาเป็นไซเล็มปฐมภูมิและโฟล เอมปฐมภูมิ นอกจากนี้ยังสร้างแคมเบียมหลอดเลือดและแคมเบียมเปลือกไม้ (ส่วนหนึ่งของเมริสเต็มทุติยภูมิ แต่เป็นลูกหลานของเซลล์เมริสเต็มปลายยอด) แคมเบียมเปลือกไม้จะแตกต่างไปเป็นเฟลโลเดอร์มหรือเปลือกไม้ (ด้านใน) และเฟลเล็มหรือเปลือกไม้ (ด้านนอก) ทั้งสามชั้นนี้ (แคมเบียมเปลือกไม้ เฟลเล็ม และเฟลโลเดอร์ม) ประกอบกันเป็นเพอริเดอร์มในราก โปรแคมเบียมยังสามารถก่อให้เกิดเพอริไซเคิลซึ่งสร้างรากข้างในยูไดคอต[ 5 ]
- เนื้อเยื่อเจริญพื้นฐาน: ประกอบด้วยเซลล์ พาเรนไค มาคอลเลนไคมาและสเคลเรนไคมาของเนื้อเยื่อพื้นฐาน[ 5 ]ที่พัฒนาเป็นคอร์เทกซ์และพิธ
เนื้อเยื่อเจริญทุติยภูมิ
หลังจากระยะการเจริญเติบโตหลักแล้ว เนื้อเยื่อเจริญด้านข้างจะพัฒนาขึ้นเป็นระยะการเจริญเติบโตทุติยภูมิ การเจริญเติบโตนี้จะเพิ่มขนาดเส้นผ่านศูนย์กลางของพืชจากลำต้นเดิม แต่ไม่ใช่พืชทุกต้นที่จะมีการเจริญเติบโตทุติยภูมิ เนื้อเยื่อเจริญทุติยภูมิมีสองประเภท ได้แก่ แคมเบียมหลอดเลือดและแคมเบียมเปลือก
- แคมเบียมหลอดเลือด (Vascular cambium ) สร้างไซเล็มทุติยภูมิและโฟลเอมทุติยภูมิ กระบวนการนี้อาจเกิดขึ้นต่อเนื่องตลอดอายุของพืช ซึ่งเป็นสิ่งที่ทำให้เกิดเนื้อไม้ในพืช พืชเหล่านี้เรียกว่าพืชยืนต้น (arboraceous plants) กระบวนการนี้จะไม่เกิดขึ้นในพืชที่ไม่มีการเจริญเติบโตทุติยภูมิ ซึ่งเรียกว่าพืชล้มลุก (herbaceous plants)
- เนื้อเยื่อคอร์กแคมเบียมซึ่งพัฒนาไปเป็นเนื้อเยื่อเพอริเดอร์ม ซึ่งจะมาแทนที่เนื้อเยื่อเอพิเดอร์มิส และสร้างเปลือกไม้และเนื้อไม้คอร์ก เป็นต้น
เนื้อเยื่อเจริญปลายยอด

- เขตกลาง
- เขตชายขอบ
- เนื้อเยื่อเจริญส่วนกลาง (เช่น เนื้อเยื่อเจริญไขกระดูก)
- เนื้อเยื่อไขกระดูก
เนื้อเยื่อเจริญปลายยอดเป็นเนื้อเยื่อเจริญที่ยังไม่แบ่งตัวอย่างสมบูรณ์ (ไม่จำกัดการเจริญเติบโต) ของพืช ทำหน้าที่สร้างการเจริญเติบโตขั้นต้น ทำให้ลำต้นและรากยืดตัวออก เนื้อเยื่อเจริญปลายยอดจะสร้างเนื้อเยื่อเจริญขั้นต้นได้ 3 ชนิด ซึ่งต่อมาจะพัฒนาไปเป็นเนื้อเยื่อเจริญขั้นทุติยภูมิหรือเนื้อเยื่อเจริญด้านข้าง ซึ่งมีส่วนช่วยในการขยายตัวด้านข้างของพืช
เนื้อเยื่อเจริญปลายยอดมีสองประเภทหลัก ได้แก่ เนื้อเยื่อ เจริญปลายยอดของลำต้น (SAM) และ เนื้อเยื่อเจริญปลาย ราก (RAM) SAM อยู่ที่ปลายยอดและทำหน้าที่สร้างใบ ลำต้น และดอก ในขณะที่ RAM อยู่ที่ปลายรากและทำหน้าที่สร้างเนื้อเยื่อรากใหม่ ทั้งสองประเภทประกอบด้วยเซลล์ที่แบ่งตัวอย่างรวดเร็วและไม่จำกัดระยะการแบ่งตัว หมายความว่าพวกมันสร้างเซลล์ใหม่ตลอดเวลาโดยไม่มีระยะสุดท้ายที่กำหนดไว้ล่วงหน้า คล้ายกับเซลล์ต้นกำเนิดในสัตว์ ซึ่งมีพฤติกรรมและหน้าที่คล้ายคลึงกัน
ในเชิงโครงสร้าง เนื้อเยื่อเจริญปลายยอดจะจัดเรียงเป็นโซนที่แตกต่างกัน โซนกลางทำหน้าที่เป็นแหล่งสะสมของเซลล์ที่ยังไม่แตกต่าง ในขณะที่โซนรอบนอกสร้างอวัยวะและเนื้อเยื่อใหม่ เนื้อเยื่อเจริญไขกระดูกมีส่วนช่วยในการพัฒนาหลอดเลือด โดยสร้างเนื้อเยื่อไขกระดูกซึ่งเป็นโครงสร้างส่วนกลางของพืช ชั้นของเนื้อเยื่อเจริญยังแตกต่างกันไปตามชนิดของพืช ชั้นนอกสุดที่เรียกว่าทูนิกาจะกำหนดขอบและแนวขอบของใบในพืชใบเลี้ยงเดี่ยวในขณะที่ในพืชใบเลี้ยงคู่ ชั้นที่สองของคอร์ปัสจะมีอิทธิพลต่อลักษณะของใบ
โดยทั่วไปเนื้อเยื่อเจริญปลายยอดจะพบที่ปลายรากและลำต้น แต่ในพืชบางชนิดที่อาศัยอยู่ในเขตอาร์กติกเนื้อเยื่อเจริญปลายยอดจะอยู่ที่ส่วนล่างหรือส่วนกลางของพืช เชื่อกันว่า การปรับตัว เช่นนี้ ช่วยให้พืชมีข้อได้เปรียบในสภาพแวดล้อมที่รุนแรง
เนื้อเยื่อเจริญปลายยอด



เนื้อเยื่อเจริญปลายยอดเป็นแหล่งกำเนิดของอวัยวะเหนือดินทั้งหมด เช่น ใบและดอก เซลล์ที่ส่วนยอดของเนื้อเยื่อเจริญปลายยอดทำหน้าที่เป็นเซลล์ต้นกำเนิดสำหรับบริเวณรอบข้าง ซึ่งเซลล์เหล่านี้จะขยายพันธุ์อย่างรวดเร็วและถูกรวมเข้ากับใบหรือดอกที่กำลังพัฒนา
เนื้อเยื่อเจริญปลายยอดเป็นบริเวณที่เกิดการเจริญเติบโตของเอ็มบริโอส่วนใหญ่ในพืชดอกเนื้อเยื่อเริ่มต้นของใบ กลีบเลี้ยง กลีบดอก เกสรตัวผู้ และรังไข่ เริ่มต้นขึ้นที่นี่ในอัตราหนึ่งหน่วยต่อช่วงเวลาที่เรียกว่าพลาสโตครอน บริเวณนี้เป็นที่ที่แสดงสัญญาณแรกของการเจริญเติบโตของดอก หนึ่งในสัญญาณเหล่านี้อาจเป็นการสูญเสียการควบคุมของปลายยอดและการปลดปล่อยเซลล์ที่อยู่เฉยๆ ให้เจริญเติบโตเป็นเนื้อเยื่อเจริญปลายยอดเสริม ในบางชนิดอาจเกิดขึ้นที่ซอกใบของเนื้อเยื่อเริ่มต้นที่อยู่ใกล้กับปลายยอดเพียงสองหรือสามหน่วย
เนื้อเยื่อเจริญปลายยอดประกอบด้วยกลุ่มเซลล์ที่แตกต่างกันสี่กลุ่ม:
- เซลล์ต้นกำเนิด
- เซลล์ลูกสาวโดยตรงของเซลล์ต้นกำเนิด
- ศูนย์จัดระเบียบที่อยู่เบื้องล่าง
- เซลล์ต้นกำเนิดสำหรับการสร้างอวัยวะในบริเวณโดยรอบ
โซนที่แตกต่างกันทั้งสี่นี้ได้รับการรักษาไว้โดยเส้นทางการส่งสัญญาณที่ซับซ้อน ในArabidopsis thaliana ยีน CLAVATAที่โต้ตอบกัน 3 ยีนจำเป็นต่อการควบคุมขนาดของ แหล่งเก็บ เซลล์ต้นกำเนิด ในเนื้อเยื่อเจริญปลายยอดโดยการ ควบคุมอัตราการแบ่งเซลล์[ 6 ] CLV1และ CLV2 คาดว่าจะก่อตัวเป็นคอมเพล็กซ์ตัวรับ (ของ ตระกูล ไคเนสคล้ายตัวรับ LRR ) ซึ่ง CLV3 เป็นลิแกนด์ [ 7 ] [ 8 ] [ 9 ] CLV3มีความคล้ายคลึงกับโปรตีน ESR ของข้าวโพด โดยมีบริเวณกรดอะมิโน 14 ตัวสั้นๆ ที่ได้ รับการอนุรักษ์ไว้ระหว่างโปรตีน[ 10 ] [ 11 ]โปรตีนที่มีบริเวณที่ได้รับการอนุรักษ์เหล่านี้ถูกจัดกลุ่มเป็นโปรตีนตระกูล CLE [ 10 ] [ 11 ]
CLV1 ได้รับการแสดงให้เห็นว่ามีปฏิสัมพันธ์กับ โปรตีน ไซโตพลาสมิก หลายชนิด ที่น่าจะเกี่ยวข้องกับการส่งสัญญาณปลายทางตัวอย่างเช่น พบว่าคอมเพล็กซ์ CLV เกี่ยวข้องกับโปรตีนที่เกี่ยวข้องกับ GTPase ขนาดเล็ก Rho/Rac [ 6 ] โปรตีนเหล่านี้อาจทำหน้าที่เป็นตัวกลางระหว่างคอมเพล็กซ์ CLV และโปรตีนไคเนสที่กระตุ้นด้วยไมโทเจน (MAPK) ซึ่งมักเกี่ยวข้องกับลำดับการส่งสัญญาณ[ 12 ] KAPP เป็น โปรตีนฟอสฟาเทสที่เกี่ยวข้องกับ ไคเนสซึ่งได้รับการแสดงให้เห็นว่ามีปฏิสัมพันธ์กับ CLV1 [ 13 ]เชื่อกันว่า KAPP ทำหน้าที่เป็นตัวควบคุมเชิงลบของ CLV1 โดยการกำจัดฟอสเฟตออกจากมัน[ 13 ]
ยีนสำคัญอีกตัวหนึ่งในการบำรุงรักษาเมริสเต็มของพืชคือWUSCHEL (ย่อเป็นWUS ) ซึ่งเป็นเป้าหมายของการส่งสัญญาณ CLV นอกเหนือจากการควบคุม CLV ในเชิงบวก จึงก่อให้เกิดวงจรป้อนกลับ[ 14 ] WUSถูกแสดงออกในเซลล์ที่อยู่ใต้เซลล์ต้นกำเนิดของเมริสเต็ม และการมีอยู่ของมันจะป้องกันการแบ่งตัวของเซลล์ต้นกำเนิด[ 14 ] CLV1 ทำหน้าที่ส่งเสริมการแบ่งตัวของเซลล์โดยการยับยั้ง กิจกรรม ของ WUSนอกโซนกลางที่มีเซลล์ต้นกำเนิด[ 6 ]
หน้าที่ของWUSในเนื้อเยื่อเจริญปลายยอดเชื่อมโยงกับฮอร์โมนพืชไซโตไคนิน ไซโตไคนินกระตุ้นฮิสติดีนไคเนสซึ่งจะทำการฟอสโฟรีเลตโปรตีนถ่ายโอนฟอสเฟตฮิสติ ดีน [ 15 ]ต่อมาหมู่ฟอสเฟตจะถูกถ่ายโอนไปยังตัวควบคุมการตอบสนองของ Arabidopsis (ARRs) สองประเภท ได้แก่ ARRS ประเภท B และ ARRs ประเภท ARRs ทำหน้าที่เป็นปัจจัยการถอดรหัสเพื่อกระตุ้นยีนที่อยู่ปลายน้ำของไซโตไคนินรวมถึง A-ARRs ด้วย A-ARRs มีโครงสร้างคล้ายกับ B-ARRs แต่ A-ARRs ไม่มีโดเมนการจับกับ DNA ที่ B-ARRs มี ซึ่งจำเป็นต่อการทำหน้าที่เป็นปัจจัยการถอดรหัส[ 16 ]ดังนั้น A-ARRs จึงไม่มีส่วนช่วยในการกระตุ้นการถอดรหัส และโดยการแข่งขันกับฟอสเฟตจากโปรตีนถ่ายโอนฟอสเฟต จึงยับยั้งการทำงานของ B-ARRs [ 17 ]ใน SAM, B-ARRs กระตุ้นการแสดงออกของWUSซึ่งกระตุ้นเอกลักษณ์ของเซลล์ต้นกำเนิด[ 18 ] จากนั้น WUSจะยับยั้ง A-ARRs [ 19 ]ส่งผลให้ B-ARRs ไม่ถูกยับยั้งอีกต่อไป ทำให้เกิดการส่งสัญญาณไซโตไคนินอย่างต่อเนื่องในใจกลางของเนื้อเยื่อเจริญปลายยอด เมื่อรวมกับการส่งสัญญาณ CLAVATA ระบบนี้ทำงานเป็น วงจร ป้อนกลับเชิงลบ การส่งสัญญาณไซโตไคนินได้รับการเสริมแรงในเชิงบวกโดย WUS เพื่อป้องกันการยับยั้งการส่งสัญญาณไซโตไคนิน ในขณะที่ WUS ส่งเสริมตัวยับยั้งของตัวเองในรูปของ CLV3 ซึ่งในที่สุดจะควบคุม WUS และการส่งสัญญาณไซโตไคนิน[ 20 ]
เนื้อเยื่อเจริญปลายราก

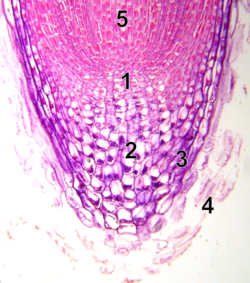
- ศูนย์กลางที่สงบ
- แคลิปโตรเจน (เซลล์ปลายรากที่มีชีวิต)
- รูทแคป
- เซลล์หมวกรากที่ตายแล้วหลุดลอกออกไป
- โปรแคมเบียม
แตกต่างจากเนื้อเยื่อเจริญปลายยอด เนื้อเยื่อเจริญปลายรากสร้างเซลล์ในสองมิติ โดยมีกลุ่มเซลล์ต้นกำเนิด สองกลุ่ม อยู่รอบศูนย์กลางการจัดระเบียบที่เรียกว่าเซลล์ศูนย์กลางสงบ (QC) และร่วมกันสร้างเซลล์ส่วนใหญ่ในรากที่โตเต็มที่[ 21 ] [ 22 ]ที่ปลายสุด เนื้อเยื่อเจริญปลายรากถูกปกคลุมด้วยหมวกราก ซึ่งทำหน้าที่ปกป้องและนำทางการเจริญเติบโต เซลล์จะหลุดออกจากผิวด้านนอกของหมวกราก อย่างต่อเนื่อง เซลล์ QC มีลักษณะเฉพาะคือมีกิจกรรมไมโทซิสต่ำ หลักฐานชี้ให้เห็นว่า QC รักษาเซลล์ต้นกำเนิดโดยรอบโดยการป้องกันการเปลี่ยนแปลงสภาพของเซลล์เหล่านั้น ผ่านสัญญาณที่ยังไม่ถูกค้นพบ ซึ่งช่วยให้มีเซลล์ใหม่ในเนื้อเยื่อเจริญอย่างต่อเนื่องที่จำเป็นสำหรับการเจริญเติบโตของรากอย่างต่อเนื่อง การค้นพบล่าสุดบ่งชี้ว่า QC ยังสามารถทำหน้าที่เป็นแหล่งเก็บเซลล์ต้นกำเนิดเพื่อเติมเต็มสิ่งที่สูญเสียหรือเสียหายได้[ 23 ]เนื้อเยื่อปลายรากและรูปแบบเนื้อเยื่อจะถูกสร้างขึ้นในเอ็มบริโอในกรณีของรากหลัก และในต้นอ่อนรากแขนงใหม่ในกรณีของรากรอง
เนื้อเยื่อเจริญแทรกกลาง
ในพืชดอก (angiosperms ) เนื้อเยื่อเจริญแทรกกลาง (บางครั้งเรียกว่าเนื้อเยื่อเจริญฐาน) พบได้ในลำต้นของพืชใบเลี้ยงเดี่ยว (โดยเฉพาะหญ้า ) ที่โคนข้อและใบ หญ้า หางม้าและเวลวิตเชีย (Welwitschia)ก็มีการเจริญเติบโตแบบแทรกกลางเช่นกัน เนื้อเยื่อเจริญแทรกกลางสามารถแบ่งเซลล์ได้ และช่วยให้พืชใบเลี้ยงเดี่ยวหลายชนิดเจริญเติบโตและงอกใหม่ได้อย่างรวดเร็ว เนื้อเยื่อเจริญแทรกกลางที่ข้อของต้นไผ่ช่วยให้ลำต้นยืดตัวได้อย่างรวดเร็ว ในขณะที่เนื้อเยื่อเจริญแทรกกลางที่โคนใบของหญ้าส่วนใหญ่ช่วยให้ใบที่เสียหายงอกใหม่ได้อย่างรวดเร็ว การงอกใหม่ของใบในหญ้าวิวัฒนาการขึ้นเพื่อตอบสนองต่อความเสียหายจากสัตว์กินพืชและ/หรือไฟป่า
เนื้อเยื่อเจริญของดอกไม้
เมื่อพืชเริ่มออกดอก เนื้อเยื่อเจริญปลายยอดจะเปลี่ยนไปเป็นเนื้อเยื่อเจริญช่อดอก ซึ่งจะพัฒนาต่อไปเป็นเนื้อเยื่อเจริญดอก ที่สร้างกลีบเลี้ยงกลีบดอกเกสรตัวผู้และเกสรตัวเมียของดอกไม้
ตรงกันข้ามกับเมริสเต็มปลายยอดพืชและเมริสเต็มช่อดอกบางส่วน เมริสเต็มดอกไม่สามารถเจริญเติบโตต่อไปได้เรื่อยๆการเจริญเติบโตของมันถูกจำกัดไว้ที่ดอกที่มีขนาดและรูปร่างเฉพาะ การเปลี่ยนจากเมริสเต็มยอดไปเป็นเมริสเต็มดอกต้องอาศัยยีนกำหนดเอกลักษณ์ของเมริสเต็มดอก ซึ่งทั้งกำหนดอวัยวะดอกและทำให้การสร้างเซลล์ต้นกำเนิดสิ้นสุดลงAGAMOUS ( AG ) เป็นยีนโฮมีโอติกของดอกที่จำเป็นสำหรับการสิ้นสุดของเมริสเต็มดอกและจำเป็นสำหรับการพัฒนาที่เหมาะสมของเกสรตัวผู้และเกสรตัวเมีย [ 6 ] AGจำเป็นต่อการป้องกันการเปลี่ยนเมริสเต็มดอกไปเป็นเมริสเต็มยอดช่อดอก แต่เป็นยีนกำหนดเอกลักษณ์LEAFY ( LFY ) และWUSและถูกจำกัดไว้ที่ศูนย์กลางของเมริสเต็มดอกหรือวงกลีบ ด้านในสอง วง[ 24 ]ด้วยวิธีนี้จึงสามารถกำหนดเอกลักษณ์ของดอกและความเฉพาะเจาะจงของภูมิภาคได้ WUS กระตุ้น AG โดยการจับกับลำดับคอนเซนซัสในอินทรอนที่สองของ AG และ LFY จับกับไซต์การรับรู้ที่อยู่ติดกัน[ 24 ]เมื่อ AG ถูกกระตุ้นแล้ว มันจะยับยั้งการแสดงออกของ WUS ซึ่งนำไปสู่การสิ้นสุดของเมริสเต็ม[ 24 ]
ตลอดหลายปีที่ผ่านมา นักวิทยาศาสตร์ได้ทำการดัดแปลงเนื้อเยื่อเจริญของดอกไม้เพื่อผลประโยชน์ทางเศรษฐกิจ ตัวอย่างเช่น ต้นยาสูบกลายพันธุ์ "แมริแลนด์แมมมอธ" ในปี 1936 กระทรวงเกษตรของสวิตเซอร์แลนด์ได้ทำการทดสอบทางวิทยาศาสตร์หลายอย่างกับพืชชนิดนี้ "แมริแลนด์แมมมอธ" มีลักษณะพิเศษคือมันเติบโตเร็วกว่าต้นยาสูบชนิดอื่นมาก
การครอบงำของปลายยอด
การครอบงำของยอดคือภาวะที่เนื้อเยื่อเจริญหนึ่งส่วนยับยั้งหรือขัดขวางการเจริญเติบโตของเนื้อเยื่อเจริญส่วนอื่น ส่งผลให้พืชมีลำต้นหลักที่เห็นได้ชัดเจนเพียงลำต้นเดียว ตัวอย่างเช่น ในต้นไม้ ปลายลำต้นหลักจะมีเนื้อเยื่อเจริญยอดที่เด่นกว่า ดังนั้นปลายลำต้นจึงเจริญเติบโตอย่างรวดเร็วและไม่ถูกบดบังด้วยกิ่งก้าน หากตัดเนื้อเยื่อเจริญที่เด่นกว่าออก ปลายกิ่งหนึ่งหรือหลายกิ่งจะเข้ามาครอบงำแทน กิ่งนั้นจะเริ่มเจริญเติบโตเร็วขึ้นและส่วนที่งอกใหม่จะตั้งตรง เมื่อเวลาผ่านไป กิ่งนั้นอาจเริ่มดูเหมือนเป็นส่วนต่อขยายของลำต้นหลักมากขึ้นเรื่อยๆ บ่อยครั้งที่หลายกิ่งจะแสดงพฤติกรรมนี้หลังจากตัดเนื้อเยื่อเจริญส่วนปลายออก ทำให้พืชเจริญเติบโตเป็นพุ่ม
กลไกการควบคุมการเจริญเติบโตของยอดนั้นอาศัยฮอร์โมน ออกซิน ซึ่งเป็นสารควบคุมการเจริญเติบโตของพืช ฮอร์โมนเหล่านี้ผลิตขึ้นในเนื้อเยื่อเจริญปลายยอดและถูกลำเลียงไปยังรากในเนื้อเยื่อแคมเบียมหากการควบคุมการเจริญเติบโตของยอดสมบูรณ์ ฮอร์โมนเหล่านี้จะป้องกันไม่ให้กิ่งก้านแตกออกมาตราบใดที่เนื้อเยื่อเจริญปลายยอดยังทำงานอยู่ แต่หากการควบคุมไม่สมบูรณ์ กิ่งก้านด้านข้างก็จะแตกออกมา
การตรวจสอบล่าสุดเกี่ยวกับการควบคุมยอดและการควบคุมการแตกกิ่งได้เปิดเผยกลุ่มฮอร์โมนพืชใหม่ที่เรียกว่าสตรีกอลแลคโตนสารประกอบเหล่านี้เคยทราบกันดีว่ามีส่วนเกี่ยวข้องกับการงอกของเมล็ดและการสื่อสารกับเชื้อราไมคอร์ไรซาและขณะนี้พบว่ามีส่วนเกี่ยวข้องกับการยับยั้งการแตกกิ่ง[ 25 ]
ความหลากหลายในโครงสร้างของเนื้อเยื่อเจริญ
SAM ประกอบด้วยประชากรของเซลล์ต้นกำเนิดที่สร้างเนื้อเยื่อเจริญด้านข้างในขณะที่ลำต้นยืดออก ปรากฏว่ากลไกการควบคุมจำนวนเซลล์ต้นกำเนิดอาจได้รับการอนุรักษ์ทางวิวัฒนาการยีนCLAVATA CLV2ที่รับผิดชอบในการรักษาประชากรเซลล์ต้นกำเนิดใน Arabidopsis thaliana มีความสัมพันธ์อย่างใกล้ชิดกับยีน FASCIATED EAR 2 (FEA2) ของข้าวโพดซึ่งเกี่ยวข้องกับหน้าที่เดียวกัน [ 26 ] ในทำนองเดียวกัน ในข้าว ระบบ FON1-FON2 ดูเหมือนจะมีความสัมพันธ์อย่างใกล้ชิดกับระบบส่งสัญญาณ CLV ในArabidopsis thaliana [ 27 ]การศึกษาเหล่านี้ชี้ให้เห็นว่าการควบคุมจำนวนเซลล์ต้นกำเนิดเอกลักษณ์และการแยกแยะ อาจเป็นกลไกที่ได้รับการอนุรักษ์ทางวิวัฒนาการในพืชใบเลี้ยงเดี่ยวหากไม่ใช่ในพืชดอกข้าวยังมีระบบทางพันธุกรรมอีกระบบหนึ่งที่แตกต่างจากFON1-FON2ซึ่งเกี่ยวข้องกับการควบคุมจำนวนเซลล์ต้นกำเนิด[ 27 ]ตัวอย่างนี้เน้นย้ำถึงนวัตกรรมที่เกิดขึ้นในโลกแห่งชีวิตตลอดเวลา
บทบาทของยีนในกลุ่ม KNOX


การตรวจคัดกรองทางพันธุกรรมได้ระบุยีนใน กลุ่ม KNOX ที่ มีหน้าที่นี้ ยีนเหล่านี้ทำหน้าที่รักษาสภาพของเซลล์ต้นกำเนิดให้อยู่ในสภาวะที่ไม่แตกต่าง กลุ่มยีน KNOX มีวิวัฒนาการที่หลากหลายมาก แต่กลไกโดยรวมยังคงคล้ายคลึงกัน พบยีนในกลุ่ม KNOX ในพืชหลากหลายชนิด เช่นอาราบิโดปซิส ทาเลียน่าข้าว ข้าวบาร์เลย์และมะเขือเทศ นอกจากนี้ยังพบยีนที่คล้ายกับ KNOX ในสาหร่าย บางชนิด มอส เฟิร์น และพืชเมล็ดเปลือย การแสดงออกที่ผิดปกติของยีนเหล่านี้ทำให้เกิดลักษณะทางสัณฐานวิทยาที่น่าสนใจ ตัวอย่างเช่น ในกลุ่มพืช วงศ์ย่อย Antirrhineae มีเพียงสกุล Antirrhinumเท่านั้นที่ไม่มีโครงสร้างที่เรียกว่าเดือยในบริเวณดอก เดือยถือเป็นนวัตกรรม ทางวิวัฒนาการ เพราะมันกำหนด ความจำเพาะและการดึงดูด ของแมลงผสมเกสร นักวิจัยได้ทำการกลาย พันธุ์ ทรานสโพซอนในAntirrhinum majusและพบว่าการแทรกบางส่วนทำให้เกิดการสร้างเดือยที่คล้ายกับสมาชิกอื่น ๆ ของAntirrhineae มาก [ 28 ] ซึ่งบ่งชี้ว่าการสูญเสียเดือยใน ประชากร Antirrhinum majus ในป่า อาจเป็นนวัตกรรมทางวิวัฒนาการ
ตระกูล KNOX ยังมีส่วนเกี่ยวข้องกับวิวัฒนาการของรูปร่างใบด้วย(ดูรายละเอียดเพิ่มเติมด้านล่าง) งานวิจัยชิ้นหนึ่งศึกษาแบบแผนการแสดงออกของยีน KNOX ในA. thalianaซึ่งมีใบเรียบง่าย และCardamine hirsutaซึ่งเป็นพืชที่มีใบซับซ้อนในA. thalianaยีน KNOX จะถูกปิดใช้งานอย่างสมบูรณ์ในใบ แต่ในC. hirsutaการแสดงออกยังคงดำเนินต่อไป ทำให้เกิดใบที่ซับซ้อน[ 29 ]นอกจากนี้ ยังมีการเสนอว่ากลไกการทำงานของยีน KNOX นั้นได้รับการอนุรักษ์ไว้ในพืชมีท่อลำเลียง ทั้งหมด เนื่องจากมีความสัมพันธ์ อย่างใกล้ชิด ระหว่างการแสดงออกของ KNOX และสัณฐานวิทยาของใบที่ซับซ้อน[ 30 ]
การเจริญเติบโตแบบไม่จำกัดของเนื้อเยื่อเจริญ
แม้ว่าพืชแต่ละชนิดจะเจริญเติบโตตามกฎเกณฑ์ที่กำหนดไว้ แต่เนื้อเยื่อเจริญของรากและยอด ใหม่แต่ละส่วน สามารถเจริญเติบโตต่อไปได้ตราบเท่าที่มันยังมีชีวิตอยู่ ในพืชหลายชนิด การเจริญเติบโตของเนื้อเยื่อเจริญอาจไม่มีขีดจำกัดทำให้รูปร่างโดยรวมของพืชไม่แน่นอน นี่คือการเจริญเติบโตขั้นต้นการเจริญเติบโตขั้นต้นนำไปสู่การยืดตัวของลำต้นพืชและการสร้างอวัยวะ อวัยวะของพืชทั้งหมดเกิดขึ้นจากกระบวนการแบ่งเซลล์ในเนื้อเยื่อเจริญส่วนปลาย ตามด้วยการขยายตัวและการเปลี่ยนแปลงรูปร่างของเซลล์ การเจริญเติบโตขั้นต้นทำให้เกิดส่วนปลายของพืชหลายชนิด
การเจริญเติบโตของ ปุ่มรากตรึงไนโตรเจนในพืชตระกูลถั่วเช่นถั่วเหลืองและถั่วลันเตาอาจเป็นแบบจำกัดหรือแบบไม่จำกัด ดังนั้น ถั่วเหลือง (หรือถั่วและโลตัสญี่ปุ่น ) จะสร้างปุ่มรากแบบจำกัด (ทรงกลม) โดยมีระบบท่อลำเลียงแตกแขนงล้อมรอบบริเวณที่ติดเชื้อตรงกลาง บ่อยครั้งที่ เซลล์ที่ติดเชื้อ ไรโซเบียม จะมี แวคิวโอลขนาดเล็กเท่านั้นในทางตรงกันข้าม ปุ่มรากบนถั่วลันเตา โคลเวอร์ และเมดิคาโก ทรันคาทูลาจะเป็นแบบไม่จำกัด เพื่อรักษาสภาพเนื้อเยื่อเจริญที่ทำงานอยู่ (อย่างน้อยก็ในช่วงเวลาหนึ่ง) ซึ่งจะสร้างเซลล์ใหม่สำหรับการติดเชื้อไรโซเบียม ดังนั้นจึงมีโซนของความเจริญเต็มที่อยู่ในปุ่มราก เซลล์ที่ติดเชื้อโดยทั่วไปจะมีแวคิวโอลขนาดใหญ่ ระบบท่อลำเลียงของพืชจะแตกแขนงและอยู่บริเวณรอบนอก
การโคลนนิ่ง
ภายใต้สภาวะที่เหมาะสม เนื้อเยื่อเจริญปลายยอดแต่ละอันสามารถพัฒนาเป็นพืชใหม่หรือโคลน ที่สมบูรณ์ได้ พืชใหม่เหล่านี้สามารถปลูกได้จากกิ่งปักชำที่มีเนื้อเยื่อเจริญปลายยอด อย่างไรก็ตาม เนื้อเยื่อเจริญปลายรากนั้นไม่สามารถโคลนได้ง่าย การโคลนนี้เรียกว่าการสืบพันธุ์แบบไม่อาศัยเพศหรือการสืบพันธุ์แบบไม่อาศัยเพศและมีการปฏิบัติกันอย่างแพร่หลายในด้านพืชสวนเพื่อผลิตพืชจำนวนมากที่มีจีโนไทป์ ที่ต้องการ กระบวนการนี้เรียกว่าเมริโคลนนิ่ง ซึ่งแสดงให้เห็นว่าสามารถลดหรือกำจัดไวรัสที่มีอยู่ในพืชแม่ในพืชหลายชนิดได้[ 31 ] [ 32 ]
การขยายพันธุ์โดยการปักชำเป็นอีกรูปแบบหนึ่งของการขยายพันธุ์แบบไม่อาศัยเพศที่เริ่มต้นการสร้างรากหรือยอดจากเซลล์แคมเบียมเมริสเต็มรอง ซึ่งอธิบายได้ว่าทำไมการ 'บาดเจ็บ' ที่โคนของกิ่งปักชำที่มาจากยอดจึงมักช่วยในการสร้างราก[ 33 ]
เนื้อเยื่อเจริญที่ถูกชักนำ
เนื้อเยื่อเจริญ (meristem) อาจถูกชักนำให้เกิดขึ้นในรากของพืชตระกูลถั่วเช่นถั่วเหลือง , Lotus japonicus , ถั่วลันเตาและMedicago truncatulaหลังจากติดเชื้อแบคทีเรียในดินที่เรียกกันทั่วไปว่าไรโซเบียมเซลล์ของคอร์เทกซ์ชั้นในหรือชั้นนอกในบริเวณที่เรียกว่า "ช่องสร้างปุ่มราก" ซึ่งอยู่ด้านหลังปลายรากที่กำลังเจริญเติบโต จะถูกชักนำให้แบ่งตัว สารส่งสัญญาณที่สำคัญคือ ไลโปโอลิโกแซ็กคาไรด์น็อดแฟคเตอร์ (Nod factor ) ซึ่งมีหมู่ข้างเคียงเพื่อช่วยให้เกิดความจำเพาะในการปฏิสัมพันธ์ โปรตีนตัวรับน็อดแฟคเตอร์ NFR1 และ NFR5 ถูกโคลนมาจากพืชตระกูลถั่วหลายชนิด รวมถึงLotus japonicus , Medicago truncatulaและถั่วเหลือง ( Glycine max ) การควบคุมเนื้อเยื่อเจริญของปุ่มรากใช้การควบคุมระยะไกลที่เรียกว่าการควบคุมตนเองของการสร้างปุ่มราก (Autoregulation of Nodulation: AON) กระบวนการนี้เกี่ยวข้องกับ ตัวรับไคเนสLRR ที่อยู่ในเนื้อเยื่อหลอดเลือดใบ(LjHAR1, GmNARK และ MtSUNN) การส่งสัญญาณของ เปปไทด์ CLE และปฏิกิริยาของ KAPP คล้ายกับที่พบในระบบ CLV1,2,3 นอกจากนี้ LjKLAVIER ยังแสดง ลักษณะการควบคุมปุ่มรากด้วยแม้ว่าจะยังไม่ทราบว่าเกี่ยวข้องกับตัวรับไคเนส AON อื่นๆ อย่างไร
เนื้อเยื่อเจริญด้านข้าง
เนื้อเยื่อเจริญด้านข้าง ซึ่งเป็นรูปแบบของการเจริญเติบโตทุติยภูมิของพืช จะเพิ่มขนาดเส้นผ่านศูนย์กลางให้กับลำต้นของพืช โดยส่วนใหญ่จะพบในพืชใบเลี้ยงคู่หลายปีที่อยู่รอดได้จากปีต่อปี เนื้อเยื่อเจริญด้านข้างมีสองประเภท ได้แก่ แคมเบียมหลอดเลือดและแคมเบียมเปลือก
ในแคมเบียมหลอดเลือด โฟลเอ็มและไซเล็มปฐมภูมิถูกสร้างขึ้นโดยเนื้อเยื่อเจริญปลายยอด หลังจากพัฒนาการเบื้องต้นนี้ โฟลเอ็มและไซเล็มทุติยภูมิถูกสร้างขึ้นโดยเนื้อเยื่อเจริญด้านข้าง ทั้งสองเชื่อมต่อกันผ่านชั้นบางๆ ของเซลล์พาเรนไคมาซึ่งแตกต่างไปเป็นแคมเบียมมัด แคมเบียมมัดแบ่งตัวเพื่อสร้างโฟลเอ็มและไซเล็มทุติยภูมิใหม่ หลังจากนั้น พาเรนไคมาคอร์ติคัลระหว่างทรงกระบอกหลอดเลือดจะแตกต่างไปเป็นแคมเบียมระหว่างมัด กระบวนการนี้จะเกิดขึ้นซ้ำๆ สำหรับการเจริญเติบโตแบบไม่จำกัด[ 34 ]
คอร์กแคมเบียมสร้างชั้นป้องกันรอบนอกของพืช กระบวนการนี้เกิดขึ้นหลังจากที่ไซเล็มและโฟลเอมทุติยภูมิขยายตัวแล้ว เซลล์พาเรนไคมัลคอร์ติคัลจะแตกต่างไปเป็นคอร์กแคมเบียมใกล้กับเอพิเดอร์มิสซึ่งสร้างเซลล์ใหม่ที่เรียกว่าเฟลโลเดอร์มและเซลล์คอร์ก เซลล์คอร์กเหล่านี้ไม่สามารถซึมผ่านน้ำและก๊าซได้เนื่องจากมีสารที่เรียกว่าซูเบอรินเคลือบอยู่[ 35 ]
ดูเพิ่มเติม
แหล่งที่มา
- ห้องปฏิบัติการกายวิภาคศาสตร์พืชมหาวิทยาลัยเท็กซัส ; ห้องปฏิบัติการของ เจ.ดี. เมาเซธภาพถ่ายจุลภาคของเซลล์และเนื้อเยื่อพืช พร้อมคำอธิบาย
- Schoof, Heiko; Lenhard, M; Haecker, A; Mayer, KF; Jürgens, G; Laux, T (2000). " เนื้อเยื่อเจริญปลายยอด ของ Arabidopsisถูกรักษาไว้ด้วยวงจรควบคุมระหว่างยีน Clavata และ Wuschel" . Cell . 100 (6): 635– 644. doi : 10.1016/S0092-8674(00)80700-X . PMID 10761929 . S2CID 8963007 .
- Scofield และ Murray (2006). แนวคิดที่เปลี่ยนแปลงไปของเนื้อเยื่อเจริญ. Plant Molecular Biology 60:v–vii.
ลิงก์ภายนอก
- Meristemania.org – งานวิจัยเกี่ยวกับเนื้อเยื่อเจริญ
สรุปเนื้อหา
ข้อมูลสำคัญจากบทความ
ข้อมูลสำคัญเกี่ยวกับ เนื้อเยื่อเจริญ
ในชีววิทยาของเซลล์เนื้อเยื่อเจริญเป็นโครงสร้างที่ประกอบด้วยเนื้อเยื่อ เฉพาะ ที่พบในพืช...
เนื้อเยื่อเจริญขั้นต้น
เนื้อเยื่อเจริญปลายยอด หรือที่เรียกว่าเนื้อเยื่อเจริญปฐมภูมิ ก่อให้เกิดลำต้นหลักของพืชและมีหน้าที่ใน การเจริญเติบโตขั้นต้น หรือการเพิ่มความยาวหรือความสูง [ 3 ] [ 4 ] เนื้อเยื่อเจริญปลายยอดอาจแบ่งตัวออกเป็นเนื้อเยื่อเจริญปฐมภูมิได้ 3 ชนิด:
เนื้อเยื่อเจริญทุติยภูมิ
หลังจากระยะการเจริญเติบโตหลักแล้ว เนื้อเยื่อเจริญด้านข้างจะพัฒนาขึ้นเป็นระยะการเจริญเติบโตทุติยภูมิ การเจริญเติบโตนี้จะเพิ่มขนาดเส้นผ่านศูนย์กลางของพืชจากลำต้นเดิม แต่ไม่ใช่พืชทุกต้นที่จะมีการเจริญเติบโตทุติยภูมิ เนื้อเยื่อเจริญทุติยภูมิมีสองประเภท ได้แก่...
เนื้อเยื่อเจริญปลายยอด
เนื้อเยื่อเจริญปลายยอดเป็นเนื้อเยื่อเจริญที่ยังไม่แบ่งตัวอย่างสมบูรณ์ (ไม่จำกัดการเจริญเติบโต) ของพืช ทำหน้าที่สร้างการเจริญเติบโตขั้นต้น ทำให้ลำต้นและรากยืดตัวออก เนื้อเยื่อเจริญปลายยอดจะสร้างเนื้อเยื่อเจริญขั้นต้นได้ 3 ชนิด...